
Abstract
Sexually transmitted infections (STIs) have remained a worldwide public health threat. It is difficult to control the spread of STIs, not only because of heterogeneous sexual transmission between men and women but also because of the complicated effects of sexual transmission among men who have sex with men (MSM) and mother-to-child transmission. Many studies point to the existence of a ‘bisexual bridge’, where STIs spread from the MSM network via bisexual connections. However, it is unclear how the MSM network affects heterosexual networks as well as mother-to-child transmission. To analyse the effect of MSM on the spread of STIs, we divided the population into four subpopulations: (i) women, (ii) men who have sex with women only (MSW), (iii) men who have sex with both men and women (MSMW), (iv) men who have sex with men exclusively (MSME). We calculated the type-reproduction numbers of these four subpopulations, and our analysis determined what preventive measures may be effective. Our analysis shows the impact of bisexual bridge on the spread of STIs does not outweigh their population size. Since MSM and mother-to-child transmission rates do not have a strong synergistic effect when combined, complementary prevention measures are needed. The methodologies and findings we have provided here will contribute greatly to the future development of public health.
Similar content being viewed by others


Introduction
The relationship between men who have sex with men (MSM) and sexually transmitted infections (STIs) has been extensively studied because the infection rates of various STIs in MSM are greater than those in women and men who have sex with women only (MSW) [1]. MSM have been widely studied in research on HIV; however, MSM were exposed to many STIs before the spread of HIV [2]. The term MSM has been used since 1990 to reflect the epidemiological belief that behaviour is responsible for the risk of STIs [3]. MSM are defined only by their behaviour, not their sexual identity. Many studies have estimated the proportion of MSM among the male population in various countries [4,5,6,7,8,9,10,11,12] (Table 1). According to these studies, the proportion of MSM among the male population is approximately 1% to 5% in many countries. MSM should be classified into two subpopulations: men who have sex with men exclusively (MSME) and men who have sex with men and women (MSMW). In the research studies shown in Table 1, MSME and MSMW are not distinguished; thus, the proportions of MSME and MSMW are unknown.
When we consider the persistence of STIs in human society, transmission routes are key factors [13, 14]. Hepatitis B virus (HBV) and human T-cell leukaemia virus type I (HTLV-1) are endemic STIs that are spread by sexual (horizontal) transmission and mother-to-child (vertical) transmission. What makes these viruses so persistent is that vertically infected infants can develop to reproductive age and spread the infections through sexual transmission [15,16,17,18]. We suspect that the presence of STIs, which have robust intragenerational routes of transmission, combined with mother-to-child transmission, which allows transmission across generations, have contributed significantly to the persistence of these viruses. If we can mathematically estimate the spreading efficiency of each of these transmission routes, we can understand persistence strategies in the context of the ecological and evolutionary biology of STIs. This will also help us develop vaccination strategies for HBV, for which a safe vaccine exists. To address the question of whether an STI can persist, we need to take a long-term view, and it is necessary to consider not only transmission due to MSM but also mother-to-child transmission.
There are many mathematical models of STIs focusing on human sexual networks [19, 20]. It is well known that human sexual networks are heterogeneous; most people have sexual contact with only a few partners, but a small number of sexually active people have sexual contact with hundreds of partners. This heterogeneity is believed to contribute to the persistence of STIs [21]. In a simple epidemic model of a network that does not consider sex [22], the basic reproduction number \({R}_{0}\), which measures the number of infections produced by an infected individual on average, is determined by the fluctuation in sexual activity \(a\):
Here, \(a\) is the degree of sexual activity, which is proportional to the number of sexual contacts, and thus, \(C\) is the variation of \(a\) [19]. If we consider heterosexual contacts between men and women (i.e., the network is bipartite), \({R}_{0}\) is proportional to the geometric mean of the value \(C\) for men and women [19]. If the contact frequency distribution is a power distribution and the network is scale-free [23, 24], then \({R}_{0}\) and \(C\) can approach infinity. Many studies have noted that sexual contact networks are scale-free [25,26,27,28].
There are few models that consider the following two points simultaneously. The first point is mother-to-child (vertical) transmission. Mother-to-child transmission (e.g., via the placenta, birth blood exposure, breast feeding) is an important transmission route of STIs [14, 29, 30]. Therefore, in our previous studies, we built models that simultaneously considered sexual (horizontal) and mother-to-child (vertical) transmission under the scale-free property of sexual contact frequency [31, 32]. In particular, a realistic model presented in our latest work took into account adult and infant mortality from infection, infertility and stillbirth caused by infection, as well as recovery with treatment [32]. Although many STIs can cause serious harm to infants infected via mother-to-child transmission, HBV and HTLV-1 do not immediately adversely affect infected infants, and infected infants can grow and spread the infection [33, 34].
The second point is how the MSM network indirectly affects mother-to-child transmission through heterosexual networks via bisexual connections, which are called ‘bisexual bridges’ [35]. Bisexual and homosexual contacts play an important role in the spread of STIs [36]. Fernando argued that the current Centers for Disease Control (CDC) risk subpopulation classification, in which MSMW and MSME are included in a single MSM subpopulation, makes it impossible to know the extent of STI (e.g., HIV) transmission from MSMW to heterosexual women [37]. Thus, to model MSM accurately, we must divide MSM into men who have sex with men exclusively (MSME) and men who have sex with men and women (MSMW) because the effects of these two behaviours on public health are very different.
In this study, we simultaneously considered (1) the network heterogeneity of human sexual contacts, (2) mother-to-child (vertical) transmission and (3) MSM (i.e., MSMW and MSME) to formulate type-reproduction numbers; the type-reproduction number is defined as an extension of the basic reproduction number, \({R}_{0}\). The type-reproduction number rather than \({R}_{0}\) is required when the population is classified into several subpopulations according to epidemiological characteristics [38, 39]. Here, the type-reproduction number for type \(i\), \({T}_{i}\), is the average number of secondary cases of type \(i\) produced by a primary case of type \(i\). Since \({T}_{i}<1\Leftrightarrow {R}_{0}<1\) regardless of type \(i\), a \({T}_{i}\) value less than one indicates that STIs will be eliminated. The spread of epidemics is prevented if we effectively vaccinate at least a fraction \((1-1/{T}_{i})\) of the susceptible target type [38]. We considered four subpopulations, women, MSMW, MSME, and MSW, and derived a formula to identify which types should be targeted for public health interventions. When it is possible to concentrate vaccination on a subpopulation, the target of vaccination is not necessarily determined by the relative sizes of the type-reproduction numbers, because the cost of vaccines depends on the size of the subpopulation. However, since public health authorities do not know who is in which subpopulation, all they can do is to promote the vaccine and educate people about safe sex. It is difficult to focus promotion solely on MSME or MSMW, because they cannot be identified by the others. Therefore, even if the size of the subpopulation is small, the cost may not be so small. Thus, the type-reproduction number can be a good indicator of public health.
Material and methods
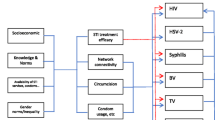
The outline of the model is illustrated in Fig. 1. To clarify the effect of MSM, we adapt a susceptibility-infection-susceptibility (SIS) model where the population is divided into four subpopulations: \(\{\mathrm{w},\overset{\sim }{\mathrm{m}},\overline{\mathrm{m} },\mathrm{m}\}\): women, MSMW, MSME, and MSW, respectively. It is assumed that a type \(\overset{\sim }{\mathrm{m}}\) person has contact not only with types \(\overset{\sim }{\mathrm{m}}\) and \(\overline{\mathrm{m} }\) but also with type \(\mathrm{w}\), while a type \(\overline{\mathrm{m} }\) person has sexual contact with only types \(\overset{\sim }{\mathrm{m}}\) and \(\overline{\mathrm{m} }\). Here, we ignore sexual contact between women; thus, a person of type \(\mathrm{w}\) can have sexual contact with persons of only types \(\mathrm{m}\) and \(\overset{\sim }{\mathrm{m}}\). Furthermore, the effect of generational change is also taken into consideration. Here, \(B\) is the number of births per unit time, \(\delta\) is the rate of infertility or stillbirths, and a newborn individual is infected at a probability of vertical transmission rate α times the female infection rate. The proportion of types of births is \({\gamma }_{\mathrm{w}}:{\gamma }_{\mathrm{m}}:{\gamma }_{\overset{\sim }{\mathrm{m}}}:{\gamma }_{\overline{\mathrm{m}} }\). Since it is not known when and how sexual orientation is determined, this model assumes that the types are fixed at birth for simplicity. The natural death rate is \({\mu }_{i}\), and the death rate for infected persons is \({\mu }_{i}^{^{\prime}}\) for type \(i\in \{\mathrm{w},\overset{\sim }{\mathrm{m}},\overline{\mathrm{m} },\mathrm{m}\}\). Thus, in the equilibrium state in the absence of disease (in the case of \({I}_{i}\left(t\right)=0\), as we will see later), the population in each type is

Conceptual diagram of the model. The population was divided into four subpopulations: women (w), MSMW (\(\overset{\sim }{\mathrm{m}}\)), MSME (\(\overline{\mathrm{m} }\)) and MSW (m). The subpopulation i individuals are born per unit time in \(B{\gamma }_{i}\) and die at a rate of \({\mu }_{i}\), where \({\gamma }_{i}\) represents the proportion of subpopulation \(i\) at the time of birth (\({\gamma }_{\mathrm{w}}+{\gamma }_{\overset{\sim }{\mathrm{m}}}+{\gamma }_{\overline{\mathrm{m}} }+{\gamma }_{\mathrm{m}}=1\)). δ is the rate of infertility or stillbirth. The number per unit time of sexual contacts between individuals in subpopulation i and individuals in subpopulation j is assumed to be \({f}_{ij}\) (white arrows in both directions). Sexual transmission occurs at a rate of \({\beta }_{i\to j}\), and mother-to-child transmission occurs at birth at a rate of \(\alpha\) (black arrows)
The total number per unit time of sexual contacts between individuals of type \(i\) and individuals of type \(j\) is \({f}_{ij}\) for \(i\ne j\) and \(\frac{1}{2}{f}_{ii}\) when \(i=j\). Although not specified for simplicity, \({f}_{ij}\) is a function of \({N}_{i}\) and \({N}_{j}\). From the definition, \({f}_{ij}={f}_{ji}\). From the assumptions of the model, \({f}_{\mathrm{ww}}={f}_{\mathrm{mm}}={f}_{\mathrm{m}\overset{\sim }{\mathrm{m}}}={f}_{\overset{\sim }{\mathrm{m}}\mathrm{m}}={f}_{\mathrm{w}\overline{\mathrm{m}} }={f}_{\overline{\mathrm{m}}\mathrm{w} }={f}_{\mathrm{m}\overline{\mathrm{m}} }={f}_{\overline{\mathrm{m}}\mathrm{m} }=0.\)
We assume that each individual in type \(i\) has a sexual activity value of \({a}_{i}\), and the value of \({a}_{i}\) follows the distribution of \({p}_{i}({a}_{i})\), where the mean of sexual activity is set to one:
The number per unit time of sexual contacts of an individual is proportional to the individual’s sexual activity value \({a}_{i}\) (thus \({a}_{i}\) is dimensionless). It is thought that the amount of \({a}_{i}\) changes with age, but in the model, where age is ignored, it is assumed to be constant for each individual. Thus, this model assumes that the values of \({a}_{i}\) is fixed at birth. The fluctuation in sexual activity defined as Eq. (1) is given as follows:
It is assumed that sexual contact is well distributed, ignoring monogamy and marriage, where the rate at which individuals with \({a}_{i}\) in subpopulation \(i\) sexually contact someone in subpopulation \(j\) is \({a}_{i}{f}_{ij}/{N}_{i}\).
The infection dynamics are as follows: \({S}_{i}(t)\) and \({I}_{i}(t)\) represent the numbers of susceptible and infected individuals in subpopulation \(i\), respectively. Assuming a susceptible-infected-susceptible model (SIS model) without carriers of immunity, the number of individuals in subpopulation \(i\) is given as \({N}_{i}\left(t\right)={S}_{i}\left(t\right)+{I}_{i}(t)\). The numbers of susceptible and infected individuals in subpopulation \(i\), whose sexual activities comprise infinitesimal interval \([{a}_{i},{a}_{i}+d{a}_{i}]\), are denoted as \({S}_{i}(t,{a}_{i})d{a}_{i}\) and \({I}_{i}(t,{a}_{i})d{a}_{i}\), respectively. Thus, the numbers of susceptible and infected individuals in type \(i\in \{\mathrm{w},\overset{\sim }{\mathrm{m}},\overline{\mathrm{m} },\mathrm{m}\}\) are given in integral form as follows:
The dynamics of \({S}_{i}(t,{a}_{i})\) and \({I}_{i}(t,{a}_{i})\) for \(i\in \{\mathrm{w},\overset{\sim }{\mathrm{m}},\overline{\mathrm{m} },\mathrm{m}\}\) are as follows:
where the parameter \({\beta }_{j\to i}\) is the probability of transmission per sexual contact from a person in subpopulation \(j\) to a person subpopulation \(i\), the parameter \({\eta }_{i}\) is the cure rate, and \({\Theta }_{i}\left(t\right)/{N}_{i}\left(t\right)\) represents the probability that the sexual partners are infected:
Results
In the absence of infection (\({I}_{i}\left(t,{a}_{i}\right)=0\)), we obtain a stationary solutions of Eq. (7):
To calculate the type-reproduction number, we consider the linear dynamics of the infected state, following the methods of the previous studies (for example [39, 40]). In other words, we linearize Eq. (8) near the disease-free solution given by Eq. (10). Here, we use Eqs. (6) and (9) to derive the dynamics of \({I}_{i}(t)\) and \({\Theta }_{i}(t)\) for \(i\in \{\mathrm{w},\overset{\sim }{\mathrm{m}},\overline{\mathrm{m} },\mathrm{m}\}\):
Here, \({\tilde{f }}_{ij}={f}_{ij}\left({\tilde{N }}_{i},{\tilde{N }}_{j}\right)\). Thus, the situation with very few infected persons is represented by a linear differential equation system closed by eight variables. According to the traditional method of calculating the type-reproduction number by Diekmann et al. [39], the Jacobi matrix \(J\) presented in Eq. (11) is divided into part \(T\), which is related to infection of the target type, and part \(Q=J-T\), and the dominant eigenvalue of \(-{TQ}^{-1}\) is calculated. Thus, we need to perform complicated algebraic operations on an eight-dimensional matrix.
We use the network diagram method according to the work by Lewis et al. [41], which is an intuitive and easy-to-understand method. Infected persons of each type are further divided into those by mother-to-child (vertical) transmission and those by sexual (horizontal) transmission. Thus, there are eight infection states, and we consider their network (see Fig. 2). In Fig. 2, red arrows indicate that infected persons in the state at the end of the arrow are infected from infected persons in the state at the beginning of the arrow. Blue arrows indicate that infected persons in the state at the ends of the arrow are born from an infected mother in the state at the beginning of the arrow. The quantities near the arrows give the number of new infections born per unit time divided by the duration of the original infected state. \({R}_{ij}^{\mathrm{v}}\) and \({R}_{ij}^{\mathrm{h}}\) represent the average number of the original persons in subpopulation \(j\) infected through sexual transmissions from a typical vertically and horizontally infected person in subpopulation \(i\), respectively, as follows [32]:

Network diagram among eight types of infection states. The red arrows represent the type-reproduction numbers of all sexual (horizontal) transmission events that occur between subpopulations. The blue arrows indicate the type-reproduction numbers of all mother-to-child (vertical) transmission events caused by infected women. The thin arrows indicate that the contribution is small when the proportion of homosexual individuals is small (see Table 3)
where \({C}_{i}\) is the fluctuation of sexual activity \({a}_{i}\) in subpopulation \(i\) (see Eq. (4)), and \({b}_{i}\) represents direct vertical transmission as follows:
The network diagram in Fig. 2 is represented by the following transition matrix:
The dominant eigenvalue of the matrix \(A\) gives the basic reproduction number, but it cannot be given in an analytical form because the characteristic equation becomes a quartic equation. On the other hand, the type-reproduction number can be written in a relatively simple formula. For example, if sexually infected females are the focus, the target matrix \(B\) has only nonzero entries \({B}_{2k}={A}_{2k}\) for \(1\le \mathrm{k}\le 8\). In this case, the type-reproduction number is given by the dominant eigenvalue of the matrix \(B(I-A+B)\) [39]. Since the rank of the matrix \(B(I-A+B)\) is one, the calculation of eigenvalues is easy. The type-reproduction number can be calculated using numerical-analysis software (the Mathematica source code is included in the Additional file 1). By performing some troublesome formula transformations, we obtain the type-reproduction numbers of persons in the four subpopulations who are infected sexually as follows if the denominators are positive:
Here, \({A}_{i}\) represents the average number of persons in each subpopulation (\(i\in\left\{\mathrm{w},\mathrm{m},\overset{\sim }{\mathrm{m}},\overline{\mathrm{m}}\right\}\)) who are infected through consecutive vertical transmissions from a sexually infected adult woman:
Note that in Eq. (15), \({R}_{i\to j}^{v}\) are always multiplied by \({A}_{i}\), such as \({A}_{i}{R}_{i\to j}^{v}\). Therefore, no vertical transmission (\(\alpha =0\)) always makes \({A}_{i}{R}_{i\to j}^{v}=0\), even if \({R}_{i\to j}^{v}\) is positive. If the denominators in Eq. (15) are not positive, they are not well defined, which means that infectious diseases are not extinct although the infection rate for the type of interest can approach zero. In particular, if the STI increases in only MSM,
the infection cannot be controlled without measures for MSM. On the other hand, if Eq. (16) does not hold, it is difficult to eradicate infections by targeting only MSM.
Figure 3 illustrates some typical results, and we make additional assumptions as follows:
-
The total numbers per unit time of sexual contacts are given as
$$\begin{array}{c}{f}_{\mathrm{wm}}={f}_{\mathrm{mw}}={k}_{\mathrm{hetero}}{N}_{\mathrm{m}}, {f}_{\mathrm{w}\overset{\sim }{\mathrm{m}}}={f}_{\overset{\sim }{\mathrm{m}}\mathrm{w}}={k}_{\mathrm{hetero}}{N}_{\overset{\sim }{\mathrm{m}}},\\ {f}_{\overset{\sim }{\mathrm{m}}\overset{\sim }{\mathrm{m}}}={k}_{\mathrm{homo}}\frac{{N}_{\overset{\sim }{\mathrm{m}}}^{2}}{{N}_{\overset{\sim }{\mathrm{m}}}+{N}_{\overline{\mathrm{m}}} }, {f}_{\overline{\mathrm{m} }\overline{\mathrm{m}} }={k}_{\mathrm{homo}}\frac{{N}_{\overline{\mathrm{m}} }^{2}}{{N}_{\overset{\sim }{\mathrm{m}}}+{N}_{\overline{\mathrm{m}}} },\\ {f}_{\overset{\sim }{\mathrm{m}}\overline{\mathrm{m}} }={f}_{\overline{\mathrm{m}}\overset{\sim }{\mathrm{m}} }={k}_{\mathrm{homo}}\frac{{N}_{\overset{\sim }{\mathrm{m}}}{N}_{\overline{\mathrm{m}}}}{{N }_{\overset{\sim }{\mathrm{m}}}+{N}_{\overline{\mathrm{m}}} }.\end{array}$$(17) -
As a result, for a man of types \(\mathrm{m}\) and \(\overset{\sim }{\mathrm{m}}\) and women, the average number of heterosexual contacts per unit time is \({k}_{\mathrm{hetero}}\) and \({k}_{\mathrm{hetero}}{N}_{\mathrm{m}}/{N}_{\mathrm{w}}\), respectively. They are approximately equal to each other because the sex ratio is approximately equal. For a man of types \(\overset{\sim }{\mathrm{m}}\) and \(\overline{\mathrm{m} }\), the average number of homosexual contacts per unit time is \({k}_{\mathrm{homo}}\).
-
Life history parameters (\({\gamma }_{i},{\eta }_{i},{\mu }_{i}^{^{\prime}}\)) and the fluctuation in sexual activity are set to \({C}_{i}=3\) for all categories \(i\in \left\{\mathrm{w},\mathrm{m},\overset{\sim }{\mathrm{m}},\overline{\mathrm{m}}\right\}\). The observational basis for the value of C will be explained later. Increasing \({C}_{i}\) reduces the effect of mother-to-child transmission (\({b}_{\mathrm{w}}\)), but the results do not change qualitatively.
-
Consider only three values of infection rates: female to male (\({\beta }_{\mathrm{w}\to \mathrm{m}}={\beta }_{\mathrm{w}\to \overset{\sim }{\mathrm{m}}}={\beta }_{\mathrm{w}\to \overline{\mathrm{m}} }\)), male to female (\({\beta }_{\mathrm{m}\to \mathrm{w}}={\beta }_{\overset{\sim }{\mathrm{m}}\to \mathrm{w}}={\beta }_{\overline{\mathrm{m}}\to \mathrm{w} }\)), and male to male (\({\beta }_{\overset{\sim }{\mathrm{m}}\to \overset{\sim }{\mathrm{m}}}={\beta }_{\overline{\mathrm{m}}\to \overset{\sim }{\mathrm{m}} }={\beta }_{\overset{\sim }{\mathrm{m}}\to \overline{\mathrm{m}} }={\beta }_{\overline{\mathrm{m} }\to \overline{\mathrm{m}} }\)).

The phase plane of the STI survival region and smallest type-reproduction numbers. The region above the black bold curve indicates the survival region of STIs (i.e., \({T}_{\mathrm{w}},{T}_{\mathrm{m}},{T}_{\overset{\sim }{\mathrm{m}}},{T}_{\overline{\mathrm{m}} }>1\)), and the four colours indicate which type-reproduction numbers are the smallest. The black dashed curves indicate that \({T}_{\overset{\sim }{\mathrm{m}}}\) is not well defined above them. In the cases of a and c the dashed curves are identical to the bold ones. In the colourless area, no type-reproduction numbers are well defined. The other parameters are set as \(({\gamma }_{\mathrm{w}},{\gamma }_{\mathrm{m}},{\gamma }_{\overset{\sim }{\mathrm{m}}},{\gamma }_{\overline{\mathrm{m}} })=(\mathrm{0.5,0.48,0},\mathrm{01,0.01})\)
Under the above assumption, the relative sizes of \({R}_{i\to j}^{\mathrm{v}}\) and \({A}_{i}\) are as shown in Table 2. In the phase plane in Fig. 3, the region above the bold curve indicates the persistence region of STIs (i.e., \({T}_{\mathrm{w}},{T}_{\mathrm{m}},{T}_{\overset{\sim }{\mathrm{m}}},{T}_{\overline{\mathrm{m}} }>1\)), the four colours indicate which type-reproduction numbers are the smallest, and the colourless region indicates that all \({T}_{i}\) are not well defined. The dashed curves indicate that \({T}_{\overset{\sim }{\mathrm{m}}}\) is not well defined above them (in Fig. 3a and c, the dashed curves coincide with the solid ones). If there are relatively few sexual contacts between men, it is more effective to focus infection control on women (w) (\({R}_{\overset{\sim }{\mathrm{m}}\to \overline{\mathrm{m}} }^{\mathrm{h}}={R}_{\overset{\sim }{\mathrm{m}}\to \overset{\sim }{\mathrm{m}}}^{\mathrm{h}}={R}_{\overline{\mathrm{m} }\to \overline{\mathrm{m}} }^{\mathrm{h}}={R}_{\overline{\mathrm{m}}\to \overset{\sim }{\mathrm{m}} }^{\mathrm{h}}=0.25\) in Fig. 3a and c) than on men. In this case, measures focused only on MSMW will not be able to suppress the STIs, because \({T}_{\overset{\sim }{\mathrm{m}}}\) is not well defined. On the other hand, if there are relatively more sexual contacts between men, there are some parameter areas where focusing infection control on MSMW (\(\overset{\sim }{\mathrm{m}}\)) is most effective (\({R}_{\overset{\sim }{\mathrm{m}}\to \overline{\mathrm{m}} }^{\mathrm{h}}={R}_{\overset{\sim }{\mathrm{m}}\to \overset{\sim }{\mathrm{m}}}^{\mathrm{h}}={R}_{\overline{\mathrm{m} }\to \overline{\mathrm{m}} }^{\mathrm{h}}={R}_{\overline{\mathrm{m}}\to \overset{\sim }{\mathrm{m}} }^{\mathrm{h}}=0.49\) in Fig. 3b and d). However, such a parameter range is narrow, and \({T}_{\overset{\sim }{\mathrm{m}}}\) is not well define in most of the blue regions where \({T}_{\mathrm{w}}\) is minimized. Moreover, Fig. 3e and f show that it would be most efficient to focus prevention measures on women (w) if there is little transmission among men and on MSME (\(\overline{\mathrm{m} }\)) otherwise. If there are many cases of both transmission types, infection cannot be suppressed by taking measures for only one subpopulation. This suggests that it is important to simultaneously prevent both homosexual and heterosexual transmission to suppress STIs. Moreover, in Fig. 4, we compare the influence of MSMW and MSME. Figure 4d-f show that when homosexuals contribute more to infectious diseases than heterosexuals, higher proportions of MSME require more measures against MSME than MSMW. On the other hand, Fig. 4a-c show that if the contributions to homosexual and heterosexual infections are equal, measures for women are important in any case.

The phase plane of the STI survival region and smallest type-reproduction numbers. The region above the black bold curve indicates the survival region of STIs (i.e., \({T}_{\mathrm{w}},{T}_{\mathrm{m}},{T}_{\overset{\sim }{\mathrm{m}}},{T}_{\overline{\mathrm{m}} }>1\)), and the four colours indicate which type-reproduction numbers are the smallest. The black dashed curves indicate that \({T}_{\overset{\sim }{\mathrm{m}}}\) is not well defined above them. In the colourless area, no type-reproduction numbers are well defined. In the top three figures, homosexual and heterosexual transmission rates are the same, whereas in the bottom three figures, homosexual transmission is twice as high as heterosexual transmission. The ratio of MSMW and MSME is set as (a,d) \({\gamma }_{\overset{\sim }{\mathrm{m}}}/{\gamma }_{\overline{\mathrm{m}} }=3\), (b,e) \({\gamma }_{\overset{\sim }{\mathrm{m}}}/{\gamma }_{\overline{\mathrm{m}} }=1\) and (c,f) \({\gamma }_{\overset{\sim }{\mathrm{m}}}/{\gamma }_{\overline{\mathrm{m}} }=1/3\) (set \({\gamma }_{\overset{\sim }{\mathrm{m}}}+{\gamma }_{\overline{\mathrm{m}} }=0.02\))
Discussion
In our previous paper [31], we proposed a simple STI model with heterosexual and vertical transmission and studied their mutual effect on the spread of STIs. In this model, people were divided into two types by sex. Moreover, we extended this model to include a juvenile type and showed that it is not necessary to include the juvenile type in the model because its effect can be mathematically reduced to postnatal effects [32]. In this paper, we formulated a type-reproduction number for the STI model that simultaneously considered (1) the network heterogeneity of human sexual contacts, (2) mother-to-child (vertical) transmission and (3) MSMW and MSME. These three factors greatly influence the spread of STIs, and we expect that the current approach will contribute to a comprehensive understanding of STI infection dynamics. It should be emphasized that the result given by Eq. (15) does not depend on the details of the model, such as the addition of the childhood stage.
In the current model, we assume a well-mixed population without consideration of the specific network structure, and we assume that each individual has a different level of sexual activity. This type of approximation is good for epidemic models with complex networks (e.g., [31, 42]). Although our model does not take into account the details of personal relationships (e.g., marital status, distinguishing between primary and casual sexual partners, repeated sexual contacts, and parent–child relationships), the result provides a good reference theory for complicated situations. Our model can reveal various trends in the population by changing the parameters. Several types of sexual contact (oral, anal and genital) can be considered differences in the level of sexual activity. Moreover, the transmission rates (\({\beta }_{i\to \mathrm{j}}\)) are dependent on sexual culture, which changes over time. In this study, we neglected women who have sex with women (WSW). Although WSW can potentially transmit STIs from current and prior male and female partners [42], it is unlikely that the WSW sexual network is a large reservoir of STIs, in contrast with the MSM network, because the prevalence of STIs in women who have sex with women exclusively (WSWE) is not higher than the prevalence in heterosexual women for many STIs [43, 44]. We considered only MSM, which has been confirmed to contribute significantly to the spread of STIs.
This model assumed that MSMW and MSME are innate, and the proportion of the sum of MSMW and MSME was set to 4% according to the previous studies shown in Table 1. Again, it is difficult to distinguish between MSMW and MSME, and the population ratio of MSMW and MSME cannot be determined definitively. This problem makes the estimation of the cost-effectiveness of prevention measures for each subpopulation difficult. The fluctuation of sexual activity is set to \({C}_{i}=3\) for all subpopulations according to the data of previous studies (see Table 3) [28, 45,46,47,48,49,50,51,52]. Although the values of \({C}_{i}\) may be slightly larger, our result does not change qualitatively when \({C}_{i}\) increases. There was almost no difference in \({C}_{\mathrm{i}}\) between men and women in Finland and Russia. In the UK, there was a large difference between men and women, with \({C}_{\mathrm{m}}=68\) for men and \({C}_{\mathrm{w}}=15\) for women. The C in Japan is large because the surveys cover all ages; thus, the variance in the total number of sexual partners is large. However, C in Japan may not be that large in reality since sexual activity in the model is assumed to be innate.
If the activity of homosexual individuals is similar to that of heterosexual individuals, the contribution of MSM to STI transmission is less than we suspected (as seen in Fig. 4). Under the realistic MSM population estimated from the previous studies shown in Table 1, prevention measures focusing on MSM are not efficient as long as there is not an explosive spread among the MSME population (Eq. (16) does not hold). It should be noted that in this model, MSMW is nearly twice as active as the other groups because it includes both homosexuality and heterosexuality. This seems to be an overestimation, but nevertheless, the impact of MSMW is not necessarily significant in this model. In the case that MSM is not important, it is more efficient to concentrate measures on women than men. Interestingly, a study published by Kahn et al. [53] in 1997 estimated that the annual number of HIV infections in the United States was approximately 40,000 and that infections transmitted by bisexual persons accounted for only 200–600 of those. Therefore, they concluded that transmission via bisexual contact was a relatively minor component of all HIV transmissions in the United States, and it seems that their findings are consistent with our model results [53].
Conclusion
In the current study, we constructed an STD model in which the population is divided into four subpopulations, women, MSMW, MSME, and MSW, and derived the analytical formula for type-reproduction numbers. As research on actual sexual contacts, including homosexuality, progresses in the future, this formula will be useful for developing preventive strategies. What we can say now is that it is important to simultaneously prevent both homosexual and heterosexual transmission to suppress STIs because MSM and mother-to-child transmission rates do not have a strong synergistic effect. Furthermore, our study is the first to quantify the effects of bisexual bridges on the spread of STIs. Understanding the potential role of MSMW and MSME in STI transmission from MSM to women is epidemiologically important. Our model shows the impact of bisexual bridge on the spread of STIs does not outweigh their population size.
Availability of data and materials
The authors declare that all data supporting the findings of this study are available within the article.
Abbreviations
- STIs:
-
Sexually transmitted infections
- MSW:
-
Men who have sex with women only
- MSM:
-
Men who have sex with men
- MSME:
-
Men who have sex with men exclusively
- MSMW:
-
Men who have sex with men and women
References
Centers for Disease Control and Prevention. Sexually transmitted disease surveillance 2017. Men who have sex with men. 2020. https://www.cdc.gov/std/stats17/msm.htm. Accessed 6 Aug 2020.
O’Leary D. The syndemic of AIDS and STDS among MSM. Linacre Q. 2014;81:12–37. https://doi.org/10.1179/2050854913y.0000000015.
Young RM, Meyer IH. The trouble with “MSM” and “WSW”: erasure of the sexual-minority person in public health discourse. Am J Public Health. 2005;95:1144–9. https://doi.org/10.2105/ajph.2004.046714.
Lieb S, Fallon SJ, Friedman SR, Thompson DR, Gates GJ, Liberti TM, Malow RM. Statewide estimation of racial/ethnic populations of men who have sex with men in the U.S. Public Health Rep. 2011;126:60–72. https://doi.org/10.1177/003335491112600110.
Grey JA, Bernstein KT, Sullivan PS, Purcell DW, Chesson HW, Gift TL, Rosenberg ES. Estimating the population sizes of men who have sex with men in US states and counties using data from the american community survey. JMIR Public Health Surveill. 2016;2:e14. https://doi.org/10.2196/publichealth.5365.
Rich AJ, Lachowsky NJ, Sereda P, Cui Z, Wong J, Wong S, Jollimore J, Raymond HF, Hottes TS, Roth EA, Hogg RS, Moore DM. Estimating the size of the MSM population in metro Vancouver, Canada, using multiple methods and diverse data sources. J Urban Health. 2018;95:188–95. https://doi.org/10.1007/s11524-017-0176-8.
Ezoe S, Morooka T, Noda T, Sabin ML, Koike S. Population size estimation of men who have sex with men through the network scale-up method in Japan. PLoS ONE. 2012;7:e31184. https://doi.org/10.1371/journal.pone.0031184.
Son VH, Safarnejad A, Nga NT, Linh VM, Tu LTC, Manh PD, Long NH, Abdul-Quader A. Estimation of the population size of men who have sex with men in vietnam: social app multiplier method. JMIR Public Health Surveill. 2019;5:e12451. https://doi.org/10.2196/12451.
Marcus U, Hickson F, Weatherburn P, Schmidt AJ. Estimating the size of the MSM populations for 38 European countries by calculating the survey-surveillance discrepancies (SSD) between self-reported new HIV diagnoses from the European MSM internet survey (EMIS) and surveillance-reported HIV diagnoses among MSM in 2009. BMC Public Health. 2013;13:919. https://doi.org/10.1186/1471-2458-13-919.
Torres TS, Marins LMS, Veloso VG, Grinsztejn B, Luz PM. How heterogeneous are MSM from Brazilian cities? An analysis of sexual behavior and perceived risk and a description of trends in awareness and willingness to use pre-exposure prophylaxis. BMC Infect Dis. 2019;19:1067. https://doi.org/10.1186/s12879-019-4704-x.
Edwards JK, Hileman S, Donastorg Y, Zadrozny S, Baral S, Hargreaves JR, Fearon E, Zhao J, Datta A, Weir SS. Estimating sizes of key populations at the national level: considerations for study design and analysis. Epidemiology. 2018;29:795–803. https://doi.org/10.1097/ede.0000000000000906.
Datta A, Lin W, Rao A, Diouf D, Kouame A, Edwards JK, Bao L, Louis TA, Baral S. Bayesian estimation of MSM population size in Côte d’Ivoire. Stat Public Policy (Phila). 2019;6:1–13. https://doi.org/10.1080/2330443x.2018.1546634.
Oriel JD. The scars of Venus: a history of venereology. London: Springer-Verlag; 1994.
Gross G, Tyring SK. Sexually transmitted infections and sexually transmitted diseases. Berlin, Heidelberg: Springer-Verlag; 2011.
World Health Organization. Hepatitis B. https://www.who.int/en/news-room/fact-sheets/detail/hepatitis-b. Accessed 10 June 2020.
Williams CKO, Cancer and AIDS. Part II: cancer pathogenesis and epidemiology. Switzerland: Springer Nature; 2019.
Iwanaga M, Watanabe T, Utsunomiya A, Okayama A, Uchimaru K, Koh K-R, Ogata M, Kikuchi H, Sagara Y, Uozumi K, Mochizuki M, Tsukasaki K, Saburi Y, Yamamura M, Tanaka J, Moriuchi Y, Hino S, Kamihira S, Yamaguchi K, for the Joint Study on Predisposing Factors of ATL Development investigators. Human T-cell leukemia virus type I (HTLV-1) proviral load and disease progression in asymptomatic HTLV-1 carriers: a nationwide prospective study in Japan. Blood. 2010;116:1211–9. https://doi.org/10.1182/blood-2009-12-257410.
Satake M, Iwanaga M, Sagara Y, Watanabe T, Okuma K, Hamaguchi I. Incidence of human T-lymphotropic virus 1 infection in adolescent and adult blood donors in Japan: a nationwide retrospective cohort analysis. Lancet Infect Dis. 2016;16:1246–54. https://doi.org/10.1016/S1473-3099(16)30252-3.
Anderson RM, May RM. Infectious diseases of humans: dynamics and control. Oxford: Oxford University Press; 1991.
Garnett GP. An introduction to mathematical models in sexually transmitted disease epidemiology. Sex Transm Infect. 2002;78:7–12. https://doi.org/10.1136/sti.78.1.7.
Hamilton DT, Handcock MS, Morris M. Degree distributions in sexual networks: a framework for evaluating evidence. Sex Transm Dis. 2008;35:30–40. https://doi.org/10.1097/olq.0b013e3181453a84.
May RM, Anderson RM. The transmission dynamics of human immunodeficiency virus (HIV). Philos Trans R Soc Lond B Biol Sci. 1988;321:565–607. https://doi.org/10.1098/rstb.1988.0108.
Newman MEJ. Networks: an introduction. New York: Oxford University Press; 2010.
Castellano C, Pastor-Satorras R. Thresholds for epidemic spreading in networks. Phys Rev Lett. 2010;105:218701. https://doi.org/10.1103/PhysRevLett.105.218701.
Liljeros F, Edling CR, Amaral LA, Stanley HE, Aberg Y. The web of human sexual contacts. Nature. 2001;411:907–8. https://doi.org/10.1038/35082140.
Schneeberger A, Mercer CH, Gregson SA, Ferguson NM, Nyamukapa CA, Anderson RM, Johnson AM, Garnett GP. Scale-free networks and sexually transmitted diseases: a description of observed patterns of sexual contacts in Britain and Zimbabwe. Sex Transm Dis. 2004;31:380–7. https://doi.org/10.1097/00007435-200406000-00012.
Latora V, Nyamba A, Simpore J, Sylvette B, Diane S, Sylvére B, Musumeci S. Network of sexual contacts and sexually transmitted HIV infection in Burkina Faso. J Med Virol. 2006;78:724–9. https://doi.org/10.1002/jmv.20614.
Ito H, Tamura K, Wada T, Yamamoto T, Morita S. Is the network of heterosexual contact in Japan scale free? PLoS ONE. 2019;14:e0221520. https://doi.org/10.1371/journal.pone.0221520.
Goldenberg RL, Andrews WW, Yuan A, MacKey HT, Louis MS. Pregnancy outcomes related to sexually transmitted diseases. In: Hitchcock PJ, MacKay HT, Wasserheit JN, editors. Sexually transmitted diseases and adverse outcomes of pregnancy. Washington DC: ASM Press; 1999. p. 1–24.
Klein JO, Remington JS. Current concepts of infections of the fetus and newborn infant. In: Remington JS, Klein JO, editors. infection diseases of fetus and newborn infant. Philadelpia: W.B. Saunders Company; 2000. p. 1–23.
Ito H, Yamamoto T, Morita S. Demography of sexually transmitted infections with vertical transmission. Appl Math Comput. 2019;348:363–70. https://doi.org/10.1016/j.amc.2018.12.002.
Ito H, Yamamoto T, Morita S. The type-reproduction number of sexually transmitted infections through heterosexual and vertical transmission. Sci Rep. 2019;9:17408. https://doi.org/10.1038/s41598-019-53841-8.
Tajima K. The 4th nation-wide study of adult T-cell leukemia/lymphoma (ATL) in Japan: estimates of risk of ATL and its geographical and clinical features. The T- and B-Cell Malignancy Study Group. Int J Cancer. 1990;45:237–43. https://doi.org/10.1002/ijc.2910450206.
Eriksen EM, Perlman JA, Miller A, Marcy SM, Lee H, Vadheim C, Zangwill KM, Chen RT, DeStefano F, Lewis E, Black S, Shinefield H, Ward JI. Lack of association between hepatitis B birth immunization and neonatal death: a population-based study from the vaccine safety datalink project. Pediatr Infect Dis J. 2004;23:656–62. https://doi.org/10.1097/01.inf.0000130953.08946.d0.
Jeffries WL IV. Beyond the bisexual bridge: sexual health among U.S. men who have sex with men and women. Am J Prev Med. 2014;47:320–9. https://doi.org/10.1016/j.amepre.2014.05.002.
Everett BG. Sexual orientation disparities in sexually transmitted infections: examining the intersection between sexual identity and sexual behavior. Arch Sex Behav. 2013;42:225–36. https://doi.org/10.1007/s10508-012-9902-1.
Fernando D. Men who have sex with men and women (MSMW), biphobia and the CDC: a bridge ignored?! Prev Med. 2017;105:368–9. https://doi.org/10.1016/j.ypmed.2017.06.013.
Heesterbeek JA, Roberts MG. The type-reproduction number T in models for infectious disease control. Math Biosci. 2007;206:3–10. https://doi.org/10.1016/j.mbs.2004.10.013.
Diekmann O, Heesterbeek JA, Roberts MG. The construction of next-generation matrices for compartmental epidemic models. J R Soc Interface. 2010;7:873–85. https://doi.org/10.1098/rsif.2009.0386.
Morita S. Type reproduction number for epidemic models on heterogeneous networks. arXiv:2006.03278. 2020.
Lewis MA, Shuai Z, van den Driessche P. A general theory for target reproduction numbers with applications to ecology and epidemiology. J Math Biol. 2019;78:2317–39. https://doi.org/10.1007/s00285-019-01345-4.
Pastor-Satorras R, Castellano C, Van Mieghem P, Vespignani A. Epidemic processes in complex networks. Rev Mod Phys. 2015;87:925. https://doi.org/10.1103/RevModPhys.87.925.
Gorgos LM, Marrazzo JM. Sexually transmitted infections among women who have sex with women. Clin Infect Dis. 2011;53(Suppl 3):S84–91. https://doi.org/10.1093/cid/cir697.
Bailey JV, Farquhar C, Owen C, Mangtani P. Sexually transmitted infections in women who have sex with women. Sex Transm Infect. 2004;80:244–6. https://doi.org/10.1136/sti.2003.007641.
Gouveia-Oliveira R, Pedersen AG. Higher variability in the number of sexual partners in males can contribute to a higher prevalence of sexually transmitted diseases in females. J Theor Biol. 2009;261:100–6. https://doi.org/10.1016/j.jtbi.2009.06.028.
Handcock MS, Jones JH. Likelihood-based inference for stochastic models of sexual network formation. Theor Popul Biol. 2004;65:413–22. https://doi.org/10.1016/j.tpb.2003.09.006.
Ashenhurst JR, Wilhite ER, Harden KP, Fromme K. Number of sexual partners and relationship status are associated with unprotected sex across emerging adulthood. Arch Sex Behav. 2017;46:419–32. https://doi.org/10.1007/s10508-016-0692-8.
Johnson AM, Wadsworth J, Wellings K, Bradshaw S, Field J. Sexual lifestyles and HIV risk. Nature. 1992;360:410–2. https://doi.org/10.1038/360410a0.
Johnson AM, Mercer CH, Erens B, Copas AJ, McManus S, Wellings K, Fenton KA, Korovessis C, Macdowall W, Nanchahal K, Purdon S, Field J. Sexual behaviour in Britain: partnerships, practices, and HIV risk behaviours. Lancet. 2001;358:1835–42. https://doi.org/10.1016/S0140-6736(01)06883-0.
Lewin B, Fugl-Meyer K, Helmius G, Lalos A, Månson S. Sex i Sverige: Om sexuallivet i Sverige 1996 [Sex in Sweden: on sex life in Sweden in 1996]. Stockholm: Statens Folkhälsoinstitut (Swedish National Institute of Public Health); 1998.
Haavio-Mannila E, Kontula O, Kuusi E. Trends in sexual life, measured by national sex surveys in Finland in 1971, 1992 and 1999, and a comparison to a sex survey in St. Petersburg in 1996. Helsinki: The Population Research Institute, The Family Federation of Finland; 2001.
Sewankambo NK, Gray RH, Ahmad S, Serwadda D, Wabwire-Mangen F, Nalugoda F, Kiwanuka N, Lutalo T, Kigozi G, Li CJ, Meehan MP, Brahmbatt H, Wawer MJ. Mortality associated with HIV infection in rural Rakai District, Uganda. AIDS. 2000;14:2391–400.
Kahn JG, Gurvey J, Pollack LM, Binson D, Catania JA. How many HIV infections cross the bisexual bridge? An estimate from the United States. AIDS. 1997;11:1031–7. https://doi.org/10.1097/00002030-199708000-00012.
Acknowledgements
Not applicable.
Funding
This work was supported by the JSPS KAKENHI (Grant nos. 17H04731, 19KK0262 and 21H01575 to H.I.; 17H04659 and 19H05737 to T.Y.; 18K03453 and 21K03387 to S.M.), the Joint Usage/Research Center on Tropical Disease, Institute of Tropical Medicine, Nagasaki University (2020-Ippan-01) and the Japan Science and Technology Agency Crest to S.M.
This funding source had no role in the design of this study and or in its execution, analyses, interpretation of the data, or decision to submit results.
Author information
Authors and Affiliations
Contributions
H.I. and S.M. conceived the study and wrote the manuscript. S.M. constructed the mathematical model. H.I., T.Y. and S.M. assisted in the interpretation of the results. H.I. and S.M. generated the figures. H.I., T.Y., and S.M. revised the references and data. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Mathematica code example for calculating the type-reproduction number.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ito, H., Yamamoto, T. & Morita, S. The effect of men who have sex with men (MSM) on the spread of sexually transmitted infections. Theor Biol Med Model 18, 18 (2021). https://doi.org/10.1186/s12976-021-00148-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12976-021-00148-9