
Abstract
Background
Several studies have reported that combination antiretroviral therapy (cART) enhances the hepatitis B surface antigen (HBsAg) clearance rate in Human Immunodeficiency Virus-1/Hepatitis B Virus (HIV/HBV) coinfected patients, yet the associated immunological characteristics remain unclear.
Methods
Global and specific immune phenotypic profiles were examined in 48 patients with HIV/HBV coinfection before cART and at 1-year, and 3-year after cART using flow cytometry. In addition, 61 patients with HBV monoinfection were included for comparison.
Results
HBsAg response (sAg-R) was defined as > 0.5 log decrease within six months of cART initiation, and 16 patients achieved it. Patients with sAg-R (the sAg-R group) exhibited distinct immune phenotypes compared to those of HBsAg-retained patients (the sAg-NR group). Notably, patients with sAg-R had lower CD4+ T cell counts and a higher number of HBcAg-specific T cells. Further, the sAg-R group exhibited upregulation of HLA-DR, Ki67, and PD-1 in CD4+ T cells and heightened HLA-DR and T-bet in CD8+ T cells. However, the sAg-R group had fewer TEMRA cells but more TEM and Th17 cells than those in the sAg-NR group. Expression of various markers, including HLA-DR+CD4+, Ki67+CD4+, PD-1+CD4+, CD38+CD8+, HLA-DR+CD8+, TIM-3+CD8+, HBV-specific CD4+ T cell secreting IFN-γ and IL-2, and specific CD8+ T cell secreting IFN-γ and IL-2, correlated with HBsAg decrease.
Conclusion
The decline in HBsAg levels during cART in HIV/HBV coinfection involves significant alterations in CD4+ and CD8+ T cells phenotypes, offering a novel perspective on a functional HBV cure.
Similar content being viewed by others

Background
Despite remarkable progress in preventive vaccines and antiviral therapies, chronic hepatitis B (CHB) still affects approximately 296 million people worldwide [1]. Achieving hepatitis B surface antigen (HBsAg) clearance is the ideal treatment endpoint for CHB [2, 3]. However, the HBsAg clearance rate in current nucleos(t)ide analog treated CHB remains low at 1–2% [4, 5]. HBV infections are common among patients living with human immunodeficiency virus (PLWH) worldwide [6]. Notably, several studies have reported a higher HBsAg clearance rate in patients with HIV/HBV coinfection after combination antiretroviral therapy (cART) initiation than in those with HBV monoinfection, especially with tenofovir disoproxil fumarate/ tenofovir alafenamide (TDF/TAF) treatment regimens [7,8,9]. While cohort studies have suggested that immune reconstitution under antiretroviral therapy may contribute to an improved HBsAg clearance rate, information on the immunological characteristics of patients with HIV/HBV coinfection is limited.
The impaired adaptive immune response plays a crucial role in the pathogenesis of persistent HBV infection [10], especially the dysfunction and exhaustion of HBV-specific T cells [11, 12]. HBsAg clearance in HBV monoinfection is partially explained by the restoration of specific T cells, increased cytokines such as IL-12, IL-17 A, and IP-10, and reduced T cell depletion [13,14,15,16]. Previous studies have demonstrated that individuals with HIV/HBV coinfection display reduced HBV-specific T-cell responses compared to those with HBV monoinfection [17, 18]. While cART promotes the recovery of CD4+ T cells, the restoration of HBV-specific T cells following cART in HIV/HBV coinfection remains inconclusive [19,20,21]. Importantly, the absence of TDF/TAF-based cART regimens in these may account for their inability to explain higher HBsAg clearance rates observed in patients with HIV/HBV following TDF/TAF-based cART. Therefore, further investigations are required to understand the immunological features associated with HBsAg clearance in patients with HIV/HBV coinfection undergoing modern cART therapy.
To investigate the immunological mechanisms involved in HBsAg decline in patients with HIV/HBV coinfection, we performed a longitudinal analysis of the immunological profiles of patients with HIV/HBV coinfection following cART. Our focus was on comparing the immunological profiles of HIV/HBV coinfected patients with and without rapid HBsAg decline after the cART initiation.
Methods
Study participants
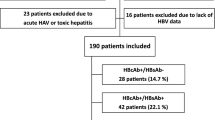
We previously studied a cohort of 51 people living with HIV/HBV coinfection who were enrolled in the Department of Infectious Disease at Peking Union Medical College Hospital (PUMCH) [22]. In this study, we enrolled 48 participants of these participants, with 3 excluded due to insufficient blood samples. All 48 patients were treatment-naïve for both HIV and HBV at the time of enrollment and subsequently received combination antiretroviral therapy (cART) based on 3TC or 3TC + TDF, with a follow-up period of at least six months.
For comparison of immunophenotypes, we also recruited 61 individuals with HBV monoinfection, which included 21 treatment-naïve patients (NT group) and 40 patients who were undergoing nucleos(t)ide analog (NAs) treatment (NAs group). Patients with hepatitis C coinfection, alcoholic and auto-autoimmune liver diseases, malignancy, and those receiving Peg-IFNα, corticosteroids, immunosuppressants, or chemotherapeutic drugs were excluded.
Study design
Participants with HIV/HBV coinfection were analyzed longitudinally. Routine blood tests, liver function tests, and absolute CD4+ and CD8+ T lymphocyte counts were determined at the PUMCH laboratory. Routine clinical data and blood samples were collected from each coinfected participant before cART (baseline) and at the first and thrid years after cART initiation.
HBV monoinfection was analyzed cross-sectionally, and samples were collected at only one-time point. The immunophenotypes of treatment-naïve HBV-infected patients (NT group) were analyzed as controls for HIV/HBV patients before cART. Participants with HBV infection receiving NAs treatment (NAs group) received treatment for a median period of 12 months. Immunophenotypes of the NAs group were analyzed as the control for HIV/HBV patients at the time of 1 year after initiation of cART. The characteristics of the participants were listed in Table 1.
Definition of HBsAg response
HBsAg response (sAg-R) was defined as a significant decrease in HBsAg level (> 0.5 log decrease from baseline levels) within six months from the initiation of cART. Participants living with HIV/HBV coinfection were stratified into sAg-R and HBsAg non-response (sAg-NR) groups according to whether or not they demonstrated a sAg-R.
Blood sample collection
Human peripheral blood mononuclear cells (PBMCs) were isolated using Ficoll density gradient centrifugation (TBD Science, Tianjin, China) and cryopreserved in liquid nitrogen for deferred use.
Serological assays
Hepatitis B surface antigen (HBsAg), anti-HBs, hepatitis B e-antigen, anti-HBe, and anti-HBc levels were detected using an Abbott Architect i2000 SR immunoassay analyzer (Abbott Diagnostics, Illinois, USA). HBsAg levels were quantified using the Abbott Architect i2000 platform.
HIV-1 total DNA measurement
HIV-1 total DNA levels were quantified using the HIV-1 DNA Quantification kit (Supbio, Guangzhou, China) on the ROCHE RT-qPCR platform, as previously described [23].
Phenotypic analysis using flow cytometry
To explore the phenotype of T cells, frozen PBMCs were thawed and rested overnight, then were incubated with Fixable Viability Stain 700 (FVS700; BD Biosciences, San Jose, USA). Surface staining was performed for 30 min at 4℃. For detection of intranuclear proteins, fixation and permeabilization (562574#, BD Biosciences) were performed before staining with intracellular antibodies. The antibodies used for phenotypic analysis are listed in Table S1. To perform intracellular cytokine staining (ICS) on global T cells, PMA (2 ng/mL; Beyotime, Shanghai, China), Ionomycin (1 µg/mL; Beyotime) and Glogistop Protein Transport Inhibitor (BD Biosciences) were added for 6 h following cell resuscitation. Surface and intracellular staining were performed as described above. A minimum of 100,000 events were acquired per sample tube by using a Fortessa LSR flow cytometer (BD Biosciences). Flow cytometry data were analyzed using FlowJo™ software version 10 (BD, Ashland, OR, USA). Single compensation tubes and fluorescent minus-one (FMO) tubes were used as controls. The analysis strategy for T cells and their subsets is illustrated in Figure S1-S3.
HBV-Specific T cell response experiments
Thawed PBMCs were stimulated with HBcAg overlapping peptides (genotypes B and C, GenBank accession numbers: AF121243 and AF112063, Genescript) for 10 days in R10. Thirty-seven core-specific peptides, 20 U/ml of human recombinant IL-2 (rhIL-2; R&D system, Bio-Techne, Minneapolis, MN, USA), 20 U/ml of human recombinant IL-7 (R&D system, Bio-Techne), 0.5 µg/mL of CD28/CD49a (BioLegend, San Diego, CA, USA) were used for stimulation and expansion on day 0. The culture was changed and replaced with half the original volume and supplemented with rhIL-2 on days 3 and 7. On day 10, PBMCs were restimulated for 6 h with the HBV core-peptide pool. Glogistop (BD Biosciences) was then used for 5 h. Then PBMCs were stained and tested for expression of IFN-γ, TNF-α, and IL-2 using flow cytometry analysis.
RNA sequencing and functional enrichment analysis
To analyze the differential gene expression profile determined by the HBsAg decline rate in patients with HIV/HBV coinfection, total RNA was extracted from PBMC using TRIzol (Invitrogen Life Technologies, Carlsbad, CA, USA), and samples were shipped frozen to Biomarker Technologies (Beijing, China) for transcriptome sequencing. Transcriptome sequencing was performed using a HiSeq 2500 platform (Illumina, San Diego, CA, USA). Differential expression analyses were performed using the BMKCloud platform (https://www.biocloud.net).
Statistical analysis
Statistical significance was determined using GraphPad Prism version 8.3 (GraphPad Software, Inc., CA, USA) or SPSS version 26 (IBM, Chicago, IL, USA) using the tests stated in the figure legends. Significant differences between the two groups were determined using the Mann-Whitney U test. Significant differences among three or more groups were determined using a mixed-effects model. Correlations between variables were determined using Spearman’s rank correlation test. The radar plot was depicted using Origin version 10.0 (Northampton, MA, USA), and a heat map was generated using Chiplot (https://www.chiplot.online). All reported P-values were two-sided, and a P value less than 0.05 was considered statistically significant.
Results
Characteristics of patients
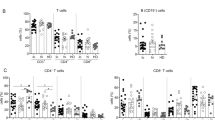
Forty-eight patients with HIV/HBV coinfection were included in this study, none of whom had prior exposure to HIV or HBV treatment. Nineteen patients were analyzed for HBV genotypes: thirteen were genotype C and six were genotype B. Thirty-eight patients (81.3%) were started on a 3TC + TDF-based antiviral regimen, nine patients received 3TC-based antiviral regimens, and one patient received a TAF/FTC-based antiviral regimen. After the initiation of cART, 16 patients achieved more than 0.5 log10 IU reduction in HBsAg levels from baseline within six months (HBsAg response group, sAg-R group), and 32 patients had HBsAg levels comparable to baseline during follow-up, considered to be in the HBsAg non-response group (HBsAg non-response group, sAg-NR group). In addition, patients with hepatitis B Virus patients (NAs and NT group) were included as controls. The demographic and clinical characteristic profiles of participants are provided in Table 1. At baseline, no differences were detected in HBeAg status, HBsAg level, HBV DNA, HBV pgRNA, and ALT levels between the sAg-R and sAg-NR groups.
Differential expression of activation and exhaustion markers in patients with different rates of HBsAg decline
Flow cytometry analysis showed that the proportion of CD4+ T cells in the sAg-R group was significantly lower than that in the sAg-NR group at baseline but recovered by the end of the third year (Fig. 1A). To understand the alteration of the immunophenotype during HBsAg clearance, we analyzed the expression levels of activation markers CD38 and HLA-DR; proliferation marker Ki67; exhaustion markers PD-1, TIM-3, and KLRG1; transcription factors T-bet and EOMES; and chemokine receptor CXCR5 in T cells using flow cytometry. As shown in Fig. 1B and Figure S4, HIV/HBV coinfection upregulated the expression of several activation and exhaustion markers compared to HBV monoinfection, such as CD38, HLA-DR, Ki67, PD-1, TIM-3, and T-bet on CD4+ T cells and CD38, HLA-DR, Ki67, PD-1, KLRG1, and T-bet on CD8+ T cells, whereas CXCR5, TIM-3, and EOMES were downregulated on CD8+ T cells (Fig. 1B). After initiation of cART, the expression levels of CD38, HLA-DR, Ki67, PD-1, TIM-3, KLRG1, and T-bet decreased in coinfected patients, whereas CXCR5 increased in sAg-R patients (Fig. 1C), indicating that antiviral treatment and suppression of viral load improved the immune exhaustion status among coinfected patients.

Expression of activation and exhaustion markers in patients with different rates of HBsAg decline. (A) Bar chart displays the median frequency of CD4+ and CD8+ T cells in HIV/HBV coinfected patients with HBsAg response (HIV.sAg-R) and non-response (HIV.sAg-NR) at baseline, 1 year, and 3 years after starting treatment, as well as treatment-naïve chronic hepatitis B virus-infected patients (NT) and nucleos(t)ide analogue (NAs)-treated hepatitis B virus patients (NAs Group). The Mann-Whitney test was used for analysis. (B) Radar plots illustrate the median expression of activation and exhaustion markers on CD4+ and CD8+ T cells for HIV.sAg-R, sAg-NR patients, NT, and NA patients. (C) Median marker expression on CD4+ and CD8+ T cells of sAg-R and sAg-NR patients at baseline, 1 year, and 3 years after starting treatment. Mixed-effect analysis was employed. (D-H) The median expression of HLA-DR+ CD4+, Ki67+ CD4+, PD-1+ CD4+, HLA-DR+ CD8+, and T-bet+ CD8+ in participants, along with representative flow cytometry analyses
*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. Abbreviation: ns, not significant
Notably, a different T cell immunophenotype was observed in the sAg-R patients compared to the sAg-NR patients. CD4+ T cells from the sAg-R group had significantly higher expression of HLA-DR (Fig. 1D; at baseline: 23.80% vs. 12.50%, P = 0.0111; at the first-year follow-up assessment: 24.9% vs. 14.9%, P = 0.0300), Ki67(Fig. 1E; at baseline: 3.3% vs. 2.4%, P = 0.0228), and PD-1 (Fig. 1F; at baseline: 14.9% vs. 9.1%, P = 0.0357) than those in the sAg-NR group. The expression of HLA-DR (Fig. 1G; at the first-year follow-up assessment: 42.2% vs. 29.1%, P = 0.0258) and T-bet (Fig. 1H; at baseline: 51.0% vs.42.5%, P = 0.0491; at the first-year follow-up, 49.5% vs. 39.3%, P = 0.0440) on CD8+ T cells in the sAg-R group was also significantly increased compared to those in the sAg-NR group. Taken together, these data demonstrated that the sAg-R group had higher T-cell activation and proliferation levels at baseline and longitudinally.
Increased effector memory T cell subsets in patients with rapid HBsAg decline
The differentiation memory phenotype was detected using CD45RA and CCR7 staining, followed by t-SNE and conventional flow cytometry analysis (Fig. 2). As expected, the t-SNE plot showed that the altered T subsets of HIV/HBV coinfection demonstrated higher proportions of CD8+ subsets and CD8+ effector memory T cells (CD45RA−CCR7−, TEM), and lower proportions of naïve T cells (CD45RA+CCR7+, Naïve) compared to those of HBV monoinfection. In contrast to the dramatic alterations in T cell subsets between coinfection and monoinfection, only minor changes were observed when the phenotype was characterized by different rates of HBsAg decline (sAg-R vs. sAg-NR group) following cART initiation (Fig. 2A). Further, the proportions of naïve T cells, TCM cells, and Tregs were similar between the sAg-R and sAg-NR patients. However, sAg-R patients had significantly higher frequencies of CD4+ TEM cell subsets after one year of cART and lower frequencies of CD4+ terminally differentiated effector memory T cells (CD45RA+CCR7−, TEMRA) before and during cART compared to sAg-NR patients (Fig. 2B). For CD8+ T cells, sAg-R patients also had significantly higher frequencies of TEM cell subsets and lower TEMRA than sAg-NR patients after one year of cART (Fig. 2B). These results suggested that HBV strongly impacts the differentiation of memory T cells.

Characterization of Memory T Cell Subsets in HBV and HIV/HBV Coinfected Patients. (A) t-SNE plots illustrating differences in CD4+ and CD8+ memory T cell populations at baseline, 1 year, and 3 years after treatment initiation. (B) Floating bars displaying the minimum and maximum values with medians for memory T cell subsets in HIV.sAg-R and sAg-NR patients at baseline, 1 year, and 3 years after starting treatment. Unpaired t-tests and Mann-Whitney tests were utilized for statistical analysis
*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001
Elevated IL17A and Th17/Treg ratio in patients with rapid HBsAg decline
Global T cell cytokines reflect the magnitude of inflammation and immune activation and contribute to viral control. To understand the extent of HBsAg clearance in HIV/HBV-coinfected patients, we evaluated the frequencies of global T cell-producing cytokine profiles in HIV/HBV coinfected and HBV monoinfected patients (Figure S5). Statistically significant differences were observed in the frequency of CD4+ T cells producing IL-17 between the sAg-R and sAg-NR groups during cART, with a higher frequency in the sAg-R than in the sAg-NR group (Fig. 3A, first year of cART: 2.6% vs. 1.2%, P = 0.0033). According to the results of IFN-γ, IL-4, and IL-17 A staining, cell subsets were divided into Th1 cells (CD4+ IFN-γ+, Th2 cells (CD4+ IL-4+), and Th17 cells (CD4+ IL-17 A+). As Fig. 3B showed, the Th17/Treg ratios were more imbalanced in the HIV/HBV coinfection group than in the HBV monoinfection group. Notably, the sAg-R group had a significantly higher Th17/Treg ratio than the sAg-NR group (0.35 vs. 0.21, P = 0.0463). Increased IL-17 levels and Th17/Treg ratios indicated an inflammatory response in patients with sAg-R.

The IL17 secreting CD4 T cell and the Th17/Treg in in Patients with Different Rates of HBsAg Decline. (A)Summary bar charts display the median with interquartile range of IL17 secreting CD4 T cell levels; and (B) the median with interquartile range of Th17/Treg ratio in NT, NAs, and HIV/HBV coinfected participants at baseline, 1 year, and 3 years after treatment initiation. Mann-Whitney tests were used for statistical analysis
*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001
Kinetic analysis of HBV-specific T cells in patients with HIV/HBV coinfection
We conducted a longitudinal analysis of HBV-specific CD4+ and CD8+ T cell responses in patients with HIV/HBV coinfection. As shown in Fig. 4A, the proliferation level (measured by Ki67 expression) of HBV-specific CD4+ T cells was significantly lower in HIV/HBV coinfection compared to HBV monoinfection, both pre-and post-antiretroviral treatment. Within coinfected patients, the sAg-R group exhibited a lower proliferation level of HBV-specific CD4+ T cells than the sAg-NR group, although the difference was not statistically significant. Additionally, the proliferative capacity of CD4+ T cells responding to HBcAg was restored in the sAg-R group. We also assessed the cytokine-secreting T cells that responded to HBV core peptides. We observed that HBV core-specific CD4+ T cells in HIV/HBV coinfected patients secreted lower IFN-γ but a higher IL-2 than HBV monoinfection. Interestingly, in the first year, the sAg-R group displayed higher levels of IFN-γ (3.2% vs. 1.5%, P = 0.0131) and IL-2 (1.72% vs. 1.06%, P = 0.0408) compared to the sAg-NR group.

Kinetic Analysis of HBV-Specific T Cells and Relationship Between Clinical Characteristics of Patients and Altered T-Cell Phenotype. The phenotypes of HBcAg-specific CD4+ (A), CD8+ T (B), and CEF-specific CD8+ T (C) cell responses were longitudinally analyzed. On the left, the frequency of specific cytokine-producing T cells is shown, while on the right, kinetic changes in specific T cell responses are demonstrated at indicated time points for sAg-R and sAg-NR patients. Horizontal bars depict median values. Mann-Whitney U tests were used for statistical analysis.*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. (D) Association between the clinical characteristics of patients and altered T cell phenotypes. On the left, this association is examined at baseline, and on the right, at 1 year after treatment. Spearman’s Rank correlation test was utilized
To assess specific CD8 + T cells, they were analyzed using PBMCs stimulated with CEF peptides as controls (Fig. 4B and C). The proliferative capacity of specific CD8+ T cells was lower in HIV/HBV coinfected patients than in HBV monoinfected patients, regardless of stimulation by HBV core or CEF peptides. CEF-specific CD8+ cells secreted significantly higher levels of IFN-γ and TNF-α in HBV-monoinfected patients than in HIV/HBV-coinfected patients. However, the secretion of IFN-γ and TNF-α by HBV core-specific CD8+ cells did not differ between HIV/HBV coinfection (sAg-R and sAg-NR) and monoinfection, both remaining at low levels. This result suggests that HIV infection affected the CD8+ cell response to CEF but did not further impair the poor specific CD8+ cell response to HBV. In addition, no recovery of specific CD8+ cell proliferation or secretory functions was observed. The results suggest that the HBsAg decline in HIV/HBV coinfected patients might be linked more closely to HBV-specific CD4+ T cells than CD8+ T cells.
Association between clinical characteristics of patients and T cell phenotype alterations
We investigated potential correlations between the T cell phenotype and clinical markers in HIV/HBV-coinfected patients using Spearman’s rank correlation test. Figure 4D illustrates that baseline level of HBV-specific CD4+ secreting IFN-γ (OR=-0.341, P = 0.032), IL-2 (OR=-0.452, P = 0.003), and specific CD8+ secreting IFN-γ (OR=-0.442, P = 0.003) were negatively correlated with HBsAg levels. After 1-year of treatment, levels of specific CD4+ secreting IL-2 (OR=-0.406, P = 0.021) and specific CD8+ secreting IL-2 (OR=-0.396, P = 0.025) remained negatively correlated with the HBsAg. This result suggested that HBsAg levels may suppress host HBV immunity both pre-and post-treatment. Moreover, positive associations were observed between HBV-specific T-cell responses and HBsAg decline post-treatment. HBV-specific CD4+ secreting IFN-γ (OR = 0.339, P = 0.028) and IL-2 (OR = 0.352, P = 0.022), and specific CD8+ secreting IFN-γ (OR = 0.292, P = 0.061) and IL-2 (OR = 0.338, P = 0.029) were positively associated with HBsAg decline. Furthermore, the expression levels of HLA-DR+ CD4+, Ki67+ CD4+, PD-1+ CD4+, CD38+ CD8+, HLA-DR+ CD8+, TIM-3+ CD8+ were positively correlated with HBsAg decline.
RNA transcription
To understand the immune alterations during HBsAg decline in coinfected patients, mRNA sequencing analysis was performed on the PBMC of 6 sAg-R and 9 sAg-NR patients, one year after cART initiation. A total of 120 genes were upregulated, and 271 genes were downregulated in the sAg-R group compared to the sAg-NR group (Fig. 5A, Table S2). A KEGG mapper was used for pathway enrichment analysis (Fig. 5B), which revealed an upregulation of the IL-17 signaling pathway in patients with sAg-R.

RNA-Seq and KEGG Analysis between sAg-R and sAg-NR Groups. (A) The volcano chart displays differentially expressed genes (DEGs) identified by RNA-Seq between sAg-R and sAg-NR groups. DEGs are defined as those with a p-value < 0.05 and a fold change (FC) > |1.5|. (B) The top-enriched pathways following KEGG analysis. The y-axis represents the pathways, and the horizontal axis indicates the enrichment factor. The color of the circle corresponds to the q value, which is the multiple hypothesis-corrected p-value. The size of the circle reflects the number of genes enriched in the pathway
Discussion
Recently, the phenomenon of HBsAg seroclearance in HIV/HBV coinfection following cART has received greater attention. This is primarily due to the noted higher rates of HBsAg seroclearance in coinfections than in cases of HBV mono-infections. Despite this emerging focus, there remains a notable gap in our understanding of the specific cellular immune responses in this context. We conducted a comprehensive longitudinal study of T cell phenotypic profiles and HBV-specific immune responses in patients with HIV/HBV coinfection three years after initiating cART. Our data suggest that the rapid HBsAg decline observed in coinfected patients is associated with T cell activation, especially in CD4+ T cells, along with changes in the frequency of TEM, Th17 cell populations, and HBV-specific CD4+ T cell responses.
Our findings of increased expression of HLA-DR+ CD4+, Ki67+ CD4+, and HLA-DR+ CD8+ in HBsAg decline, which suggest that the rapid decrease in HBsAg is associated with T cell activation in HIV/HBV coinfection. Consistent with our finding, a recent study measuring the T cell phenotype of patients with CHB with rapid HBsAg decrease and HBsAg seroclearance showed that the activated T cell phenotype was positively correlated with the degree of HBsAg decline [15]. Further, Rinker et al. revealed that patients with HBsAg loss had a higher T cell-activated phenotype than patients with retained HBsAg after structural drug discontinuation [24].
Furthermore, we observed a higher proportion of TEM and Th17 subsets but a lower TEMRA subpopulation in HIV/HBV coinfected patients with rapid HBsAg decline than in the sustained HBsAg group. TEM cells could respond rapidly to reinfection by producing effector cytokines, important for viral clearance [25]. A recent study by Islam et al. also reported an increased proportion of the TEM population in patients with HBsAg seroclearance in CHB [13]. Th17 cells play a dual role in viral clearance and liver inflammation during chronic HBV infection by eliciting the cytokines IL-17 and IL-2126. The higher levels of TEM and Th17 cells in the HBsAg-responsive group suggest that these cells may also play a role in HBsAg decline during HIV/HBV coinfection.
The correlation analysis indicated that HBsAg decline was associated with the level of HBV-specific CD4+ T cells secreting IFN-γ and IL-2, as well as HBV-specific CD8+ T cell secretion of IFN-γ. Moreover, after one year of treatment, we observed a higher level of HBV-specific CD4+ T cells secreting IFN-γ and IL-2 in coinfected patients within the sAg-R group. These findings suggest a connection between HBsAg clearance and HBV-specific CD4+ cell responses. Notably, conflicting data exists regarding the recovery of HBV-specific T cells in HIV/HBV-coinfected patients after the initiation of treatment [19, 27,28,29]. In this study, we observed the recovery of the proliferative capacity of HBV-specific T cells post-treatment without a concurrent increase in the overall quantity of cytokines secreted by specific T cells. However, the frequency of HBV-specific T cells in the peripheral blood remained low in both HIV/HBV coinfection and HBV monoinfection, with CEF-specific responses being higher in HBV monoinfected patients than in those with HIV/HBV coinfection.
In transcriptomic studies of patients with CHB, interferon-stimulated genes, immune activation/depletion genes and chemokine genes were significantly upregulated during the immune clearance phase [30, 31]. Our DEG analysis showed that patients in the sAg-R group exhibited an upregulated expression of chemokine and inflammatory factor genes enriched in the IL-17 signaling pathway compared to the sAg-NR group. This finding suggests that patients with HBsAg-responsive coinfection display characteristics indicative of the immune clearance phase.
Based on these findings, we speculate that immune activation and HBV-specific CD4+ cells may contribute to the clearance of HBsAg in coinfected patients. Previous studies have shown that patients who achieve HBsAg loss during NA therapy tend to have higher levels of HBV-specific CD4+ T cells compared to HBsAg-positive patients [32]. Additionally, HBV env-specific CD4+ T cells are linked to HBsAg loss following therapy withdrawal [33]. Theoretically, CD4+ T cells can enhance HBV-specific CD8+ T responses and facilitate HBsAb production by B cells [34]. Therefore, therapeutic strategies aimed at enhancing HBV-specific CD4+ T cell responses may increase the likelihood of achieving higher rates of HBsAg clearance. Moreover, in two preclinical mouse models of HBV carriers, the activation status of HBV-specific CD4+ T cells was critical in determining the success of therapeutic hepatitis B vaccination [35]. Thus, interventions targeting the enhancement of CD4+ T cell activation and response could contribute to the functional cure of HBV.
This study has several limitations. First, we did not measure the immunological phenotype of HIV/HBV coinfected patients at 12 and 24 weeks post-treatment, which may have led to missing important immunological changes during the early phase of treatment. Second, our study lacks liver tissue samples, preventing a comparative analysis of the phenotypic characteristics between peripheral blood and liver immune cells. Additionally, we did not include patients with acute HIV infection, which restricts our ability to evaluate the dynamics of HBsAg clearance and the corresponding immune phenotypes in chronic HBV-infected individuals who initiate treatment during the acute phase of HIV infection.
In summary, our findings offer a comprehensive analysis of immunophenotypic changes in HIV/HBV coinfected patients, suggesting that post-treatment immunophenotypic changes in coinfected patients may be associated with HBsAg clearance. This study provides crucial insights into the role of immune activation and HBV-specific CD4+ T cells as a functional cure for HBV infections. Our findings may partially explain the higher HBsAg seroclearance rate in coinfected patients.
Data availability
The original contributions presented in this study are included in the article and supplementary material. Further inquiries can be directed at the corresponding author.
References
World Health Organization: Hepatitis B. Updated 24 June 2022. Accessed 01. May 2023, https://www.who.int/news-room/fact-sheets/detail/hepatitis-b
Lim SG, Baumert TF, Boni C, et al. The scientific basis of combination therapy for chronic hepatitis B functional cure. Nat Rev Gastroenterol Hepatol Apr. 2023;20(4):238–53. https://doi.org/10.1038/s41575-022-00724-5.
Cornberg M, Lok AS, Terrault NA, Zoulim F, Faculty E-AHTEC. Mar. Guidance for design and endpoints of clinical trials in chronic hepatitis B - Report from the 2019 EASL-AASLD HBV Treatment Endpoints Conference(double dagger). J Hepatol. 2020;72(3):539–557. https://doi.org/10.1016/j.jhep.2019.11.003
Hsu YC, Yeh ML, Wong GL, et al. Incidences and determinants of functional cure during Entecavir or Tenofovir Disoproxil Fumarate for Chronic Hepatitis B. J Infect Dis Dec. 2021;1(11):1890–9. https://doi.org/10.1093/infdis/jiab241.
Hsu YC, Jun DW, Peng CY, et al. Effectiveness of entecavir vs tenofovir disoproxil fumarate for functional cure of chronic hepatitis B in an international cohort. Hepatol Int Dec. 2022;16(6):1297–307. https://doi.org/10.1007/s12072-022-10411-x.
Ahmadi Gharaei H, Fararouei M, Mirzazadeh A, et al. The global and regional prevalence of hepatitis C and B co-infections among prisoners living with HIV: a systematic review and meta-analysis. Infect Dis Poverty Jul. 2021;1(1):93. https://doi.org/10.1186/s40249-021-00876-7.
van Bremen K, Hoffmann C, Mauss S, et al. Obstacles to HBV functional cure: late presentation in HIV and its impact on HBV seroconversion in HIV/HBV coinfection. Liver Int Dec. 2020;40(12):2978–81. https://doi.org/10.1111/liv.14684.
Yoshikawa S, Yoshio S, Yoshida Y, et al. Impact of Immune reconstitution-Induced hepatic flare on Hepatitis B Surface Antigen loss in Hepatitis B Virus/Human Immunodeficiency Virus-1 coinfected patients. J Infect Dis Jun. 2021;15(12):2080–9. https://doi.org/10.1093/infdis/jiaa662.
Audsley J, Avihingsanon A, Littlejohn M, et al. Long-term TDF-Inclusive ART and progressive rates of HBsAg loss in HIV-HBV Coinfection-lessons for functional HBV cure? J Acquir Immune Defic Syndr. Aug. 2020;15(5):527–33. https://doi.org/10.1097/QAI.0000000000002386.
Ferrari C, Boni C, Rossi M, et al. T cell regulation in HBV-related chronic liver disease. J Hepatol May. 2017;66(5):1096–8. https://doi.org/10.1016/j.jhep.2016.10.002.
Maier H, Isogawa M, Freeman GJ, Chisari FV. PD-1:PD-L1 interactions contribute to the functional suppression of virus-specific CD8 + T lymphocytes in the liver. J Immunol Mar. 2007;1(5):2714–20. https://doi.org/10.4049/jimmunol.178.5.2714.
Raziorrouh B, Schraut W, Gerlach T, et al. The immunoregulatory role of CD244 in chronic hepatitis B infection and its inhibitory potential on virus-specific CD8 + T-cell function. Hepatol Dec. 2010;52(6):1934–47. https://doi.org/10.1002/hep.23936.
Islam M, Sevak JK, Sharma MK, et al. Immune predictors of hepatitis B surface antigen seroconversion in patients with hepatitis B reactivation. Aliment Pharmacol Ther Mar. 2023;57(6):689–708. https://doi.org/10.1111/apt.17306.
Kim SW, Yoon JS, Lee M, Cho Y. Toward a complete cure for chronic hepatitis B: novel therapeutic targets for hepatitis B virus. Clin Mol Hepatol Jan. 2022;28(1):17–30. https://doi.org/10.3350/cmh.2021.0093.
Xiong S, Zhu D, Liang B, et al. Longitudinal characterization of phenotypic profile of T cells in chronic hepatitis B identifies immune markers associated with HBsAg loss. EBioMedicine Jul. 2021;69:103464. https://doi.org/10.1016/j.ebiom.2021.103464.
Tout I, Loureiro D, Mansouri A, Soumelis V, Boyer N, Asselah T. Hepatitis B surface antigen seroclearance: Immune mechanisms, clinical impact, importance for drug development. J Hepatol Aug. 2020;73(2):409–22. https://doi.org/10.1016/j.jhep.2020.04.013.
Chang JJ, Sirivichayakul S, Avihingsanon A, et al. Impaired quality of the hepatitis B virus (HBV)-specific T-cell response in human immunodeficiency virus type 1-HBV coinfection. J Virol Aug. 2009;83(15):7649–58. https://doi.org/10.1128/JVI.00183-09.
Zhang X, Xing H, Feng X, Zhang H, Wang Y, Yan H. Hepatitis B virus (HBV)-specific T-cell responses to recombinant HBV core protein in patients with normal liver function and co-infected with chronic HBV and human immunodeficiency virus 1 (HIV-1). Virol J Jul. 2013;12:10:232. https://doi.org/10.1186/1743-422X-10-232.
Crane M, Sirivichayakul S, Chang JJ, et al. No increase in hepatitis B virus (HBV)-specific CD8 + T cells in patients with HIV-1-HBV coinfections following HBV-active highly active antiretroviral therapy. J Virol Mar. 2010;84(6):2657–65. https://doi.org/10.1128/JVI.02124-09.
Lascar RM, Lopes AR, Gilson RJ, et al. Effect of HIV infection and antiretroviral therapy on hepatitis B virus (HBV)-specific T cell responses in patients who have resolved HBV infection. J Infect Dis Apr. 2005;1(7):1169–79. https://doi.org/10.1086/428502.
Lascar RM, Gilson RJ, Lopes AR, Bertoletti A, Maini MK. Reconstitution of hepatitis B virus (HBV)-specific T cell responses with treatment of human immunodeficiency virus/HBV coinfection. J Infect Dis Dec. 2003;15(12):1815–9. https://doi.org/10.1086/379896.
Li X, Xu L, Lu L, et al. CD4(+) T cell counts and soluble programmed death-1 at baseline correlated with hepatitis B surface antigen decline in HIV/HBV coinfection during combined antiretroviral therapy. Front Cell Infect Microbiol. 2023;13:1178788. https://doi.org/10.3389/fcimb.2023.1178788.
Lin L, Yue YS, Wang ND, et al. Whole blood as an alternative to peripheral blood mononuclear cell for detection of total HIV-1 DNA. BMC Infect Dis Dec. 2020;10(1):941. https://doi.org/10.1186/s12879-020-05675-3.
Rinker F, Zimmer CL, Honer Zu Siederdissen C, et al. Hepatitis B virus-specific T cell responses after stopping nucleos(t)ide analogue therapy in HBeAg-negative chronic hepatitis B. J Hepatol Sep. 2018;69(3):584–93. https://doi.org/10.1016/j.jhep.2018.05.004.
Jabbari A, Harty JT. The generation and modulation of antigen-specific memory CD8 T cell responses. J Leukoc Biol Jul. 2006;80(1):16–23. https://doi.org/10.1189/jlb.0206118.
Huang Z, van Velkinburgh JC, Ni B, Wu Y. Pivotal roles of the interleukin-23/T helper 17 cell axis in hepatitis B. Liver Int Jul. 2012;32(6):894–901. https://doi.org/10.1111/j.1478-3231.2012.02764.x.
Boni C, Penna A, Bertoletti A, et al. Transient restoration of anti-viral T cell responses induced by lamivudine therapy in chronic hepatitis B. J Hepatol Oct. 2003;39(4):595–605. https://doi.org/10.1016/s0168-8278(03)00292-7.
Gotto J, Webster GJ, Brown D, Jenkins J, Dusheiko GM, Bertoletti A. The impact of HBV-DNA fluctuations on virus-specific CD8 + T cells in HBeAg + chronic hepatitis B patients treated with a steroid and lamivudine. J Viral Hepat Jun. 2006;13(6):415–25. https://doi.org/10.1111/j.1365-2893.2005.00716.x.
Lau GK, Cooksley H, Ribeiro RM, et al. Impact of early viral kinetics on T-cell reactivity during antiviral therapy in chronic hepatitis B. Antivir Ther. 2007;12(5):705–18.
Vanwolleghem T, Hou J, van Oord G, et al. Re-evaluation of hepatitis B virus clinical phases by systems biology identifies unappreciated roles for the innate immune response and B cells. Hepatol Jul. 2015;62(1):87–100. https://doi.org/10.1002/hep.27805.
Montanari NR, Ramirez R, Aggarwal A, et al. Multi-parametric analysis of human livers reveals variation in intrahepatic inflammation across phases of chronic hepatitis B infection. J Hepatol Aug. 2022;77(2):332–43. https://doi.org/10.1016/j.jhep.2022.02.016.
Boni C, Laccabue D, Lampertico P, et al. Restored function of HBV-specific T cells after long-term effective therapy with nucleos(t)ide analogues. Gastroenterology. 2012;143(4):963–e739. https://doi.org/10.1053/j.gastro.2012.07.014.
Li Y, Wen C, Gu S, et al. Differential response of HBV envelope-specific CD4 + T cells is related to HBsAg loss after stopping nucleos(t)ide analogue therapy. Hepatology. 2023;78(2):592–606. https://doi.org/10.1097/HEP.0000000000000334.
Bertoletti A, Ferrari C. Adaptive immunity in HBV infection. J Hepatol. 2016;64(1 Suppl):S71–83. https://doi.org/10.1016/j.jhep.2016.01.026.
Su J, Brunner L, Ates Oz E, et al. Activation of CD4 T cells during prime immunization determines the success of a therapeutic hepatitis B vaccine in HBV-carrier mouse models. J Hepatol. 2023;78(4):717–30. https://doi.org/10.1016/j.jhep.2022.12.013.
Acknowledgements
The authors express their gratitude to all the participants for their contributions to this study.
Funding
The research was supported by the National Key Technologies R&D Program for the 13th Five-year Plan (2017ZX10202101‐001), the Chinese Academy of Medical Sciences (CAMS) Innovation Fund for Medical Sciences (2021-I2M-1-037), the Key Clinical Specialties Program of Beijing and China and National High-Level Hospital Clinical Research Funding (2022-PUMCH-D-008).
Author information
Authors and Affiliations
Contributions
Xiaodi Li: Conceptualization(equal); data Curation(equal), formal analysis(equal), investigation(equal), methodology (equal), visualization(equal), resources (equal); software (equal); writing – original draft; writing-review and editing (equal). Ling Xu, Lianfeng Lu, Xiaosheng Liu, Yang Yang, and Yuanni Wu: formal analysis (equal); methodology (equal); validation (equal); visualization(equal); software (equal) writing – review and editing (equal). Ting Zhu, Xiaoxia Li, Yanling Li, Xiaojing Song, Yang Han, Wei Lyu: Data curation (equal); formal analysis (equal); investigation(equal), validation(equal); writing – review and editing (equal). Wei Cao, Taisheng Li: conceptualization(equal); data curation(equal); formal analysis (equal); funding acquisition(equal); investigation (equal); methodology (equal); project administration(equal); supervision(equal); validation (equal); writing –Original draft preparation; writing- review and editing (equal).
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was conducted in compliance with the Declaration of Helsinki and approved by the Institutional Review Board of PUMCH. Written informed consent was obtained from all participants.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Li, X., Xu, L., Lu, L. et al. Differential T-cell profiles determined by Hepatitis B surface antigen decrease among people with Human Immunodeficiency Virus /Hepatitis B Virus coinfection on treatment. J Transl Med 22, 901 (2024). https://doi.org/10.1186/s12967-024-05681-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12967-024-05681-y