
Abstract
Background
The primary goal of radiotherapy (RT) is to induce cellular damage on malignant cells; however, it is becoming increasingly recognized the important role played by the tumor microenvironment (TME) in therapy outcomes. Therapeutic irradiation of tumor lesions provokes profound cellular and biological reconfigurations within the TME that ultimately may influence the fate of the therapy.
Main content
Cancer-associated fibroblasts (CAFs) are known to participate in all stages of cancer progression and are increasingly acknowledged to contribute to therapy resistance. Accumulated evidence suggests that, upon radiation, fibroblasts/CAFs avoid cell death but instead enter a permanent senescent state, which in turn may influence the behavior of tumor cells and other components of the TME. Despite the proposed participation of senescent fibroblasts on tumor radioprotection, it is still incompletely understood the impact that RT has on CAFs and the ultimate role that irradiated CAFs have on therapy outcomes. Some of the current controversies may emerge from generalizing observations obtained using normal fibroblasts and CAFs, which are different cell entities that may respond differently to radiation exposure.
Conclusion
In this review we present current knowledge on the field of CAFs role in radiotherapy; we discuss the potential tumorigenic functions of radiation-induced senescent fibroblasts and CAFs and we make an effort to integrate the knowledge emerging from preclinical experimentation with observations from the clinics.
Video Abstract
Plain English summary
Radiotherapy (RT) is currently used to treat more than 50% of all diagnosed cancer cases and counts for around 40% of all cure rates. The primary goal of RT has always been to induce damage on tumor cells; thus, it has been traditionally believed that the therapeutic efficacy of RT is mediated exclusively by its capacity to directly kill malignant cells. However, tumors are complex tissues composed of multiple cellular and acellular elements often referred as the tumor microenvironment (TME) or tumor stroma. Both, the malignant and the non-malignant cells in tumors receive the same prescribed radiation dose during treatment. In recent years, it has become more and more evident the fundamental role played by elements of the TME on therapy outcomes. One of the most abundant and influential elements of the reactive stroma in tumors are cancer-associated fibroblasts (CAFs). CAFs are also exposed to the full prescribed radiation dose during the course of radiotherapy. Accumulated evidence suggests that, upon radiation, fibroblasts/CAFs avoid cell death but instead enter a permanent senescent (growth arrest) state, which in turn may influence the behavior of tumor cells and other components of the TME. Despite the proposed participation of senescent fibroblasts on tumor radioprotection, it is still incompletely understood the impact that RT has on CAFs and the ultimate role that irradiated CAFs have on therapy outcomes. Further research using appropriate experimental models to study CAFs, and more information from clinical research is needed to unveil the ultimate role played by CAFs on radiotherapy.
Similar content being viewed by others
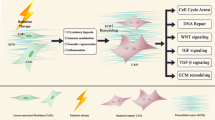
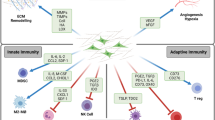
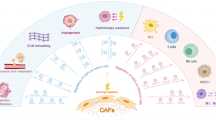
Background
Radiotherapy is currently used to treat more than 50% of all diagnosed cancer cases and counts for around 40% of all cure rates [1]. Traditionally, it has been thought that the therapeutic efficacy of RT is mediated predominantly by its capacity to directly kill malignant cells. Consequently, research aiming for improved therapeutic outcomes have focused almost entirely on the cancer cell itself, encouraging dose escalation strategies or the use of radiosensitizers for enhancing the antitumor effects [1, 2]. However, in recent years, it has become increasingly evident the fundamental role played by elements of the tumor stroma on therapy outcomes [3, 4]. Hence, today, several components of the stroma have been identified to interact with the response to RT. The extracellular matrix (ECM) can dictate radiation responsiveness by influencing oxygen availability, inducing pro-survival signals via cell-matrix interactions or by controlling the bioavailability of growth factors and cytokines [5,6,7,8]. Radiation can also induce endothelial cell dysfunctions, with altered expression of cell adhesion molecules and changes in the vascular network in tumors, which altogether may translate into impaired tumor perfusion, altered immune infiltration and/or increased hypoxia [9,10,11,12]. Moreover, the inexorable tissue damage provoked by RT is normally associated with an inflammatory response [13, 14], characterized by recruitment and activation of innate and adaptive immune cells and release of potent pro-inflammatory mediators [15,16,17].
As one of the most abundant elements of the reactive stroma in solid tumors, cancer-associated fibroblasts (CAFs) [18,19,20] are exposed to the full prescribed radiation dose during the course of clinical radiotherapy [21, 22]. However, the impact that radiation has on CAFs and the potential downstream effects of radiation-induced changes on therapy outcomes remain unsettled [11, 23,24,25]. Studies investigating cytotoxic effects of ionizing radiation (IR) on stromal cells have revealed the intrinsic radio-resistant nature of fibroblasts [26, 27]. Several in vitro observations have confirmed that following exposure to high radiation doses, CAFs [26, 28, 29] and normal tissue fibroblasts evade cell death [30, 31], but acquire a senescent phenotype [26, 30] accompanied by impaired proliferative and migratory functions [26]. A role of senescent cells in cancer development has been proposed, however such effects can be ascribed as beneficial or detrimental depending on the context [32,33,34,35,36,37,38]. Based on current knowledge on the topic of radiation effects in fibroblasts, it exists a generalized view postulating the acquisition of enhanced pro-malignant functions in irradiated (senescent) fibroblasts/CAFs, irrespective of their origin and functional status [39, 40]. However, most of the existing knowledge in this field emerge from studies conducted with normal tissue fibroblasts or fibroblast cell lines. In contrast to quiescent “normal” tissue fibroblasts (NFs), CAFs (in non-irradiated conditions) actively produce numerous tumor-promoting molecules such as matrix metalloproteinases (MMPs), inflammatory cytokines, pro-angiogenic factors and miscellaneous tumor-promoting growth factors [18, 20, 41]. Thus, to understand the contribution of CAFs to therapeutic outcomes post-RT, the activated status of the cells before treatment should be taken into consideration. Additionally, in contemporary high-precision image-guided radiotherapy settings with steep dose gradients, only cells residing within the delineated tumor volume (i.e. the planning target volume), or its periphery, are exposed to the full prescribed radiation dose (i.e. CAFs) [42], whereas healthy tissues located outside the target field (NFs) normal Fbs typically receive only residual radiation doses [21, 22, 43]. In this review, we discuss the controversies and misconceptions that have emerged in this field, due to the unrecognized fact that normal tissue fibroblasts and CAFs represent two different cell entities, and that radiation effects and responses may differ substantially between them.
Normal fibroblasts versus CAFs
Fibroblasts are spindle-shaped cells of mesenchymal origin which reside in connective tissues and are responsible for the production of ECM components and connective tissue homeostasis [44]. The embryonic origin of tissue resident fibroblasts is shared by other mesenchymal lineages including adipocytes, pericytes, chondrocytes and osteoblasts [45]. The difficulty in defining fibroblasts results largely from the lack of unique markers that are not expressed in other cell types [19, 45, 46]. In practical terms, fibroblasts are often defined by their specific cellular functions (ECM synthesis and remodeling), their lack of mutations and their lack of lineage markers for epithelial cells, endothelial cells and leukocytes. When quiescent, fibroblasts and stellate cells from specific organs do not proliferate, they form few cell-to-cell connections, they synthesize little collagen and secrete low amounts of cytokines and other signal mediators [18, 47].
Activation of tissue resident fibroblasts can occur in physiological conditions, such as wound healing or acute organ repair, or in pathologic conditions such as chronic infections, organ fibrosis, autoimmune diseases, or cancer [44, 45]. Activated fibroblasts can originate from multiple cell types including tissue-resident fibroblasts, mesenchymal progenitor cells, pericytes, organ-specific stellate cells or from trans-differentiation from different stromal cell types [18,19,20, 48]. Activation is triggered by multiple stimuli including hypoxia, injury-induced platelet activation and the secretion of potent factors by epithelial cells such as transforming growth factor- (TGF-β), platelet-derived growth factor (PDGF) and basic fibroblast growth factor (bFGF), resulting in increased proliferation, increased contractility, and expression of activation markers such as αSMA, PDGFRα/β, FAP, podoplanin (PDPN) and desmin [18, 20]. In the context of cancer, chronically activated fibroblasts/CAFs contribute to aberrant ECM deposition and desmoplasia, promote angiogenesis and regulate the infiltration and polarization of myeloid cells attracted to tumors [19]. A complex and intricate network of signaling pathways and crosstalk with epithelial cells and other stromal cells perpetuate the activation status of CAFs in tumors [18, 49, 50]. Most of the well-described tumor-promoting functions of CAFs happen in the context of paracrine and juxtracrine signaling via the secretion of multiple growth factors including TGF-β, connective tissue growth factor (CTGF), stromal-derived growth factor (SDF-1) vascular endothelial growth factor (VEGF), osteopontin (OPN) or hepatocyte growth factor (HGF) just to mention some, as well as a myriad of cytokines and chemokines such as IL-6, IL-8, IL-1, CXCL2, CXCL5, CXCL12/SDF-1a, CCL20 and others [18, 20, 50].
With the advent of single-cell sequencing, the complexity of CAF biology during tumor development and across different tumor entities has gradually become more evident [44, 51, 52]. CAFs, which were originally perceived as a homogenous population, are now understood to be a mixture of different fibroblast phenotypes with distinct behavior, comprising both tumor-promoting and tumor-restraining subtypes [46, 53,54,55]. Despite the proven existence of CAF diversity in both preclinical and clinical settings [56], the context-dependent roles of different CAF populations and their interchangeable plasticity remain largely unknown [56]. In this review, we have not put emphasis on the CAF diversity aspect since CAF heterogeneity has not been studied in the context of radiation or radiotherapy yet.
Role of senescent fibroblasts in cancer development
Cellular senescence is an established tumor suppressive mechanism that halts the proliferation of premalignant cells [37]. Replicative senescence in a physiological context is frequently associated with aging [38, 57], and it is attributed to the gradual loss of telomere length after repeated cell divisions, whereas premature cell senescence or accelerated senescence is normally triggered by exogenous stresses, including genotoxic anticancer treatments such as chemotherapy or radiotherapy [58, 59]. Despite its tumor suppressive nature, mounting evidence indicate that senescent cells can also promote tumor progression via the senescence-associated secretory phenotype or SASP [60].
In the context of cancer, senescent cells can act as a double-edged sword. On one hand, SASP promotes tissue repair through the induction of plasticity and stemness and participates in the clearance of cancer and damaged cells by attracting phagocytes, NK cells and other immune cells [61, 62]. Induction of cell senescence in fibroblasts also limit the development of desmoplastic reactions and fibrosis [63]. On the other hand, the presence of senescent cells in the tumor stroma or its surroundings can aid in the establishment of an immunosuppressive [64, 65], pro-angiogenic [66], pro-inflammatory and catabolic microenvironment [33, 67, 68] that stimulate tumor growth and cancer cell dissemination.
Senescence-associated alterations in the secretion of matricellular proteins and ECM constituents by fibroblasts have been shown to create a favorable milieu for tumor development [33]. The observed upregulation of MMPs, cathepsins, ADAMTs (a disintegrin and metalloproteinase with thrombospondin motifs) and other proteases, together with the downregulation of tissue inhibitors of matrix metalloproteinases (TIMPs) contribute to a catabolic environment that supports tumor cell invasion and metastasis. In contrast to ECM-degrading constituents, which are mainly overexpressed in senescent fibroblasts, ECM molecules have been reported to be downregulated in general [69, 70]. Senescent fibroblasts and mesenchymal cells can also promote cancer development by favoring epithelial-to-mesenchymal transition in premalignant and malignant cells [71, 72]. This phenomenon is mediated in a paracrine fashion by the release of molecules such as MMPs, IL-1β, IL-6 and reactive oxygen species (ROS). Additionally, senescent macrophages can directly or indirectly promote tumor vascularization by overexpressed secretion of pro-angiogenic factors and by promoting the recruitment and polarization of M2-macrophages [73]. Last, the SASP from both stromal and tumor cells promotes tumor growth by establishing a microenvironment that is immunosuppressive. Such induction is mediated by the secretion of immunosuppressive (Th2) cytokines that favor the recruitment of myeloid-derived suppressor cells (MDSCs) and consequently inhibition of CD8 + T-lymphocyte-mediated killing of tumor cells [64].
Pro-tumorigenic functions of irradiated (senescent) normal fibroblasts
Ionizing radiation has been shown to drive both stromal fibroblasts and cancer cells to premature senescence [30, 74, 75]. Accordingly, therapy-induced cell senescence has become a recognized side effect of anticancer treatments with potential to mediate substantial impact on therapy outcomes [59]. Soluble SASP elements such as MMPs have been associated with pro-tumorigenic effects from radiation-induced senescent fibroblasts in vitro and in vivo [30, 76, 77]. Moreover, insoluble (cell-associated) molecules such as syndecan-1 have also been shown to be overexpressed in radiation-induced senescent breast stromal fibroblasts, which reportedly have been demonstrated to mediate enhanced tumor progression [78].
In the context of radiotherapy, accumulated evidence reveals that on one hand, radiation-induced premature senescence instead of apoptosis is a major mode of cell fate in irradiated fibroblasts, and on the other hand, radiation-induced senescence is a dose-dependent factor, with higher doses being more effective in inducing premature senescence [26, 28]. Experiments related to the involvement of radiation-induced senescent fibroblasts in cancer promotion have usually been performed using normal human fibroblasts. In fact, the pro-malignant phenotype acquired by senescent normal fibroblasts post-RT have been thoroughly documented. Numerous in vitro studies have demonstrated increased invasiveness, proliferation rates and radio-resistance of tumor cells exposed to irradiated fibroblast cell lines (or their conditioned medium) when compared to non-irradiated cells (Table 1). Different soluble signal molecules have been proposed to be responsible for the radiation-induced enhanced effects including HGF [79], MMPs [80], TGF-β [81] as well as interleukin (IL) 6 and IL-8 [82]. There are also reports of pro-tumorigenic effects exerted by irradiated fibroblasts upon co-transplantation in in vivo models [30, 83]. Additionally, increased tumor incidence and growth have been observed in animal models when both pre-malignant and malignant cells are injected in pre-irradiated tissues [80, 84].
Collectively, the accumulated knowledge clearly indicate that radiation of normal tissue fibroblasts may turn them pro-tumorigenic via acquisition of a senescent phenotype and the associated pro-tumorigenic SASP. Also, the accumulation of senescent fibroblasts in premalignant tissues by processes of normal aging or genotoxic stress (radiation) may create a favorable environment for tumor initiation and growth (Table 1).
Role of (non-senescent) CAFs in radioprotection
Aiming at understanding the contribution of CAFs in tumor radio-resistance, some groups have investigated the potential radioprotective effects exerted by CAFs (non-irradiated/non-senescent) on cancer cells (Table 2). Several in vitro studies have demonstrated radioprotective effects of CAF conditioned medium on the survival and colony-forming abilities of cervical cancer [89] and pancreatic cancer cells [90]. Also, radioprotective effects from CAFs in co-culture with NSCLC cell lines have been described [91]. In preclinical models of melanoma and lung cancer, elevated expression of insulin-like growth factor 1 (IGF-1) and the chemokine CXCL12/SDF-1 by non-irradiated CAFs have been shown to be responsible for radioprotective effects on cancer cells [92]. In a study by Zhang et al. [93], CAF-derived CXCL1 was suggested to be accountable for the induction of a radioresistant phenotype in esophageal squamous cancer (ESCC), by promoting a reduction in ROS scavenging enzyme superoxide dismutase 1 (SOD-1) in cancer cells. Whereas in a pancreatic cancer model, it has been proposed that increased expression of TGF-β and possible other soluble signal molecules from pancreatic stellate cells (PSCs) promoted EMT changes in tumor cells and acquisition of a radioresistant phenotype [83]. Moreover, in a recent study by Ebbing et al. [94], using organoids and in vivo PDX models of esophageal cancer, authors point to CAF-derived IL-6 as a major soluble factor responsible for EMT induction and therapy resistance.
In clinical settings, the enhanced expression of CAF-specific markers or CAF-related gene signatures in tumor specimens have shown potential to predict responses to radio(chemo)therapy, and is persistently associated with poor prognosis in different types of cancers, including prostate cancer [95], cervical cancer [96], breast cancer [97], colon cancer [98], oral squamous cell carcinoma [99] and rectal cancer [100] (Table 3). Such consistency in clinical findings suggest that CAFs may play important roles in the conferral of radiation resistance phenotypes across many different solid malignancies.
Effects of RT on CAF tumorigenic functions
In addition to the general radioprotective functions assigned to CAFs, some studies claim that radiation exposure is amplifying the intrinsic radioprotective and pro-malignant effects exerted by CAFs (Table 4). In a study by Li et al. [101], irradiated CAFs provoked induction of epithelial mesenchymal transition (EMT) and enhanced invasive capacity of pancreatic cancer cells in co-cultures. RT-activated CAFs were found to excrete increased levels of CXCL12/SDF-1, ultimately promoting a mesenchymal phenotype in cancer cells and aiding to the overall tumor progression. Again, in a pancreatic cancer model, Mantoni et al. [102] demonstrated that pancreatic stellate cells (PSC) promote radioprotection and stimulate proliferation of pancreatic cancer cells in direct co-cultures and after co-injections in animal models. In this study, interfering with β1-integrin signaling abolished the radioprotective effects. In a third preclinical study performed on pancreatic cancer, authors show that induction of iNOS and nitric oxide (NO) in CAFs by RT is responsible for elevation of intratumoral pH and induction of a proinflammatory phenotype in tumor cells in a NFκB-dependent manner [103]. In a colorectal cancer model, Tommelein et al. [104] found increased IGF signaling from irradiated CAFs, and both IGF1 and IGF-binding proteins (IGFBP2) levels were elevated in supernatants from irradiated versus non-irradiated CAFs. In another preclinical study of colorectal cancer (CRC), upon irradiation, tumor-derived interleukin1α (IL-1α) mediated polarization of cancer-associated fibroblasts (CAFs) towards a pro-inflammatory pro-tumorigenic phenotype [105]. Authors demonstrated that IL-1-dependent signaling elevates oxidative DNA damage in iCAFs, which upon irradiation undergo senescence. This causes tissue remodeling and therapy resistance that can be overcome by inhibiting IL-1. Finally, in a recent study by Meng et al. [106], authors demonstrated that radiation-induced senescent CAFs promote non-small cell lung cancer (NSCLC) cell proliferation and radio-resistance through activation of the JAK/STAT pathway in tumor cells. Selective targeting of senescent fibroblast with a senotoxic agent was able to radio-sensitize tumors.
Conversely to the current view proposing a radiation-enhanced activation of CAFs, some studies document a loss of CAF pro-tumorigenic functions after irradiation (Table 4). In an in vivo study by Grinde et al. [107], the tumor enhancing effects exerted by NSCLC-CAFs after co-injection with A549 cells in nude mice was lost when CAFs were irradiated prior to implantation. In an in vitro study performed with NSCLC-CAFs, authors demonstrate changes in the secretory profile CAFs upon a single-high dose (1 × 18 Gy) radiation. However, in functional assays, they report no effects of conditioned media from irradiated CAFs on the proliferative or migratory capacity of tumor cells, and reduced migration rates on endothelial cells (HUVECs) [108]. Arshad et al. [87] reported similar findings, showing that murine lung CAFs did not affect the intrinsic radio-sensitivity of cancer cells. In contrast, reduced expression of TGF-β and MMPs were observed in co-culture supernatants after exposure to (1 × 10 Gy) radiation. In another study by Steer et al., the radio-protective and long-term survival effects of CAFs over cancer cells were studied in 2D and 3D in vitro systems, using different sets of fibroblasts and tumor cell lines [88]. The outcomes were inconsistent among different fibroblast-tumor cell combinations. Similar observations were obtained after co-implantation of cells in xenografts. Some authors have suggested that cancer promotion by senescent stromal cells may be restricted to certain organs and tissue types and claim that the tumorigenic properties of senescent cells need to be validated in other tissues than subcutaneous lesions [109].
CAFs impact on RT and vice-versa: observations from the clinics
In clinical settings, radiotherapy continue being a safe and efficient way to treat most known solid malignancies [1]. Exacerbated tumor growth after curative, adjuvant or neoadjuvant (chemo)radiotherapy is scantly observed, even though local tumor progression under or post-treatment may occur in minor subgroups of patients. In contrast, improved outcomes are frequently observed when radiation is used pre- or post-operative as opposed to surgery alone. Such long-term benefit from RT treatment have been reported for many different types of malignancies, ranging from early-stage breast cancer [110] to locally advanced colorectal cancer [111]. Considering that nearly all solid neoplasms contain tumor stroma and CAFs at all developmental stages, the evidence from the clinics suggests that the potential activation of CAFs post-RT (if happening at all) is not a dominant force that dictate the fate of the therapy. On the other hand, there is not much evidence on the impact of radiotherapy on CAFs in clinical settings. A study by Verset et al. on rectal cancer explored the impact of (chemo)radiotherapy on CAFs by comparing α-SMA/KI-67 ratios in tumor specimens collected before and after radiotherapy. Results showed that the α-SMA/neoplastic epithelial area ratio was higher after neoadjuvant therapy, and that α-SMA/epithelial area ratio was an adverse prognostic factor regarding recurrence-free survival [98]. Studies showing prognostic and predictive potential of CAF-markers in the clinics have been presented in the previous chapter (Table 3).
Concluding remarks
Scientific efforts over the last couple of decades have uncovered the important role played by the different elements of the tumor stroma in (radio)therapy outcomes. Cancer-associated fibroblasts have been proposed to participate significantly in tumor responses to radiotherapy. However, despite their prominent participation in tumor development and therapy resistance, the impact of RT on CAFs and the role of CAFs on RT outcomes remain elusive. Most existing literature in this field argue that radiation mediate changes in the mesenchymal components that favor tumor establishment and progression, however the impact that RT has on CAFs in vivo and the downstream effects of the potential RT-mediated changes remain controversial and still insufficiently investigated.
A potential source of controversy may emerge from the unrecognized fact that CAFs and normal fibroblasts are different cell entities, behaving differently, and probably responding differently to radiation exposure even when considering that both may survive and turn senescent after exposure to substantial radiation doses. Collectively, most published studies on the effects of radiation performed with normal tissue fibroblasts or fibroblast cell lines demonstrate enhanced pro-tumorigenic functions from irradiated (senescent) cells induced primarily in a paracrine fashion, by increased secretion of soluble growth factors, inflammatory mediators, and proteolytic enzymes (Table 1). Moreover, numerous studies have documented enhanced radioprotection of tumor cells cultured in the presence of (non-irradiated) CAFs (Table 2). These observations are in accordance with other studies performed on clinical specimens, demonstrating clear associations between high expression levels of CAF markers or CAF signature genes and poor prognosis (Table 3).
Findings on the impact of RT on CAFs and on the role of irradiated CAFs on tumor cell behavior are more controversial. While some studies claim that radiation exposure may affect CAF negatively through growth arrest and impaired mobility, others argue that exposure of CAFs to radiation can promote a more aggressive phenotype capable of conferring enhanced radio-resistance on tumor cells (Table 4). Most published studies have been performed on in vitro culture conditions, using single radiation doses or regimens, and have collected data at specific time-points, normally few hours/days post IR. These approaches gloss over potential differences related to different RT-regimens, that are crucial factors in modern radiotherapy [112] and radiotherapy-immunotherapy settings [4, 113,114,115,116] and also overlook potential long-term effects of RT on CAFs. Besides, most preclinical in vivo studies use co-injections of tumor cells and fibroblasts/CAFs orthotopically or in subcutaneous pockets. In transplantation experiments with admixed cells, it is frequently observed that non-tumoral cells disappear quickly after implantation [76, 107, 117]. Tumor growth effects in such experiments are believed to be related to initial tumor engraftment rather than tumor growth, and are therefore not optimal to study effects from transplanted (irradiated) cells. Additionally, there is little or no evidence on the impact that RT has on CAFs in vivo (preclinical) or in clinical settings. Preclinical models reproducing stroma-rich tumors resembling the human scenario, where endogenous CAFs can be targeted, tracked and/or regulated, could represent attractive models to explore CAF-mediated effects from RT and vice versa. In future efforts, the CAF heterogeneity aspect, including aged-fibroblasts, should also be taken into consideration. It is still unknown which CAF subtype (if any) is mostly responsible for conferring tumor cell radio-resistance. We still don’t know if radiation affects all CAF subtypes in the same way, or if the induction of cell senescence by RT unifies all subpopulations into a single CAF cell phenotype with specific functions.
In the clinics, radiotherapy is considered to be a safe and effective way to treat non-metastatic cancers with curative intent. Moreover, the use of RT in adjuvant and neoadjuvant settings has been proved to give better survival rates than surgery alone in many different tumor types [1]. Considering that basically all solid neoplasms contain CAFs at all developmental stages, the evidence from the clinics suggests that the potential activation of CAFs post-RT is not a dominant force that dictate the fate of the therapy. At least, it remains unexplained why local tumor recurrence or exacerbated tumor growth is not more frequently observed following RT, if we assume that radiation triggers enhanced pro-tumorigenic functions in CAFs. Further research using appropriate models to study CAFs and more information from clinical research is needed to clarify the ultimate role played by CAFs on radiotherapy.
Search strategy and selection criteria
Data for this review were identified by searches of MEDLINE, PubMed and references from relevant articles using search terms such as “cancer-associated fibroblasts”, “tumor microenvironment”, “radiation”, “radiotherapy”, “cell senescence”, “stress-induced senescence”. Only articles published between 1999 and 2022 have been included.
Availability of data and materials
Data sharing not applicable to this article as no datasets have been generated or analyzed for the creation of the study.
Abbreviations
- α-SMA:
-
Alpha-smooth muscle actin
- CAFs:
-
Cancer-associated fibroblasts
- ECM:
-
Extracellular matrix
- EMT:
-
Epithelial-mesenchymal transition
- IR:
-
Ionizing radiation
- MMPs:
-
Matrix metalloproteinases
- NFs:
-
Normal fibroblasts
- NSCLC:
-
Non-small cell lung cancer
- RT:
-
Radiotherapy
- SASP:
-
Senescence-associated secretory phenotype
- TME:
-
Tumor microenvironment
References
Sharma RA, Plummer R, Stock JK, Greenhalgh TA, Ataman O, Kelly S, et al. Clinical development of new drug-radiotherapy combinations. Nat Rev Clin Oncol. 2016;13(10):627–42.
Begg AC, Stewart FA, Vens C. Strategies to improve radiotherapy with targeted drugs. Nat Rev Cancer. 2011;11(4):239–53.
Barker HE, Paget JT, Khan AA, Harrington KJ. The tumour microenvironment after radiotherapy: mechanisms of resistance and recurrence. Nat Rev Cancer. 2015;15(7):409–25.
Hellevik T, Martinez-Zubiaurre I. Radiotherapy and the Tumor Stroma: The Importance of Dose and Fractionation. Front Oncol. 2014;4:1.
Nia HT, Munn LL, Jain RK. Physical traits of cancer. Science. 2020;370(6516):eaaz0868.
Barcellos-Hoff MH. The radiobiology of TGFbeta. Semin Cancer Biol. 2022;86(Pt 3):857–67.
Lan Y, Moustafa M, Knoll M, Xu C, Furkel J, Lazorchak A, et al. Simultaneous targeting of TGF-beta/PD-L1 synergizes with radiotherapy by reprogramming the tumor microenvironment to overcome immune evasion. Cancer Cell. 2021;39(10):1388–403.
Park CC, Zhang HJ, Yao ES, Park CJ, Bissell MJ. Beta1 integrin inhibition dramatically enhances radiotherapy efficacy in human breast cancer xenografts. Cancer Res. 2008;68(11):4398–405.
Garcia-Barros M, Paris F, Cordon-Cardo C, Lyden D, Rafii S, Haimovitz-Friedman A, et al. Tumor response to radiotherapy regulated by endothelial cell apoptosis. Science. 2003;300(5622):1155–9.
Park HJ, Griffin RJ, Hui S, Levitt SH, Song CW. Radiation-induced vascular damage in tumors: implications of vascular damage in ablative hypofractionated radiotherapy (SBRT and SRS). Radiat Res. 2012;177(3):311–27.
Martinez-Zubiaurre I, Chalmers AJ, Hellevik T. Radiation-induced transformation of immunoregulatory networks in the tumor stroma. Front Immunol. 2018;9:1679.
Stapleton S, Jaffray D, Milosevic M. Radiation effects on the tumor microenvironment: implications for nanomedicine delivery. Adv Drug Deliv Rev. 2017;109:119–30.
Surace L, Guckenberger M, van den Broek M. Radiation holidays stimulate tumor immunity. Oncotarget. 2015;6(18):15716–7.
Schaue D, Micewicz ED, Ratikan JA, Xie MW, Cheng G, McBride WH. Radiation and inflammation. Semin Radiat Oncol. 2015;25(1):4–10.
Gupta A, Probst HC, Vuong V, Landshammer A, Muth S, Yagita H, et al. Radiotherapy promotes tumor-specific effector CD8+ T cells via dendritic cell activation. J Immunol. 2012;189(2):558–66.
Burnette BC, Liang H, Lee Y, Chlewicki L, Khodarev NN, Weichselbaum RR, et al. The efficacy of radiotherapy relies upon induction of type i interferon-dependent innate and adaptive immunity. Cancer Res. 2011;71(7):2488–96.
Rodriguez-Ruiz ME, Vitale I, Harrington KJ, Melero I, Galluzzi L. Immunological impact of cell death signaling driven by radiation on the tumor microenvironment. Nat Immunol. 2020;21(2):120–34.
Kalluri R. The biology and function of fibroblasts in cancer. Nat Rev Cancer. 2016;16(9):582–98.
Sahai E, Astsaturov I, Cukierman E, DeNardo DG, Egeblad M, Evans RM, et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat Rev Cancer. 2020;20(3):174–86.
Biffi G, Tuveson DA. Diversity and biology of cancer-associated fibroblasts. Physiol Rev. 2021;101(1):147–76.
Stieb S, McDonald B, Gronberg M, Engeseth GM, He R, Fuller CD. Imaging for target delineation and treatment planning in radiation oncology: current and emerging techniques. Hematol Oncol Clin N Am. 2019;33(6):963–75.
Giglioli FR, Clemente S, Esposito M, Fiandra C, Marino C, Russo S, et al. Frontiers in planning optimization for lung SBRT. Phys Med. 2017;44:163–70.
Wang Z, Tang Y, Tan Y, Wei Q, Yu W. Cancer-associated fibroblasts in radiotherapy: challenges and new opportunities. Cell Commun Signal. 2019;17(1):47.
Ji X, Zhu X, Lu X. Effect of cancer-associated fibroblasts on radiosensitivity of cancer cells. Future Oncol. 2017;13(17):1537–50.
Hellevik T, Berzaghi R, Lode K, Islam A, Martinez-Zubiaurre I. Immunobiology of cancer-associated fibroblasts in the context of radiotherapy. J Transl Med. 2021;19(1):437.
Hellevik T, Pettersen I, Berg V, Winberg JO, Moe BT, Bartnes K, et al. Cancer-associated fibroblasts from human NSCLC survive ablative doses of radiation but their invasive capacity is reduced. Radiat Oncol. 2012;7:59.
Tachiiri S, Katagiri T, Tsunoda T, Oya N, Hiraoka M, Nakamura Y. Analysis of gene-expression profiles after gamma irradiation of normal human fibroblasts. Int J Radiat Oncol Biol Phys. 2006;64(1):272–9.
Berzaghi R, Islam A, Hellevik T, Martinez-Zubiaurre I. Secretion rates and protein composition of extracellular vesicles released by cancer-associated fibroblasts after radiation. J Radiat Res. 2021;62(3):401–13.
Yang N, Lode K, Berzaghi R, Islam A, Martinez-Zubiaurre I, Hellevik T. Irradiated tumor fibroblasts avoid immune recognition and retain immunosuppressive functions over natural killer cells. Front Immunol. 2020;11: 602530.
Papadopoulou A, Kletsas D. Human lung fibroblasts prematurely senescent after exposure to ionizing radiation enhance the growth of malignant lung epithelial cells in vitro and in vivo. Int J Oncol. 2011;39(4):989–99.
Pereira BI, Devine OP, Vukmanovic-Stejic M, Chambers ES, Subramanian P, Patel N, et al. Senescent cells evade immune clearance via HLA-E-mediated NK and CD8(+) T cell inhibition. Nat Commun. 2019;10(1):2387.
Lecot P, Alimirah F, Desprez PY, Campisi J, Wiley C. Context-dependent effects of cellular senescence in cancer development. Br J Cancer. 2016;114(11):1180–4.
Mavrogonatou E, Pratsinis H, Kletsas D. The role of senescence in cancer development. Semin Cancer Biol. 2020;62:182–91.
Wang L, Lankhorst L, Bernards R. Exploiting senescence for the treatment of cancer. Nat Rev Cancer. 2022;22(6):340–55.
Wyld L, Bellantuono I, Tchkonia T, Morgan J, Turner O, Foss F, et al. Senescence and cancer: a review of clinical implications of senescence and senotherapies. Cancers. 2020;12(8):2134-53.
Rodier F, Campisi J. Four faces of cellular senescence. J Cell Biol. 2011;192(4):547–56.
Gorgoulis V, Adams PD, Alimonti A, Bennett DC, Bischof O, Bishop C, et al. Cellular senescence: defining a path forward. Cell. 2019;179(4):813–27.
Campisi J. Aging, cellular senescence, and cancer. Annu Rev Physiol. 2013;75:685–705.
Bao CH, Wang XT, Ma W, Wang NN, Un Nesa E, Wang JB, et al. Irradiated fibroblasts promote epithelial-mesenchymal transition and HDGF expression of esophageal squamous cell carcinoma. Biochem Biophys Res Commun. 2015;458(2):441–7.
Hawsawi NM, Ghebeh H, Hendrayani SF, Tulbah A, Al-Eid M, Al-Tweigeri T, et al. Breast carcinoma-associated fibroblasts and their counterparts display neoplastic-specific changes. Cancer Res. 2008;68(8):2717–25.
Erez N, Truitt M, Olson P, Arron ST, Hanahan D. Cancer-associated fibroblasts are activated in incipient neoplasia to orchestrate tumor-promoting inflammation in an NF-kappaB-dependent manner. Cancer Cell. 2010;17(2):135–47.
Hoffman D, Dragojevic I, Hoisak J, Hoopes D, Manger R. Lung stereotactic body radiation therapy (SBRT) dose gradient and PTV volume: a retrospective multi-center analysis. Radiat Oncol. 2019;14(1):162.
Filice A, Casali M, Ciammella P, Galaverni M, Fioroni F, Iotti C, et al. Radiotherapy planning and molecular imaging in lung cancer. Curr Radiopharm. 2020;13(3):204–17.
Buechler MB, Pradhan RN, Krishnamurty AT, Cox C, Calviello AK, Wang AW, et al. Cross-tissue organization of the fibroblast lineage. Nature. 2021;593(7860):575–9.
Lendahl U, Muhl L, Betsholtz C. Identification, discrimination and heterogeneity of fibroblasts. Nat Commun. 2022;13(1):3409.
Ohlund D, Elyada E, Tuveson D. Fibroblast heterogeneity in the cancer wound. J Exp Med. 2014;211(8):1503–23.
Lynch MD, Watt FM. Fibroblast heterogeneity: implications for human disease. J Clin Invest. 2018;128(1):26–35.
Arina A, Idel C, Hyjek EM, Alegre ML, Wang Y, Bindokas VP, et al. Tumor-associated fibroblasts predominantly come from local and not circulating precursors. Proc Natl Acad Sci U S A. 2016;113(27):7551–6.
Mishra P, NBanerjee D, Ben-Baruch A. Chemokines at the crossroads of tumor-fibroblast interactions that promote malignancy. J Leukoc Biol. 2011;89:31–9.
Jung JG, Le A. Targeting Metabolic cross talk between cancer cells and cancer-associated fibroblasts. Adv Exp Med Biol. 2018;1063:167–78.
Dominguez CX, Muller S, Keerthivasan S, Koeppen H, Hung J, Gierke S, et al. Single-Cell RNA sequencing reveals stromal evolution into LRRC15(+) myofibroblasts as a determinant of patient response to cancer immunotherapy. Cancer Discov. 2020;10(2):232–53.
Joseph DB, Henry GH, Malewska A, Reese JC, Mauck RJ, Gahan JC, et al. Single-cell analysis of mouse and human prostate reveals novel fibroblasts with specialized distribution and microenvironment interactions. J Pathol. 2021;255(2):141–54.
Ohlund D, Handly-Santana A, Biffi G, Elyada E, Almeida AS, Ponz-Sarvise M, et al. Distinct populations of inflammatory fibroblasts and myofibroblasts in pancreatic cancer. J Exp Med. 2017;214(3):579–96.
Menezes S, Okail MH, Jalil SMA, Kocher HM, Cameron AJM. Cancer-associated fibroblasts in pancreatic cancer: new subtypes, new markers, new targets. J Pathol. 2022;257(4):526–44.
Jenkins BH, Buckingham JF, Hanley CJ, Thomas GJ. Targeting cancer-associated fibroblasts: challenges, opportunities and future directions. Pharmacol Ther. 2022;240: 108231.
Chen Y, McAndrews KM, Kalluri R. Clinical and therapeutic relevance of cancer-associated fibroblasts. Nat Rev Clin Oncol. 2021;18(12):792–804.
Harley CB, Futcher AB, Greider CW. Telomeres shorten during ageing of human fibroblasts. Nature. 1990;345(6274):458–60.
Toussaint O, Dumont P, Remacle J, Dierick JF, Pascal T, Frippiat C, et al. Stress-induced premature senescence or stress-induced senescence-like phenotype: One in vivo reality, two possible definitions? ScientificWorldJournal. 2002;2:230–47.
Gewirtz DA, Holt SE, Elmore LW. Accelerated senescence: an emerging role in tumor cell response to chemotherapy and radiation. Biochem Pharmacol. 2008;76(8):947–57.
Kuilman T, Peeper DS. Senescence-messaging secretome: SMS-ing cellular stress. Nat Rev Cancer. 2009;9(2):81–94.
Giannoula Y, Kroemer G, Pietrocola F. Cellular senescence and the host immune system in aging and age-related disorders. Biomed J. 2023.
Kale A, Sharma A, Stolzing A, Desprez PY, Campisi J. Role of immune cells in the removal of deleterious senescent cells. Immun Ageing. 2020;17:16.
Krizhanovsky V, Yon M, Dickins RA, Hearn S, Simon J, Miething C, et al. Senescence of activated stellate cells limits liver fibrosis. Cell. 2008;134(4):657–67.
Toso A, Revandkar A, Di Mitri D, Guccini I, Proietti M, Sarti M, et al. Enhancing chemotherapy efficacy in Pten-deficient prostate tumors by activating the senescence-associated antitumor immunity. Cell Rep. 2014;9(1):75–89.
Buechler MB, Turley SJ. A short field guide to fibroblast function in immunity. Semin Immunol. 2018;35:48–58.
Davalos AR, Coppe JP, Campisi J, Desprez PY. Senescent cells as a source of inflammatory factors for tumor progression. Cancer Metastasis Rev. 2010;29(2):273–83.
Coppe JP, Patil CK, Rodier F, Sun Y, Munoz DP, Goldstein J, et al. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008;6(12):2853–68.
Rodier F, Munoz DP, Teachenor R, Chu V, Le O, Bhaumik D, et al. DNA-SCARS: distinct nuclear structures that sustain damage-induced senescence growth arrest and inflammatory cytokine secretion. J Cell Sci. 2011;124(Pt 1):68–81.
Mavrogonatou E, Pratsinis H, Papadopoulou A, Karamanos NK, Kletsas D. Extracellular matrix alterations in senescent cells and their significance in tissue homeostasis. Matrix Biol. 2019;75–76:27–42.
Bizot-Foulon V, Bouchard B, Hornebeck W, Dubertret L, Bertaux B. Uncoordinate expressions of type I and III collagens, collagenase and tissue inhibitor of matrix metalloproteinase 1 along in vitro proliferative life span of human skin fibroblasts. Regulation by all-trans retinoic acid. Cell Biol Int. 1995;19(2):129–35.
Parrinello S, Coppe JP, Krtolica A, Campisi J. Stromal-epithelial interactions in aging and cancer: senescent fibroblasts alter epithelial cell differentiation. J Cell Sci. 2005;118(Pt 3):485–96.
Di GH, Liu Y, Lu Y, Liu J, Wu C, Duan HF. IL-6 secreted from senescent mesenchymal stem cells promotes proliferation and migration of breast cancer cells. PLoS ONE. 2014;9(11): e113572.
Kelly J, Ali Khan A, Yin J, Ferguson TA, Apte RS. Senescence regulates macrophage activation and angiogenic fate at sites of tissue injury in mice. J Clin Invest. 2007;117(11):3421–6.
Jones KR, Elmore LW, Jackson-Cook C, Demasters G, Povirk LF, Holt SE, et al. p53-Dependent accelerated senescence induced by ionizing radiation in breast tumour cells. Int J Radiat Biol. 2005;81(6):445–58.
Mirzayans R, Scott A, Cameron M, Murray D. Induction of accelerated senescence by gamma radiation in human solid tumor-derived cell lines expressing wild-type TP53. Radiat Res. 2005;163(1):53–62.
Liu D, Hornsby PJ. Senescent human fibroblasts increase the early growth of xenograft tumors via matrix metalloproteinase secretion. Cancer Res. 2007;67(7):3117–26.
Coppe JP, Desprez PY, Krtolica A, Campisi J. The senescence-associated secretory phenotype: the dark side of tumor suppression. Annu Rev Pathol. 2010;5:99–118.
Liakou E, Mavrogonatou E, Pratsinis H, Rizou S, Evangelou K, Panagiotou PN, et al. Ionizing radiation-mediated premature senescence and paracrine interactions with cancer cells enhance the expression of syndecan 1 in human breast stromal fibroblasts: the role of TGF-beta. Aging. 2016;8(8):1650–69.
Patel ZS, Grugan KD, Rustgi AK, Cucinotta FA, Huff JL. Ionizing radiation enhances esophageal epithelial cell migration and invasion through a paracrine mechanism involving stromal-derived hepatocyte growth factor. Radiat Res. 2012;177(2):200–8.
Tsai KK, Chuang EY, Little JB, Yuan ZM. Cellular mechanisms for low-dose ionizing radiation-induced perturbation of the breast tissue microenvironment. Cancer Res. 2005;65(15):6734–44.
Kamochi N, Nakashima M, Aoki S, Uchihashi K, Sugihara H, Toda S, et al. Irradiated fibroblast-induced bystander effects on invasive growth of squamous cell carcinoma under cancer-stromal cell interaction. Cancer Sci. 2008;99(12):2417–27.
Rodier F, Coppe JP, Patil CK, Hoeijmakers WA, Munoz DP, Raza SR, et al. Persistent DNA damage signalling triggers senescence-associated inflammatory cytokine secretion. Nat Cell Biol. 2009;11(8):973–9.
Al-Assar O, Demiciorglu F, Lunardi S, Gaspar-Carvalho MM, McKenna WG, Muschel RM, et al. Contextual regulation of pancreatic cancer stem cell phenotype and radioresistance by pancreatic stellate cells. Radiother Oncol. 2014;111(2):243–51.
Barcellos-Hoff MH, Ravani SA. Irradiated mammary gland stroma promotes the expression of tumorigenic potential by unirradiated epithelial cells. Cancer Res. 2000;60(5):1254–60.
Ohuchida K, Mizumoto K, Murakami M, Qian LW, Sato N, Nagai E, et al. Radiation to stromal fibroblasts increases invasiveness of pancreatic cancer cells through tumor-stromal interactions. Cancer Res. 2004;64(9):3215–22.
Tsai KK, Stuart J, Chuang YY, Little JB, Yuan ZM. Low-dose radiation-induced senescent stromal fibroblasts render nearby breast cancer cells radioresistant. Radiat Res. 2009;172(3):306–13.
Arshad A, Deutsch E, Vozenin MC. Simultaneous irradiation of fibroblasts and carcinoma cells repress the secretion of soluble factors able to stimulate carcinoma cell migration. PLoS ONE. 2015;10(1): e0115447.
Steer A, Cordes N, Jendrossek V, Klein D. Impact of cancer-associated fibroblast on the radiation-response of solid xenograft tumors. Front Mol Biosci. 2019;6:70.
Chu TY, Yang JT, Huang TH, Liu HW. Crosstalk with cancer-associated fibroblasts increases the growth and radiation survival of cervical cancer cells. Radiat Res. 2014;181(5):540–7.
Hwang RF, Moore T, Arumugam T, Ramachandran V, Amos KD, Rivera A, et al. Cancer-associated stromal fibroblasts promote pancreatic tumor progression. Cancer Res. 2008;68(3):918–26.
Ji X, Ji J, Shan F, Zhang Y, Chen Y, Lu X. Cancer-associated fibroblasts from NSCLC promote the radioresistance in lung cancer cell lines. Int J Clin Exp Med. 2015;8(5):7002–8.
Wang Y, Gan G, Wang B, Wu J, Cao Y, Zhu D, et al. Cancer-associated fibroblasts promote irradiated cancer cell recovery through autophagy. EBioMedicine. 2017;17:45–56.
Zhang H, Yue J, Jiang Z, Zhou R, Xie R, Xu Y, et al. CAF-secreted CXCL1 conferred radioresistance by regulating DNA damage response in a ROS-dependent manner in esophageal squamous cell carcinoma. Cell Death Dis. 2017;8(5): e2790.
Ebbing EA, van der Zalm AP, Steins A, Creemers A, Hermsen S, Rentenaar R, et al. Stromal-derived interleukin 6 drives epithelial-to-mesenchymal transition and therapy resistance in esophageal adenocarcinoma. Proc Natl Acad Sci U S A. 2019;116(6):2237–42.
Zhang R, Liu F. Cancer-associated fibroblast-derived gene signatures predict radiotherapeutic survival in prostate cancer patients. J Transl Med. 2022;20(1):453.
Kim KH, Chang JS, Byun HK, Kim YB. A novel gene signature associated with poor response to chemoradiotherapy in patients with locally advanced cervical cancer. J Gynecol Oncol. 2022;33(1): e7.
Strell C, Stenmark Tullberg A, Jetne Edelmann R, Akslen LA, Malmstrom P, Ferno M, et al. Prognostic and predictive impact of stroma cells defined by PDGFRb expression in early breast cancer: results from the randomized SweBCG91RT trial. Breast Cancer Res Treat. 2021;187(1):45–55.
Verset L, Tommelein J, Moles Lopez X, Decaestecker C, Boterberg T, De Vlieghere E, et al. Impact of neoadjuvant therapy on cancer-associated fibroblasts in rectal cancer. Radiother Oncol. 2015;116(3):449–54.
Matsuoka Y, Yoshida R, Nakayama H, Nagata M, Hirosue A, Tanaka T, et al. The tumour stromal features are associated with resistance to 5-FU-based chemoradiotherapy and a poor prognosis in patients with oral squamous cell carcinoma. APMIS. 2015;123(3):205–14.
Saigusa S, Toiyama Y, Tanaka K, Yokoe T, Okugawa Y, Fujikawa H, et al. Cancer-associated fibroblasts correlate with poor prognosis in rectal cancer after chemoradiotherapy. Int J Oncol. 2011;38(3):655–63.
Li D, Qu C, Ning Z, Wang H, Zang K, Zhuang L, et al. Radiation promotes epithelial-to-mesenchymal transition and invasion of pancreatic cancer cell by activating carcinoma-associated fibroblasts. Am J Cancer Res. 2016;6(10):2192–206.
Mantoni TS, Lunardi S, Al-Assar O, Masamune A, Brunner TB. Pancreatic stellate cells radioprotect pancreatic cancer cells through beta1-integrin signaling. Cancer Res. 2011;71(10):3453–8.
Pereira PMR, Edwards KJ, Mandleywala K, Carter LM, Escorcia FE, Campesato LF, et al. iNOS regulates the therapeutic response of pancreatic cancer cells to radiotherapy. Cancer Res. 2020;80(8):1681–92.
Tommelein J, De Vlieghere E, Verset L, Melsens E, Leenders J, Descamps B, et al. Radiotherapy-activated cancer-associated fibroblasts promote tumor progression through paracrine IGF1R activation. Cancer Res. 2018;78(3):659–70.
Nicolas AM, Pesic M, Engel E, Ziegler PK, Diefenhardt M, Kennel KB, et al. Inflammatory fibroblasts mediate resistance to neoadjuvant therapy in rectal cancer. Cancer Cell. 2022;40(2):168–84.
Meng J, Li Y, Wan C, Sun Y, Dai X, Huang J, et al. Targeting senescence-like fibroblasts radiosensitizes non-small cell lung cancer and reduces radiation-induced pulmonary fibrosis. JCI Insight. 2021;6(23).
Grinde MT, Vik J, Camilio KA, Martinez-Zubiaurre I, Hellevik T. Ionizing radiation abrogates the pro-tumorigenic capacity of cancer-associated fibroblasts co-implanted in xenografts. Sci Rep. 2017;7:46714.
Hellevik T, Pettersen I, Berg V, Bruun J, Bartnes K, Busund LT, et al. Changes in the secretory profile of NSCLC-associated fibroblasts after ablative radiotherapy: potential impact on angiogenesis and tumor growth. Transl Oncol. 2013;6(1):66–74.
Liu D, Hornsby PJ. Fibroblast stimulation of blood vessel development and cancer cell invasion in a subrenal capsule xenograft model: stress-induced premature senescence does not increase effect. Neoplasia. 2007;9(5):418–26.
van Maaren MC, de Munck L, de Bock GH, Jobsen JJ, van Dalen T, Linn SC, et al. 10 year survival after breast-conserving surgery plus radiotherapy compared with mastectomy in early breast cancer in the Netherlands: a population-based study. Lancet Oncol. 2016;17(8):1158–70.
van Gijn W, Marijnen CA, Nagtegaal ID, Kranenbarg EM, Putter H, Wiggers T, et al. Preoperative radiotherapy combined with total mesorectal excision for resectable rectal cancer: 12-year follow-up of the multicentre, randomised controlled TME trial. Lancet Oncol. 2011;12(6):575–82.
Ramroth J, Cutter DJ, Darby SC, Higgins GS, McGale P, Partridge M, et al. Dose and fractionation in radiation therapy of curative intent for non-small cell lung cancer: meta-analysis of randomized trials. Int J Radiat Oncol Biol Phys. 2016;96(4):736–47.
Demaria S, Guha C, Schoenfeld J, Morris Z, Monjazeb A, Sikora A, et al. Radiation dose and fraction in immunotherapy: One-size regimen does not fit all settings, so how does one choose? Journal for immunotherapy of cancer. 2021;9(4).
Dovedi SJ, Cheadle EJ, Popple AL, Poon E, Morrow M, Stewart R, et al. Fractionated radiation therapy stimulates antitumor immunity mediated by both resident and infiltrating polyclonal T-cell populations when combined with PD-1 blockade. Clin Cancer Res. 2017;23(18):5514–26.
Frey B, Hehlgans S, Rodel F, Gaipl US. Modulation of inflammation by low and high doses of ionizing radiation: Implications for benign and malign diseases. Cancer Lett. 2015;368(2):230–7.
Klug F, Prakash H, Huber PE, Seibel T, Bender N, Halama N, et al. Low-dose irradiation programs macrophage differentiation to an iNOS(+)/M1 phenotype that orchestrates effective T Cell immunotherapy. Cancer Cell. 2013;24(5):589–602.
Fabris VT, Sahores A, Vanzulli SI, Colombo L, Molinolo AA, Lanari C, et al. Inoculated mammary carcinoma-associated fibroblasts: contribution to hormone independent tumor growth. BMC Cancer. 2010;10:293.
Acknowledgements
Not applicable.
Funding
Open access funding provided by UiT The Arctic University of Norway (incl University Hospital of North Norway) This work was supported by grants from the Norwegian Regional Health Authorities (Grant #: HNF1423-18); The Norwegian Cancer Society (project ID#: 198164) and the Aakre Foundation at UiT.
Author information
Authors and Affiliations
Contributions
IM was the main contributor to the conception of the study. IMZ and TH drafted the manuscript and revised it critically for important intellectual content. The two authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Martinez-Zubiaurre, I., Hellevik, T. Cancer-associated fibroblasts in radiotherapy: Bystanders or protagonists?. Cell Commun Signal 21, 108 (2023). https://doi.org/10.1186/s12964-023-01093-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12964-023-01093-5