
Abstract
Background
Previous studies had shown that microRNA-638 (miR-638) exhibited different effects in malignant tumors. Moreover, the function of miR-638 has not been reported in breast cancer. Hence, we designed this research to explore the function of miR-638 in breast cancer.
Methods
Firstly, miR-638 expressions were measured in breast cancer tissues via RT-qPCR. Protein expressions were detected through immunocytochemical (IHC) assay and western blot analysis. Then, Cell Counting Kit-8 (CCK-8) assay and Transwell assay were conducted to observe proliferation and motility of the cells. Dual luciferase assay was performed to confirm the binding site between miR-638 and Homeobox protein Hox-A9 (HOXA9).
Results
Reduced expression of miR-638 was detected in breast cancer. And low miR-638 expression was related to poor prognosis in patients with breast cancer. Functionally, the viability, migration, and invasion of the breast cancer cells were suppressed by miR-638 overexpression. Furthermore, miR-638 can directly bind to HOXA9, and increased expression of HOXA9 was also detected in breast cancer. In particular, HOXA9 upregulation can impair anti-tumor effect of miR-638 in breast cancer, and miR-638 can hinder the Wnt/β-cadherin pathway and epithelial-mesenchymal transition (EMT) in breast cancer.
Conclusion
miR-638 inhibits breast cancer progression through binding to HOXA9.
Similar content being viewed by others

Introduction
Breast cancer usually occurs in the epithelium of the breast gland that seriously endangers women’s lives [1]. Breast cancer usually occurs in women, and only about 1-2% of the patients with breast cancer are men [2]. Breast cancer accounts for 7-10% of human malignant tumors, second only to cervical cancer in women [3]. Among them, the incidence of postmenopausal women ranging from 40 to 60 years old is higher [3]. Similar to a few types of tumors, such as thyroid cancer, the natural development of breast cancer is usually long, and it takes 7 to 8 years for the tumor to reach a sphere with a diameter of 1 cm [4]. Moreover, surgical treatment is the main therapy for breast cancer, and radiotherapy is one of the local treatment methods [5]. Prior to tumor metastasis, surgery and radiation alone can cure the vast majority of breast cancer patients. Once metastasis occurs, routine treatment can only cure a small number of breast cancer patients [6]. In addition, there are many factors influencing the prognosis of breast cancer patients, among which tumor invasion scope and pathological and biological characteristics are the main ones [7]. And the best way to reduce the mortality of breast cancer is early diagnosis and early treatment.
MicroRNAs (miRNAs) have been reported to affect the expressions of their targets through binding with their 3′-UTR. Besides, miRNAs are widely recognized as powerful regulators which involve many processes of breast cancer. For instance, miR-384 was found to inhibit breast cancer progression by targeting ACVR1 [8]. Inversely, miR-374a promoted tumor metastasis and progression by downregulating LACTB and predicts unfavorable prognosis in breast cancer [9]. In addition, it was reported that miR-17-5p could serve as a novel predictor for breast cancer recurrence [10]. Recently, previous studies showed that miR-638 was an important regulator for the occurrence and development of human cancers. For instance, miR-638 was found to restrain the tumorigenesis of gastric cancer [11]. On the contrary, miR-638 was found to promote cell metastasis and inhibit apoptosis in melanoma [12]. miR-638 can also predict the outcome of the patients with non-small cell lung cancer after chemotherapy [13]. Moreover, Zhao et al. found that potentiation of docetaxel sensitivity was regulated by miR-638 via regulation of STARD10 pathway in human breast cancer cells [14]. However, the function of miR-638 in breast cancer remains unclear.
Besides that, it has been found that the activation of Wnt/β-catenin pathway can promote the progression of human cancers [15]. β-catenin belonging to the Wnt/β-catenin signaling pathway was reported to participate in tumorigenesis [16]. Wnt/β-catenin signaling pathways was also identified as therapeutic targets in cancer [17]. Moreover, it was demonstrated that the dysfunction of Wnt and Notch signaling pathways regulated the progression of breast cancer [18]. The activation of oncogene HOXA9 was demonstrated to associate with the prognosis of human glioblastoma [19]. Brumatti G et al. demonstrated that HOXA9 could mediate the survival of myeloid progenitors [20]. HOXA9 had been found to modulate human breast tumor phenotype [21]. In addition, it was found that Wnt/β-catenin pathway was associated with HOXA2 in mouse embryo [22]. However, the functions of Wnt/β-catenin pathway and HOXA9 were not reported in breast cancer.
Moreover, epithelial to mesenchymal transition (EMT) had a great effect on human cancer cell metastasis [23]. Han et al. proposed that miR-30d could mediate breast cancer invasion, migration, and EMT by targeting KLF11 [24]. But the relationship between miR-638 and EMT is still elusive in breast cancer. Therefore, this study attempted to investigate the role of miR-638 in breast cancer, and illustrate the regulation mechanism of miR-638 on breast cancer.
Materials and methods
Experimental samples
The experimental tissues were acquired from fifty-two patients with breast cancer in CaoXian People’s Hospital. These patients provided informed consents and only received surgery before this study. The tissues were frozen in liquid nitrogen and stored in a −80 °C refrigerator. This research was approved by the Institutional Ethics Committee of CaoXian People’s Hospital.
Cell lines culture
The MDA-MB-468, MDA-MB-231, MCF-7 cell lines, and human breast epithelial cell line MCF10A were purchased from ATCC (Manassas, VA). These cell lines were then cultured in DMEM medium with 10% fetal bovine serum (FBS) at 37 °C with 5% CO2.
Cell transfection
miR-638 mimics or inhibitor and negative control (NC, GenePharma, Shanghai, China) were respectively transfected into MCF-7 cells with Lipofectamine 2000 (Invitrogen, Carlsbad, USA).
RT-qPCR analysis
TRIzol reagent (Invitrogen, Carlsbad, USA) was applied to extract total RNA. The reverse transcription of cDNA was performed by PrimeScript reverse transcription kit (Qiagen, Valencia, USA). RT-qPCR was conducted using SYBR green Supermix (Bio-Rad, Richmond, USA) on ABI 7500 Fast system (Applied Biosystems, CA, USA). The internal control for miR-638 or HOXA9 is U6 or GAPDH. And the mRNA expression was calculated by the 2−△△ct method.
CCK-8 assay
The transfected cells (4 × 104) were put in 96-well plates and incubated for 0, 24, 48, and 72 h in an incubator at 37 °C with 5% CO2. Next, each well was added with 10 μl CCK-8 reagents for 2 h (Dojindo, Tokyo, Japan). Finally, the absorbance of each well at 450 nm was detected by a microplate reader (Molecular Devices).
Transwell assay
Cell migration and invasion were detected using Transwell chambers (8-μm pore size membranes). Matrigel (BD Biosciences, USA) was added in the upper surface for observing cellular invasion. The lower chamber was added with 10% FBS. MCF-7 cells with miR-638 mimic or inhibitor were incubated in the upper chamber with serum-free medium for 48 h at 37 °C with 5% CO2. The moving cells were fixed with methanol and stained with crystal violet. Then, the number of removed cells was observed using a microscope.
Dual luciferase assay
The psiCHECK-2 vector (Promega, Madison, USA) containing the 3′-UTR of wild type or mutant HOXA9 was constructed. Then, the whole vector and miR-638 mimics were transfected into MCF-7 cells. The luciferase activity was detected using dual luciferase assay system (Promega, USA).
Immunohistochemistry (IHC)
After dewaxing, hydrating, and washing the ovary tissues section, the cells were blocked with 5% goat serum (diluted in PBS). Next, the cells were incubated with anti-HOXA9 (nucleus) antibody at 37 °C for 1-2 h. Then, the cells were incubated with appropriate secondary antibodies at 37 °C for 1 h. After washing, the color development of this section was performed using DAB mixture. Next, we washed, counterstained, dehydrated, transparentized, and mounted the section. Microscope was used to capture images.
Western blot analysis
The protein samples were obtained using RIPA lysis buffer. The protein was separated by 10% SDS-PAGE. After blocking with 5% skim milk, the protein was transferred in PVDF membranes. The membranes were then incubated with E-cadherin, N-cadherin, vimentin, β-cadherin, p-β-cadherin, HOXA9, and GAPDH antibodies overnight at 4°C. The protein was then incubated with secondary antibodies for 2 h at room temperature. Finally, protein expression was detected by Electro-Chemi-Luminescence system (ECL, Pierce Biotechnology, USA).
Statistical analysis
Data are analyzed using SPSS 19.0 or Graphpad Prism 6. Chi-squared test was adopted to analyze the relationship between HOXA9 and clinical features in breast cancer patients. Tukey’s one-way ANOVA was used to calculate the difference between multiple groups. The survival differences were compared by Kaplan-Meier analysis (log-rank test). P < 0.05 indicates significant difference.
Results
miR-638 expression was decreased in breast cancer tissues
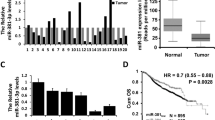
Firstly, miR-638 expressions were observed in breast cancer tissues. Compared to normal tissues, miR-638 expressions were downregulated in breast cancer tissues (Fig. 1A). Moreover, miR-638 was found to be associated with tumor size, TNM stage, and lymph node metastasis (Table 1). Furthermore, low miR-638 expression predicted unfavorable prognosis in breast cancer patients (Fig. 1B). The results suggested that abnormal miR-638 expression was related to the tumorigenesis and prognosis of breast cancer.

miR-638 expression was decreased in breast cancer tissues. (A) The expressions of miR-638 in breast cancer tissues. (B) High miR-638 expression was correlated with longer overall survival of breast cancer patients. *P < 0.05, **P < 0.01
Overexpression of miR-638 inhibited breast cancer progression
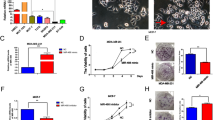
Then, miR-638 expressions were detected in MDA-MB-468, MDA-MB-231, MCF-7, and MCF10A cell lines. Similarly, downregulation of miR-638 was also identified in MDA-MB-468, MDA-MB-231, and MCF-7 cell lines in contrast to MCF10A cells (Fig. 2A). Next, miR-638 mimics or inhibitors were transfected into MCF-7 cells to detect its role in breast cancer. The transfection efficiency was detected by RT-qPCR (Fig. 2B). Functionally, miR-638 overexpression inhibited cell proliferation in breast cancer (Fig. 2C). On the contrary, the knockout of miR-638 promoted the proliferation of MCF-7 cells (Fig. 2D). Moreover, miR-638 overexpression also repressed cell migration while knockout of miR-638 promoted the migration of MCF-7 cells (Fig. 2E). Similarly, the same results of cell invasion regulated by miR-141-3p were also identified in MCF-7 cells (Fig. 2F). All these results suggested that miR-638 overexpression inhibited breast cancer progression.

Overexpression of miR-638 inhibited the progression of breast cancer. (A) The miR-638 expression in MCF-7, MDA-MB-231, MDA-MB-468, and MCF10A cell lines. (B) The expression of miR-638 was examined in MCF-7 cells with miR-638 mimics or inhibitor. (C, D) The cell proliferation was measured in cells containing miR-638 mimics or inhibitor. (E, F) Cell migration and invasion analysis in cells containing miR-638 mimics or inhibitor *P <0.05, **P <0.01
miR-638 can directly bind to HOXA9 in breast cancer
The downstream target of miR-638 was identified in this study. HOXA9 was found to have binding sites with miR-638 predicted by TargetScan (http://www.targetscan.org/) (Fig. 3A). Then, luciferase reporter assay suggested that miR-638 mimics decreased the luciferase activity of Wt-HOXA9. But miR-638 mimics had little effect on that of Mut-HOXA9 (Fig. 3B). Moreover, HOXA9 expression was found to have negative correlation with miR-638 in breast cancer tissues (R2 = 0.4964, Fig. 3C). We also found that miR-638 mimics declined HOXA9 expression (Fig. 3D), and miR-638 inhibitor enhanced HOXA9 expression in MCF-7 cells (Fig. 3E). In a word, miR-638 can directly bind to HOXA9 and negatively regulate HOXA9 expression in breast cancer.

HOXA9 was a direct target of miR-638 in breast cancer. (A) HOXA9 had binding sites with miR-638. (B) Luciferase reporter assay. (C) miR-638 had negative correlation with HOXA9. (D, E) The expression of HOXA9 were observed in MCF-7 cells containing miR-638 mimics or inhibitor **P <0.01
HOXA9 was involved in breast cancer progression
Next, HOXA9 expression was identified in breast cancer tissues. IHC showed that positive detection of HOXA9 protein expression was in the nucleus of breast cancer cells (Fig. 4A). Moreover, the upregulation of HOXA9 was detected in breast cancer tissues (Fig. 4B). Furthermore, breast cancer patients with high HOXA9 expression had shorter overall survival (Fig. 4C). According to these findings, HOXA9 was considered to participate in the pathogenesis and prognosis of breast cancer.

HOXA9 was upregulated in breast cancer tissues. (A, B) The protein expression of HOXA9 in breast cancer tissues detected by immunohistochemistry. (C) High HOXA9 expression was related to shorter overall survival of breast cancer patients. *P < 0.05
miR-638 impeded the progression of breast cancer through targeting HOXA9
Then, miR-638 mimics and HOXA9 vector were transfected in MCF-7 cells to explore the relationship between miR-638 and HOXA9. The qRT-PCR assay suggested that the reduced HOXA9 expression induced by miR-638 mimics was recovered by HOXA9 vector in MCF-7 cells (Fig. 5A). Moreover, HOXA9 overexpression impaired the suppressive effect of miR-638 on MCF-7 cell proliferation (Fig. 5B). Consistently, the same results were also identified for cell migration (Fig. 5C) and invasion (Fig. 5D) in breast cancer. In brief, miR-638 was speculated to impede breast cancer progression by targeting HOXA9.

miR-638 impeded the progression of breast cancer through targeting HOXA9. (A) The expression of HOXA9 was measured in MCF-7 cells with HOXA9 vector and miR-638. (B) The cell proliferation was measured in MCF-7 cells with HOXA9 vector and miR-638. (C, D) The cell migration and invasion in MCF-7 cells with HOXA9 vector and miR-638 **P < 0.01
miR-638 hindered EMT and Wnt/β-cadherin pathway in breast cancer
Furthermore, we investigated the effect of miR-638 on EMT and Wnt/β-cadherin pathway. Upregulation of miR-638 was found to promote E-cadherin expression and inhibit N-cadherin and Vimentin expressions in MCF-7 cells (Fig. 6). And downregulation of miR-638 showed opposite results (Fig. 6). Moreover, miR-638 overexpression inhibited p-β-cadherin expression (Fig. 6), while miR-638 downregulation enhanced p-β-cadherin expression (Fig. 6). But β-cadherin expression was not affected by miR-638 in MCF-7 cells. Therefore, miR-638 may inhibit breast cancer tumorigenesis by hindering EMT and Wnt/β-cadherin pathway.

miR-638 hindered EMT and Wnt/β-cadherin pathway in breast cancer. Western blot analysis of E-cadherin, N-cadherin, Vimentin, β-cadherin, and p-β-cadherin in MCF-7 cells contained miR-638 mimics or inhibitor
Discussion
The global incidence of breast cancer has been on the rise since the late 1970s. Although China is not a country with high incidence of breast cancer, it should not be optimistic. In recent years, the growth rate of breast cancer incidence in China is 1-2% higher than that in countries with high incidence of breast cancer [25]. Therefore, people pay more and more attention to breast cancer. Recently, increasing miRNAs have been reported to participate in the pathogenesis of breast cancer, such as miR-22 [26] and miR-1271 [27]. In present study, downregulation of miR-638 was identified in breast cancer. And miR-638 exhibited suppressive effect on the development of breast cancer.
MiRNAs serve as important participants in regulating the behaviors of the cells, such as differentiation and proliferation. In recent 10 years, increasingly studies have indicated that miRNA dysfunction is one of the major causes leading the malignant progression of the tumors. In breast cancer, downregulated miR-143 is related with the poor prognosis of the patients with breast cancer, and miR-143 can effectively inhibit the malignant behaviors of the breast cancer cells via targeting HMGA2 [28]. In this study, miR-638 downregulation was related to worse prognosis in breast cancer patients. In previous studies, low miR-638 expression was found in hepatocellular carcinoma and related to its clinical significance [29]. Similarly, decreased miR-638 expression was also related to the clinic pathological characteristics in patients with breast cancer. Shi et al. proposed that decreased levels of miR-638 predicted poor prognosis in hepatocellular carcinoma [30]. In addition, miR-638 inhibited cell proliferation and invasion in human colorectal cancer [31], which is consistent with our results. Besides, miR-638 downregulation promoted EMT and cell invasion in colorectal carcinoma [32]. The same results were also identified in this study. Furthermore, it was observed that overexpression of miR-638 could hinder EMT and Wnt/β-cadherin pathways in breast cancer. Similarly, it was indicated that miR-638 inhibited the Wnt/β-catenin pathway in cervical cancer [33]. Furthermore, miR-638 directly binds to HOXA9 in breast cancer, and miR-638 impeded breast cancer progression through targeting HOXA9.
HOXA9 is a kind of homeoproteins which abnormally expressed aberrantly in human cancers [34]. Liu et al. proposed that HOXA9 was upregulated and functioned as an oncogene in chronic myeloid leukemogenesis [35]. In current research, we also found the upregulation of HOXA9 in breast cancer. Additionally, HOXA9 has been reported to be regulated by some miRNAs, such as miR-126 [36], miR-155 [37], and miR-196b [38]. Same as our findings, Wang et al. demonstrated that miR-133b suppressed metastasis by targeting HOXA9 in human colorectal cancer [39]. Zhang et al. indicated that miR-182 blocked Wnt/β-catenin signaling and suppressed cell proliferation in human osteosarcoma via targeting HOXA9 [40]. The dysfunction of Wnt/β-catenin pathway is involved in the formation and development of breast cancer. Jiang et al. have indicated that miR-449b-5p-mediated inactivation of Wnt/β-catenin pathway could effectively reduce the proliferation and invasion of the breast cancer cells [41]. In this study, miR-638 was found to suppress the tumorigenesis of breast cancer through targeting HOXA9 and suppressing Wnt/β-cadherin pathway. Therefore, this study suggested that miR-638 served as a tumor inhibitor to impede the malignant progression of breast cancer via suppressing the expression of HOXA9 and activation of Wnt/β-cadherin pathway.
Conclusion
In conclusion, miR-638 downregulation was identified in breast cancer and related to adverse prognosis in breast cancer patient. Moreover, miR-638 inhibited breast cancer progression via targeting HOXA9 and suppressing EMT/Wnt/β-cadherin pathway. The findings may provide new thoughts for the diagnosis and therapies of breast cancer.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Yang J, Zhang JY, Chen J, Chen C, Song XM, Xu Y, et al. Prognostic role of microRNA-145 in various human malignant neoplasms: a meta-analysis of 18 related studies. World J Surg Oncol. 2014;12(1):254. https://doi.org/10.1186/1477-7819-12-254.
Liang F, Qu H, Lin Q, Yang Y, Ruan X, Zhang B, et al. Molecular biomarkers screened by next-generation RNA sequencing for non-sentinel lymph node status prediction in breast cancer patients with metastatic sentinel lymph nodes. World J Surg Oncol. 2015;13(1):258. https://doi.org/10.1186/s12957-015-0642-2.
Qiu Z, Wang L, Liu H. Hsa_circ_0001982 promotes the progression of breast cancer through miR-1287-5p/MUC19 axis under hypoxia. World J Surg Oncol. 2021;19(1):161. https://doi.org/10.1186/s12957-021-02273-8.
Mao XH, Ye Q, Zhang GB, Jiang JY, Zhao HY, Shao YF, et al. Identification of differentially methylated genes as diagnostic and prognostic biomarkers of breast cancer. World J Surg Oncol. 2021;19(1):29. https://doi.org/10.1186/s12957-021-02124-6.
Perri F, Longo F, Giuliano M, Sabbatino F, Favia G, Ionna F, et al. Epigenetic control of gene expression: potential implications for cancer treatment. Crit Rev Oncol Hematol. 2017;111:166–72. https://doi.org/10.1016/j.critrevonc.2017.01.020.
Li Y, Shan F, Chen J. Lipid raft-mediated miR-3908 inhibition of migration of breast cancer cell line MCF-7 by regulating the interactions between AdipoR1 and Flotillin-1. World J Surg Oncol. 2017;15(1):69. https://doi.org/10.1186/s12957-017-1120-9.
Zhou M, Zhong L, Xu W, Sun Y, Zhang Z, Zhao H, et al. Discovery of potential prognostic long non-coding RNA biomarkers for predicting the risk of tumor recurrence of breast cancer patients. Sci Rep. 2016;6(1):31038. https://doi.org/10.1038/srep31038.
Wang Y, Zhang Z, Wang J. MicroRNA-384 inhibits the progression of breast cancer by targeting ACVR1. Oncol Rep. 2018;39(6):2563–74. https://doi.org/10.3892/or.2018.6385.
Zhang J, He Y, Yu Y, Chen X, Cui G, Wang W, et al. Upregulation of miR-374a promotes tumor metastasis and progression by downregulating LACTB and predicts unfavorable prognosis in breast cancer. Cancer Med. 2018;7(7):3351–62. https://doi.org/10.1002/cam4.1576.
Wang Y, Li J, Dai L, Zheng J, Yi Z, Chen L. MiR-17-5p may serve as a novel predictor for breast cancer recurrence. Cancer Biomark. 2018;22(4):721–6. https://doi.org/10.3233/CBM-181228.
Shen Y, Chen H, Gao L, Zhang W, He J, Yang X, et al. MiR-638 acts as a tumor suppressor gene in gastric cancer. Oncotarget. 2017;8(64):108170–80. https://doi.org/10.18632/oncotarget.22567.
Bhattacharya A, Schmitz U, Raatz Y, Schonherr M, Kottek T, Schauer M, et al. miR-638 promotes melanoma metastasis and protects melanoma cells from apoptosis and autophagy. Oncotarget. 2015;6(5):2966–80. https://doi.org/10.18632/oncotarget.3070.
Wang F, Lou JF, Cao Y, Shi XH, Wang P, Xu J, et al. miR-638 is a new biomarker for outcome prediction of non-small cell lung cancer patients receiving chemotherapy. Exp Mol Med. 2015;47:e162.
Zhao G, Li Y, Wang T. Potentiation of docetaxel sensitivity by miR-638 via regulation of STARD10 pathway in human breast cancer cells. Biochem Biophys Res Commun. 2017;487(2):255–61. https://doi.org/10.1016/j.bbrc.2017.04.045.
Li CY, Cui ZS, Lu Y, Zhang Y, Gao J, Wang EH. The roles of important molecules of Wnt signaling pathway in non-small-cell lung cancer. Zhonghua Bing Li Xue Za Zhi. 2005;34(9):599–600.
Umbreit C, Aderhold C, Faber A, Sommer JU, Sauter A, Hofheinz RD, et al. Unexpected alteration of beta-catenin and c-KIT expression by 5-FU and docetaxel in p16-positive squamous cell carcinoma compared to HPV-negative HNSCC cells in vitro. Anticancer Res. 2013;33(6):2457–65.
Anastas JN, Moon RT. WNT signalling pathways as therapeutic targets in cancer. Nat Rev Cancer. 2013;13(1):11–26. https://doi.org/10.1038/nrc3419.
Zardawi SJ, O’Toole SA, Sutherland RL, Musgrove EA. Dysregulation of Hedgehog, Wnt and Notch signalling pathways in breast cancer. Histol Histopathol. 2009;24(3):385–98. https://doi.org/10.14670/HH-24.385.
Costa BM, Smith JS, Chen Y, Chen J, Phillips HS, Aldape KD, et al. Reversing HOXA9 oncogene activation by PI3K inhibition: epigenetic mechanism and prognostic significance in human glioblastoma. Cancer Res. 2010;70(2):453–62. https://doi.org/10.1158/0008-5472.CAN-09-2189.
Brumatti G, Salmanidis M, Kok CH, Bilardi RA, Sandow JJ, Silke N, et al. HoxA9 regulated Bcl-2 expression mediates survival of myeloid progenitors and the severity of HoxA9-dependent leukemia. Oncotarget. 2013;4(11):1933–47. https://doi.org/10.18632/oncotarget.1306.
Gilbert PM, Mouw JK, Unger MA, Lakins JN, Gbegnon MK, Clemmer VB, et al. HOXA9 regulates BRCA1 expression to modulate human breast tumor phenotype. J Clin Invest. 2010;120(5):1535–50. https://doi.org/10.1172/JCI39534.
Donaldson IJ, Amin S, Hensman JJ, Kutejova E, Rattray M, Lawrence N, et al. Genome-wide occupancy links Hoxa2 to Wnt-beta-catenin signaling in mouse embryonic development. Nucleic Acids Res. 2012;40(9):3990–4001. https://doi.org/10.1093/nar/gkr1240.
Kalluri R, Weinberg RA. The basics of epithelial-mesenchymal transition. J Clin Invest. 2009;119(6):1420–8. https://doi.org/10.1172/JCI39104.
Han M, Wang Y, Guo G, Li L, Dou D, Ge X, et al. microRNA-30d mediated breast cancer invasion, migration, and EMT by targeting KLF11 and activating STAT3 pathway. J Cell Biochem. 2018;119(10):8138–45. https://doi.org/10.1002/jcb.26767.
Zuo TT, Zheng RS, Zeng HM, Zhang SW, Chen WQ. Female breast cancer incidence and mortality in China, 2013. Thorac Cancer. 2017;8(3):214–8. https://doi.org/10.1111/1759-7714.12426.
Liu H, Huang X, Ye T. MiR-22 down-regulates the proto-oncogene ATP citrate lyase to inhibit the growth and metastasis of breast cancer. Am J Transl Res. 2018;10(3):659–69.
Du HY, Liu B. MiR-1271 as a tumor suppressor in breast cancer proliferation and progression via targeting SPIN1. Eur Rev Med Pharmacol Sci. 2018;22(9):2697–706. https://doi.org/10.26355/eurrev_201805_14966.
Mansoori B, Duijf PHG, Mohammadi A, Safarzadeh E, Ditzel HJ, Gjerstorff MF, et al. MiR-142-3p targets HMGA2 and suppresses breast cancer malignancy. Life Sci. 2021;276:119431. https://doi.org/10.1016/j.lfs.2021.119431.
Cheng J, Chen Y, Zhao P, Li N, Lu J, Li J, et al. Dysregulation of miR-638 in hepatocellular carcinoma and its clinical significance. Oncol Lett. 2017;13(5):3859–65. https://doi.org/10.3892/ol.2017.5882.
Shi M, Jiang Y, Yang L, Yan S, Wang YG, Lu XJ. Decreased levels of serum exosomal miR-638 predict poor prognosis in hepatocellular carcinoma. J Cell Biochem. 2018;119(6):4711–6. https://doi.org/10.1002/jcb.26650.
Zhang J, Fei B, Wang Q, Song M, Yin Y, Zhang B, et al. MicroRNA-638 inhibits cell proliferation, invasion and regulates cell cycle by targeting tetraspanin 1 in human colorectal carcinoma. Oncotarget. 2014;5(23):12083–96. https://doi.org/10.18632/oncotarget.2499.
Ma K, Pan X, Fan P, He Y, Gu J, Wang W, et al. Loss of miR-638 in vitro promotes cell invasion and a mesenchymal-like transition by influencing SOX2 expression in colorectal carcinoma cells. Mol Cancer. 2014;13(1):118. https://doi.org/10.1186/1476-4598-13-118.
Wei H, Zhang JJ, Tang QL. MiR-638 inhibits cervical cancer metastasis through Wnt/beta-catenin signaling pathway and correlates with prognosis of cervical cancer patients. Eur Rev Med Pharmacol Sci. 2017;21(24):5587–93. https://doi.org/10.26355/eurrev_201712_13999.
Collins CT, Hess JL. Role of HOXA9 in leukemia: dysregulation, cofactors and essential targets. Oncogene. 2016;35(9):1090–8. https://doi.org/10.1038/onc.2015.174.
Liu Y, Zheng W, Song Y, Ma W, Yin H. Low expression of miR-196b enhances the expression of BCR-ABL1 and HOXA9 oncogenes in chronic myeloid leukemogenesis. PLoS One. 2013;8(7):e68442. https://doi.org/10.1371/journal.pone.0068442.
Shen WF, Hu YL, Uttarwar L, Passegue E, Largman C. MicroRNA-126 regulates HOXA9 by binding to the homeobox. Mol Cell Biol. 2008;28(14):4609–19. https://doi.org/10.1128/MCB.01652-07.
Hu YL, Fong S, Largman C, Shen WF. HOXA9 regulates miR-155 in hematopoietic cells. Nucleic Acids Res. 2010;38(16):5472–8. https://doi.org/10.1093/nar/gkq337.
Yu SL, Lee DC, Sohn HA, Lee SY, Jeon HS, Lee JH, et al. Homeobox A9 directly targeted by miR-196b regulates aggressiveness through nuclear Factor-kappa B activity in non-small cell lung cancer cells. Mol Carcinog. 2016;55(12):1915–26. https://doi.org/10.1002/mc.22439.
Wang X, Bu J, Liu X, Wang W, Mai W, Lv B, et al. miR-133b suppresses metastasis by targeting HOXA9 in human colorectal cancer. Oncotarget. 2017;8(38):63935–48. https://doi.org/10.18632/oncotarget.19212.
Zhang ZF, Wang YJ, Fan SH, Du SX, Li XD, Wu DM, et al. MicroRNA-182 downregulates Wnt/beta-catenin signaling, inhibits proliferation, and promotes apoptosis in human osteosarcoma cells by targeting HOXA9. Oncotarget. 2017;8(60):101345–61. https://doi.org/10.18632/oncotarget.21167.
Jiang J, Yang X, He X, Ma W, Wang J, Zhou Q, et al. MicroRNA-449b-5p suppresses the growth and invasion of breast cancer cells via inhibiting CREPT-mediated Wnt/β-catenin signaling. Chem Biol Interact. 2019;302:74–82. https://doi.org/10.1016/j.cbi.2019.02.004.
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
QX designed the study and drafted the manuscript. QZ and MD were responsible for the collection and analysis of the experimental data. QX and YY revised the manuscript critically for important intellectual content. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was approved by the Ethics Committee of Weifang People’s Hospital, China. Signed written informed consents were obtained from the patients and/or guardians.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Xu, Q., Zhang, Q., Dong, M. et al. MicroRNA-638 inhibits the progression of breast cancer through targeting HOXA9 and suppressing Wnt/β-cadherin pathway. World J Surg Onc 19, 247 (2021). https://doi.org/10.1186/s12957-021-02363-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12957-021-02363-7