
Abstract
Pancreatic ductal adenocarcinoma (PDAC) is one of the most fatal cancers worldwide with high mortality, which is mainly due to the lack of reliable biomarkers for PDAC diagnosis/prognosis in the early stages and effective therapeutic strategies for the treatment. Cancer-derived small extracellular vesicles (sEVs), which carry various messages and signal biomolecules (e.g. RNAs, DNAs, proteins, lipids, and glycans) to constitute the key features (e.g. genetic and phenotypic status) of cancer cells, are regarded as highly competitive non-invasive biomarkers for PDAC diagnosis/prognosis. Additionally, new insights on the biogenesis and molecular functions of cancer-derived sEVs pave the way for novel therapeutic strategies based on cancer-derived sEVs for PDAC treatment such as inhibition of the formation or secretion of cancer-derived sEVs, using cancer-derived sEVs as drug carriers and for immunotherapy. This review provides a comprehensive overview of the most recent scientific and clinical research on the discovery and involvement of key molecules in cancer-derived sEVs for PDAC diagnosis/prognosis and strategies using cancer-derived sEVs for PDAC treatment. The current limitations and emerging trends toward clinical application of cancer-derived sEVs in PDAC diagnosis/prognosis and treatment have also been discussed.
Similar content being viewed by others


Introduction
Pancreatic ductal adenocarcinoma (PDAC), accounting for more than 90% of pancreatic cancers, is one of the most aggressive malignancies with a 5-year survival rate of 8-9%.[1,2,3,4,5,6] It has been reported that only 15–20% of PDAC patients can be surgically resected; the other 80–85% of patients present with unresectable metastatic or locally progressed diseases.[7, 8] Most PDAC patients still suffer local recurrence or systematic metastasis in 12 months after surgery, with an overall 5-year survival rate between 20% and 30%.[9] There were over 55,989 new cases (29,673 men and 26,316 women) of PDAC diagnosed in the United States in 2022 according to cancer statistics of American Cancer Society.[10] And PDAC has been estimated to surpass colon cancer as the second leading cause of cancer-related death in the United States by 2030.[11] The majority of patients failed to be diagnosed in the early stage of PDAC, which caused approximately 48,220 deaths.[12] Despite various factors that influence cancer patient outcomes, there remain two essential problems, namely reliable PDAC diagnosis/prognosis and effective treatment regimes. The poor diagnosis/prognosis of PDAC is attributed to multiple reasons, including non-specific symptoms or even no symptoms in the early stage of PDAC, lack of sensitive and specific PDAC biomarkers, and difficulties in imaging early-stage tumors.[5] While the treatment failure of PDAC is typically caused by its insidious onset, high invasiveness and metastasis[13], detecting PDAC at an early stage is crucial to improve the therapeutic effect and thereby significantly increase the overall survival of PDAC patients.
Currently, there are no validated and specific tests to reliably diagnose PDAC in clinic, particularly during early stages. PDAC is usually diagnosed by biochemical examination, imaging examination and tissue biopsy.[14] The most extensively evaluated biomarker for biochemical examination of PDAC is carbohydrate antigen 19−9 (CA19-9). However, CA19-9 has insufficient sensitivity and specificity to distinguish the PDAC patients from healthy people or patients with other pancreatic disease (chronic pancreatitis, acute pancreatitis, etc.).[15, 16] Studies have shown that multiple biomarkers provide more accurate results than individual biomarkers.[17, 18] For example, Shreya et al. identified a diagnostic panel of 4 serum biomarkers (S100A2, A100A4, CA-125 and CA19-9) which had higher diagnostic potential (AUC 0.913) than CA19-9 alone (AUC 0.869) in a small study of 120 PDAC patients and 80 healthy controls.[19] However, larger clinical trials are still essential to validate its accuracy and investigate the potential for early PDAC detection. Imaging examination, such as contrast-enhanced computed tomography (CT), magnetic resonance imaging (MRI), and endoscopic ultrasound are quite expensive and inefficient to detect early lesions or to differentiate benign from malignant lesions.[20] On the other hand, tissue biopsies are invasive, show low sensitivity and require specialized surgical skills and facilities for sampling.[21, 22] Therefore, PDAC patients can only gain very limited benefit from the advanced surgical techniques, perioperative management and oncological treatments due to the weaknesses of the current diagnostic methods for early PDAC diagnosis. There is an urgent need for reliable, specific and sensitive PDAC biomarkers and diagnosis methods to improve the diagnostic accuracy of PDAC at early stages.
Earlier diagnosis of cancer would give patients more time for treatment, but patient outcomes will not be significantly improved without efficient treatment plans. The currently available therapeutic options for PDAC involve the combination of chemotherapy, surgery, radiation and immunotherapy, most of which are palliative treatments aiming to relieve the symptoms and prolong the patient survival rate.[23] However, PDAC can survive under these harsh conditions and increase proliferative ability because of its genetic and metabolic remodeling.[24, 25] Furthermore, a dense and diffuse stroma forming around the tumor can increase its resistance to treatments and affect the tumor progression.[25] PDAC may also develop chemoresistance during treatment due to tumor heterogeneity and plasticity.[26, 27] These characteristics make PDAC resistant to traditional treatment approaches and lead to poor clinical outcomes, thus innovative therapies are required to improve the prospects of PDAC patients. Fortunately, researchers have discovered emerging biomarkers and treatment candidates for both PDAC diagnosis and therapy after intensive studies of extracellular vesicles (EVs) in recent years.
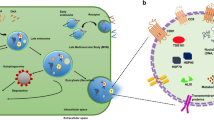
EVs released from a variety of cell types are classified into three broad groups according to their size, pathway of origin, and biomolecules: small EVs (sEVs) (namely exosomes, 40–200nm) (Fig.1), microvesicles (microparticles or ectosomes, 50-2000nm) and apoptotic bodies (500–4000nm).[28, 29] In the past decade, sEVs have attracted worldwide attention among researchers from various fields of life sciences because of their special and important roles in various biological functions (angiogenesis, cell apoptosis, inflammation and immune regulation, etc.) at normal physiological condition as well as pathological condition, determining by which cells they originate as well as the status of these cells at time of sEV generation.[30] sEVs derived from cancer cells played a crucial role in PDAC biology, including tumorigenesis, cancer progression, cancer metastasis, immune regulation and therapeutic resistance, etc., showing great value in cancer studies.[31, 32] Cancer-derived sEVs are small, lipid bilayer membrane vesicles generated inside the cell in multivesicular bodies (MVBs), which release cancer-derived sEVs into the extracellular microenvironment by fusion with the cell membrane (Fig.1).[1, 33] These cancer-derived small vesicles contain numerous biomolecules including DNAs, RNAs, proteins, glycans and lipids, which can be transported from donor cells to other recipient cells (adjacent or distant cells) mainly by receptor-ligand binding, endocytosis and direct fusion, to establish a desired small-scale environment for modifying the functions (gene expression, signaling, and overall functions) in states of cancers.[34] Although the mechanism of cancer-derived sEV for tumorigenesis is complex, it is generally accepted that the interactions of cancer associated proteins and oncogenes between cancer cells and healthy cells promote the process. [35] These cancer associated biomolecules in cancer derived sEVs can activate the signal transduction pathways and induce cellular change within recipient cells to regulate cancer growth and metastasis.[36] Body fluids such as blood in cancer patients contain diverse mixture of EV subsets, among which cancer-derived sEVs (~ 23–66% of total sEVs in plasma) are substantial and important EV subset, acting as an indicator of tumor and holding a significant potential to serve as a liquid biopsy tool for cancer diagnosis/prognosis.[37,38,39,40] These sEVs secreted by cancer cells can be collected and provide the dynamic information from the tumors at the time of blood drawing. Thus, cancer-derived sEVs are promising cancer biomarkers for non-invasive cancer diagnosis/prognosis.[41,42,43,44] In addition, cancer-derived sEVs have also been explored for their use in cancer therapeutics. Using sEVs in therapeutics has been studied for preventing the formation and release of cancer-derived sEVs; using cancer-derived sEVs as drug delivery vesicles; as well as using cancer-derived sEVs in immunotherapy. Compared with synthetic nanoparticles, cancer-derived sEVs encompass several desirable attributes: intrinsic ability to carry biomolecules such as RNAs, DNAs, and proteins ; immune tolerance when using autologous-derived sEVs; desirable stability in body fluids; natural targeting property of cancer cells; ease of surface modification and ability to cross biological barriers, such as the blood-brain barrier (BBB).[45,46,47] Overall, these advantageous features allow cancer-derived sEVs to be promising candidates for providing novel therapeutic strategies for cancer treatment.

Biogenesis and identification of small extracellular vesicles (sEVs). sEVs originate from the endosomal pathway by the formation of endosomes and MVBs. When MVBs fuse with cell membrane, sEVs are released into extracellular milieu. sEVs are composed of a lipid bilayer vesicle containing nucleic acids, proteins, lipids, glycans, and other small molecules
Cancer-derived sEVs have been reported to play different roles in PDAC, including cancer initiation, progression, metastasis, drug resistance, cancer diagnosis/prognosis and treatment. [32, 35, 48] In this review, we will mainly focus on the most recent progress in the use of key molecules of cancer-derived sEVs as emerging biomarkers for PDAC diagnosis/prognosis and cancer-derived sEVs based therapeutic strategies for PDAC treatment, as well as discussing the current hurdles and perspectives for further clinical applications, with the aim of gaining new insights for researchers working on sEVs in cancer diagnosis/prognosis and treatment.
PDAC diagnosis/prognosis
Given the absence of non-invasive and robust biomarkers for PDAC diagnosis, there has recently been significant interest in the use of PDAC-derived sEVs as biomarkers due to their diverse molecular contents. The biogenesis of sEVs enables the packing of these complex extracellular and intracellular molecular contents into sEVs (or on the surface of sEVs) in a cell specific manner.[49] These molecular contents can reflect the key features of cells from which they originate.[50] As the contents of PDAC-derived sEVs are cell-type specific, PDAC-derived sEVs may provide a unique ‘signature’ of genetic and phenotypic status of the tumor.[51] This molecular signature is able to discriminate cancer-derived sEVs from different types of cancer cells, as well as distinguish cancer-derived sEVs from healthy sEVs. Cancer-derived sEVs also carry specific oncogenes and oncoproteins (mutant KRAS, etc.), which can be used to detect cancer-derived sEVs from other sources of sEVs as well.[52] Moreover, sEVs secreted by PDAC cells can be easily collected from body fluids, such as blood. Under the protection of the endogenous membrane of the sEVs, these biomolecules carried by sEVs can remain stable inside the blood circulation, which makes the PDAC diagnosis/prognosis more reliable. PDAC-derived sEVs in blood can be enriched using different isolation approaches (e.g. ultracentrifugation, immunoaffinity isolation, polymeric precipitation isolation and size exclusion chromatography) and their molecular components (e.g. RNAs, DNAs, proteins, lipids and glycans) can be analyzed by corresponding techniques (e.g. polymerase chain reaction, gel electrophoresis, flow cytometry and mass spectroscopy) for PDAC diagnosis/prognosis (Fig.2). There are plenty of excellent review articles [30, 31, 49, 53] on sEVs’ origin, isolation, characterization and analysis techniques, which are not the focus of this review article. Here we will initially focus on the key molecules that are carried by PDAC-derived sEVs and discuss their potential as biomarkers in PDAC diagnosis/prognosis .

Molecular analysis of components in/on sEVs for PDAC diagnosis/prognosis. PDAC-derived sEVs circulating in blood can be enriched by techniques such as ultracentrifugation. Molecular components including RNAs, DNAs, proteins, lipids and glycans can be analyzed to generate the unique molecular signature for PDAC diagnosis/prognosis

Schematic of the formation and release of sEVs. MVB biogenesis is associated with ESCRT-dependent and ESCRT-independent pathways, agents such as ROCK, RAB, SNARE, Ca2+ affect the release of sEVs from cells
RNAs
PDAC-derived sEVs contain different forms of RNAs, including micro RNAs (miRNAs), messenger RNAs (mRNAs), long non-coding RNAs (lncRNAs) and circular RNAs (circRNAs).[54, 55] RNAs in sEVs have been extensively studied, due to their critical roles in regulating almost all aspects of cancer-related metabolism and function.[56] Among these RNAs, miRNAs are the most studied target for cancer diagnosis/prognosis, whereas other RNAs may also act as potential biomarkers for PDAC diagnosis/prognosis. [57] Table1 provides the reported RNA biomarkers for PDAC diagnosis/prognosis.
miRNAs are small, noncoding RNAs of ~ 19–24 nucleotide length and regulate about 70% of mRNA transcripts in humans, playing vital roles in a variety of cellular processes such as cancer development. [78, 79] Oncogenic and cancer-suppressor miRNAs in sEVs may be of high diagnostic/prognostic value in PDAC because of their differential expression between cancer cells and normal cells. For instance, Takahasi et al. profiled the expression of miRNAs of sEVs from plasma of 50 PDAC patients and 20 healthy volunteers using real-time quantitative reverse transcription (qRT-PCR) and found that miR-451a showed higher upregulation in the patients and was associated with PDAC stages.60 In another study, Xu et al. analyzed miRNA expression in sEVs from plasma of 15 PDAC patients and 15 healthy people using qRT-PCR and discovered that both miR-196a and miR-1246 were increased in the serum-derived sEVs of PDAC patients, as compared to the controls.[58] Interestingly, few studies indicated that some miRNAs might decrease in patients, which can also be used as biomarkers for PDAC diagnosis/prognosis. For example, Lai et al. compared the miRNA levels in the sEVs from healthy people and PDAC patients. They discovered that PDAC patients expressed higher levels of some miRNAs such as miR-21, miR-10b, miR-30c, and miR-181a and lower levels of some miRNAs such as miR-let7a and miR-122 in sEVs from serum, which could be used to differentiate healthy controls and PDAC patients.[13] According to ROC curves, miR-10b, -21, -30c, -181a, and -let7a in sEVs all have 100% sensitivity and specificity in detecting PDAC from normal controls, while miR-106b and − 483 failed to distinguish these two groups. Furthermore, another 3 serum-derived sEV RNAs (ANLN, ITGA6, and KRT18) were recently reported lower expression in PDAC than benign pancreatic diseases or healthy controls, while sEV RNA MMP9 showed relatively higher level in advanced PDAC patients than in early stage patients.[80]To further improve the detection accuracy and promote the clinical application of sEVs for PDAC diagnosis, the combination of miRNAs and other molecules such as proteins have been applied for PDAC diagnosis. For instance, a study indicated a combination of proteins (CD44v6, Tspan8, EpCAM, MET, and CD104) and miRNAs (miR-1246, miR-4644, miR-3976, and miR-4306) in serum-derived sEVs could improve the diagnostic accuracy of PDAC with a sensitivity of 1.00 (CI: 0.95–1) and specificity of 0.80 (CI: 0.67–0.90).[66] Results of these studies demonstrated that these miRNAs could serve as diagnostic and prognostic indicators for PDAC.
mRNAs in sEVs have also been reported to be PDAC biomarkers.[81] In a study by Hu et al., glypican-1 (GPC-1) mRNA in sEVs from serum with an AUC of 1.0 or 100% specificity and 100% sensitivity in each stage of PDAC were identified by a biochip, distinguishing patients with early- and late-stage PDAC from healthy donors and patients with benign pancreatic disease.[69] IncRNAs are nonprotein-coding RNAs with more than 200 nucleotides, playing an important role in regulation of gene expression and pathogenesis in cancers.[82] Kenji et al. analyzed lncRNA “highly upregulated in liver cancer (HULC)” expression from serum sEVs of 20 PDAC patients, 22 intraductal papillary mucinous neoplasm (IPMN) patents and 21 heathy individuals, showing significantly higher HULC level in PDAC patients than others.[73] Apart from above RNAs, circRNAs, which was recognized as a novel class of highly stable noncoding RNA species, may also act as a biomarker for PDAC diagnosis. [83] Specifically, Li et al. found an elevated expression level of circRNAs such as circPDE8A and circIARS, in tumor tissues and sEVs from the plasma of PDAC patients. The overexpression of the circRNAs in sEVs was speculated to contribute to tumor invasion and metastasis.[76, 77].
All in all, RNAs in PDAC-derived sEVs showed promising performance in PDAC diagnosis/prognosis. However, there is still some disputes. John et al. found most individual sEVs isolated by ultracentrifugation contained biologically insufficient quantities of miRNAs, accompanied by a small proportion of free miRNAs from plasma, making them unlikely to serve as miRNA-based communication vehicles.[84] Currently, there are no RNA-based detection method for PDAC in clinic due to various reasons, such as heterogeneous nature of sEVs, difficulty in pure RNA extraction from sEVs and lack of validated RNA biomarkers. To realize their potential value as biomarkers for clinical application, more efforts are needed to discover new RNA biomarkers, develop highly sensitive and specific detection techniques and evaluate their significance in PDAC diagnosis/prognosis. Excitedly, two clinical trials (NCT03821909 and NCT04636788) were started to investigate the diagnostic and prognostic values of small RNA biomarkers in sEVs for PDAC diagnosis in August of 2018 in affiliated Nanjing drum tower hospital of Nanjing University medical school and in November of 2020 in Tongji hospital (Tongji medical college), respectively.
DNAs
Owing to their ability to carry information regarding cancer-associated mutations, DNAs in cancer-derived sEVs are of great value as a diagnostic/prognosis tool.[85, 86] Thus, detection of DNA mutations in sEVs from PDAC patients can be potentially used for PDAC diagnosis/prognosis.
Wan et al. developed a device utilizing sEV size-matched silica nanostructures and a surface-conjugated lipid nanoprobe to enrich sEVs from the plasma of 3 PDAC patients and 2 healthy controls. They confirmed that the concentration of DNA with the KRAS mutation was higher in patients than controls. [87] In a study with higher number of PDAC patient cohorts by Allenson et al., sEV KRAS mutations were detected in 66.7% (22/33), 80% (12/15), and 85% (17/20) of localized, locally advanced, and metastatic PDAC, respectively, and in 7.4% (4/54) of healthy controls in the discovery cohort. In the validation cohort of 121 individuals, mutant KRAS DNA in sEVs was detected in 43.6% (17/39) of early-stage PDAC patients and 20% (17/82) of healthy controls. [88] Furthermore, Vincent et al. also reported that KRAS mutant allele fraction (MAF) from sEV DNA provides both predictive and prognostic information for PDAC based on data from 123 serial blood samples. [89]
Multiple DNA mutations in cancer-derived sEVs were also explored for PDAC diagnosis/prognosis. F.A. San et al. identified multiple DNA mutations from plasma-derived sEVs of 2 PDAC patients and found NOTCH1 and BRCA2 in the samples. [90] In another study with bigger cohort, Yang et al. investigated the potential clinical utility of sEV DNA from the serum of 114 healthy subjects, 7 IPMN patients, 9 CP patients, 48 PDAC patients and 12 other patients (diseases such as autoimmune pancreatitis, common bile duct cancer) for identification of both KRASG12D and TP53R273H mutations.[91] They found that sEV DNA harbors KRASG12D mutation in 39.6% of cases, and TP53R273H mutation in 4.2% of cases of PDAC patients, while 2.6% of healthy subjects presented with KRASG12D mutation and none with TP53R273H mutation in the sEVs, indicating the strong potential of circulating sEV DNA for cancer diagnosis/prognosis.
All these studies demonstrate the value of DNA mutations in sEVs as potential biomarkers for PDAC diagnosis/prognosis. Considering that some healthy people also had these DNA mutations in sEVs, this approach may also be used to predict the development of PDAC. However, it should be noted that a DNA mutation does not indicate the presence or prognosis of cancer. In addition, DNA methylation has also been found to play an important role in the initiation and progression of many tumors, such as gastric cancer.[92] The role of DNA methylation in PDAC diagnosis/prognosis will need further investigations.
Proteins
sEVs contain a great number of cytosolic proteins (enzymes, cytokines, apoptotic proteins, oncoproteins, etc.) and surface proteins (adhesion molecules, integrins, tetraspanins, etc.). According to the current version of sEV content database, 9769 proteins have been identified associated with sEVs and 745 of them are relevant to pancreatic cancer.[93] Analysis of these sEV proteins is a powerful tool for PDAC diagnosis/prognosis, due to their unique characteristics compared with traditional serological markers. For example, sEV proteins show higher stability, as they are protected by the lipid bilayer from degradation by extracellular proteases and enzymes.[94] Notably, abundant cancer-associated proteins have been identified in PDAC-derived sEVs, and their types and expression levels are strongly correlated with the presence and progression of PDAC.[95] The aberrantly expressed proteins in PDAC-derived sEVs distinguish them from sEVs of healthy donors or patients with other diseases, and make them a novel means to identify PDAC.[96,97,98] Table2 summarizes the sEV protein biomarkers for PDAC diagnosis/prognosis.
In 2015, Melo et al. reported that the level of GPC-1 on circulating blood sEVs was significantly higher in PDAC patients compared to healthy people, with the sensitivity and specificity of GPC-1 detection both at 100% for PDAC diagnosis.[96] In another study, Buscail et al. modified magnetic beads with CD63 antibody to collect PDAC-derived sEVs from the blood and detected GPC-1 positive sEVs by flow cytometry with sensitivity of 64% and specificity of 90%.[117] Furthermore, Liang et al. found that ephrin type-A receptor 2 (EphA2), which expressed on the surface of sEVs from plasma, could be used to distinguish PDAC patients and healthy subjects.[101] Mutant proteins can also be biomarkers for PDAC diagnosis. In a recent study, Scott et al. found the mutant proteins KRASmut and/or P53mut were positive in plasma-derived sEVs from 15 to 16 stage I PDAC patients using single-sEV analysis technique, showing potential of mutant sEV proteins for early stage cancer diagnosis.[102].
Different proteins played different roles in biologic functions, thus simultaneous detection of multiple proteins on/in sEVs may provide much richer information than each one alone, which may more accurately reflect a molecular signature of the cell from which they originate, comparing to the presence of only one biomarker.[50] For example, Yang et al. used a multiplexed plasmonic assay to analyze circulating cancer-derived sEV biomarkers in the training cohort involving 22 PDAC patients and 10 healthy donors. [95] They found that the five surface membrane proteins (EGFR, EPCAM, MUC1, GPC1, and WNT2) showed relatively different individual expression levels in each subject. The authors also found that GPC-1 had a sensitivity of 55% and a specificity of 60%, whereas the PDAC sEV signature of the five surface proteins showed a sensitivity of 100% and a specificity of 100%. In a more recent study, Juan et al. used bead-based multiplex immunoassay kits to test the isolated sEVs from plasma of stage I-II PDAC patients and healthy donors and found the six protein biomarkers could identify the PDAC with AUC of 0.997 in the training cohort (controls, n = 146; PDAC cases, n = 33) and AUC of 0.978 in the validation cohort (controls, n = 139; PDAC cases, n = 35).[118] The PDAC EV protein signature of sEVs offered higher sensitivity, specificity, and accuracy than the single sEV protein biomarker.
Apart from the membrane proteins on sEVs, proteins inside the sEVs can also serve as biomarkers. Zheng et al. identified the protein complement of sEVs from pancreatic duct fluid of patients with PDAC (n = 13), IPMN (n = 8) and benign pancreatic diseases (n = 5) using mass spectroscopy, and validated the expression by immunohistochemistry. Among all the proteins, the top 35 proteins were significantly associated with PDAC, showing strong potential to be PDAC diagnosis biomarkers.[116] Hiroyuki et al. detected the proteins of sEVs from endoscopic ultrasound-fine needle aspiration (EUS-FNA) biopsy PDAC patients (n = 40) and autoimmune pancreatitis (AIP) patients (n = 6) using nano liquid chromatography tandem-mass spectrometry.[119] 1071 sEV proteins were identified only in PDAC and 153 of them were significantly different between PDAC and AIP, indicating the specific sEV protein barcode of PDAC was promising biomarkers for diagnosis. Mass spectrometry has the potential to greatly promote the proteomics research on cancer-derived sEVs, facilitating the discovery of specific and sensitive protein biomarkers for PDAC diagnosis/prognosis.
Lipids
Compared to the great attention being paid to nucleic acids and proteins in sEVs, lipids represent other less-explored bioactive molecules abundantly present in sEVs. According to a report of 2019, research into sEV lipidomics accounted for less than 4.3% of the sEV genomics studies, and approximately 5.5% of the sEV proteomics work, indicating low scientific interest in sEV lipid research.[120] However, lipid is one of the most important components in sEVs, playing indispensable roles on the structural and regulatory functions of sEV biogenesis, release, targeting and cellular uptake.[120, 121] Therefore, lipidomic studies of sEVs may be an innovative direction for the discovery of new biomarkers for cancer diagnosis/prognosis.
Currently, lipids of sEVs have been reported to be biomarkers for cancer diagnosis/prognosis in diverse cancers, including prostate cancer,[122] non-small cell lung cancer,[123] and colorectal cancer.[124] So far, very limited studies on lipids in sEVs for PDAC diagnosis/prognosis have been published. Raghava et al. found phosphatidylserine positive sEVs in blood increased significantly in PDAC bearing mouse, suggesting the potential of phosphatidylserine positive sEVs for PDAC detection.[125] Furthermore, Samuel et al. extracted lipid from different PDAC cell-derived sEVs and analyzed them by mass spectroscopy.[126] They found that sEVs derived from AsPC-1, Panc-1, BxPC-3 and HDPE cells all had significantly different lipid expression profiles, including phosphatidylcholine, phosphatidylserine, phosphatidylethanolamine, phosphatidic acid and phosphatidylinositol, which showed potential use for PDAC diagnosis. Charles et al. analyzed 1021 lipid species from sEVs of different pancreatic cancer cell lines (Panc-1, Capan-1, SW-1990, Mia PaCa-2, PPCL-68 and PPCL-46) and normal cell lines (hTERT-HPNE, HPDE-H6c7) dysregulated lipids were observed between cancer-dervied sEVs and normal sEVs, especially these lipid species containing palmitic acid (16:0) and sphingomyelin.[127] In another study by Tao et al., liquid chromatography-data dependent acquisition-mass spectroscopy (LC-DDA-MS) based lipidomic analysis was used to analyze the lipid expression profile in sEVs derived from peripheral blood of 22 PDAC patients and 17 healthy people.[128] The authors found that about 270 lipids were significantly dysregulated between the sEVs of PDAC patients and healthy controls, and 61 significantly dysregulated lipids were further analyzed by LC-MRM-MS to verify the results of lipidomic analysis in sEVs from 24 PDAC patients and 40 healthy donors. They discovered that LysoPC 22:0, PC (P-14:0/22:2) and PE (16:0/18:1) were all associated with tumor stage, CA19-9, CA242 and tumor diameter. These findings revealed that dysregulated lipids in sEVs from PDAC patients show potential as biomarkers for diagnosis/prognosis. The research into the role of lipids in sEVs offers a un-explored avenue for PDAC biomarker discovery.
Glycans
Glycans consist of oligosaccharides linked glycosidically to proteins, lipids and proteoglycans which are displayed on the exterior surfaces of cells and sEVs. They can be classified into different types of glycans. Specifically, they can be divided into N-linked glycans (attached to the Asn of glycoproteins in a particular Asp-X-Ser/Thr sequon), O-linked glycans (attached to the Ser or Thr of glycoproteins and predominate on mucins), glycosaminoglycans (attached to Ser residues of proteoglycans), and glycolipids (attached variously to lipids).[129] Different types of glycans and their receptor binding lead to different physical properties and cellular functions which attribute to the development and progression of cancer and other diseases.[130, 131] Moreover, glycans play significant roles in intracellular interactions that depend on cellular conditions and the onset of diseases such as PDAC. [132] Therefore, glycans on PDAC-derived sEVs may also have the potential to be a useful biomarker for PDAC diagnosis/prognosis.
To study the roles of glycan changes in pancreatic diseases, Engle et al. inducibly expressed human fucosyltransferase 3 and β-1,3-galactosyltransferase 5 in mice to reconstitute the glycan sialyl-Lewisa (also known as CA19-9) which was a carbohydrate antigen attached to O-glycans on the surface of pancreatic cells. 133They found that CA19-9 expression in mice resulted in rapid and severe pancreatitis with hyperactivation of epidermal growth factor receptor (EGFR) signaling to promote PDAC. Yokose and his colleagues analyzed the differential glycomic profiling of sEVs derived from serum (two cohorts including 117 PDAC patients and 98 normal controls) using lectin microarrays.[134] The glyco-candidates of PDAC-specific sEVs were quantified using a highly-sensitive sEV-counting system, ExoCounter. Quantitative analysis using ExoCounter revealed that the O-glycan-binding lectins, Agaricus bisporus agglutinin (ABA) and Amaranthus caudatus agglutinin (ACA), positive sEVs were significantly increased in the culture of PDAC cell lines and in the serum of PDAC patients. These specific sEVs with O-glycans recognized by ABA/ACA were elevated in PDAC sera and could act as potential biomarkers in a liquid biopsy for PDAC patient screening. Choi et al. attached lectins with a high and specific affinity for sialic acid or fucose to bifunctional Janus nanoparticles (JNPs) for glycan detection on sEVs by a microfluidic device.[135] sEVs derived from PDAC cell lines and plasma samples of PDAC patients were successfully captured on the lectin-conjugated JNPs. The relatively higher glycan recognition of sEVs from patient plasma by Sambucus nigra agglutinin (SNA) and Aleuria aurantia lectin (AAL) modified JNPs could thus potentially be used for cancer diagnosis. The abnormal expression of specific glycans on sEVs, their presence in patient serum and plasma, and their possible ability to facilitate metastases, suggests that glycans could contribute to cancer diagnosis/prognosis. To fully exploit glycans for further clinical use, it will be vital to fully profile the glycans of PDAC-derived sEVs and determine how this varies between different cancer stages and healthy people.
Biomolecules such as nucleic acids, proteins, lipids and glycans in PDAC-derived sEVs have all shown the potential to act as biomarkers for PDAC diagnosis/prognosis. RNAs and proteins serve as highly specific biomarker candidates for PDAC detection and represent the most important biomolecules associated with PDAC-derived sEV studies. RNAs affect the cancer associated metabolism and functions. They could be easily extracted and analyzed using the well-developed RNA profiling techniques such as polymerase chain reaction and next-generation sequencing. There are three ongoing clinical trials using RNAs for PDAC detection as mentioned above. sEV proteins include membrane proteins and inner proteins, analysis of which could help us to understand the mechanisms of sEV biogenesis and functions. Membrane proteins are more studied than inner proteins for PDAC detection due to easier sample preparation and rapid and sensitive analysis approaches (e.g. flow cytometry and enzyme-linked immunosorbent assay), which are based on high specific of antibodies. DNA mutations exist in sEVs from both healthy donors and PDAC patients, their role for clinic use need more investigation. sEV glycans and lipids have been explored for cancer diagnosis/prognosis due to the fast development of glycomics and lipidomics in recent years, screening and validation of clinical biomarkers for PDAC diagnosis/prognosis require more efforts. There is no sEV based clinical assays for PDAC detection now. To realize clinical use of these biomolecules, more intensive studies are still required to determine their clinical sensitivity, specificity and accuracy. Moreover, the lack of clinically applicable, reliable detection techniques for these biomolecules makes them difficult to be utilized. These potential problems may be solved by gaining more knowledge about how these biomolecules transport into sEVs, optimizing the extraction procedures and improving the detection techniques of these active molecules, which could eventually lead to the development of a heterogeneous panel of biomarkers for clinical application in PDAC diagnosis/prognosis. Although cancer-derived sEVs as biomarkers for cancer diagnosis/prognosis in patients have been reported in a flurry of papers, very few clinical diagnostic/prognostic assays are implemented. To date, two sEV-based clinical assays have reached the stage of clinical validation and one assay called ExoDx Prostate (IntelliScore) has been launched onto the market in United States since 2016.[136, 137] Briefly, the ClarityDx™ System (NCT03957252) is used for prostate cancer diagnosis by detecting GHSR (ghrelin receptor), PSMA (prostate-specific membrane antigen) and polysialic acid in blood-derived sEVs by micro flow cytometry. Sentinel™ PCC4 test (NCT04100811, NCT04661176) is used for prostate cancer by detecting 442 small non-coding RNAs (sncRNAs) in urinary sEVs. Another test Sentinel™ (NCT04155359) detecting 280 sncRNAs is used for bladder cancer diagnosis. The commercial available assay ExoDx Prostate (IntelliScore) (NCT03235687, NCT03031418, NCT04720599, NCT02702856) is based on the detection of three RNAs (PCA3, SPDEF, ERG) in urinary sEVs from prostate cancer patients by quantitative reverse transcription-polymerase chain reaction (RT-PCR).
Cancer therapy
The exploration of cancer-derived sEVs for therapeutic purposes is still in its infancy due to the risk of endogenous cargo molecules which may activate pathological pathways.[138] Over the past few years, several cancer-derived sEV based therapeutic approaches have been developed, including inhibition of cancer-derived sEV formation or secretion, using cancer-derived sEVs as a potential drug carrier for target therapy and for immunotherapy. As compared to other therapeutic strategies, cancer-derived sEV based therapies are likely to have targetable ability, high stability, cross biological barrier ability and low toxic side effects.[139] All of these characteristics support the potential application of cancer-derived sEVs in cancer treatment. In this section, we will mainly review the advances in therapeutic strategies using cancer-derived sEVs for PDAC treatment (Table3).
Inhibition of cancer-derived sEV formation and secretion
Cancer-derived sEVs play key roles in PDAC progression and in the onset of cancer drug resistance.[35] It has also been reported that cancer-derived sEVs can transport epidermal growth factors to macrophages, thus interfering with the innate immune system’s function.[157] These cancer-derived sEVs not only promoted the cancer development, but also affected the immune system and drug resistance in order to protect the cancer cells. Thus, inhibiting the formation and release of cancer-derived sEVs may delay cancer progression and improve cancer treatment. A growing number of studies have indicated that disrupting the signaling pathway of cancer-derived sEVs was able to block the formation and secretion of cancer-derived sEVs, thus inhibiting tumor growth and metastasis in cancers.[158,159,160,161] As aforementioned, the biogenesis of sEVs occurs inside MVBs, and is driven mainly by two mechanisms: endosomal sorting complexes required for transport (ESCRT)-dependent and ESCRT-independent pathways (Fig.3). Manumycin A is an antibiotic that can inhibit RAS (small GTPases) activation, thus disrupting the biogenesis of sEVs by the ESCRT-dependent pathway. GW4869 is able to suppress neutral sphingomyelinase (nSMase), which is an important enzyme for MVB biogenesis, resulting in the inhibition of sEV formation by the ESCRT-independent pathway.[162] Furthermore, the transportation of MVBs, the fusion between MVBs and cell membrane, and the release of sEVs from MVBs, are regulated by many agents, such as the RAB family (a member of the RAS superfamily of small G proteins) for recruiting cytosolic tethers to MVB membranes, soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) for MVB and cell membrane fusion, Ca2+ for calpain activation, and Rho-associated protein kinases (ROCK) for cytoskeleton re‐organization (Fig.3).[162,163,164].
Cancer-associated fibroblasts (CAFs) are the dominant components of PDAC tumor bulk. Richard et al. found that CAFs played a critical role in promoting the proliferation of PDAC cells through sEV signaling.[140] While CAFs were intrinsically resistant to gemcitabine (GEM), GEM-treated CAFs significantly increased the release of sEVs, which resulted in elevation of a chemoresistance-inducing factor in recipient epithelial cells and promoted proliferation and drug resistance. The treated GEM exposed CAFs were then treated with sEV release inhibitor GW4869 and the survival of epithelial cells was significantly reduced, indicating an important role of cancer-derived sEVs in chemotherapeutic drug resistance. Weikun et al. built a bio-mimetic 3D co-culture model to integrate the complex tumor organoids and used sEV release inhibitor climbazole and imipramine to treat the PDAC organoids, finding that climbazole and imipramine inhibited the growth of organoids and chemoresistance by preventing the release of CAF tumor organoid-derived sEVs. [141] In another study, Mikamori et al. found GEM resistance in PDAC cells might be relevant to miR-155, which controlled genes required for sEV synthesis.[142] The authors used RAB27B siRNA (siRAB27B) to transfect PDAC cells and downregulated RAB27B effectively. The resultant RAB27B knockdown resulted in the remarkable reduction of the amounts of sEVs and GEM resistance in PDAC cells with or without transfection with pre miR-155. These results indicated that targeted miR-155 therapy and inhibition of cancer-derived sEV secretion may be an effective way to reduce or eliminate GEM resistance and to help improve the therapy of PDAC. PDAC patients normally have altered tumor suppressor phosphatase and tensin homolog (PTEN) and low expression of PTEN may promote PDAC progression.[165, 166] Katherine et al. discovered GEM treated CAFs could release PTEN targeting miRNAs (miR-21, miR-181a, miR-221, miR-222, and miR-92a) and the suppression of CAF-derived sEV secretion with inhibitor GW4869 could reduce these PTEN targeting miRNAs and restore the PTEN expression.[143] However, these strategies involve the inhibition of sEVs being secreted from both healthy cells and cancer cells. Therefore, their use in cancer treatment should be carefully considered and investigated, as sEVs also play an essential role in regulating normal biological functions as well.

Strategies of using cancer-derived sEVs for drug delivery. Chemotherapy drugs, nucleic acids and/or proteins can be loaded into sEVs by direct or indirect methods
It would be safer to only inhibit or remove the cancer cells-derived sEVs. For example, Hoshino et al. found that ITGβ4 or ITGβ5 genes were overexpressed in pancreatic cancer cell-derived sEVs; knocking down or inactivating these genes dramatically reduced the metastatic ability of pancreatic cancer cells.[144] A more recent study by Capello et al. indicated that PDAC-derived sEVs displayed a wide variety of tumor-associated antigens which could bind circulating autoantibodies.[167] PDAC-derived sEVs exerted a decoy function and possibly attenuated complement-dependent cytotoxicity against PDAC cells, which suggested that it may be possible to inhibit PDAC sEV secretion or selectively eliminate the circulating PDAC-derived sEVs through affinity capture as one way to treat PDAC. One interesting finding is that the inhibition of a specific molecule on cancer-derived sEVs could also improve cancer treatment. Chun-an et al. found vascular endothelial growth factor-C (VEGF-C) on PDAC-derived sEVs promoted lymphangiogenesis and thus blocking of the production of VEFG-C could inhibit PDAC early dissemination and cancer malignancy.[145].
PDAC-derived sEVs have been reported to promote immune suppression, cancer metastasis and chemoresistance in PDAC. The suppression of PDAC sEV secretion or disruption of the signaling pathways is thus an achievable strategy to treat PDAC. To bring these therapeutic approaches to clinical application, it is essential to comprehensively understand the biogenesis of PDAC-derived sEVs as well as their signaling pathways and elucidation of the mechanisms as to how PDAC-derived sEVs contribute to cancer metastasis and chemoresistance.
Cancer-derived sEVs as drug carrier vesicles
In recent years, researchers have made great progress in the development of sEVs as drug carriers. Compared with liposomes and other nanoparticles, sEVs possess better biocompatibility and are considered as a natural way for drug delivery. Injected sEVs shed from endogenous cells of the body are tolerated with minimal immune reaction and toxicity.[139, 168] The therapeutic cargos can be efficiently delivered into the tumor microenvironment using sEVs since these vesicles display efficient target-homing capabilities and easily penetrate through natural biological barriers due to their small particle size.[139, 168] In addition, sEVs express the integrin-associated transmembrane protein CD47 that protects sEVs from phagocytosis by monocytes and macrophages, thus increasing the sEVs’ half-life.[169] Because of these unique advantages, sEVs have emerged as ideal candidates for the delivery of therapeutic cargos, such as chemotherapy drugs, nucleic acids, and proteins.[170, 171] Therapeutic agents can be loaded into sEVs by direct methods such as incubation, sonication and electroporation, or by indirect methods using a drug to treat parent cells and collecting the released sEVs containing the drugs.[172] A series of clinical trials in phase I and II have demonstrated that sEVs hold strong potential for drug delivery.[173] sEVs from different sources such as mesenchymal stem cells, macrophages, dendritic cells, embryonic kidney cells, cancer cells have been used for drug delivery.[174] For example, Kamerkar et al. loaded short interfering RNA or short hairpin RNA in sEVs from fibroblast-like mesenchymal cells to target oncogenic KRASG12D in Panc-1 tumor bearing mouse models and moved forward to a Phase-I clinical trial by the M.D. Anderson Cancer Center in January of 2021 (NCT03608631) [169, 175].
Cancer-derived sEVs accumulate preferentially in the tumor tissue via the so-called passive targeting ability, which may be ascribed to the presence of several classes of proteins (e.g. integrins, tetraspanins and cancer-specific antigens) on their surface.[47, 176] For example, Xu et al. investigated the uptake properties of sEVs derived from Panc-1 and other cell lines (B16-F10, HEK-293) and Panc-1-derived sEVs showed significant higher uptake in Panc-1 cells than other types of sEVs in vitro and in vivo (Panc-1 tumor bearing mouse model), demonstrating the passive homing ability of Panc-1 sEVs.[177]Additionally, modification the surface of cancer-derived sEVs with targeting molecules such as ligands, aptamers and antibodies can improve the targeting ability (active targeting ability).[176] Furthermore, some researchers proposed that fusing sEVs with conventional synthetic nanoparticles such as liposomes to form hybrid particles (combination of two unique entities) could achieve the beneficial properties associated with both sEVs and synthetic nanoparticles. Fig.4 illustrates the strategies of using cancer-derived sEVs for drug delivery.
Li et al. loaded GEM into autologous Panc-1 cell-derived sEVs for targeted chemotherapy of PDAC.[146] GEM-loaded sEVs facilitated the cellular drug uptake and significantly suppressed tumor growth in tumor bearing mice; even tumors in several mice disappeared without recurrence after treatment. PDAC treatment response however may be affected by cancer associated macrophages. Cristina et al. demonstrated these cancer associated macrophage-derived sEVs decreased the PDAC cell sensitivity to GEM, which was caused by chitinase 3-like-1 (CHI3L1) and fibronectin (FN1).[178] In order to overcome GEM resistance in PDAC, Aspe JR et al. delivered survivin T34A by melanoma cell-derived sEVs to restore GEM sensitivity to PDAC cell lines, inducing a significant increase in apoptotic cancer cell deaths, compared with using GEM alone.[147] More recently, Hasan et al. engineered the surface of PDAC (Panc-1)-derived sEVs by conjugating with functional ligand RGD (peptide composed of several repetitions of Arg-Gly-Asp) and magnetic nanoparticles, then loaded PTX into the modified sEVs to treat xenograft mice bearing Panc-1 tumor.[148] RGD showed good affinity for αvβ3 that was highly expressed in pancreatic cancer cells, thus RGD modification on sEVs could improve the tumor cell targeting ability. The authors also found one important molecule integrin β3, which was expressed in Panc-1 cells and Panc-1 sEVs, involved in the home-driving properties of Panc-1 sEVs to their parent cancer cells. This therapeutic formulation based on the passive and active targeting ability of cancer-derived sEVs could efficiently deliver the drug to the cancer cells and significantly reduced the tumor size when comparing with the control groups. In addition to chemical drugs, RNA drugs have also been shown to be highly effective for PDAC treatment. Xu et al. encapsulated siRNA (siPAK4) into PDAC-derived sEVs by electroporation for PDAC treatment in a mouse model. The siPAK4 induced significant apoptosis of tissue and prolonged survival of PDAC bearing mice with minimal toxicity.[149] Another study by Song et al. found selective packaging of miRNAs into EVs could lead to enrichment of stromal specific miR-145 in sEVs from tumor-associated stroma (TAS). The TAS-derived miR-145 was able to be delivered into PDAC cells by the sEVs and suppressed the cancer development.[150] sEVs have also been engineered to enhance the loading ability of therapeutic cargo or to improve the targeting ability for treatment of other cancers [179], [180].
Collectively, these results indicated an interesting potential utility of cancer-derived sEVs as candidates for different therapeutic agent delivery that could be used to develop innovative treatment strategies for PDAC. However, it should be noted that the endogenous cargos such as oncogenes in cancer-derived sEVs may be associated with cancer progression and migration,[174] thus their potential safety concerns still remain. Several clinical trials have been conducted using cancer-derived sEVs and no serious safety issues were reported in these studies [181,182,183]. For example, Guo et al. used autologous tumor cell-derived nanoparticles loaded with a chemotherapeutic drug (methotrexate) to treat lung cancer patients with pleural effusions (4 females and 8 males).[182] Six mild adverse events (grades 1 to 2) were observed through the treatments and no acute autoimmune reactions were recorded. In another clinic trial (NCT01854866), Huang et al. investigated the safety and effectiveness of cancer-derived sEVs loaded with chemotherapeutic drugs for malignant ascites and pleural effusion treatment and reported no typical side effects.[184] Furthermore, the development of new techniques that remove the harmful contents in cancer-derived sEVs or preparation of sEV mimetics might helpful to improve the safety. The drug loading capacity of cancer-derived sEVs is also one of the obstacles. Thus a few studies reported the fused sEVs with other nanoparticles such as liposomes or synthesis of sEV mimetics to enhance the loading capacity.[45] Moreover, the improvement of the drug loading method such as electroporation, saponin-assisted loading and extrusion could further increase the loading capacity of sEVs.[185] Another major problem for drug loading using cancer-derived sEVs is their low yield. The effective sEV dose is 10–500µg sEV proteins to each mouse or 0.5–1.4 × 1011 sEVs to each patient in one clinic trial.[186,187,188] Thus, the strategies to obtain large-scale clinical-grade sEVs are required for in vitro basic research, in vivo preclinical animal models, and clinical trials.
Using cancer-derived sEVs in immunotherapy
Antitumor immunity is triggered when immune effector cells such as natural killer (NK) cells are activated. sEVs derived from immune cells, such as NK cells, dendritic cells (DCs) and bone marrow mesenchymal stem cells (BM-MSC), can inhibit PDAC progression.[189, 190]. Similarly, sEVs derived from cancer cells carrying immunosuppressive molecules such as Fas ligand (FasL), TNF-related apoptosis-inducing ligand (TRAIL), programmed death-ligand 1 (PD-L1), and enzymes engaged in the adenosine pathway (CD39 and D73) were found to mediate the functions of immune cells, such as activation of regulatory T-cells (T regs), DCs, macrophages and immature myeloid-derived suppressor cells (MDSCs) (Fig.5), thus playing important roles in the establishment of an immunosuppressive microenvironment.[191,192,193]. For example, these cancer-derived sEVs caused the apoptosis of CD8 + T cells, which is critical for immune defense against cancer, by activating death receptor pathways.[194] Although cancer-derived sEVs have thus been shown to have an immunosuppressive effect on various immune cells, some studies also indicate these sEVs may contain immunostimulatory factors (hear shock protein 70/90 (HSP70/90), major histocompatibility complex class I/II (MHC I/II), tumor-associated antigens (TAAs), etc.) and serve as anti-tumor agonists or vaccines to increase cancer antigen recognition and promote tumor clearance.[152, 195196197] Therefore, cancer-derived sEVs are considered to be valuable agents for the treatment of cancer. Although current progress of using cancer-derived sEVs for PDAC immunotherapy is very limited, it is an emerging field for study. Fig. 5 illustrates the effect of cancer-derived sEVs on immune cells and possible therapeutic strategies for cancer treatment by immunotherapy.

Effects of cancer-derived sEVs on immune cells and therapeutic strategies for cancer treatment by immunotherapy. Cancer-derived sEVs contain immunosuppressive and immunostimulatory molecules, which can be used to activate immune cells. Loading immune drugs into sEVs or inhibition of cancer-derived sEV secretion are two other strategies for immunotherapy
Que et al. investigated the proteins of PDAC-derived sEVs and their effect on immune cells DCs/ cytokine-induced killer cells (CIKs) for anti-tumor activity.[151] They isolated sEVs from Panc-1 cell supernatants, then ruptured and ultrafiltered the sEV lysate. 128 proteins, including several immune-activating proteins (attractin, complement C3, C4 and C5, integrin and lactotransferrin), remained in the lysate, while the miRNAs from sEVs were removed. The authors found that these PDAC-derived sEV lysates increased the tumor-killing capacity of DCs/CIKs towards PDAC cells, suggesting that miRNA-depleted cancer-derived sEVs might represent valuable immunotherapeutic tools in PDAC treatment. In line with this study, Gastpar et al. found one protein, HSP70, enriched from PDAC-derived sEVs, stimulated NK cell migration and caused cytotoxicity against cancer cells.[152] Another study discovered that PDAC-derived sEVs, from rats, supported leukocyte effector functions by strengthening NK and cytotoxic T cell activity and showed a minor effect on leukocyte activation. Thus, these cancer-derived sEVs might be used as adjuvant in immunotherapy.[153].
Cancer-derived sEVs can also be used as vectors of immune drugs for cancer immunotherapy. Mahmoodzadeh et al. used PDAC-derived sEVs as carriers to load staphylococcal enterotoxin B (SEB), which was used as a potent immune stimulator for T cell activation.[154] They demonstrated that SEB-loaded sEVs could significantly induce cancer cell apoptosis and no cytotoxic effect was observed in both human kidney embryonic cells and peripheral human white blood cells, indicating that such a strategy might be used to stimulate an immune response against PDAC cells. The authors also investigated the expression of anti-apoptotic genes including Bax, Bak and Fas in cells and found the induction of these genes after treatment with SEB-loaded sEVs. In a more recent study, Wenxi et al. modified sEVs derived from immunogenically dying tumor cells with MART-1 peptide (sequence: ELAGIGILTV), which displayed immunogenic properties and was able to expand CD8 + T cells for adoptive T cell transfer.[155] The modified sEVs could enhance antitumor immune response. The authors also loaded CCL22 siRNA into the modified sEVs to suppress Treg expansion, the results demonstrated them as an effective prophylactic vaccine in delaying tumor growth and good adjuvant for chemotherapeutic drugs in PDAC treatment. As cancer-derived sEVs can suppress the functions of some immune cells, the inhibition of these cancer-derived sEV secretion may also improve cancer treatment by immunotherapy. Lau et al. reported that PDAC-derived sEVs from tumor-bearing mice affected the expression of genes in the salivary glands and subsequently induced changed contents of salivary sEVs in vivo.[156] The salivary sEVs from PDAC tumor-bearing mice regulated the phenotype of NK cells, resulting in inhibition of the antitumor cytotoxicity of NK cells. When these saliva sEVs were inhibited, they lost the ability to inhibit NK cells.
Overall, studies using cancer-derived sEVs as potential immunity enhancers against PDAC have so far provided some preliminary but promising results, which encourages further investigations in this field. There has however been some controversy regarding the biological roles of cancer-derived sEVs in immunotherapy, as these sEVs mediate both immunosuppressive or immunostimulatory responses and it is challenging to reconcile these two aspects. The safety concerns of cancer-derived sEVs is another problem and substantial research is required to evaluate this issue.
Conclusion and perspectives
In summary, PDAC is a highly aggressive and lethal malignancy mostly due to its late-stage presentation and lack of curative therapies. This malignancy is difficult to diagnose/prognose, monitor and treat. Hence, the development of novel diagnostic/prognostic biomarkers and better therapeutic strategies are urgently needed. Cancer-derived sEVs are abundant in both the tumor environment and circulation system. They act as a non-invasive biomarker and provide diverse biological functional information and medical applications in PDAC diagnosis/prognosis and treatment. The biomolecules in/on sEVs including RNAs, mutated DNAs, proteins, lipids and glycans provide the feasibility for developing non-invasive liquid biopsy for low-risk and routine screening for PDAC diagnosis/prognosis. However, it seems unrealistic to detect a single biomarker for accurate PDAC diagnosis/prognosis as a highly sensitive and specific test. Multiplex biomarker detection of sEVs with simple, sensitive and specific assay is a promising area for further development. Researchers should continue to investigate the use of cancer-derived sEVs as biomarkers to detect PDAC. In addition, these cancer-derived sEVs play vital roles in cancer environment, including cancer progression and metastasis, drug resistance, immune regulation, etc., and hold unique properties for cancer treatment such as intrinsic homing ability to tumor tissues, which make them an advantageous mechanism for cancer therapy. Thus new therapeutic approaches based on the understanding of biological functions of these cancer-derived sEVs may facilitate the development of new PDAC treatments. The targeting ability, safety and drug loading efficiency of cancer-derived sEVs should be further explored to enable effective cancer treatment.
To further implement the clinical application of cancer-derived sEVs for PDAC diagnosis/prognosis, there are several issues to be considered: (1) lack of reliable techniques for sEV separation in high-purity homogeneous form; (2) isolation of cancer-derived sEVs from bodily fluids containing healthy sEVs; (3) screening of multiple and reliable biomarkers; (4) development of highly sensitive, specific sEV detection methods. Given that the lipoproteins and other type of EVs have similar particle size with sEVs, it is impossible to enrich high-purity homogeneous sEVs using single traditional isolation method (e.g. ultracentrifugation and size exclusion chromatography). The combination of multiple isolation methods for specific sEV subtype enrichment might be one of the solutions. For example, particles with similar size of sEVs can be enriched using size exclusion chromatography then purified by immunocapture of the specific subtype of sEVs using antibodies. However, the lack of high discriminating cancer biomarker slow down the progress. With the development of multi-omics tools, multiple and reliable biomarker can be screened and used for cancer detection in the near future. In addition, new techniques are still under exploration to improve the sensitivity and specificity for the detection of sEVs for cancer diagnosis/prognosis. The development of new analysis methods such as machine learning are expected to advance the cancer diagnosis/prognosis as well. To progress the development of cancer-derived sEVs in cancer treatment towards clinic application, there are several tasks researchers should pay more attention: (1) large-scale production and isolation approaches for sEVs; (2) technical difficulties in production of clinical-grade sEVs and establishment of corresponding quality control standards; (3) drug loading capacity; (4) cancer cell targeting efficiency; and (5) safety in human trials. Cancer-derived sEVs based therapeutic strategies are still in the primary research stage, there still a long way to go before they can be applied in clinic. For application of cancer-derived sEVs in therapy, it is essential to develop strict good manufacturing practice (GMP) procedures that include all the parameters such as the type of parent cells, the sample volume and isolation conditions (temperature, speed, time, etc.). Corresponding quality control standards should also be developed to release the qualified sEV products and maintain reproducible manufacturing process, including the acceptable range of particle size, concentration and biomarker profiles. Hybrid particles (e.g. sEV and liposome hybrid) or sEV mimetics which carry various payloads may be an interesting area to explore to improve the drug loading capacity. The development of different drug loading techniques such as electroporation could also increase the loading efficiency. As for the cancer cell targeting efficiency and safety of cancer-derived sEVs in human trials, there are quite limited studies being reported. Further collaborative efforts in multiple disciplines are needed to transform cancer-derived sEVs based therapeutic strategies into practical applications for the benefit of cancer patients.
Data availability
Not applicable.
References
Court CM, Ankeny JS, Hou S, Tseng HR, Tomlinson JS. Improving pancreatic cancer diagnosis using circulating tumor cells: prospects for staging and single-cell analysis. Expert Rev Mol Diagn. 2015;15:1491–504.
Neoptolemos JP, Kleeff J, Michl P, Costello E, Greenhalf W, Palmer DH. Therapeutic developments in pancreatic cancer: current and future perspectives. Nat reviews Gastroenterol Hepatol. 2018;15:333–48.
Buscail L. Commentary. Pancreatic cancer: is the worst to come? Int J Epidemiol. 2017;46:1774–5.
Siegel RL, Miller KD, Jemal A. Cancer statistics. 2020. CA: a cancer journal for clinicians. 2020; 70: 7–30.
Kleeff J, Korc M, Apte M, La Vecchia C, Johnson CD, Biankin AV, et al. Pancreatic cancer. Nat Reviews Disease Primers. 2016;2:16022.
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Cancer J Clin. 2021;71:209–49.
Humphris JL, Chang DK, Johns AL, Scarlett CJ, Pajic M, Jones MD, et al. The prognostic and predictive value of serum CA19.9 in pancreatic cancer. Ann Oncol. 2012;23:1713–22.
Zhou B, Xu J-W, Cheng Y-G, Gao J-Y, Hu S-Y, Wang L, et al. Early detection of pancreatic cancer: Where are we now and where are we going? J Cancer. 2017;141:231–41.
Garrido-Laguna I, Hidalgo M. Pancreatic cancer: from state-of-the-art treatments to promising novel therapies. Nat reviews Clin Oncol. 2015;12:319–34.
Cancer statistics of American Cancer Society. https://www.cancer.org/cancer/pancreatic-cancer/about/key-statistics.html. Accessed 20 August 2022.
Rahib L, Smith BD, Aizenberg R, Rosenzweig AB, Fleshman JM, Matrisian LM. Projecting cancer incidence and deaths to 2030: the unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014;74:2913–21.
Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer Statistics. 2021. CA: a cancer journal for clinicians. 2021; 71: 7–33.
Lai X, Wang M, McElyea SD, Sherman S, House M, Korc M. A microRNA signature in circulating exosomes is superior to exosomal glypican-1 levels for diagnosing pancreatic cancer. Cancer Lett. 2017;393:86–93.
Zhu Y, Zhang H, Chen N, Hao J, Ma XJM. Diagnostic value of various liquid biopsy methods for pancreatic cancer: A systematic review and meta-analysis. Med (Baltim). 2020;99:e18581.
Poruk KE, Gay DZ, Brown K, Mulvihill JD, Boucher KM, Scaife CL, et al. The clinical utility of CA 19 – 9 in pancreatic adenocarcinoma: diagnostic and prognostic updates. Curr Mol Med. 2013;13:340–51.
Ballehaninna UK, Chamberlain RS. The clinical utility of serum CA 19 – 9 in the diagnosis, prognosis and management of pancreatic adenocarcinoma: An evidence based appraisal. J Gastrointest Oncol. 2012;3:105–19.
Mellby LD, Nyberg AP, Johansen JS, Wingren C, Nordestgaard BG, Bojesen SE, et al. Serum Biomarker Signature-Based Liquid Biopsy for Diagnosis of Early-Stage Pancreatic Cancer. J Clin Oncol. 2018;36:2887–94.
Mayerle J, Kalthoff H, Reszka R, Kamlage B, Peter E, Schniewind B, et al. Metabolic biomarker signature to differentiate pancreatic ductal adenocarcinoma from chronic pancreatitis. Gut. 2018;67:128–37.
Mehta S, Bhimani N, Gill AJ, Samra JS, Sahni S, Mittal A. Serum Biomarker Panel for Diagnosis and Prognosis of Pancreatic Ductal Adenocarcinomas. Front Oncol. 2021;11:708963.
Capurso G, Signoretti M, Valente R, Arnelo U, Lohr M, Poley JW, et al. Methods and outcomes of screening for pancreatic adenocarcinoma in high-risk individuals. World J Gastrointest endoscopy. 2015;7:833–42.
Baek HW, Park MJ, Rhee YY, Lee KB, Kim MA, Park IA. Diagnostic accuracy of endoscopic ultrasound-guided fine needle aspiration cytology of pancreatic lesions. J Pathol translational Med. 2015;49:52–60.
Kitano M, Yoshida T, Itonaga M, Tamura T, Hatamaru K, Yamashita Y. Impact of endoscopic ultrasonography on diagnosis of pancreatic cancer. J Gastroenterol. 2019;54:19–32.
Adamska A, Domenichini A, Falasca M. Pancreatic Ductal Adenocarcinoma: Current and Evolving Therapies. Int J Mol Sci. 2017;18:1338.
Neesse A, Krug S, Gress TM, Tuveson DA, Michl P. Emerging concepts in pancreatic cancer medicine: targeting the tumor stroma. Onco Targets Ther. 2014;7:33–43.
Xu Z, Pothula SP, Wilson JS, Apte MV. Pancreatic cancer and its stroma: a conspiracy theory. World J gastroenterology: WJG. 2014;20:11216.
Gu Z, Du Y, Zhao X, Wang C. Tumor microenvironment and metabolic remodeling in gemcitabine-based chemoresistance of pancreatic cancer. Cancer Lett. 2021;521:98–108.
Zeng S, Pöttler M, Lan B, Grützmann R, Pilarsky C, Yang H. Chemoresistance in Pancreatic Cancer. Int J Mol Sci. 2019;20:4504.
Théry C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses. Nat Rev Immunol. 2009;9:581–93.
Kalra H, Drummen GPC, Mathivanan S. Focus on Extracellular Vesicles: Introducing the Next Small Big Thing. Int J Mol Sci. 2016;17:170.
Gurunathan S, Kang M-H, Jeyaraj M, Qasim M, Kim J-H. Review of the isolation, characterization, biological function, and multifarious therapeutic approaches of exosomes. Cells. 2019;8:307.
Shao H, Im H, Castro CM, Breakefield X, Weissleder R, Lee H. New Technologies for Analysis of Extracellular Vesicles. Chem Rev. 2018;118:1917–50.
Chang C-H, Pauklin S. Extracellular vesicles in pancreatic cancer progression and therapies. Cell Death Dis. 2021;12:1–12.
Tkach M, Théry C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell. 2016;164:1226–32.
Kourembanas S. Exosomes. Vehicles of Intercellular Signaling, Biomarkers, and Vectors of Cell Therapy. Annu Rev Physiol. 2015;77:13–27.
Armstrong EA, Beal EW, Chakedis J, Paredes AZ, Moris D, Pawlik TM, et al. Exosomes in Pancreatic Cancer: from Early Detection to Treatment. J Gastrointest surgery: official J Soc Surg Aliment Tract. 2018;22:737–50.
De Lellis L, Florio R, Di Bella MC, Brocco D, Guidotti F, Tinari N, et al. Exosomes as pleiotropic players in pancreatic cancer. Biomedicines. 2021;9:275.
Pisitkun T, Shen R-F, Knepper MA. Identification and proteomic profiling of exosomes in human urine. Proc Natl Acad Sci USA. 2004;101:13368.
Bonnerot C, Vincendeau-Scherrer C, Lankar D, Raposo G, Caby M-P. Exosomal-like vesicles are present in human blood plasma. Int Immunol. 2005;17:879–87.
Ogawa Y, Miura Y, Harazono A, Kanai-Azuma M, Akimoto Y, Kawakami H, et al. Proteomic Analysis of Two Types of Exosomes in Human Whole Saliva. Biol Pharm Bull. 2011;34:13–23.
Sharma P, Diergaarde B, Ferrone S, Kirkwood JM, Whiteside TL. Melanoma cell-derived exosomes in plasma of melanoma patients suppress functions of immune effector cells. Sci Rep. 2020;10:1–11.
Simpson RJ, Lim JWE, Moritz RL, Mathivanan S. Exosomes: proteomic insights and diagnostic potential. Expert Rev Proteomics. 2009;6:267–83.
Choi D-S, Lee J, Go G, Kim Y-K, Gho YS. Circulating Extracellular Vesicles in Cancer Diagnosis and Monitoring. Mol Diagn Ther. 2013;17:265–71.
Alderton GK. Fishing for exosomes. Nat Rev Cancer. 2015;15:453.
Zhang W, Jiang L, Diefenbach RJ, Campbell DH, Walsh BJ, Packer NH, et al. Enabling Sensitive Phenotypic Profiling of Cancer-Derived Small Extracellular Vesicles Using Surface-Enhanced Raman Spectroscopy Nanotags. ACS Sens. 2020;5:764–71.
Lu M, Huang Y. Bioinspired exosome-like therapeutics and delivery nanoplatforms. Biomaterials. 2020;242:119925.
Jiang L, Gu Y, Du Y, Liu J. Exosomes: diagnostic biomarkers and therapeutic delivery vehicles for cancer. Mol Pharm. 2019;16:3333–49.
Qiao L, Hu S, Huang K, Su T, Li Z, Vandergriff A, et al. Tumor cell-derived exosomes home to their cells of origin and can be used as Trojan horses to deliver cancer drugs. Theranostics. 2020;10:3474.
Massoumi RL, Hines OJ, Eibl G, King JC. Emerging Evidence for the Clinical Relevance of Pancreatic Cancer Exosomes. Pancreas. 2019;48:1–8.
Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367:eaau6977.
Li W, Li C, Zhou T, Liu X, Liu X, Li X, et al. Role of exosomal proteins in cancer diagnosis. Mol Cancer. 2017;16:145.
Erb U, Zöller M. Progress and potential of exosome analysis for early pancreatic cancer detection. Expert Rev Mol Diagn. 2016;16:757–67.
Tai YL, Chu PY, Lee BH, Chen KC, Yang CY, Kuo WH, et al. Basics and applications of tumor-derived extracellular vesicles. J Biomed Sci. 2019;26:35.
Zhang Y, Bi J, Huang J, Tang Y, Du S, Li P. Exosome. A Review of Its Classification, Isolation Techniques, Storage, Diagnostic and Targeted Therapy Applications. Int J Nanomedicine. 2020;15:6917–34.
Qian L, Yu S, Chen Z, Meng Z, Huang S, Wang P. Functions and clinical implications of exosomes in pancreatic cancer. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer. 2019; 1871: 75–84.
Zhao G, Zhou A, Li X, Zhu S, Wang Y, Zhang S, et al. The Significance of Exosomal RNAs in the Development, Diagnosis, and Treatment of Gastric Cancer. Genes. 2021;12:73.
Zhao Z, Zhao G, Yang S, Zhu S, Zhang S, Li P. The significance of exosomal RNAs in the development, diagnosis, and treatment of pancreatic cancer. Cancer Cell Int. 2021;21:1–16.
Yee NS, Zhang S, He H-Z, Zheng S-Y. Extracellular Vesicles as Potential Biomarkers for Early Detection and Diagnosis of Pancreatic Cancer. 2020; 8: 581.
Xu Y-F, Hannafon BN, Zhao YD, Postier RG, Ding W-Q. Plasma exosome miR-196a and miR-1246 are potential indicators of localized pancreatic cancer. 2017; 8: 77028–40.
Joshi GK, Deitz-McElyea S, Liyanage T, Lawrence K, Mali S, Sardar R, et al. Label-Free Nanoplasmonic-Based Short Noncoding RNA Sensing at Attomolar Concentrations Allows for Quantitative and Highly Specific Assay of MicroRNA-10b in Biological Fluids and Circulating Exosomes. ACS Nano. 2015;9:11075–89.
Takahasi K, Iinuma H, Wada K, Minezaki S, Kawamura S, Kainuma M, et al. Usefulness of exosome-encapsulated microRNA-451a as a minimally invasive biomarker for prediction of recurrence and prognosis in pancreatic ductal adenocarcinoma. 2018; 25: 155–61.
Marin AM, Mattar SB, Amatuzzi RF, Chammas R, Uno M, Zanette DL, et al. Plasma Exosome-Derived microRNAs as Potential Diagnostic and Prognostic Biomarkers in Brazilian Pancreatic Cancer Patients. Biomolecules. 2022;12:769.
Wang C, Wang J, Cui W, Liu Y, Zhou H, Wang Y, et al. Serum Exosomal miRNA-1226 as Potential Biomarker of Pancreatic Ductal Adenocarcinoma. Onco Targets Ther. 2021;14:1441–51.
Que R, Ding G, Chen J, Cao L. Analysis of serum exosomal microRNAs and clinicopathologic features of patients with pancreatic adenocarcinoma. World J Surg Oncol. 2013;11:219.
Verel-Yilmaz Y, Fernández JP, Schäfer A, Nevermann S, Cook L, Gercke N, et al. Extracellular Vesicle-Based Detection of Pancreatic Cancer. Frontiers in Cell and Developmental Biology. 2021; 9.
Goto T, Fujiya M, Konishi H, Sasajima J, Fujibayashi S, Hayashi A, et al. An elevated expression of serum exosomal microRNA-191, – 21, – 451a of pancreatic neoplasm is considered to be efficient diagnostic marker. BMC Cancer. 2018;18:116.
Madhavan B, Yue S, Galli U, Rana S, Gross W, Müller M, et al. Combined evaluation of a panel of protein and miRNA serum-exosome biomarkers for pancreatic cancer diagnosis increases sensitivity and specificity. Int J Cancer. 2015;136:2616–27.
Machida T, Tomofuji T, Maruyama T, Yoneda T, Ekuni D, Azuma T, et al. miR–1246 and miR–4644 in salivary exosome as potential biomarkers for pancreatobiliary tract cancer. Oncol Rep. 2016;36:2375–81.
Nakamura S, Sadakari Y, Ohtsuka T, Okayama T, Nakashima Y, Gotoh Y, et al. Pancreatic Juice Exosomal MicroRNAs as Biomarkers for Detection of Pancreatic Ductal Adenocarcinoma. Ann Surg Oncol. 2019;26:2104–11.
Hu J, Sheng Y, Kwak KJ, Shi J, Yu B, Lee LJ. A signal-amplifiable biochip quantifies extracellular vesicle-associated RNAs for early cancer detection. Nat Commun. 2017;8:1683.
Yang Z, LaRiviere MJ, Ko J, Till JE, Christensen T, Yee SS, et al. A Multianalyte Panel Consisting of Extracellular Vesicle miRNAs and mRNAs, cfDNA, and CA19-9 Shows Utility for Diagnosis and Staging of Pancreatic Ductal Adenocarcinoma. 2020; 26: 3248–58.
Lau C, Kim Y, Chia D, Spielmann N, Eibl G, Elashoff D, et al. Role of Pancreatic Cancer-derived Exosomes in Salivary Biomarker Development. J Biol Chem. 2013;288:26888–97.
Li Z, Jiang P, Li J, Peng M, Zhao X, Zhang X, et al. Tumor-derived exosomal lnc-Sox2ot promotes EMT and stemness by acting as a ceRNA in pancreatic ductal adenocarcinoma. Oncogene. 2018;37:3822–38.
Takahashi K, Ota Y, Kogure T, Suzuki Y, Iwamoto H, Yamakita K, et al. Circulating extracellular vesicle-encapsulated HULC is a potential biomarker for human pancreatic cancer. Cancer Sci. 2020;111:98–111.
Kumar SR, Kimchi ET, Manjunath Y, Gajagowni S, Stuckel AJ, Kaifi JT. RNA cargos in extracellular vesicles derived from blood serum in pancreas associated conditions. Sci Rep. 2020;10:2800.
Yu S, Li Y, Liao Z, Wang Z, Wang Z, Li Y, et al. Plasma extracellular vesicle long RNA profiling identifies a diagnostic signature for the detection of pancreatic ductal adenocarcinoma. 2020; 69: 540–50.
Li J, Li Z, Jiang P, Peng M, Zhang X, Chen K, et al. Circular RNA IARS (circ-IARS) secreted by pancreatic cancer cells and located within exosomes regulates endothelial monolayer permeability to promote tumor metastasis. J Experimental Clin Cancer Res. 2018;37:177.
Li Z, Yanfang W, Li J, Jiang P, Peng T, Chen K, et al. Tumor-released exosomal circular RNA PDE8A promotes invasive growth via the miR-338/MACC1/MET pathway in pancreatic cancer. Cancer Lett. 2018;432:237–50.
Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136:215–33.
Chiang HR, Schoenfeld LW, Ruby JG, Auyeung VC, Spies N, Baek D, et al. Mammalian microRNAs: experimental evaluation of novel and previously annotated genes. Genes Dev. 2010;24:992–1009.
Zhang X, Xia D, Wang R-X, Zhang Y-T, Zhang S-Y, Yang C, et al. Identification of potential biomarkers for digestive system cancers from serum-derived extracellular vesicle RNA. Clin Chim Acta. 2022;531:36–47.
Liu B, Yang H, Taher L, Denz A, Grützmann R, Pilarsky C, et al. Identification of Prognostic Biomarkers by Combined mRNA and miRNA Expression Microarray Analysis in Pancreatic Cancer. Translational Oncol. 2018;11:700–14.
Gupta RA, Shah N, Wang KC, Kim J, Horlings HM, Wong DJ, et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010;464:1071–6.
Costa-Silva B, Aiello NM, Ocean AJ, Singh S, Zhang H, Thakur Basant K, et al. Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver. Nat Cell Biol. 2015;17:816–26.
Chevillet JR, Kang Q, Ruf IK, Briggs HA, Vojtech LN, Hughes SM, et al. Quantitative and stoichiometric analysis of the microRNA content of exosomes. Proceedings of the National Academy of Sciences. 2014; 111: 14888-93.
Kahlert C, Melo SA, Protopopov A, Tang J, Seth S, Koch M, et al. Identification of double-stranded genomic DNA spanning all chromosomes with mutated KRAS and p53 DNA in the serum exosomes of patients with pancreatic cancer. J Biol Chem. 2014;289:3869–75.
Thakur BK, Zhang H, Becker A, Matei I, Huang Y, Costa-Silva B, et al. Double-stranded DNA in exosomes: a novel biomarker in cancer detection. Cell Res. 2014;24:766–9.
Wan Y, Maurer M, He H-Z, Xia Y-Q, Hao S-J, Zhang W-L, et al. Enrichment of extracellular vesicles with lipid nanoprobe functionalized nanostructured silica. Lab Chip. 2019;19:2346–55.
Allenson K, Castillo J, San Lucas FA, Scelo G, Kim DU, Bernard V, et al. High prevalence of mutantKRAS in circulating exosome-derived DNA from early-stage pancreatic cancer patients. Ann Oncol. 2017;28:741–7.
Bernard V, Kim DU, San Lucas FA, Castillo J, Allenson K, Mulu FC, et al. Circulating Nucleic Acids Are Associated With Outcomes of Patients With Pancreatic Cancer. Gastroenterology. 2019; 156: 108 – 18.e4.
San Lucas FA, Allenson K, Bernard V, Castillo J, Kim DU, Ellis K, et al. Minimally invasive genomic and transcriptomic profiling of visceral cancers by next-generation sequencing of circulating exosomes. Ann Oncol. 2016;27:635–41.
Yang S, Che SPY, Kurywchak P, Tavormina JL, Gansmo LB, Correa de Sampaio P, et al. Detection of mutant KRAS and TP53 DNA in circulating exosomes from healthy individuals and patients with pancreatic cancer. Cancer Biol Ther. 2017;18:158–65.
Yamamoto H, Watanabe Y, Oikawa R, Morita R, Yoshida Y, Maehata T, et al. BARHL2Methylation Using Gastric Wash DNA or Gastric Juice Exosomal DNA is a Useful Marker For Early Detection of Gastric Cancer in anH. pylori-Independent Manner. Clin Translational Gastroenterol. 2016;7:e184.
ExoCarta. http://wwwexocartaorg/. Accessed 20 August 2022.
Wu H, Chen X, Ji J, Zhou R, Liu J, Ni W, et al. Progress of Exosomes in the Diagnosis and Treatment of Pancreatic Cancer. Genetic Test Mol biomarkers. 2019;23:215–22.
Yang KS, Im H, Hong S, Pergolini I, Del Castillo AF, Wang R, et al. Multiparametric plasma EV profiling facilitates diagnosis of pancreatic malignancy. Sci Transl Med. 2017;9:eaal3226.
Melo SA, Luecke LB, Kahlert C, Fernandez AF, Gammon ST, Kaye J, et al. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature. 2015;523:177–82.
Jin H, Liu P, Wu Y, Meng X, Wu M, Han J, et al. Exosomal zinc transporter ZIP4 promotes cancer growth and is a novel diagnostic biomarker for pancreatic cancer. Cancer Sci. 2018;109:2946–56.
Liu C, Zhao J, Tian F, Cai L, Zhang W, Feng Q, et al. Low-cost thermophoretic profiling of extracellular-vesicle surface proteins for the early detection and classification of cancers. Nat Biomedical Eng. 2019;3:183–93.
Frampton AE, Prado MM, López-Jiménez E, Fajardo-Puerta AB, Jawad ZAR, Lawton P, et al. Glypican-1 is enriched in circulating-exosomes in pancreatic cancer and correlates with tumor burden. Oncotarget. 2018;9:19006–13.
Giampieri R, Piva F, Occhipinti G, Bittoni A, Righetti A, Pagliaretta S, et al. Clinical impact of different exosomes’ protein expression in pancreatic ductal carcinoma patients treated with standard first line palliative chemotherapy. PLoS ONE. 2019;14:e0215990.
Liang K, Liu F, Fan J, Sun D, Liu C, Lyon CJ, et al. Nanoplasmonic Quantification of Tumor-derived Extracellular Vesicles in Plasma Microsamples for Diagnosis and Treatment Monitoring. Nat Biomed Eng. 2017;1:0021.
Ferguson S, Yang KS, Zelga P, Liss AS, Carlson JC, Del Castillo CF, et al. Single-EV analysis (sEVA) of mutated proteins allows detection of stage 1 pancreatic cancer. Sci Adv. 2022;8:eabm3453.
Lennon KM, Wakefield DL, Maddox AL, Brehove MS, Willner AN, Garcia-Mansfield K, et al. Single molecule characterization of individual extracellular vesicles from pancreatic cancer. J Extracell Vesicles. 2019;8:1685634.
Lewis JM, Vyas AD, Qiu Y, Messer KS, Heller MJJAN. Integrated Analysis of Exosomal Protein Biomarkers on Alternating Current Electrokinetic Chips Enables Rapid Detection of Pancreatic Cancer in Patient Blood. 2018; 12: 3311–20.
Zhang W, Wang L, Li D, Campbel DH, Walsh BJ, Packer NH, et al. Phenotypic profiling of pancreatic ductal adenocarcinoma plasma-derived small extracellular vesicles for cancer diagnosis and cancer stage prediction: a proof-of-concept study. Anal Methods. 2022;14:2255–65.
Dong C, Huang L, Melo SA, Kurywchak P, Peng Q, Kahlert C, et al. Multiple antibodies identify glypican-1 associated with exosomes from pancreatic cancer cells and serum from patients with pancreatic cancer. bioRxiv; 2018: 145706.
Moutinho-Ribeiro P, Adem B, Batista I, Silva M, Silva S, Ruivo CF, et al. Exosomal glypican-1 discriminates pancreatic ductal adenocarcinoma from chronic pancreatitis. Dig Liver Disease. 2022;54:871–7.
Lux A, Kahlert C, Grützmann R, Pilarsky C. c-Met and PD-L1 on Circulating Exosomes as Diagnostic and Prognostic Markers for Pancreatic Cancer. Int J Mol Sci. 2019;20:3305.
Kimura H, Yamamoto H, Harada T, Fumoto K, Osugi Y, Sada R, et al. CKAP4, a DKK1 Receptor, Is a Biomarker in Exosomes Derived from Pancreatic Cancer and a Molecular Target for Therapy. Clin Cancer Res. 2019;25:1936–47.
Leca J, Martinez S, Lac S, Nigri J, Secq V, Rubis M, et al. Cancer-associated fibroblast-derived annexin A6 + extracellular vesicles support pancreatic cancer aggressiveness. J Clin Investig. 2016;126:4140–56.
Odaka H, Hiemori K, Shimoda A, Akiyoshi K, Tateno H. CD63-positive extracellular vesicles are potential diagnostic biomarkers of pancreatic ductal adenocarcinoma. BMC Gastroenterol. 2022;22:153.
Xie Z, Gao Y, Ho C, Li L, Jin C, Wang X, et al. Exosome-delivered CD44v6/C1QBP complex drives pancreatic cancer liver metastasis by promoting fibrotic liver microenvironment. Gut. 2022;71:568–79.
Li J, Li Y, Chen S, Duan W, Kong X, Wang Y, et al. Highly Sensitive Exosome Detection for Early Diagnosis of Pancreatic Cancer Using Immunoassay Based on Hierarchical Surface-Enhanced Raman Scattering Substrate. Small Methods. 2022;6:2200154.
Asada T, Nakahata S, Fauzi YR, Ichikawa T, Inoue K, Shibata N, et al. Integrin α6A (ITGA6A)-type Splice Variant in Extracellular Vesicles Has a Potential as a Novel Marker of the Early Recurrence of Pancreatic Cancer. Anticancer Res. 2022;42:1763.
Osteikoetxea X, Benke M, Rodriguez M, Pálóczi K, Sódar BW, Szvicsek Z, et al. Detection and proteomic characterization of extracellular vesicles in human pancreatic juice. Biochem Biophys Res Commun. 2018;499:37–43.
Zheng J, Hernandez JM, Doussot A, Bojmar L, Zambirinis CP, Costa-Silva B, et al. Extracellular matrix proteins and carcinoembryonic antigen-related cell adhesion molecules characterize pancreatic duct fluid exosomes in patients with pancreatic cancer. HPB. 2018;20:597–604.
Buscail E, Chauvet A, Quincy P, Degrandi O, Buscail C, Lamrissi I, et al. CD63-GPC1-Positive Exosomes Coupled with CA19-9 Offer Good Diagnostic Potential for Resectable Pancreatic Ductal Adenocarcinoma. Translational Oncol. 2019;12:1395–403.
Hinestrosa JP, Lewis JM, Balcer HI, Kurzrock R, Lippman S, Krishnan R. Abstract 5960: Blood-based extracellular vesicle biomarker test for detection of early-stage pancreatic cancer. Cancer Res. 2022;82:5960.
Inoue H, Eguchi A, Kobayashi Y, Usugi E, Yamada R, Tsuboi J, et al. Extracellular vesicles from pancreatic ductal adenocarcinoma endoscopic ultrasound-fine needle aspiration samples contain a protein barcode. J Hepato-Biliary-Pancreat Sci. 2022;29:394–403.
Donoso-Quezada J, Ayala-Mar S, González-Valdez J. The role of lipids in exosome biology and intercellular communication: Function, analytics and applications. 2021; 22: 204 – 20.
Record M, Carayon K, Poirot M, Silvente-Poirot S. Exosomes as new vesicular lipid transporters involved in cell-cell communication and various pathophysiologies. Biochim Biophys Acta. 2014;1841:108–20.
Skotland T, Ekroos K, Kauhanen D, Simolin H, Seierstad T, Berge V, et al. Molecular lipid species in urinary exosomes as potential prostate cancer biomarkers. European journal of cancer (Oxford, England: 1990). 2017; 70: 122 – 32.
Fan TWM, Zhang X, Wang C, Yang Y, Kang W-Y, Arnold S, et al. Exosomal lipids for classifying early and late stage non-small cell lung cancer. Anal Chim Acta. 2018;1037:256–64.
Eylem CC, Yilmaz M, Derkus B, Nemutlu E, Camci CB, Yilmaz E, et al. Untargeted multi-omic analysis of colorectal cancer-specific exosomes reveals joint pathways of colorectal cancer in both clinical samples and cell culture. Cancer Lett. 2020;469:186–94.
Sharma R, Huang X, Brekken RA, Schroit AJ. Detection of phosphatidylserine-positive exosomes for the diagnosis of early-stage malignancies. Br J Cancer. 2017;117:545–52.
Linton SS, Abraham T, Liao J, Clawson GA, Butler PJ, Fox T, et al. Tumor-promoting effects of pancreatic cancer cell exosomes on THP-1-derived macrophages. PLoS ONE. 2018;13:e0206759.
Hinzman CP, Singh B, Bansal S, Li Y, Iliuk A, Girgis M, et al. A multi-omics approach identifies pancreatic cancer cell extracellular vesicles as mediators of the unfolded protein response in normal pancreatic epithelial cells. J Extracell Vesicles. 2022;11:e12232.
Tao L, Zhou J, Yuan C, Zhang L, Li D, Si D, et al. Metabolomics identifies serum and exosomes metabolite markers of pancreatic cancer. Metabolomics. 2019;15:86.
Chen Y, Ding L, Ju H. In Situ Cellular Glycan Analysis. Acc Chem Res. 2018;51:890–9.
Scott E, Munkley J. Glycans as Biomarkers in Prostate Cancer. Int J Mol Sci. 2019;20:1389.
Díaz-Fernández A, Miranda-Castro R, de-los-Santos-Álvarez N, Lobo-Castañón MJ, Estrela P. Impedimetric aptamer-based glycan PSA score for discrimination of prostate cancer from other prostate diseases. Biosens Bioelectron. 2021;175:112872.
Munkley J. The glycosylation landscape of pancreatic cancer (Review). Oncol Lett. 2019;17:2569–75.
Engle DD, Tiriac H, Rivera KD, Pommier A, Whalen S, Oni TE, et al. The glycan CA19-9 promotes pancreatitis and pancreatic cancer in mice. Science. 2019;364:1156–62.
Yokose T, Kabe Y, Matsuda A, Kitago M, Matsuda S, Hirai M, et al. O-Glycan-Altered Extracellular Vesicles: A Specific Serum Marker Elevated in Pancreatic Cancer. Cancers. 2020;12:2469.
Choi Y, Park U, Koo H-J, Park J-s, Lee DH, Kim K, et al. Exosome-mediated diagnosis of pancreatic cancer using lectin-conjugated nanoparticles bound to selective glycans. Biosens Bioelectron. 2021;177:112980.
Tutrone R, Donovan MJ, Torkler P, Tadigotla V, McLain T, Noerholm M, et al. Clinical utility of the exosome based ExoDx Prostate(IntelliScore) EPI test in men presenting for initial Biopsy with a PSA 2–10 ng/mL. Prostate Cancer Prostatic Dis. 2020;23:607–14.
Enderle D, Noerholm M. Are extracellular vesicles ready for the clinical laboratory? LaboratoriumsMedizin. 2022.
Becker A, Thakur BK, Weiss JM, Kim HS, Peinado H, Lyden D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell. 2016;30:836–48.
Pullan JE, Confeld MI, Osborn JK, Kim J, Sarkar K, Mallik S. Exosomes as Drug Carriers for Cancer Therapy. Mol Pharm. 2019;16:1789–98.
Richards KE, Zeleniak AE, Fishel ML, Wu J, Littlepage LE, Hill R. Cancer-associated fibroblast exosomes regulate survival and proliferation of pancreatic cancer cells. Oncogene. 2017;36:1770–8.
Xiao W, Pahlavanneshan M, Eun C-Y, Zhang X, DeKalb C, Mahgoub B, et al. Matrix stiffness mediates pancreatic cancer chemoresistance through induction of exosome hypersecretion in a cancer associated fibroblasts-tumor organoid biomimetic model. Matrix Biology Plus. 2022;14:100111.
Mikamori M, Yamada D, Eguchi H, Hasegawa S, Kishimoto T, Tomimaru Y, et al. MicroRNA-155 Controls Exosome Synthesis and Promotes Gemcitabine Resistance in Pancreatic Ductal Adenocarcinoma. Sci Rep. 2017;7:42339.
Richards KE, Xiao W, Hill R, Team obotUPR. Cancer-Associated Fibroblasts Confer Gemcitabine Resistance to Pancreatic Cancer Cells through PTEN-Targeting miRNAs in Exosomes. Cancers. 2022;14:2812.
Hoshino A, Costa-Silva B, Shen T-L, Rodrigues G, Hashimoto A, Tesic Mark M, et al. Tumour exosome integrins determine organotropic metastasis. Nature. 2015;527:329–35.
Wang C-A, Chang I-H, Hou P-C, Tai Y-J, Li W-N, Hsu P-L, et al. DUSP2 regulates extracellular vesicle-VEGF-C secretion and pancreatic cancer early dissemination. J Extracell vesicles. 2020;9:1746529.
Li Y-J, Wu J-Y, Wang J-M, Hu X-B, Cai J-X, Xiang D-X. Gemcitabine loaded autologous exosomes for effective and safe chemotherapy of pancreatic cancer. Acta Biomater. 2020;101:519–30.
Aspe JR, Diaz Osterman CJ, Jutzy JMS, Deshields S, Whang S, Wall NR. Enhancement of Gemcitabine sensitivity in pancreatic adenocarcinoma by novel exosome-mediated delivery of the Survivin-T34A mutant. J Extracell Vesicles. 2014;3:23244.
Al Faruque H, Choi E-S, Kim J-H, Kim E. Enhanced effect of autologous EVs delivering paclitaxel in pancreatic cancer. J Controlled Release. 2022;347:330–46.
Xu L, Faruqu FN, Lim YM, Lim KY, Liam-Or R, Walters AA, et al. Exosome-mediated RNAi of PAK4 prolongs survival of pancreatic cancer mouse model after loco-regional treatment. Biomaterials. 2021;264:120369.
Han S, Gonzalo DH, Feely M, Rinaldi C, Belsare S, Zhai H, et al. Stroma-derived extracellular vesicles deliver tumor-suppressive miRNAs to pancreatic cancer cells. Oncotarget. 2018;9:5764.
Que R-s, Lin C, Ding G-p, Wu Z-r. Cao L-p. Increasing the immune activity of exosomes: the effect of miRNA-depleted exosome proteins on activating dendritic cell/cytokine-induced killer cells against pancreatic cancer. J Zhejiang University-SCIENCE B. 2016;17:352–60.
Gastpar R, Gehrmann M, Bausero MA, Asea A, Gross C, Schroeder JA, et al. Heat shock protein 70 surface-positive tumor exosomes stimulate migratory and cytolytic activity of natural killer cells. Cancer Res. 2005;65:5238–47.
Zech D, Rana S, Büchler MW, Zöller M. Tumor-exosomes and leukocyte activation: an ambivalent crosstalk. Cell Communication and Signaling. 2012;10:37.
Mahmoodzadeh Hosseini H, Ali Imani Fooladi A, Soleimanirad J, Reza Nourani M, Mahdavi M. Exosome/staphylococcal enterotoxin B, an anti tumor compound against pancreatic cancer. J BUON. 2014;19:440–8.
Zhou W, Chen X, Zhou Y, Shi S, Liang C, Yu X, et al. Exosomes derived from immunogenically dying tumor cells as a versatile tool for vaccination against pancreatic cancer. Biomaterials. 2022;280:121306.
Lau C, Kim Y, Chia D, Spielmann N, Eibl G, Elashoff D, et al. Role of pancreatic cancer-derived exosomes in salivary biomarker development. J Biol Chem. 2013;288:26888–97.
Wang M, Su Z, Amoah Barnie P. Crosstalk among colon cancer-derived exosomes, fibroblast-derived exosomes, and macrophage phenotypes in colon cancer metastasis. Int Immunopharmacol. 2020;81:106298.
Binenbaum Y, Fridman E, Yaari Z, Milman N, Schroeder A, Ben David G, et al. Transfer of miRNA in Macrophage-Derived Exosomes Induces Drug Resistance in Pancreatic Adenocarcinoma. Cancer Res. 2018;78:5287.
Ostenfeld MS, Jeppesen DK, Laurberg JR, Boysen AT, Bramsen JB, Primdal-Bengtson B, et al. Cellular Disposal of miR23b by RAB27-Dependent Exosome Release Is Linked to Acquisition of Metastatic Properties. Cancer Res. 2014;74:5758.
Fares J, Kashyap R, Zimmermann P. Syntenin. Key player in cancer exosome biogenesis and uptake? Cell Adhes Migr. 2017;11:124–6.
Christianson HC, Svensson KJ, van Kuppevelt TH, Li J-P, Belting M. Cancer cell exosomes depend on cell-surface heparan sulfate proteoglycans for their internalization and functional activity. Proceedings of the National Academy of Sciences. 2013; 110: 17380.
Hao Y, Song H, Zhou Z, Chen X, Li H, Zhang Y, et al. Promotion or inhibition of extracellular vesicle release: Emerging therapeutic opportunities. J Controlled Release. 2021;340:136–48.
Rezaie J, Akbari A, Rahbarghazi R. Inhibition of extracellular vesicle biogenesis in tumor cells: A possible way to reduce tumorigenesis. Cell Biochem Funct. 2022;40:248–62.
Catalano M, O’Driscoll L. Inhibiting extracellular vesicles formation and release: a review of EV inhibitors. J Extracell vesicles. 2020;9:1703244.
Rosenfeldt MT, O’Prey J, Flossbach L, Nixon C, Morton JP, Sansom OJ, et al. PTEN deficiency permits the formation of pancreatic cancer in the absence of autophagy. Cell Death & Differentiation. 2017;24:1303–4.
Si W, Liu X, Wei R, Zhang Y, Zhao Y, Cui L, et al. MTA2-mediated inhibition of PTEN leads to pancreatic ductal adenocarcinoma carcinogenicity. Cell Death Dis. 2019;10:206.
Capello M, Vykoukal JV, Katayama H, Bantis LE, Wang H, Kundnani DL, et al. Exosomes harbor B cell targets in pancreatic adenocarcinoma and exert decoy function against complement-mediated cytotoxicity. Nat Commun. 2019;10:254.
Vader P, Mol EA, Pasterkamp G, Schiffelers RM. Extracellular vesicles for drug delivery. Adv Drug Deliv Rev. 2016;106:148–56.
Kamerkar S, LeBleu VS, Sugimoto H, Yang S, Ruivo CF, Melo SA, et al. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature. 2017;546:498–503.
Kim MS, Haney MJ, Zhao Y, Mahajan V, Deygen I, Klyachko NL, et al. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomed Nanotechnol Biol Med. 2016;12:655–64.
Lamichhane TN, Jeyaram A, Patel DB, Parajuli B, Livingston NK, Arumugasaamy N, et al. Oncogene Knockdown via Active Loading of Small RNAs into Extracellular Vesicles by Sonication. Cell Mol Bioeng. 2016;9:315–24.
Rahbarghazi R, Jabbari N, Sani NA, Asghari R, Salimi L, Kalashani SA, et al. Tumor-derived extracellular vesicles: reliable tools for Cancer diagnosis and clinical applications. Cell Communication and Signaling. 2019;17:1–17.
Fais S, O’Driscoll L, Borras FE, Buzas E, Camussi G, Cappello F, et al. Evidence-Based Clinical Use of Nanoscale Extracellular Vesicles in Nanomedicine. ACS Nano. 2016;10:3886–99.
Walker S, Busatto S, Pham A, Tian M, Suh A, Carson K, et al. Extracellular vesicle-based drug delivery systems for cancer treatment. Theranostics. 2019;9:8001.
Surana R, LeBleu VS, Lee JJ, Smaglo BG, Zhao D, Lee MS, et al. Phase I study of mesenchymal stem cell (MSC)-derived exosomes with KRASG12D siRNA in patients with metastatic pancreatic cancer harboring a KRASG12D mutation. J Clin Oncol. 2022;40:TPS633-TPS.
Ou Y-H, Liang J, Czarny B, Wacker MG, Yu V, Wang J-W, et al. Extracellular Vesicle (EV) biohybrid systems for cancer therapy: Recent advances and future perspectives. Seminars in Cancer Biology: Elsevier; 2021. p.45–61.
Xu L, Faruqu FN, Liam-Or R, Abu Abed O, Li D, Venner K, et al. Design of experiment (DoE)-driven in vitro and in vivo uptake studies of exosomes for pancreatic cancer delivery enabled by copper-free click chemistry-based labelling. J Extracell vesicles. 2020;9:1779458.
Xavier CP, Castro I, Caires HR, Ferreira D, Cavadas B, Pereira L, et al. Chitinase 3-like-1 and fibronectin in the cargo of extracellular vesicles shed by human macrophages influence pancreatic cancer cellular response to gemcitabine. Cancer Lett. 2021;501:210–23.
Gilligan KE, Dwyer RM. Engineering exosomes for cancer therapy. Int J Mol Sci. 2017;18:1122.
Liang Y, Duan L, Lu J, Xia J. Engineering exosomes for targeted drug delivery. Theranostics. 2021;11:3183–95.
Dai S, Wei D, Wu Z, Zhou X, Wei X, Huang H, et al. Phase I clinical trial of autologous ascites-derived exosomes combined with GM-CSF for colorectal cancer. Mol Ther. 2008;16:782–90.
Guo M, Wu F, Hu G, Chen L, Xu J, Xu P, et al. Autologous tumor cell–derived microparticle-based targeted chemotherapy in lung cancer patients with malignant pleural effusion. Sci Transl Med. 2019;11:eaat5690.
Ma J, Zhang Y, Tang K, Zhang H, Yin X, Li Y, et al. Reversing drug resistance of soft tumor-repopulating cells by tumor cell-derived chemotherapeutic microparticles. Cell Res. 2016;26:713–27.
Tang K, Zhang Y, Zhang H, Xu P, Liu J, Ma J, et al. Delivery of chemotherapeutic drugs in tumour cell-derived microparticles. Nat Commun. 2012;3:1282.
Meng W, He C, Hao Y, Wang L, Li L, Zhu G. Prospects and challenges of extracellular vesicle-based drug delivery system: Considering cell source. Drug Delivery. 2020;27:585–98.
Yamashita T, Takahashi Y, Takakura Y. Possibility of exosome-based therapeutics and challenges in production of exosomes eligible for therapeutic application. Biol Pharm Bull. 2018;41:835–42.
de Almeida Fuzeta M, Bernardes N, Oliveira FD, Costa AC, Fernandes-Platzgummer A, Farinha JP, et al. Scalable production of human mesenchymal stromal cell-derived extracellular vesicles under serum-/xeno-free conditions in a microcarrier-based bioreactor culture system. Front cell Dev biology. 2020;8:553444.
Yi YW, Lee JH, Kim S-Y, Pack C-G, Ha DH, Park SR, et al. Advances in analysis of biodistribution of exosomes by molecular imaging. Int J Mol Sci. 2020;21:665.
Sun H, Shi K, Qi K, Kong H, Zhang J, Dai S, et al. Natural Killer Cell-Derived Exosomal miR-3607-3p Inhibits Pancreatic Cancer Progression by Targeting IL-26. Front Immunol. 2019;10:2819.
Shang S, Wang J, Chen S, Tian R, Zeng H, Wang L, et al. Exosomal miRNA-1231 derived from bone marrow mesenchymal stem cells inhibits the activity of pancreatic cancer. Cancer Med. 2019;8:7728–40.
Huber M, Brehm CU, Gress TM, Buchholz M, Alashkar Alhamwe B, von Strandmann EP, et al. The Immune Microenvironment in Pancreatic Cancer. Int J Mol Sci. 2020;21:7307.
Hofmann L, Ludwig S, Vahl JM, Brunner C, Hoffmann TK, Theodoraki M-N. The Emerging Role of Exosomes in Diagnosis, Prognosis, and Therapy in Head and Neck Cancer. Int J Mol Sci. 2020;21:4072.
Whiteside TL. Exosomes and tumor-mediated immune suppression. J Clin Investig. 2016;126:1216–23.
Sun W, Luo J-d, Jiang H, Duan DD. Tumor exosomes: a double-edged sword in cancer therapy. Acta Pharmacol Sin. 2018;39:534–41.
Gehrmann U, Näslund TI, Hiltbrunner S, Larssen P, Gabrielsson S. Harnessing the exosome-induced immune response for cancer immunotherapy. Sem Cancer Biol. 2014;28:58–67.
Gu X, Erb U, Büchler MW, Zöller M. Improved vaccine efficacy of tumor exosome compared to tumor lysate loaded dendritic cells in mice. Int J Cancer. 2015;136:E74–84.
Koyama Y, Ito T, Hasegawa A, Eriguchi M, Inaba T, Ushigusa T, et al. Exosomes derived from tumor cells genetically modified to express Mycobacterium tuberculosis antigen: a novel vaccine for cancer therapy. Biotechnology letters. 2016; 38: 1857–66.
Acknowledgements
WZ acknowledges the support from Macquarie University Research Excellence Scholarship (MQRES).
Author information
Authors and Affiliations
Contributions
WZ conceived and wrote the manuscript. DHC, BJW and NHP reviewed and edited the manuscript. DL and YW contributed to the conceptualization, revised the original draft and provided the supervision. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors have approved the final draft of this manuscript for submission and have given consent for the publication of identifiable details.
Competing interests
The authors declare no competing financial interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, W., Campbell, D.H., Walsh, B.J. et al. Cancer-derived small extracellular vesicles: emerging biomarkers and therapies for pancreatic ductal adenocarcinoma diagnosis/prognosis and treatment. J Nanobiotechnol 20, 446 (2022). https://doi.org/10.1186/s12951-022-01641-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12951-022-01641-0