
Abstract
Background
Gut microbiota plays an important role in many metabolic diseases such as diabetes and atherosclerosis. Apolipoprotein E (apoE) knock-out (KO) mice are frequently used for the study of hyperlipidemia and atherosclerosis. However, it is unknown whether apoE KO mice have altered gut microbiota when challenged with a Western diet.
Methods
In the current study, we assessed the gut microbiota profiling of apoE KO mice and compared with wild-type mice fed either a normal chow or Western diet for 12 weeks using 16S pyrosequencing.
Results
On a western diet, the gut microbiota diversity was significantly decreased in apoE KO mice compared with wild type (WT) mice. Firmicutes and Erysipelotrichaceae were significantly increased in WT mice but Erysipelotrichaceae was unchanged in apoE KO mice on a Western diet. The weighted UniFrac principal coordinate analysis exhibited clear separation between WT and apoE KO mice on the first vector (58.6%) with significant changes of two dominant phyla (Bacteroidetes and Firmicutes) and seven dominant families (Porphyromonadaceae, Lachnospiraceae, Ruminococcaceae, Desulfovibrionaceae, Helicobacteraceae, Erysipelotrichaceae and Veillonellaceae). Lachnospiraceae was significantly enriched in apoE KO mice on a Western diet. In addition, Lachnospiraceae and Ruminococcaceae were positively correlated with relative atherosclerosis lesion size in apoE KO.
Conclusions
Collectively, our study showed that there are marked changes in the gut microbiota of apoE KO mice, particularly challenged with a Western diet and these alterations may be possibly associated with atherosclerosis.
Similar content being viewed by others
Background
Excess intake of a high-fat diet or Western diet is generally considered unhealthy because such a diet can cause many metabolic disorders such as obesity, diabetes and atherosclerosis [1,2,3]. Although this notion has been widely accepted, the underlying mechanisms responsible for these pathophysiological consequences are still not fully understood. Gut microbiota has received more attention for its roles in both physiological and pathophysiological processes [4,5,6,7]. It has been shown that many metabolic diseases are related with the changes in gut microbiota. Laboratory mice are frequently used for the study of human diseases but most strains of wild-type mice do not show any metabolic disorders on a normal chow diet. However, if WT mice were fed with a diet containing 21% fat along with 0.15% cholesterol, which is often terminated as a Western diet, they show elevated plasma lipids but they do not show obvious lesions of atherosclerosis. However, if mice are genetically deficient in apolipoprotein E (apoE) such as apoE knock-out (KO) mice, they are sensitive to a Western diet and rapidly develop hyperlipidemia and atherosclerosis. ApoE is a component of all lipoproteins except for low density lipoproteins (LDL) and functions as a ligand for LDL receptors and LDL-related proteins (LRP) on the liver thereby playing an important role in the catabolism of the remnants of chylomicrons and very low-density lipoproteins. The genetic deficiency of apoE gene leads to the accumulation of cholesterol-rich remnants in plasma in both humans and animals [8,9,10,11,12]. Therefore, Western diet-fed apoE KO mice are widely used to study many facets of human hyperlipidemia and atherosclerosis [13,14,15,16].
Given the fact that the gut microbiota plays an important role in many metabolic diseases such as obesity, diabetes, metabolic syndrome [17,18,19,20] and atherosclerosis [21,22,23,24], we envisioned that the gut microbiota may be altered in apoE KO mice when challenged with a Western diet. Driven by the popularization of high throughput sequencing-based approaches, many meta-genomic studies were carried out to characterize the profile changes of the gut microbiome in atherosclerotic individuals [22, 23] and facilitated the comprehensive understanding of the interaction between the gut microbiota and host physiology. However, as the most commonly used atherogenic mouse model, the gut microbiota of apoE KO mice compared with WT mice has not been clarified and their relationship with metabolic and pathophysiological consequences remain unclear. In the current study, we investigated the gut microbiota profiling of apoE KO mice fed a chow and western diet compared with WT mice. Our data revealed that apoE KO mice exhibited unique features of the gut microbiota diversity and compositions and these features may provide novel insight into understanding the relationship between gut microbiota and atherosclerosis.
Methods
Animals
In this study, six-week-old male C57BL/6 J mice (wild type, WT) and apoE KO were used and divided into four groups: WT mice feed on normal chow diet (WT-CD), WT mice feed on western diet (WT-HC), apoE−/− mice feed on normal chow diet (apoE-CD), and apoE KO mice feed on Western diet (apoE-HC) (n = 7~ 8). The composition of normal chow diet is based on the NIH feeds standard (NIH-07, NSN-8710-00-509-7915). The Western diet contains 21% fat and 0.15% cholesterol. The diets were provided by Beijing Keao Xieli Feed Co.,Ltd. All animals were provided ad libitum access to food and water and housed under the conventional 12 h: 12 h light/dark cycle for 12 weeks. The plasma total cholesterol (TC), Triglyceride (TG) and high-density lipoprotein cholesterol (HDL-C) levels were measured at 0, 4, 8, 12 weeks from the beginning of experiment. At the end of the experiment, all mice were sacrificed by a sodium pentobarbital overdose. The heart and artery tissue and intestinal contents were collected for further processing. Animal care and experimental procedures were approved by the Animal Care and Use Committee of Xi’an Jiaotong University.
Plasma lipid measurement
Mice were fasted for 8 h before collecting the blood samples from tail veins. Then blood samples were centrifuged at 3000 g for 15 min and supernatants were collected. The plasma levels of TC, TG, and HDL-C were measured with commercially available kits from Biosino Bio-technology and Science (Beijing) according to the manufacturer’s protocols.
Analysis of atherosclerotic lesions
Mice were euthanized by a sodium pentobarbital overdose and flushed with saline at physiological pressure, then perfused with 4% paraformaldehyde through the left ventricle. The heart and aorta were dissected, removed and fixed with parafonnaldehyde. Aortas were open longitudinally, stained with Oil Red O and digitally scanned. En face lesion area was measured with WinROOF 6.5 (Mitani Co., Fukui, Japan). To quantify atherosclerosis at the aortic root, cross-cryosections were prepared according to our previous report [25]. In brief, part of each heart between the lower tips of the right and left atria were embedded in optimal cutting temperature compound and the aortic root was sectioned (8 μm) serially. Corresponding sections on separate slides were treated with Oil Red O staining and analyzed by WinROOF 6.5.
Immunohistochemistry analysis
For immunohistochemistry, corresponding sections were incubated with 3% hydrogen peroxide for 30 min and blocked with 2% horse serum for 2 h. Sections were then stained with anti-MOMA2 (Cat.No.ab-33,451, abcam) or anti-α-SMA (AA132, Beyotime Biotech Inc) at 4 °C for overnight. The sections were then incubated with a secondary antibody against rat IgG (Thermo Scientific) for 1 h at room temperature. The sections were visualized with a DAB substrate kit (Vector Laboratories). Finally, image processing was carried out using a Leica DMRE microscope equipped with Spot digital image analysis software and camera.
Gut bacterial DNA extraction and sequencing
About each of 0.5 g cecal sample was used for DNA extraction with QIAamp Fast DNA Stool Mini kit (Qiagen) according to the manufacturer’s protocols. The extracted DNA was amplified with primers targeting V4-V5 hypervariable region of bacterial 16S rRNA genes (forward: 5-CCAGCAGCYGCGGTAAN-3; reverse: 5-CCGTCAATTCNTTTRAGT-3). PCR were then performed under the following condition: denaturation (95 °C, 3 mins), amplification (25 cycles), annealing (55 °C, 30 s), and elongation (72 °C, 30 s), and final elongation (72 °C, 5 mins). Sequencing was performed on the platform of Illumina HiSeq 2000. 16S rRNA sequence were explored by the package of Quantitative Insights Into Microbial Ecology (QIIME, 1.9.0) [26]. To avoid GC bias and low-quality nucleotides, the raw sequences were filtered with the following criteria: minimum Phred score, 20; minimum number of high-quality calls, 0.75; maximum number of consecutive low-quality base calls, 3. Trimmed sequences were then assembled [27].
Gut microbiota diversity and composition analysis
Sequence with 97% similarity were clustered into one operational taxonomic unit by UCLUST [28]. The closed-reference taxonomy was assigned to genus level against the Greengenes database [29] with 0.5 confidence threshold. Sequence occurred less than three times were removed from further analysis. Estimation of Shannon-Weiner index based on 16S rRNA gene were performed to assess the gut microbiota diversity with an average sequence number of 2,4000 reads per sample. Weighted uniFrac distance [30, 31] were used for the principle coordinate analysis.
Identification of gut microbiota features that discriminate groups
Linear Discriminant Analysis Effect Size (LEfSe) were performed to identify the bacterial communities that differentiate different groups of mice [32]. Gut bacterial richness was used to calculate the effect size on either dietary or apoE deficiency. The threshold on logarithmic Linear Discriminant Analysis (LDA) score were set as 2 (default). One-way ANOVA was used to analyze differences between groups and Tukey’s multiple comparison test was applied to correct for multiple tests.
Statistics
The TG, TC and HLD-C levels were represented as mean ± SEM. The richness of dominant gut bacteria was presented by average and relative abundance with more than 0.5% were considered as dominate bacterial communities. Statistical analyses were performed by one-way ANOVA followed by Student’s t-test. P < 0.05 was considered as statistics significant. Pearson’s correlation was calculated for each pairwise combination of each gut bacterial family versus atherosclerosis lesion area ratio using Graphpad Prism 6.
Results
Dyslipidemia and atherosclerosis in apoE KO mice
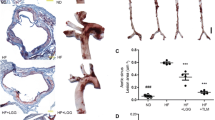
On a normal chow diet, apoE KO mice had significantly higher levels of plasma TC but lower levels of HDL-C than WT mice but TG levels were unchanged (Fig. 1a, b and c). On a Western diet, TC levels of WT mice were increased but no change in apoE KO mice compared to normal chow diet (Fig. 1a). The same trend was also found in HDL-C (Fig. 1c). In spite of this, there were no aortic lesions in WT mice on both a chow and Western diet but atherosclerosis was observed in apoE KO mice on both a chow and Western diet (Fig. 1d, e and f). Atherosclerotic lesions were significantly increased in Western diet-fed apoE KO mice compared with chow-fed apoE KO mice (Fig. 1d, e and f). Moreover, the macrophage and smooth muscle cells increased in high cholesterol feeding apoE KO mice compared to normal chow diet (Fig. 1e).

Western diet and apoE gene deletion elevate blood lipids and aggravate atherosclerosis. a Total cholesterol (TC); (b) Triglycerides (TG); (c) High-density lipoprotein cholesterol (HDL-C) levels at 12 weeks in the plasma samples from four groups of mice; (d) Cross sections of aortic roots from four groups of mice were stained with Oil-Red-O; (e) Macrophage and smooth muscle cells staining of apoE KO mice; (f) En face Oil-Red-O staining of aortic specimens with the quantitive analysis on the right panel. Data are expressed as the mean ± SEM, n = 7~ 8 for each group. ∗P < 005, ∗∗∗P < 0001 versus wild type (WT) mice
The gut microbiota alteration in apoE KO mice
To shed light on the gut microbiota distribution in mice, we sequenced V4-V5 amplicons of 16S rRNA genes. After quality control, we obtained a total of 2,766,806 sequences (69,170 ± 26,490 per sample) with the average length of 327 bp (Additional file 1: Figure S1). As expected, phylum-level composition was dominated by three phyla (Bacteroidetes, Firmicutes, and Proteobacteria), accounting for 84.7%~ 87.1% of the gut bacterial community (Additional file 2: Figure S2). Of these phyla, the most dominant phylum was Bacteroidetes with a richness of 36.4%~ 50.9%, followed by Firmicutes with 17.6%~ 30.6%, and Proteobacteria with 15.6~ 20.1% (Additional file 2: Figure S2). The other phyla such as Actinobacteria, Deferribacteres and Verrucomicrobia also were classified, but these bacteria were present at relatively very low abundance (less than 0.5%). Family-level composition was dominated by Lachnospiraceae, Porphyromonadaceae, Prevotellaceae, Ruminococcaceae, Geobacteraceae, Desulfovibrionaceae and Bacteroidaceae (Additional file 3: Figure S3). Estimation of Shannon-Weiner index, a measurement of the α-diversity of the gut microbiota, demonstrated that apoE KO mice had a tendency for decreasing the microbiota diversity when challenged with a Western diet (Fig. 2a) whereas it was unchanged in WT mice fed on a Western diet. Principal coordinate analysis based on weighted-unifrac distance showed a high degree of divergence between apoE KO and WT mice on the first vector, which accounts for 58.6% of the gut microbiota profile (Fig. 2b) although the gut microbiota change induced by different diets was indistinguishable (Fig. 2b). On normal chow diet, apoE KO mice showed significant variance of two phyla (Firmicutes and Bacteroidetes) and six families (Lachnospiraceae, Ruminococcaceae, Desulfovibrionaceae, Helicobacteraceae, Erysipelotrichaceae and Veillonellaceae) (Fig. 3) compared to WT mice. Western diet feeding led to the increase of Firmicutes and Erysipelotrichaceae in WT mice but Erysipelotrichaceae was not changed in apoE KO mice (Fig. 3).

The gut microbiota diversity and PCoA analysis of four groups of mice. a The comparison of Shannon-Weiner index, representing the microbiota diversity for four groups of mice; (b) Principal coordinate analysis (PCoA) for gut microbiota of four groups of mice

Comparison of gut microbiota among four groups of mice. T-test was applied to detect the statistical significance. Data are expressed as the mean ± SEM, n = 7~ 8 for each group. ∗P < 005 versus wild type (WT) mice
LEfSe and correlation analysis
The strict version of LEfSe (all against all) detected eight gut bacterial clades showing significant differences between aopE KO mice fed a Western diet and WT mice on a normal chow diet with LDA scores higher than two (Fig. 4a). The phylum of Firmicutes and Lachnospiraceae (from class to family) were found to be enriched in apoE KO mice on a Western diet, while Bifidobacteriales (from order to family), Negativicutes, Selenomonadales and Veillonellaceae were enriched in WT mice on a normal chow diet (Fig. 4a). In order to explore whether gut microbiota have any correlations with atherosclerosis in apoE KO mice, we performed a correlation analysis of family-level gut microbiota and aortic atherosclerosis lesion area. The relative abundances of Lachnospiraceae and Ruminococcaceae were found to be positively correlated with atherosclerosis lesion area (P < 0.05) (Fig. 4b and c).

Linear regression plots and LDA Effect Size (LEfSe) analysis. a Cladogram generated with Linear Discriminant Analysis (LAD) Effect Size (LEfSe) analysis illustrating significant shifts in the gut microbiota in the four groups of mice (n = 7~ 8). Size of yellow circles is proportionated to each taxon’s mean relative abundance; (b) Linear regression of Lachnospiracea correlated with relative atherosclerosis lesions; (c) Linear regression of Ruminococcaceae correlated with relative atherosclerosis lesions. P < 0.05 and Pearson’s correlation coefficient (r) > 0.6
Discussion
In the current study, we analyzed the gut microbiota of apoE KO mice and compared with WT mice on either a normal chow or Western diet. It is well known that apoE KO mice can develop atherosclerosis even on a normal chow diet and moreover, a Western diet is very atherogenic and accelerates the development of atherosclerosis in apoE KO mice. In spite of this, it is not clear whether Western diet feeding influences the gut microbiota in apoE KO mice or whether there are any associations between the altered gut microbiota profiling with atherosclerosis. Our results showed that the gut microbiota diversity and composition were remarkably changed in apoE KO mice, especially on a Western diet. Firmicutes and Clostridia (from class to family) were found to be enriched in apoE KO mice on a Western diet by LEfSe analysis. It has been reported that the relative abundance of Firmicutes could lead to the formation of increased amounts of metabolic endotoxins like lipopolysaccharides, which was able to enter the blood steam and cause the chronic inflammation [33]. Three species, including Clostridium asparagiforme, Clostridium hathewayi, and Clostridium sporogenes in the family of Clostridiales, were also reported to be correlated with the accumulation of trimethylamine N-oxide (TMAO) [34, 35]. Atherosclerosis is considered as a chronic inflammatory process and previous studies have identified that the gut microbiota-dependent TMAO production directly contributed to the progression of atherosclerosis, indicating that gut microbiota participates in the development of atherosclerosis [24, 35,36,37]. TMAO was able to accelerate atherosclerosis by disrupting the cholesterol transport and modifying bile acids composition [38], enhancing thrombosis formation by activating platelets [39], and increasing high blood pressure induced heart failure by adverse cardiac remodeling [40]. In our study, Lachnospiraceae and Ruminococcaceae, the two most abundant families in the order of Clostridiales, were found to be correlated with the atherosclerosis size. Although it is not possible to ascertain whether such correlation is causal or casual, the further studies are required to investigate its significance in future. Nevertheless, it has also been reported that the members of Lachnospiraceae were able to cause diabetes mellitus in germ-free mice [41]. Therefore, the gut microbiota transplantation would be performed using germ-free mice to determine the causal relationship in the future.
Conclusions
Collectively, our study showed that there are marked changes in the gut microbiota of apoE KO mice, particularly challenged with a Western diet and these alterations may be possibly associated with atherosclerosis.
References
Tencerova M, Figeac F, Ditzel N, Taipaleenmaki H, Nielsen TK, Kassem M. High-fat diet-induced obesity promotes expansion of bone marrow adipose tissue and impairs skeletal stem cell functions in mice. J Bone Miner Res. 2018;33(6):1154–65.
Namekawa J, Takagi Y, Wakabayashi K, Nakamura Y, Watanabe A, Nagakubo D, et al. Effects of high-fat diet and fructose-rich diet on obesity, dyslipidemia and hyperglycemia in the wbn/kob-lepr(fa) rat, a new model of type 2 diabetes mellitus. J Vet Med Sci. 2017;79:988–91.
Ghosh SS, Righi S, Krieg R, Kang L, Carl D, Wang J, et al. High fat high cholesterol diet (western diet) aggravates atherosclerosis, hyperglycemia and renal failure in nephrectomized ldl receptor knockout mice: role of intestine derived lipopolysaccharide. PLoS One. 2015;10:e0141109.
Andoh A. Physiological role of gut microbiota for maintaining human health. Digestion. 2016;93:176–81.
Del Chierico F, Vernocchi P, Bonizzi L, Carsetti R, Castellazzi AM, Dallapiccola B, et al. Early-life gut microbiota under physiological and pathological conditions: the central role of combined meta-omics-based approaches. J Proteome. 2012;75:4580–7.
Noureldein MH, Eid AA. Gut microbiota and mtor signaling: insight on a new pathophysiological interaction. Microb Pathog. 2018;118:98–104.
Quigley EM, Stanton C, Murphy EF. The gut microbiota and the liver. Pathophysiological and clinical implications. J Hepatol. 2013;58:1020–7.
Breslow JL. Mouse models of atherosclerosis. Science. 1996;272:685–8.
Linton MF, Atkinson JB, Fazio S. Prevention of atherosclerosis in apolipoprotein e-deficient mice by bone marrow transplantation. Science. 1995;267:1034–7.
Zhang SH, Reddick RL, Piedrahita JA, Maeda N. Spontaneous hypercholesterolemia and arterial lesions in mice lacking apolipoprotein e. Science. 1992;258:468–71.
Wang Z, Zhang J, Li H, Li J, Niimi M, Ding G, et al. Hyperlipidemia-associated gene variations and expression patterns revealed by whole-genome and transcriptome sequencing of rabbit models. Sci Rep. 2016;6:26942.
Niimi M, Yang D, Kitajima S, Ning B, Wang C, Li S, et al. Apoe knockout rabbits: a novel model for the study of human hyperlipidemia. Atherosclerosis. 2016;245:187–93.
Zhu L, Zhang D, Zhu H, Zhu J, Weng S, Dong L, et al. Berberine treatment increases akkermansia in the gut and improves high-fat diet-induced atherosclerosis in apoe(−/−) mice. Atherosclerosis. 2017;268:117–26.
Xu Y, Xu S, Liu P, Koroleva M, Zhang S, Si S, et al. Suberanilohydroxamic acid as a pharmacological kruppel-like factor 2 activator that represses vascular inflammation and atherosclerosis. J Am Heart Assoc. 2017;6(12):e007134.
Hamoud S, Shekh Muhammad R, Abu-Saleh N, Hassan A, Zohar Y, Hayek T. Heparanase inhibition reduces glucose levels, blood pressure, and oxidative stress in apolipoprotein E knockout mice. BioMed Research International. 2017;2017:7357495. https://doi.org/10.1155/2017/7357495.
Kolovou G, Anagnostopoulou K, Mikhailidis DP, Cokkinos DV. Apolipoprotein e knockout models. Curr Pharm Des. 2008;14:338–51.
Burcelin R. Gut microbiota and immune crosstalk in metabolic disease. Mol Metab. 2016;5:771–81.
Conterno L, Fava F, Viola R, Tuohy KM. Obesity and the gut microbiota: does up-regulating colonic fermentation protect against obesity and metabolic disease? Genes Nutr. 2011;6:241–60.
Nagpal R, Kumar M, Yadav AK, Hemalatha R, Yadav H, Marotta F, et al. Gut microbiota in health and disease: an overview focused on metabolic inflammation. Benef Microbes. 2016;7:181–94.
Sanduzzi Zamparelli M, Compare D, Coccoli P, Rocco A, Nardone OM, Marrone G, et al. The metabolic role of gut microbiota in the development of nonalcoholic fatty liver disease and cardiovascular disease. Int J Mol Sci. 2016;17(8):1225.
Sanchez-Alcoholado L, Castellano-Castillo D, Jordan-Martinez L, Moreno-Indias I, Cardila-Cruz P, Elena D, et al. Role of gut microbiota on cardio-metabolic parameters and immunity in coronary artery disease patients with and without type-2 diabetes mellitus. Front Microbiol. 2017;8:1936.
Karlsson FH, Fak F, Nookaew I, Tremaroli V, Fagerberg B, Petranovic D, et al. Symptomatic atherosclerosis is associated with an altered gut metagenome. Nat Commun. 2012;3:1245.
Jie ZY, Xia HH, Zhong SL, Feng Q, Li SH, Liang SS, et al. The gut microbiome in atherosclerotic cardiovascular disease. Nat Commun. 2017;8:845.
Wang Z, Klipfell E, Bennett BJ, Koeth R, Levison BS, Dugar B, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 2011;472:57–63.
Bai L, Li Z, Li Q, Guan H, Zhao S, Liu R, et al. Mediator 1 is atherosclerosis protective by regulating macrophage polarization. Arterioscler Thromb Vasc Biol. 2017;37:1470–81.
Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. Qiime allows analysis of high-throughput community sequencing data. Nat Methods. 2010;7:335–6.
Eren AM, Vineis JH, Morrison HG, Sogin ML. A filtering method to generate high quality short reads using illumina paired-end technology. PLoS One. 2013;8:e66643.
Edgar RC. Search and clustering orders of magnitude faster than blast. Bioinformatics. 2010;26:2460–1.
McDonald D, Price MN, Goodrich J, Nawrocki EP, DeSantis TZ, Probst A, et al. An improved greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012;6:610–8.
Lozupone C, Knight R. Unifrac: a new phylogenetic method for comparing microbial communities. Appl Environ Microbiol. 2005;71:8228–35.
Chang Q, Luan Y, Sun F. Variance adjusted weighted unifrac: a powerful beta diversity measure for comparing communities based on phylogeny. BMC Bioinformatics. 2011;12:118.
Segata N, Izard J, Waldron L, Gevers D, Miropolsky L, Garrett WS, et al. Metagenomic biomarker discovery and explanation. Genome Biol. 2011;12:R60.
Poppleton DI, Duchateau M, Hourdel V, Matondo M, Flechsler J, Klingl A, et al. Outer membrane proteome of veillonella parvula: a diderm firmicute of the human microbiome. Front Microbiol. 2017;8:1215.
Romano KA, Vivas EI, Amador-Noguez D, Rey FE. Intestinal microbiota composition modulates choline bioavailability from diet and accumulation of the proatherogenic metabolite trimethylamine-n-oxide. MBio. 2015;6:e02481.
Tang WH, Wang Z, Levison BS, Koeth RA, Britt EB, Fu X, et al. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N Engl J Med. 2013;368:1575–84.
Geng J, Yang C, Wang B, Zhang X, Hu T, Gu Y, et al. Trimethylamine n-oxide promotes atherosclerosis via cd36-dependent mapk/jnk pathway. Biomed Pharmacother. 2017;97:941–7.
Senthong V, Li XS, Hudec T, Coughlin J, Wu Y, Levison B, et al. Plasma trimethylamine n-oxide, a gut microbe-generated phosphatidylcholine metabolite, is associated with atherosclerotic burden. J Am Coll Cardiol. 2016;67:2620–8.
Koeth RA, Wang Z, Levison BS, Buffa JA, Org E, Sheehy BT, et al. Intestinal microbiota metabolism of l-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med. 2013;19:576–85.
Zhu W, Gregory JC, Org E, Buffa JA, Gupta N, Wang Z, et al. Gut microbial metabolite tmao enhances platelet hyperreactivity and thrombosis risk. Cell. 2016;165:111–24.
Organ CL, Otsuka H, Bhushan S, Wang Z, Bradley J, Trivedi R, et al. Choline diet and its gut microbe-derived metabolite, trimethylamine n-oxide, exacerbate pressure overload-induced heart failure. Circ Heart Fail. 2016;9:e002314.
Kameyama K, Itoh K. Intestinal colonization by a lachnospiraceae bacterium contributes to the development of diabetes in obese mice. Microbes Environ. 2014;29:427–30.
Funding
This work was supported in part by the National Natural Science Foundation of China (No. 81270348, EL), the Natural Science Foundation of Shaanxi Province (2014FWPT017, EL), the China Postdoctoral Science Foundation (2014 M562425, RW).
Availability of data and materials
The data supporting the findings of this study are available from the corresponding author upon request.
Author information
Authors and Affiliations
Contributions
BL performed bioinformatics analysis and wrote the manuscript; YZ performed the experiments and wrote the manuscript. YA, WG, LB and YL performed the experiments; RW and SZ interpreted the data and revised the manuscript; EL and JF designed the experiments and read the manuscript. All authors have approved the final version to be published.
Corresponding author
Ethics declarations
Ethics approval
Animal care and experimental procedures were approved by the Animal Care and Use Committee of Xi’an Jiaotong University.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
Figure S1. Reads number distribution of each sample based on 16S rRNA gene sequencing. (JPG 262 kb)
Additional file 2:
Figure S2. Dominant gut microbiota compositions of samples at phyla level. (JPG 96 kb)
Additional file 3:
Figure S3. Dominant gut microbiota compositions of samples at family level. (JPG 144 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

{kind=link}
{kind=link}
{kind=link}
Cite this article
Liu, B., Zhang, Y., Wang, R. et al. Western diet feeding influences gut microbiota profiles in apoE knockout mice. Lipids Health Dis 17, 159 (2018). https://doi.org/10.1186/s12944-018-0811-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12944-018-0811-8