
Abstract
Chimeric antigen receptor (CAR) T-cell therapy has made remarkable progress in cancer immunotherapy, but several challenges with unclear mechanisms hinder its wide clinical application. Single-cell sequencing technologies, with the powerful unbiased analysis of cellular heterogeneity and molecular patterns at unprecedented resolution, have greatly advanced our understanding of immunology and oncology. In this review, we summarize the recent applications of single-cell sequencing technologies in CAR T-cell therapy, including the biological characteristics, the latest mechanisms of clinical response and adverse events, promising strategies that contribute to the development of CAR T-cell therapy and CAR target selection. Generally, we propose a multi-omics research mode to guide potential future research on CAR T-cell therapy.
Similar content being viewed by others
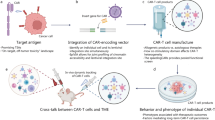

Introduction
Chimeric antigen receptor (CAR) T-cell therapy has changed the therapeutic landscape of cancer immunotherapy, especially for hematological malignancies, including B-cell acute lymphoblastic leukemia (B-ALL) [1], B-cell non-Hodgkin’s lymphoma (B-NHL) [2], chronic lymphocytic leukemia (CLL) [3], and Hodgkin’s lymphoma (HL) [4]. Although CAR T-cell therapy has impressive clinical outcomes, there are still a series of challenges, such as high cost, restricted clinical accessibility, resistance, relapse and adverse events [5, 6]. The widespread application of CAR T-cell therapy in hematological malignancies has led researchers to test its efficacy in solid tumors. [7,8,9,10,11]. However, compared with hematological diseases, CAR T-cells are less effective in solid tumors due to the inadequate tumor infiltration of CAR T-cells, the lack of stably expressed, tumor-specific antigens, the highly immunosuppressive tumor microenvironment (TME), etc. [12,13,14,15,16]. These problems are expected to be addressed by analyzing the molecular landscape and heterogeneity of CAR T-cells and their interactions with various cells in the microenvironment.
Flow cytometry has been the main metric used in CAR T-cell therapy to measure the immunophenotype and composition of circulating CAR T-cells [17, 18], but its low throughput and hypothesis-driven nature prevent unbiased exploratory screening and molecular profiling of CAR T-cells. Therefore, it is challenging to identify the key molecular drivers associated with the expansion and persistence properties of CAR T-cells that are directly related to treatment outcomes. These shortcomings in measuring clinically relevant features are being addressed with the development of single-cell RNA sequencing (scRNA-seq). Single-cell sequencing technologies enable unbiased, high-resolution, and high-throughput sequencing analysis to reveal cellular heterogeneity with unprecedented resolution and capture high levels of the molecular landscape. Functional analysis also helps to understand cellular transitions, interactions and communications of CAR T-cells [19,20,21,22,23]. This has provided new insights into the overall performance of CAR T-cells in relation to patient prognosis (Table 1). In this review, we discuss how single-cell sequencing technologies, especially scRNA-seq, decipher and advance CAR T-cells and facilitate CAR target selection. Finally, we propose a multi-omics research mode from a clinically translational perspective to help researchers better understand and explore the future directions of CAR T-cell therapy.
Basics of CAR T-cells
CAR T-cell product heterogeneity is affected by CAR structure [24,25,26], T cell subtype [27] and product manufacturing process [28], which influences the efficacy and safety of CAR T-cell therapy. Single-cell sequencing technologies can systematically evaluate the impact of the above factors on the final CAR T-cell product, guiding the rational design and optimization of CAR T-cell therapy. CAR is an engineered receptor that is composed of three main parts: the extracellular, transmembrane and intracellular domains (Fig. 1A). According to the differences in the design of intracellular domains and the adoption of cytokines and ligands, CAR molecules have been developed for five generations [29]. Different combinations of molecular modules of the CAR have different effects on the phenotype and function of CAR T-cells, such as the selection of the costimulatory domain [26] and the immunogenicity of the single-chain variable fragment (scFv) [24, 25]. For T cell subtypes, the CD4:CD8 ratio and the composition of different T cell subtypes affect the antitumor ability of CAR T-cell therapy. For instance, when the CD4:CD8 ratio is 1:1, the synergistic antitumor ability can have a better effect [30,31,32]. CAR T-cells with less-differentiated naive and early memory features are related to a higher rate of durable clinical remission [33, 34]. Exhausted T cells with higher expression of inhibitory immune checkpoint receptors are associated with poorer clinical outcomes [35]. In addition, the difference in CAR T-cell lineage clones based on the T cell receptor (TCR) is also an important factor leading to the heterogeneity of CAR T-cell products [36].

The basics of CAR T-cells. (A) Illustration of the basic structure of the conventional TCR-CD3 complex and CAR molecule. (B) Flow scheme of the CAR T-cell manufacturing process. Autologous CAR T-cell manufacturing typically begins with leukapheresis of the patient. Then, the T cells are activated and amplified with antibody-coated beads. After that, the CAR construct is introduced into the T cells, typically by viral or non-viral vectors. Finally, CAR T-cells are expanded to the required amount and then infused into the patient after quality control testing
During the manufacturing process, every step may contribute to the heterogeneity of the final CAR T-cell product [28]. The traditional manufacturing process of CAR T-cells begins with the collection of peripheral blood mononuclear cells (PBMCs) from patients undergoing leukapheresis. Next, T cells that have been further enriched from PBMCs are stimulated with anti-CD3/anti-CD28 monoclonal antibodies to induce T cell activation, and then lentiviral vectors, gamma-retroviral vectors or other delivery methods are used to engineer CAR molecules into T cells. Subsequently, these generated CAR T-cells are expanded in vitro to reach the required amount of cells for either experimental testing or clinical treatment [37, 38] (Fig. 1B). Moreover, T cells can also be extracted from healthy donors (HDs) to prepare allogeneic CAR T-cells [39].
Single-cell sequencing technologies for CAR T-cells
Single-cell sequencing technologies used in CAR T-cell research mainly include scRNA-seq, single-cell T-cell receptor sequencing (scTCR-seq), single-cell assay for transposase-accessible chromatin sequencing (scATAC-seq), cytometry by time-of-flight (CyTOF), cellular indexing of transcriptomes and epitopes by sequencing (CITE-seq) and single-cell multiplexed secretome proteomics.
scRNA-seq for CAR T-cell research
scRNA-seq is currently the most widely used single-cell sequencing technology. Every scRNA-seq experiment follows a similar basic strategy, including sample dissociation, single-cell capture, cell lysis, mRNA reverse transcription, cDNA amplification, library construction, high-throughput sequencing, and data analysis [40, 41] (Fig. 2A, B). The samples of CAR T-cell research mainly include CAR T-cell products, PBMCs, bone marrow (BM), cerebrospinal fluid (CSF) and tumor tissue. Adequate sample preparation is a prerequisite for generating reliable single-cell transcriptomics results. The general dissociation process includes tissue dissection, mechanical mincing, enzymatic/proteolytic extracellular matrix (ECM) breakdown and selective enrichment. Since CAR T-cell products and PBMCs are single-cell suspensions, the dissociation step is eliminated, avoiding the generation of stress genes in the process and retaining the proportion of various cell types, which is crucial for the reliability of single-cell sequencing data.

Single-cell sequencing technologies for CAR T-cell therapy. (A) Schematic overview of single-cell multi-omics analysis of CAR T-cell therapy. (B) scRNA-seq measures the transcriptome from the entire cell. (C) scTCR-seq measures the V(D)J sequence of TCR and gene expression profiles in tandem. (D) scATAC-seq identifies open regions of chromatin by inserting adapters with Tn5 transposase to map regions of transcription factor binding. (E) CyTOF analyzes a high-dimensional, multi-parametric quantification of protein detection with metal-isotype antibody, inductively coupled plasma ionization, and time-of-flight detector. (F) CITE-seq allows the simultaneous assessment of the transcriptome and surface or intracellular protein expression of individual cells immunostained with oligonucleotide-coupled antibodies. (G) SCBC measures multiple secreted proteins with arrays of microchambers that are decorated with ordered arrays of antibodies against target proteins
The droplet-based platform (10x Genomics BD) is the most commonly used scRNA-seq platform in the field of CAR T-cells. Using a microfluidic chamber, the droplet-based method can separate individual cells into an oil-based microdroplet. A single droplet contains gel microbeads with mRNA-capturing primers in conjunction with a unique molecular barcode alongside an enzyme/reagent mix, which is required for cell lysis and reverse transcription. Single cells in the droplets are lysed, followed by reverse transcription, cDNA amplification, and the generation of a barcoded sequencing library. Then, the samples are processed for sequencing. The obtained raw data require quality control, which is crucial to the subsequent exploration of biological questions from downstream analysis at the cell/gene level. Downstream analyses include simple quantification of gene expression to in-depth examination of cell heterogeneity, lineage transitions, cell-cell interactions, etc. Comprehensive tutorials of workflow and analysis of scRNA-seq have been established [40, 41].
Other single-cell sequencing technologies
scTCR-seq sequences the 5′-end transcript and can simultaneously detect the V(D)J sequence and transcript of a single cell (Fig. 2C). It can not only characterize the clonotypic diversity of the CAR T-cell population but also correlate T cell clones with their functional phenotypes, such as activation, memory, and exhaustion, providing information on the role of CAR T-cell clonotypic phenotypes in driving the immune response [42,43,44]. By inserting sequencing adapters into accessible regions of the genome with Tn5 transposase, scATAC-seq allows the measurement of genome-wide open chromatin regions to identify both activated genes and cis-elements (such as promoters and enhancers), as well as to postulate the TFs to which cis-elements are bound [45] (Fig. 2D). At present, scATAC-seq is mainly used to characterize the epigenetics of CAR T-cell differentiation and exhaustion [46, 47]. CyTOF, also known as mass cytometry, is a technology that combines flow cytometry with mass spectrometry for multi-parameter analysis. By conjugating more than 100 antibodies with unique isotopically pure metals to overcome the overlaps in fluorophore spectrum, CyTOF could detect intracellular and extracellular protein expression accurately at the single-cell level [48, 49] (Fig. 2E). As a platform for multiplexed data analysis, CITE-seq utilizes an oligonucleotide-barcoded antibody conjugated to the cell surface antigen that can achieve parallel characterization of cell populations on the basis of the expression of surface protein markers and the transcriptome landscape [50] (Fig. 2F). Cytokines are potent secreted regulators that play an important role in CAR T-cell function [51, 52]. The single-cell barcode chip (SCBC) is one of the most advanced single-cell proteomic devices for measuring secreted cytokines [53]. SCBC uses concentrated arrangements of microfabricated compartments (typically < 1 nL volume) with spatially barcoded capture sites (currently commercialized by Isoplexis) for highly multiplexed single-cell analysis of up to 42 cytokines related to CAR T-cell function, such as effector, stimulatory, regulatory and inflammatory molecules [54] (Fig. 2G). Since CAR T-cells exhibit significant heterogeneity in cytokine secretion, polyfunctional CAR T-cells and the polyfunctionality strength index (PSI) are used to describe CAR T-cell subsets capable of co-producing multiple cytokines at the single-cell level, which has been utilized to predict the clinical outcome of patients [55,56,57,58].
Deciphering and advancing CAR T-cell biology
CAR T-cell product heterogeneity
For CAR structures, the selection of the costimulatory domain is a critical factor to explore. Although CAR T-cells with CD28 or 4-1BB have similar clinical efficacy, they differ in kinetics and phenotype [26]. scRNA-seq showed that they had distinct transcriptional expression profiles, whether in the baseline or activated state, which indirectly reflected the different transcriptional regulatory mechanisms [59,60,61]. In addition to verifying that 4-1BB CAR T-cells have the gene encoding the memory phenotype and longer persistence compared with CD28 CAR T-cells [59, 60], multiple studies have observed that 4-1BB CAR T-cells express more MHC II genes [59,60,61]. This is presumably beneficial for co-application with tumor vaccines to increase antigen presentation to enhance epitope spread, but increases the risk of host-graft rejection of allogeneic “off-the-shelf” CAR T-cells [59].
For T cells, allogeneic CAR T-cells derived from HDs are an important means to expand the clinical accessibility of CAR T-cell therapy [62,63,64], but the intrinsic heterogeneity, function and safety of HD-derived allogeneic CAR T-cells have not been systematically evaluated. A study combined scRNA-seq and CITE-seq to characterize the differences between HD and patient-derived CAR T-cells in transcriptome, phenotype, and metabolic characteristics and found that HD-derived CAR T-cells were at a higher level of activation [61]. Moreover, the upregulation of MHC II genes indicated that HD-derived CAR T-cells may have stronger and faster antitumor efficacy but increased the risk of being cleared by the host immune system. HD-derived CAR T-cells were also associated with lower granulocyte-macrophage colony-stimulating factor (GM-CSF) expression than patient-derived CAR T-cells [61], which is a stimulator of CRS [65, 66], consistent with the clinically observed lower incidence of CRS in allogeneic CAR T-cells [67].
The difference in manufacturing processes is a crucial factor leading to CAR T-cell product heterogeneity. Previous studies have observed that the selection of fresh or cryopreserved PBMCs as primary material led to discrepancies in the efficacy of CAR T-cell products [68, 69]. A recent single-cell study suggested that this might be related to the number of Treg cells, which are notoriously intolerant of freezing [70]. In addition, the efficiency of viral transduction of CAR molecules influences CAR T-cell fitness and antitumor efficacy. Different profiles of CAR T-cells expressed by different CAR molecules (CARHigh, CARLow) at the bulk and single-cell levels confirm that CARHigh T-cells have stronger tonic signaling, activation and exhaustion [71]. Furthermore, characterizing gene regulatory networks found that CARHigh T-cells were regulated by the exhaustion-related regulators RFX5, NR4A1, and MAF [71]. Notably, cells with low or even negative expression of CAR express interferon-induced transmembrane (IFITM) 2 and IFITM3, which prevent viral vector entry and are probably potential drug targets to overcome the inefficiency of CAR transduction [72]. For vector bias to CAR T-cell function, scRNA-seq revealed that the transduction of vesicular stomatitis virus (VSV)-lentiviral vectors (LV) promoted the transition of CAR T-cells to a central memory phenotype, while the transduction of CD8-LV promoted the transition of CAR T-cells to a cytotoxic phenotype [72].
Antigen-specific stimulation of CAR T-cells
The activation mechanism of CAR T-cells is quite different from that of innate T cells. Compared with TCR-induced activation, the full molecular landscape of downstream signaling in CAR-induced activation remains elusive, which is partly due to the heterogeneity of CAR T-cell products. On the one hand, different CAR T-cell phenotypes differ in their ability to respond to antigens. On the other hand, not all cells in CAR T-cell products harbor CAR expression, which easily confuses the analysis of the cells upon antigen encounter. Several studies have used scRNA-seq and other single-cell sequencing technologies to resolve the heterogeneity of CAR T-cell products under different conditions of unstimulated, CAR-induced stimulated and TCR-induced stimulated CAR T-cells [59, 61, 73,74,75]. Due to the presence of ligand-independent tonic signaling, CAR T-cells in the unstimulated state are regulated by a mixture of early activation, exhaustion signatures, and cytotoxic activities [61]. After CAR-induced activation, CAR T-cells present highly mixed TH1/TH2 cell signaling. The levels of cytokines, such as IFN-γ, TNF-α, GM-CSF, IL-5 and IL-13, show great heterogeneity among different cell subsets [75]. Since GM-CSF is highly expressed in many CAR T-cells, GM-CSF+ CAR T-cells can be regarded as in a functionally active state, which is different from conventional T cells [61, 75]. Notably, CD4+ and CD8+ CAR T-cells showed high expression of cytotoxic cytokines, indicating that both cell types had killing functions [75]. Moreover, some CAR T-cell subsets upregulated the expression of the immune checkpoint genes CTLA4 and PD-1 and the immunosuppressive cytokine genes IL-10 and TGFB1 and downregulated the costimulator genes inducible costimulatory (ICOS) and OX40, which may be a mechanism to maintain immune homeostasis after activation [75]. In addition, some studies have also observed that some activated CAR T-cells did not change the transcriptomic profiles, and a few CAR T-cells showed signs of exhaustion in the early stages after activation [74, 76]. The relevant mechanism is still unclear, and it may be attributed to the CAR T-cell manufacturing process, tonic signaling, or cell-source specificity [74]. The gene expression was different between CAR T-cells stimulated by TCR and CAR-specific stimulation, with the former more specifically enriched in T cell activation genes (e.g., IFN-γ, IL-3, and CCL4) [59, 73].
The dynamic performance of CAR T-cells in vivo
CAR T-cells after infusion will experience rapid expansion as well as differentiation and exhibit long-term persistence of atypical patterns in metabolism and clearance [77,78,79]. The dynamic performance of CAR T-cells varied in both initial response and long-term remission, reflecting the function of CAR T-cells and the interaction between CAR T-cells and the host. Heterogeneity of CAR T-cell products affects the differentiation of CAR T-cells to different cellular dynamics and cell fates. A study utilized scRNA-seq and scTCR-seq and revealed that early CAR T-cell proliferation in tisa-cel responders was characterized by expansion of memory-like CD8+ CAR T-cell clones that differentiated into IL7R+ effector memory CAR T-cells, while axi-cel responders exhibited more heterogeneous populations. Among them, CD8+ CAR T-cells had stronger upregulation of activation marker PDCD1 and the immune checkpoint regulator SLAMF6 [70]. However, even if the same CAR T-cell product was activated by the same antigen, different CAR T-cell subpopulations led to different patterns of expansion and displayed divergent differentiation trajectories [36, 76]. However, clusters with high expression of cytotoxicity and proliferation genes usually predominated the post-infusion CAR T-cell functional groups [36, 76]. In addition, a study using scRNA-seq and scTCR-seq found an effector precursor CD8+ CAR T-cell with a unique transcriptional profile TIGIT+CD27−CD62Low in the initial infusion sample [76]. It was subsequently the main source of the majority of CAR T-cells with an effector phenotype in patients [76]. As tumor cells were cleared, most CAR T-cells at the remission phase further developed into long-lived memory cells and stayed in the “resting primed” state with minimal energy consumption to prevent relapse [36, 80]. These processes were conserved in the evolution of CAR T-cells targeting different antigens in different hematological malignancies [36, 76, 80]. Of note, in a recent study, two patients with CLL who achieved complete remission (CR) for up to 10 years had an initial response dominated by cytotoxic CD8+ CAR T-cells in their peripheral blood, followed by a long-term remission stage dominated by cytotoxic and proliferative Ki67hiCD4+ CAR T-cells [81] (Fig. 3A). The CD4+ CAR T-cells displayed a non-classical memory phenotype and a state of ongoing activation and proliferation. Meanwhile, they expressed cytotoxic genes, such as GZMA, GZMK and PRF1, as well as genes related to oxidative phosphorylation pathways. In vitro culture showed that the long-persisting CD4+ CAR T-cells were capable of killing CD19-expressing target cells directly. Nevertheless, in other studies, almost no CD4+ CAR T-cells were observed in patients in remission, and cytotoxic effects were absent [80]. Thus, the importance of CD4+ CAR T-cells in long-term disease control should be considered in conjunction with patient-specific characteristics, and the universality of the gene expression profiles needs to be further validated and explored in large-scale clinical cohort.

The dynamic biology of CAR T-cells in the peripheral blood and tumor microenvironment. (A) Schematic diagram of CAR T-cell dynamics in different phases in two CLL patients with clinical remission for more than 10 years after the infusion of CAR T-cells. (Left) The adjacent stacked bar plots exhibit the proportion of each CAR T-cell population at different time points. (Right) In the initial stage, CD4+ CAR T-cells provide support to CD8+ CAR T-cell responses via IL-2. In the long-term remission phase, the expression of the GZMA and GZMK genes was strongly upregulated, while GZMB was not highly expressed. The persistence of Ki67hiCD4+ CAR T-cells may partly be driven by healthy B cells. (B) Schematic diagram of cellular changes in bone marrow samples at different times in the TME (classified as pre-therapy, 28 days (D28) or 3 months (3M) following CAR T-cell treatment) of patients with short PFS (< 6 months) or long PFS (> 6 months). (Left) The adjacent stacked bar plots show the proportion of nontumor cells at different times. (Right) For patients with short PFS, there were terminal differentiation markers in bone marrow T cells at day 0 (D0). Monocytes/macrophages enriched with myeloid cells expressing BAFF and PD-L1 and CAR T-cells showed a more differentiated/effector phenotype at D28. At 3M, myeloma cells were increased and similar to the baseline phenotype. For patients with long PFS, the diversity of TCR was higher at D0 and increased at D28, accompanied by CAR T-cells with a more naive phenotype and enrichment of dendritic cells. CD8+ T cells at 3M had higher expression levels of genes associated with human bone marrow residence/retention, such as CXCR4 and CD69, and the myeloma cell phenotype was different from baseline
Cellular interactions with CAR T-cells
The function of CAR T-cells is closely associated with endogenous immune cells in the circulation system and tumor cells, immune cells and stromal cells in the TME. The complex cellular interactions involved in the function of CAR T-cells are critical to understanding the mechanism of action of CAR T-cells [82]. A study performed scRNA-seq on CAR T-cell products and PBMCs from plasma cell leukemia (PCL) patients after receiving CAR T-cell treatment. The ligand-receptor analysis showed the extensive interaction between proliferating CAR T-cells and cytotoxic CAR T-cells, as well as between CAR T-cells and endogenous T cells, and the gene ontology (GO) analysis found it associated with T cell activation, cell-cell adhesion, and TNF-related pathways, which indicated that CAR T-cells may establish a new immune environment by recruiting endogenous T cells [80]. Hence, CAR T-cells act as not only killers but also regulators. In fact, CAR T-cells can also reshape the TME and activate both innate and adaptive immunity to produce synergistic antitumor immunity by releasing IFN-γ [83, 84]. It was observed in two studies that IFN-γ derived from CAR T-cells promoted a more activated and less suppressive TME in two different animal models (B-cell lymphoma and glioblastoma), with concurrent activation and an increase of T lymphocytes and natural killer (NK) cells and the upregulation of myeloid cells expressing more antigen processing and presentation-related genes. Subsequent functional experiments confirmed that a lack of IFN-γ impaired the activation of host immune cells, which then affected the killing efficacy of CAR T-cells in vivo [85, 86].
CAR T-cells in continuous contact with tumor cells could easily lead to CAR T-cell exhaustion. A study performed bulk and single-cell sequencing of CAR T-cells continuously co-cultured with tumor cells in vitro (0, 20 and 28 days) and found that exhausted CD8+ CAR T-cells changed to a NK-like phenotype at the transcriptional, epigenetic and protein levels [87]. Meanwhile, gene expression levels of exhaustion markers and the NK signature increased under chronic antigen stimulation. Moreover, the transcriptional regulators ID3 and SOX4 were specifically expressed in exhausted NK-like CAR T-cell clusters, and their knockdown restored CAR T-cell potency. However, it is noteworthy that NK-like CAR T-cell clusters are not pre-existing but rather that CD8+ CAR T-cells acquire NK receptors via plasticity during prolonged antigen exposure. The causality of the transition and CAR T-cell dysfunction remains uncertain. Another study delineated the landscape for comprehensive and dynamic chromatin accessibility of CAR T-cells derived from an in vitro tumor cell co-culture system (0, 6 and 48 hours) via scATAC-seq [47]. Two subsets of exhausted CAR T-cells were defined. One is intermediate exhausted CAR T-cells with enriched motifs of transcription factors such as JUN, FOS, NFKB1, and BACH2, and the other is terminal exhausted CAR T-cells with enriched motifs of BATF, IRF4, and PRDM1. Similar changes in chromatin accessibility and transcriptional regulation patterns of exhausted CAR T-cells were also observed in patients. An in vitro experiment confirmed that CAR T-cells with reduced BATF and IRF4 expression exhibited better persistence and enhanced killing ability.
Deciphering and advancing the efficacy and safety of CAR T-cell therapy
Primary resistance
Primary resistance refers to the inability to induce CR after CAR T-cell infusion. A study performed a whole-transcriptome scRNA-seq analysis of CD19 CAR T-cell products from 24 large B-cell lymphoma (LBCL) patients and found that exhausted CD4+ and CD8+ T cells were enriched within infusion products (IPs) of patients with partial response/progressive disease (PR/PD) 3 months after infusion. Moreover, the lymphocyte activation 3 (LAG-3) and TIGIT genes and the basic leucine zipper ATF-like transcription factor (BATF), inhibitor of DNA binding 2 (ID2) and other failure-related transcription factors were highly expressed. Nevertheless, patients achieving CR had more memory CD8+ T cells [88]. Other scRNA-seq data obtained from relapsed/refractory B-cell lymphoma patients before and after CAR T-cell infusion suggested that the upregulation of the exhaustion marker TIGIT in CAR T-cells was highly correlated with a lower clinical response. Moreover, subsequent experiments both in vitro and in vivo confirmed that blocking TIGIT could delay tumor progression and restore the antitumor function of CAR T-cells. In the future, immune checkpoint inhibitors against TIGIT may be the key to improving the clinical response in patients [89]. In addition, CAR Treg cells are also involved in primary resistance [70, 90]. Another scRNA-seq study deciphered IPs and PBMCs derived from LBCL patients before and after CAR T-cell infusion and found that the increasing number of CAR Treg cells in IPs was positively correlated with the nonresponse rate of patients. In vitro and in vivo models showed that CAR Treg cells inhibited the antitumor activity of CAR T-cells and led to tumor relapse in mouse models [70]. Similarly, a study used CyTOF to examine PBMCs from LBCL patients after CAR T-cell infusion and found that higher frequencies of cytotoxic CD4+ and CD8+ subsets of CD57+ T-BET+ CAR T-cells were associated with patients achieving CR, whereas higher numbers of CD4+ HELIOS+ CAR T-cells with a Treg cell phenotype were associated with disease progression. Furthermore, a logistic regression model that combined the percentage of CAR Treg cells and the level of lactate dehydrogenase (LDH) was a powerful predictor of durable complete response versus progression [90] (Fig. 4A).

Mechanisms of resistance and toxicities. (A) Genes encoding immune checkpoints, such as lymphocyte activation 3 (LAG3) and T cell immunoglobulin and ITIM domain (TIGIT), as well as exhaustion-associated transcription factors, including basic leucine zipper ATF-like transcription factor (BATF) and inhibitor of DNA binding 2 (ID2), were highly expressed in infusion products. Among them, TIGIT has been used as a potential target with suggested beneficial efficacy in both in vitro and in vivo experiments. In addition, the number of CAR Treg cells also increased significantly in infusion products. (B) The TH2 pathway was absent, and early memory-like T cell subsets (TSCM and TCM) were decreased in infusion products. (C) scRNAseq profiling revealed that CD19-negative leukemic cells were present before CAR T-cell therapy. CD34+CD22+CD19− progenitor cells are B-ALL cells with negative CD19 expression that carry oncogenic lesions and lead to leukemia. In addition, regulatory programs of B-cell activation and germinal center reaction occurring in B-ALL cells can reduce CD19 expression. (D) IL-6, recognized as pivotal for CRS pathogenesis, is specifically and highly expressed by monocytes. (E) Targeting CD19+ mural cells, critical for BBB integrity, may lead to ICANS of CAR T-cell therapy. Reactivation of HHV-6+ CAR T-cells in the infusion product may cause ICANS. IACs, more IL-17 A polyfunctional T cells, and fewer CD4+CD57–Helios+ CAR T-cells in the infusion product are associated with high-grade ICANS. (F) Integrative analyses of single-cell sequencing datasets promote a deeper understanding of CAR antigen expression and identification of safer and more effective targets
Tumor cells can also induce primary resistance. Unbiased genome-wide loss-of-function screens in B-ALL and B lymphoma cell lines have revealed that tumor cells with inherent dysregulation of death receptor signaling could be resistant to CAR T-cell killing [91, 92]. Meanwhile, reduced expression of death receptor genes (FADD, BID, CASP8, and TNFRSF10B) was associated with patients with no response [91]. scRNA-seq results confirmed that the CAR T-cells from these patients expressed much higher levels of exhaustion markers [91].
Relapse
Relapse, also called acquired resistance, refers to the phenomenon in which patients who initially respond to CAR T-cell therapy experience disease recurrence over time. The TME plays a considerable role in promoting tumor relapse because of the expansion of the failure phenotype and immunosuppression-related cells [93,94,95]. A study leveraged multi-level single-cell sequencing technologies to analyze BM samples from multiple myeloma (MM) patients after B-cell maturation antigen (BCMA) CAR T-cell treatment and analyzed its association with the length of progression-free survival (PFS) in the responders. They found that patients with a long PFS were usually associated with the increased appearance of CLEC9A+ dendritic cells, CD27+TCF1+ T cells and T cells expressing marrow-residence genes, while the expansion of KLRG+HLA-DR+ terminally differentiated T cells and the appearance of the immunosuppressed BAFF+PD-L1+ myeloid cells were related to a short PFS. In addition, the residual myeloma cells tended to have less-differentiated and stem-like characteristics, with upregulated epithelial-mesenchymal transition (EMT) genes, which may be related to tumor relapse [96] (Fig. 3B). Another scRNA-seq study analyzed tumor tissue from glioblastoma (GBM) patients before and after CAR T-cell therapy and revealed that phenotype remodeling occurred in recurrent GBM after CAR T-cell therapy with a transition from a mesenchymal-like (MES) phenotype to a mixed MES-like, neural progenitor-like (NPC) and astrocyte-like (AC) phenotype, suggesting that the coexistence of cells characterized by different GBM subtypes undermined the efficacy of CAR T-cell therapy [97]. Similarly, a scRNAseq study analyzed PBMC and BM samples before and after CAR T-cell treatment in patients with mantle cell lymphoma (MCL) and discovered that the number of endogenous cytotoxic lymphocytes (CTLs) was significantly decreased after relapse, while the expression of the immune checkpoint TIGIT was high, which impaired their killing ability. TIGIT was also highly expressed on MCL cells. Cell-cell interaction showed that the interaction between TIGIT on MCL cells and its ligand CD155 on CD16+ monocytes significantly increased after relapse, which might attenuate the ability of myeloid cells to present tumor antigens and lead to weaker antitumor immune surveillance. In conclusion, the acquired expression of TIGIT in CTLs and MCL cells may be a central mechanism resulting in relapse in MCL patients [98]. In addition, further studies have focused on the distinct mechanisms of relapse by investigating positive and negative relapse.
Positive relapse
Positive relapse refers to tumor relapse with targeted tumor antigen expression, generally caused by limited activation and expansion, as well as poor antitumor potency and persistence of CAR T-cells in vivo [93, 94]. Single-cell sequencing technologies have not only validated the previously identified mechanisms but also helped to discover some entirely new mechanisms of positive relapse (Fig. 4B). Bai et al. [73] analyzed the IPs from some ALL patients with CR and others undergoing positive relapse via scRNA-seq and found that a deficiency of TH2 function in CAR T-cells was associated with positive relapse patients. Cytokine module analysis indicated that TH2 cytokine modules were enriched in CR patients and absent in positive relapse patients. Differential expression analysis also showed that except for the TH2 pathway and TH2-related genes, including IL-4, IL-5, IL-13 and the upstream regulator GATA3, there was no significant difference in other immune programs, such as effector, activation, inflammation and chemokine modules, between CR and positive relapse patients. Therefore, the maintenance of TH2 function may be indispensable for the long-term remission of patients receiving CAR T-cell therapy. In addition to immune function, CITE-seq analysis revealed that differentiated subsets of T cells could also make a difference. Early memory-like T cell subsets TSCM and TCM were significantly decreased in both resting and activated CAR T-cells from positive relapse patients. Furthermore, a prognostic model including three biological indicators, CAR+TH2+ frequency, TCM frequency, and TEM + TEF frequency, endowed good discrimination of patients with positive relapse and long-term remission, which may act as a biomarker for the prediction of risk and clinical response of patients.
Negative relapse
Negative relapse refers to tumor relapse without targeted tumor antigen expression [93]. The recognized mechanisms include the selection of pre-existing antigen-negative tumor cells, mutation, splicing variation, lineage switching-mediated target antigen loss and other factors affecting the presentation and expression of target antigens [99]. Several single-cell sequencing studies have validated the natural selection theory of the presence of antigen-negative tumor subclones before CAR T-cell treatment [100, 101] and unveiled a new mechanism for mediating negative relapse [102] (Fig. 4C). A study performed scRNA-seq on CD10posCD19pos cells and CD10posCD19neg cells of BM from patients with B-ALL before and after CAR T-cell treatment [101] and found that the gene expression profiles of the CD19neg B-ALL cells detected before CAR T-cell treatment were similar to those of other B-ALL cells both before and after CAR T-cell treatment but were significantly different from those of other CD19neg cells, such as NK cells and myeloid cells. Moreover, CaSpER and Ballele frequency (BAF) analysis showed that the potential pre-existing CD19neg B-ALL cells harbored genomic deletions in the same chromosome location as the other B-ALL cells, further confirming the presence of true B-ALL cells that did not express CD19 before CAR T-cell treatment [101]. Another study further characterized antigen-negative expressing cells. The researcher applied bulk RNA-seq, scRNA-seq and flow cytometry to describe the gene expression of CD34+ hematopoietic stem cell progenitor (HSCP) development across healthy fetal, neonatal, and postnatal samples and found that CD22 was expressed before CD19, which is highly expressed in PreProB and ProB progenitors. CD34+CD19−CD22+ early/immature progenitors were prevalent in patients with relapsed B-ALL and increased the probability of relapse in patients receiving CD19-targeted therapy. Fluorescence in situ hybridization (FISH) showed that CD34+CD19−CD22+ early/immature progenitors in B-ALL patients carried an oncogene lesion, and subsequent animal testing demonstrated their ability to initiate leukemogenesis [100]. Since the initial number of antigen-negative expressing cells is quite small, it is usually difficult to detect by flow cytometry [101]. However, scRNA-seq can unambiguously detect these scarce antigen-negative subclones and thus can serve as an effective strategy for CAR T-cell selection before cellular product infusion. Regarding novel mechanisms leading to negative relapse, Im et al. used live cell imaging and CD19-specific antibody fluorescence and found that when CD19 CAR T-cells interacted with B-ALL cells, the CD19 surface protein of surviving B-ALL cells accumulated and internalized into cells and subsequently downregulated CD19 expression on the surface. This process is similar to the activation of normal B cells [103]. scRNA-seq and scATAC-seq of surviving leukemic cells co-cultured with CAR T-cells showed that the leukemic cell subsets with the lowest CD19 expression were significantly enriched for gene expression and regulators correlated with B-cell activation signatures and germinal center reaction. Bruton’s tyrosine kinase (BTK) inhibitors can inhibit this process and enhance CAR T-cell killing ability [102].
Adverse events
CRS and ICANS
Cytokine release syndrome (CRS) is the most common and severe toxicity of CAR T-cell therapy [104,105,106,107,108]. CRS is generally considered to be a systemic inflammatory response induced by endothelial cell dysfunction, abnormal activation of macrophages, and the release of supraphysiological levels of various proinflammatory cytokines [106,107,108,109]. Although IL-6 is recognized as critical for CRS [104,105,106,107,108], the main cellular source releasing it remains unknown. An in vitro cytokine release assay of cultured CAR T-cells found that IL-6 secretion was closely related to monocytes, and the presence of IL-6 in monocytes was confirmed by intracellular staining results. scRNA-seq performed on CD45+ leukocytes isolated from humanized mice that developed CRS after CAR T-cell infusion confirmed that monocytes were the only cell population that consistently and specifically expressed high levels of IL-6. Overall, circulating monocytes, but not CAR T-cells, were major sources of IL-6 during CRS [108] (Fig. 4D). Moreover, single-cell cytokine profiling revealed that higher product PSI was associated with grade ≥ 3 CRS [58]. Compared to patients with grade 0-1 CRS, patients with grade ≥ 2 CRS had a higher PSI of CD4+ CAR T-cells, especially PSI of IL-8 and MCP-1 [57], which are involved in the recruitment of neutrophils and monocytes/macrophages [110, 111]. Moreover, PSI combined with CAR T-cell expansion or pretreatment serum IL-15 levels was more indicative of severe CRS [58].
Immune effector cell-associated neurotoxicity syndrome (ICANS) is another common toxicity related to CAR T-cell therapy. Activation of endothelial cells in the brain during inflammation and the destruction of the integrity of the blood brain barrier (BBB) may play a key role in the occurrence and development of ICANS [104, 105, 112]. Single-cell sequencing technologies elucidated the pathogenesis of ICANS (Fig. 4E). By analyzing human brain scRNA-seq data, Parker et al. discovered a rare population of cells that co-expressed CD19 and CD248, the cell marker of mural cells. Specific expression of CD19 in mural cells was confirmed by exclusion of B-cell confounding and immunohistochemistry of several regions of the human brain. This described a possible mechanism for ICANS, namely, CD19 CAR T-cells targeting mural cells, thus increasing BBB permeability and causing circulating inflammatory cytokines and CAR T-cells to enter the central nervous system (CNS) [113]. These findings may caution the use of intrathecal administration of CD19 CAR T-cells for primary CNS lymphoma. Another scRNA-seq study found that reactivated human herpesvirus 6 (HHV-6) carried by CAR T-cells may enter the CNS through OX40 receptors on BBB endothelial cells, resulting in the development of HHV-6 encephalitis, which has similar symptoms and requires differential diagnosis from ICANS [114]. Moreover, single-cell sequencing technologies could also identify some biomarkers from CAR T-cell products that predict an increased risk of severe ICANS, such as ICANS-associated cells (IACs) with a monocyte-like transcriptional signature [88], an increased number of polyfunctional CAR T-cells producing IL-17A [58], and low levels of CD4+Helios+ CAR T-cells [90].
On-target, off-tumor effects
On-target, off-tumor effects refer to CAR T cell-mediated recognition and lysis of non-malignant tissues expressing the target antigens because the antigens recognized by CAR T-cells are mostly tumor-associated antigens (TAAs) expressed in both normal and malignant cells [115]. The development of single-cell sequencing technologies has improved the resolution of target antigen analysis, providing valuable guidance on on-target, off-tumor effects evaluation (Fig. 4F). By integrating publicly available scRNA-seq databases, the antigen expression profiles of CAR targets were delineated in normal as well as malignant tissues, highlighting the importance of identifying differential expression levels of target antigens in patients before CAR T-cell therapy [116]. To further search for potential off-target CAR antigens, Jing et al. analyzed two single-cell databases, the human cell landscape (HCL) and adult human cell atlas (AHCA), and defined the CAR targets measurable in more than 100 nonimmune cells and more than 2% of the total cells in at least three normal tissues as a potentially risky gene (PRG) [117]. By employing this criterion, they identified PRGs neglected by bulk expression analysis, such as EGFR, PSCA and KDR (VEGFR2), and suggested that close surveillance was needed for early signs of off-target toxicity [117]. Of note, Parker et al. discovered that even 0.15% (12/7906) CD19-measurable human brain mural cells could lead to ICANS [113], but whether such a low level of expression had toxic impact on other tissues remained unknown. Moreover, different physiological and disease states may also affect the expression of target antigens to some extent. Therefore, a comprehensive atlas of human antigen expression is vital for understanding the on-target, off-target effects, and follow-up experiments may be needed to confirm the off-target toxicity.
Deciphering and advancing strategies for CAR T-cell therapy
Combination therapy
Combining CAR T-cells with other therapies, including chemotherapy, radiotherapy, hematopoietic stem cell transplantation, and other immunotherapies, is regarded as a promising strategy to overcome challenges and enhance the effectiveness of CAR T-cell therapy [118, 119]. In a study, cyclophosphamide (Cy) and oxaliplatin (Ox) were used innovatively to replace the conventional chemotherapy regimen Cy and fludarabine (Flu) prior to CAR T-cell infusion [120], which has been shown to promote T cell infiltration into tumors [121]. scRNA-seq profiles of murine lung tumors treated with Ox/Cy and CAR T-cells revealed the expression of T-cell-recruiting chemokine genes, including CXCL16 and CCL5, in multiple cell types in the TME, such as macrophages and DCs, which facilitated early infiltration of CAR T-cells into tumors partially through CXCR6 and CCR5. Subsequently, IFN-γ produced by tumor-infiltrating CAR T-cells led to the recruitment and activation of iNOS+ tumor macrophages. As the main source of chemokines, iNOS+ macrophages upregulate the expression of the CXCR3 ligands CXCL9 and CXCL10, binding to CAR T-cells expressing CXCR3 to facilitate their further infiltration and therefore initiating a positive feedback loop supporting CAR T-cell recruitment to tumors [120]. A patient receiving this combination regimen also achieved improved clinical responses [120]. Moreover, stimulator of IFN genes (STING) pathway activation enhanced T cell recruitment and effector cell function in tumors [122]. Another scRNA-seq study combined the STING agonists DMXAA/cGAMP with CAR T-cells to treat mice with breast cancer. The results demonstrated that compared to CAR T-cell infusion alone, DMXAA/cGAMP could regulate the inhibitory TME by increasing proinflammatory myeloid cells, reducing myeloid-derived suppressive cells and increasing the expression of chemokines that facilitated CAR T-cell recruitment and persistence at the tumor site. In addition, it could also promote the conversion of CAR T-cells to the proinflammatory phenotype to improve antitumor effects [123]. Furthermore, cyclin-dependent kinase 4 and 6 inhibitors (CDK4/6i) are widely used in cancer therapy [124]. In addition to inhibiting tumor proliferation by blocking the cell cycle [125], they were also found to promote the long-term antitumor effects of endogenous T cells through immunomodulatory effects [126]. A study combining scRNA-seq and CITE-seq found that T cells in the spleen of mice pretreated with CDK4/6i upregulated effector and memory-related genes, suggesting that CDK4/6i might enhance the cytotoxic effects of T cells and promote the differentiation of memory subsets that maintain long-term antitumor immunity. The combination of CDK4/6i and CAR T-cells in ovarian cancer mice significantly improved the effectiveness and persistence of tumor control [126].
Engineered CAR T-cells
Designing innovative engineered CAR T-cells to optimize CAR T-cell therapy is an important area to be explored [127,128,129]. Single-cell sequencing technologies can help to systematically evaluate the features of engineered CAR T-cells. A study using genome-wide clustered regularly interspaced short palindromic repeats (CRISPR) screening discovered that Ikaros family zinc finger protein 2 (IKZF2) and transducin-like enhancer of split 4 (TLE4) may be linked with the functional suppression and exhaustion of T cells, thus affecting the killing effects of CAR T-cells on GBM stem cells. scRNA-seq performed on IKZF2-KO (knockout) and TLE4-KO CAR T-cells revealed that these cells displayed enhanced cytotoxicity and immune stimulation transcriptional signatures while prohibiting exhaustion, indicating their superior effector function against tumor cells [130]. In addition, two pilot studies used CRISPR-Cas9 gene editing technology for CAR gene transduction instead of traditional viral transduction [131, 132]. scRNA-seq found that CAR T-cells integrating the CAR gene at the T-cell receptor α constant (TRAC) locus and the PD1 gene locus both expressed a memory-like phenotype and less exhaustion-associated transcriptional signatures, which was related to higher potency [131, 132]. Obviously, the site-directed integration of CAR genes by CRISPR may be superior to random insertion by viral vector transduction.
In addition, many studies have attempted to modify engineered CAR T-cells to induce endogenous antitumor immunity and thus reshape the TME. One study designed a novel CAR T-cell (RN7SL1 CAR T-cell) that secreted non-coding RNA RN7SL1 in the form of extracellular vesicles (EVs), which would be preferentially taken up by immune cells to enhance endogenous antitumor immunity. scRNA-seq performed on tumor tissues of mice infused with RN7SL1 CAR T-cells showed that RN7SL1 could reduce suppressive myeloid cell subsets, increase inflammatory DCs expressing costimulatory genes, and activate amplification of effector-memory endogenous CD8+ T cells. Supported by these features, RN7SL1 CAR T-cells could perform an effective killing function even in a poorly immunogenic tumor [133]. Similarly, scRNA-seq suggested that CAR T-cells overexpressing superkine IL-2 (Super2) and IL-33 (Super2+ IL-33 CAR T-cells) could induce the conversion of M2-like macrophages to M1-like macrophages that highly express antigen presentation genes in TME, upregulate the ratio between CD8+ effector T cells and immunosuppressive Tregs, recruit and activate endogenous innate and adaptive immune cells, including tumor-specific T cells. The increased antitumor efficacy of Super2+ IL-33 CAR T-cells was observed in a variety of animal models [134].
Locoregional delivery of CAR T-cells
CAR T-cells are usually administered intravenously to treat hematological malignancies, but for solid tumors, locoregional and intratumoral CAR T-cell delivery are also included [13, 135]. In the treatment of CNS tumors, intracerebroventricular and intrathecal administration of CAR T-cells has already shown positive outcomes in both preclinical and early clinical trials [135,136,137], which is probably related to the different microenvironments of CSF compared to PBMCs [7, 138, 139]. Two studies applying scRNA-seq and CyTOF respectively showed that CAR T-cells exposed to CSF can promote the formation of a memory-like phenotype through metabolic reprogramming, resulting in higher antitumor activity [138], and increase the expression of activation markers and trafficking/homing signatures, which may facilitate the migration of CAR T-cells to the CNS [140]. In the clinical study by Majzner et al. [141], glioma patients received a regional intracerebroventricular administration after the first GD2 CAR T-cell infusion through the vein. In addition to further radiographic and clinical benefits, local administration was associated with less systemic toxicity, such as CRS. Moreover, scRNA-seq found that intracerebroventricular administration reduced the number of myeloid cells with immunosuppressive properties compared with intravenous administration. In the future, by analyzing the distinct properties of CAR T-cells in PBMCs and CSF or monitoring the regional T cell dynamics more closely related to the TME, single-cell sequencing technologies may provide more insights into the treatment of CNS tumors.
Perspectives for CAR target selection
An ideal CAR T-cell target has the following advantages: (1) coverage: expression on the vast majority of tumor cells; (2) specificity: tumor-specific expression and minimal expression in healthy tissues; (3) stability: continuous and stable expression to avoid antigen escape [142]. However, identifying suitable CAR T-cell targets has been challenging, especially for solid tumors, due to the co-expression of antigens between tumor cells and non-malignant cells, as well as the highly heterogeneous expression among tumor cells. Integrating single-cell multi-dimensional omics data is a promising strategy for CAR target selection.
Previous studies have integrated transcriptomics and/or proteomics data from analyses of both malignant and non-malignant tissues for target discovery [143,144,145,146]. Bosse et al. compared bulk RNA-seq results of neuroblastomas and normal tissues and identified Glypican 2 (GPC2) as a potential CAR T-cell target antigen, which has significant differential expression and extracellular epitopes [146]. However, bulk gene or protein expression data reflect average differences across tissues. Single-cell sequencing technologies with higher resolution can facilitate accurate identification of differentially expressed markers of tumor cell subsets and even rare but important tumor subsets, including cancer stem cells. Furthermore, homogenous target antigen expression can be assessed based on the fraction of tumor cells expressing each antigen. However, the sequencing depth of single-cell sequencing is still insufficient, and the average number of detected genes in a single cell is approximately 2000, with a massive amount of genes missing. With the improvement of sequencing depth, single-cell sequencing technologies may become a powerful tool for CAR target selection.
After screening targets that are specifically overexpressed in tumor cells, the specificity of targets should be verified via multi-organ single-cell atlases, such as HCL [147] and human cell atlas (HCA) [148]. A data portal (CARTSC) integrates HCL and HCA to visualize the expression of CAR target expression in normal tissues at the single-cell level [117]. At present, since most of the single-cell atlases of human organs have been completed and are available as open resources, the single-cell data of specific tissues can be directly obtained for expression verification of CAR targets [149,150,151,152,153]. Based on integrating published scRNA-seq data of tumor and normal tissues/organs, two recent studies systematically analyzed the expression of CAR T-cell target antigens and captured the rare cell types that had previously been omitted in the evaluation of bulk tissues [116, 154]. Particularly, through a machine-learning-based algorithm and the index ECF (expressing cell fraction), Kwon et al. selected ideal gene pairs for dual-target CAR T-cells that were controlled by logical switches (that is, AND, OR and NOT), promoting further tumor coverage and specificity [154].
The selected CAR T-cell target antigens require further validation of target expression on the surface of the cell membrane through proteomics techniques (such as flow cytometry). In addition, it is necessary to evaluate the target antigens in terms of function, immunogenicity, clinical response and other aspects. In particular, CAR T-cell products with novel targets need to be carefully evaluated in rigorous preclinical studies and early clinical trials.
Conclusions and future perspectives
Although CAR T-cell therapy has achieved tremendous clinical success in hematological malignancies, challenges including high manufacturing costs, disease relapse, and adverse events remain to be overcome, and its efficacy in solid tumors requires urgent improvement. The application of single-cell sequencing technologies is a prospect to address these challenges. Single-cell sequencing technologies have natural advantages in deciphering CAR T-cell therapy since CAR T-cells are cellular products and single-cell suspensions. Currently, single-cell sequencing technologies mainly serve as evaluation platforms for deciphering the biological characteristics of CAR T-cell products and the therapeutic response of patients after CAR T-cell infusion. When identifying crucial cell populations and molecular features in certain pathophysiological processes, they can also provide potential strategies to advance CAR-T cell therapy. In addition, single-cell sequencing technologies have the potential to screen ideal CAR targets with sufficient coverage, high specificity and stable expression. Comprehensive single-cell atlases of normal and diseased tissues can be an effective tool for understanding off-target effects and validating candidate targets. It is worth noting that some limitations still exist in single-cell sequencing technologies [155], such as insufficient sequencing coverage and depth, sequencing bias, and high overall cost. Batch effect and data integration across experiments and different sequencing platforms are areas of particular attention that require continuous optimization and standardization of the data processing and analytical pipelines. To meet the demands of clinical accessibility, the accuracy, repeatability, stability and reliability of sequencing data also need to be continuously optimized. Standardized protocols for data analysis as well as physician-friendly interfaces and software should be developed for clinicians. We anticipate that the research benefits brought by single-cell sequencing technologies will push the limits of technical development and accelerate the standardization of computational analytical methods.
Based on the application and broad prospects of single-cell multi-omics technologies in the field of CAR T-cell therapy, we propose a multi-omics research mode to facilitate high-quality research and further clinical translation of CAR T-cell therapy. First, the establishment of a large clinical cohort and a complete sample library is conducive to achieving a subsequent refined experimental design, including the collection of paired samples at multiple time points before and after CAR T-cell treatment and comprehensive clinical and prognostic characteristics of patients, such as disease type, patient baseline characteristics, CAR T-cell product type, infusion dose, and clinical outcome [156]. Sample types include CAR T-cell products, PBMCs, BM, tumors, CSF and target organ tissues. Advanced single-cell multi-omics technologies can be used as a tool for research implementation, including single-cell genomics, epigenomics, transcriptomics, proteomics and spatial transcriptomics, enabling target discovery, mutual validation of the experimental results, and searching for upstream and downstream molecules and pathways in multiple dimensions (Fig. 5A). Based on the above premises and current progress, both basic and clinical research can be guided by multi-omics research mode. The former is dedicated to answering questions about the basic biological characteristics of CAR T-cells, such as the impact of the CAR structure, functional phenotype and manufacturing process on the final CAR T-cell product (Fig. 5B). Notably, from the perspective of cell types, analyzing the interaction among CAR T-cells, endogenous immune cells, stromal cells and tumor cells in the TME of BM and solid tumors would help to explore the mechanisms of tumor cell immune escape leading to relapse and to identify intervention targets. The latter is driven by clinical scientific questions, such as relapse, adverse events, and clinical efficacy, which are common in CAR T-cell therapy. Comparison groups based on clinical outcomes can be set up to identify key differential cell populations and differentially expressed genes to reveal potential mechanisms and therapeutic targets, and functional validation can be conducted in cells or animal models. Furthermore, research applying multi-omics technologies in a large-scale clinical cohort can also be conducted. Based on artificial intelligence, machine learning algorithms, and unsupervised and unbiased methods, detailed subgroup analysis of patients’ clinical information would be performed to determine risk prediction, prognosis stratification, therapeutic classification and personalized therapy (Fig. 5C).

The multi-omics research mode for CAR T-cell therapy. (A) In the multi-omics research mode, a large clinical cohort with complete clinical information as well as biological samples and single-cell multi-omics technologies would be combined to create multi-level metadata from micro to macros, which provides guidance for the development of precision medicine. In the field of CAR T-cell therapy, integration of multi-dimensional omics data promotes a comprehensive description of the molecular regulatory network during CAR T-cell functional processes, leading to the discovery of new mechanisms and targeted treatment strategies for CAR T-cell therapy and thus bringing more insights into this field at both the basic and clinical levels. (B) Basic research is dedicated to illustrating the basic biological characteristics of CAR T-cells, including the effect of each manufacturing stage on the final cellular products, the heterogeneity in the evolution of functional phenotypes during the CAR T-cell functional process, the interaction between CAR T-cells and various cells within the TME and the selection of suitable target antigens, which will advance the safety and efficacy of CAR T-cell therapy. (C) Clinical question-driven research focus on tracing the reasons for different clinical outcomes (including resistance, relapse, and toxicities) in patients treated with CAR T-cells. Grouping patients with different characteristics and comparing differences in metadata may bring more meaningful discoveries and guide further improvement of CAR T-cell therapy, such as combination therapy and engineered CAR T-cells
In summary, leveraging single-cell sequencing technologies has brought new cellular and molecular insights to deciphering and advancing CAR T-cell therapy, with the potential to answer significant scientific questions related to the efficacy and safety of CAR T-cell therapy and is likely to revolutionize the methods of diagnosis and treatment and promote further progress in precision medicine.
Data Availability
Not applicable.
Abbreviations
- BAF:
-
Ballele frequency
- B-ALL:
-
B-cell acute lymphoblastic leukemia
- BATF:
-
Basic leucine zipper ATF-like transcription factor
- BBB:
-
Blood brain barrier
- BCMA:
-
B-cell maturation antigen
- BM:
-
Bone marrow
- B-NHL:
-
B-cell non-Hodgkin’s lymphoma
- BTK:
-
Bruton’s tyrosine kinase
- CAR:
-
Chimeric antigen receptor
- CDK4i/6i:
-
Cyclin-dependent kinases 4 and 6 inhibitors
- CITE-seq:
-
Cellular indexing of transcriptomes and epitopes by sequencing
- CLL:
-
Chronic lymphocytic leukemia
- CNS:
-
Central nervous system
- CR:
-
Complete response
- CRS:
-
Cytokine release syndrome
- CSF:
-
Cerebrospinal fluid
- Cy:
-
Cyclophosphamide
- CyTOF:
-
Cytometry by time-of-flight
- ECM:
-
Extracellular matrix
- EMT:
-
Epithelial-mesenchymal transition
- EVs:
-
Extracellular vesicles
- FISH:
-
Fluorescence in situ hybridization
- Flu:
-
Fludarabine
- GBM:
-
Glioblastoma
- GM-CSF:
-
Granulocyte-macrophage colony-stimulating factor
- GO:
-
Gene ontology
- GPC2:
-
Glypican 2
- HCA:
-
Human cell atlas
- HCL:
-
Human cell landscape
- HHV-6:
-
Human herpesvirus 6
- HL:
-
Hodgkin’s lymphoma
- HSCP:
-
Hematopoietic stem cell progenitor
- IACs:
-
ICANS-associated cells
- ICANS:
-
Immune effector cell-associated neurotoxicity syndrome
- ID2:
-
Inhibitor of DNA binding 2
- ID3:
-
Inhibitor of DNA binding 3
- IFITM:
-
Interferon-induced transmembrane
- IKZF2:
-
Ikaros family zinc finger protein 2
- IPs:
-
Infusion products
- LAG-3:
-
Lymphocyte activation 3
- LBCL:
-
Large B-cell lymphoma
- LDH:
-
Lactate dehydrogenase
- LVs:
-
Lentiviral vectors
- MCL:
-
Mantle cell lymphoma
- MHC:
-
Major histocompatibility complex
- MM:
-
Multiple myeloma
- Ox:
-
Oxaliplatin
- PBMCs:
-
Peripheral blood mononuclear cells
- PCL:
-
Plasma cell leukemia
- PD:
-
Progressive disease
- PFS:
-
Progression-free survival
- PR:
-
Partial response
- PRG:
-
Potentially risky gene
- PSI:
-
Polyfunctionality strength index
- scATAC-seq:
-
Single-cell assay for transposase-accessible chromatin sequencing
- SCBC:
-
Single-cell barcode chip
- scFv:
-
Single-chain variable fragment
- scRNA-seq:
-
Single-cell RNA sequencing
- scTCR-seq:
-
Single-cell T-cell receptor sequencing
- STING:
-
Stimulator of IFN genes
- TAAs:
-
Tumor-associated antigens
- TCR:
-
T cell receptor
- TIGIT:
-
T cell immunoglobulin and ITIM domain
- TLE4:
-
Transducin like enhancer of split 4
- TME:
-
Tumor microenvironment
- TRAC:
-
T-cell receptor α constant
- VSV:
-
Vesicular stomatitis virus
References
Maude SL, Frey N, Shaw PA, Aplenc R, Barrett DM, Bunin NJ, et al. Chimeric antigen receptor T cells for sustained remissions in leukemia. N Engl J Med. 2014;371:1507–17.
Jacobson CA, Chavez JC, Sehgal AR, William BM, Munoz J, Salles G, et al. Axicabtagene ciloleucel in relapsed or refractory indolent non-hodgkin lymphoma (ZUMA-5): a single-arm, multicentre, phase 2 trial. Lancet Oncol. 2022;23:91–103.
Frey NV, Gill S, Hexner EO, Schuster S, Nasta S, Loren A, et al. Long-term outcomes from a randomized dose optimization study of chimeric Antigen receptor modified T cells in relapsed chronic lymphocytic leukemia. J Clin Oncol. 2020;38:2862–71.
Ramos CA, Grover NS, Beaven AW, Lulla PD, Wu MF, Ivanova A, et al. Anti-CD30 CAR-T cell therapy in relapsed and refractory Hodgkin Lymphoma. J Clin Oncol. 2020;38:3794–804.
Park JH, Rivière I, Gonen M, Wang X, Sénéchal B, Curran KJ, et al. Long-term follow-up of CD19 CAR therapy in Acute Lymphoblastic Leukemia. N Engl J Med. 2018;378:449–59.
Maude SL, Laetsch TW, Buechner J, Rives S, Boyer M, Bittencourt H, et al. Tisagenlecleucel in Children and Young adults with B-Cell lymphoblastic leukemia. N Engl J Med. 2018;378:439–48.
Brown CE, Alizadeh D, Starr R, Weng L, Wagner JR, Naranjo A, et al. Regression of Glioblastoma after chimeric Antigen receptor T-Cell therapy. N Engl J Med. 2016;375:2561–9.
Beatty GL, O’Hara MH, Lacey SF, Torigian DA, Nazimuddin F, Chen F, et al. Activity of mesothelin-specific chimeric Antigen receptor T cells against pancreatic carcinoma metastases in a phase 1 trial. Gastroenterology. 2018;155:29–32.
Ahmed N, Brawley VS, Hegde M, Robertson C, Ghazi A, Gerken C, et al. Human epidermal growth factor receptor 2 (HER2) -Specific chimeric Antigen receptor-modified T cells for the Immunotherapy of HER2-Positive sarcoma. J Clin Oncol. 2015;33:1688–96.
Qi C, Gong J, Li J, Liu D, Qin Y, Ge S, et al. Claudin18.2-specific CAR T cells in gastrointestinal cancers: phase 1 trial interim results. Nat Med. 2022;28:1189–98.
Narayan V, Barber-Rotenberg JS, Jung IY, Lacey SF, Rech AJ, Davis MM, et al. PSMA-targeting TGFβ-insensitive armored CAR T cells in metastatic castration-resistant prostate cancer: a phase 1 trial. Nat Med. 2022;28:724–34.
Larson RC, Maus MV. Recent advances and discoveries in the mechanisms and functions of CAR T cells. Nat Rev Cancer. 2021;21:145–61.
Hou AJ, Chen LC, Chen YY. Navigating CAR-T cells through the solid-tumour microenvironment. Nat Rev Drug Discov. 2021;20:531–50.
Shah NN, Fry TJ. Mechanisms of resistance to CAR T cell therapy. Nat Rev Clin Oncol. 2019;16:372–85.
Chow A, Perica K, Klebanoff CA, Wolchok JD. Clinical implications of T cell exhaustion for cancer immunotherapy. Nat Rev Clin Oncol. 2022;19:775–90.
Schubert ML, Schmitt M, Wang L, Ramos CA, Jordan K, Müller-Tidow C, et al. Side-effect management of chimeric antigen receptor (CAR) T-cell therapy. Ann Oncol. 2021;32:34–48.
Maryamchik E, Gallagher KME, Preffer FI, Kadauke S, Maus MV. New directions in chimeric antigen receptor T cell [CAR-T] therapy and related flow cytometry. Cytometry B Clin Cytom. 2020;98:299–327.
Sarikonda G, Pahuja A, Kalfoglou C, Burns K, Nguyen K, Ch’en IL, et al. Monitoring CAR-T cell kinetics in clinical trials by multiparametric flow cytometry: benefits and challenges. Cytometry B Clin Cytom. 2021;100:72–8.
Kharchenko PV. The triumphs and limitations of computational methods for scRNA-seq. Nat Methods. 2021;18:723–32.
Erfanian N, Derakhshani A, Nasseri S, Fereidouni M, Baradaran B, Jalili Tabrizi N, et al. Immunotherapy of cancer in single-cell RNA sequencing era: a precision medicine perspective. Biomed Pharmacother. 2022;146:112558.
Hwang B, Lee JH, Bang D. Single-cell RNA sequencing technologies and bioinformatics pipelines. Exp Mol Med. 2018;50:1–14.
Papalexi E, Satija R. Single-cell RNA sequencing to explore immune cell heterogeneity. Nat Rev Immunol. 2018;18:35–45.
Xu X, Hua X, Mo H, Hu S, Song J. Single-cell RNA sequencing to identify cellular heterogeneity and targets in cardiovascular diseases: from bench to bedside. Basic Res Cardiol. 2023;118:7.
Ghorashian S, Kramer AM, Onuoha S, Wright G, Bartram J, Richardson R, et al. Enhanced CAR T cell expansion and prolonged persistence in pediatric patients with ALL treated with a low-affinity CD19 CAR. Nat Med. 2019;25:1408–14.
Wagner DL, Fritsche E, Pulsipher MA, Ahmed N, Hamieh M, Hegde M, et al. Immunogenicity of CAR T cells in cancer therapy. Nat Rev Clin Oncol. 2021;18:379–93.
Cappell KM, Kochenderfer JN. A comparison of chimeric antigen receptors containing CD28 versus 4-1BB costimulatory domains. Nat Rev Clin Oncol. 2021;18:715–27.
Meyran D, Terry RL, Zhu JJ, Haber M, Ziegler DS, Ekert PG, et al. Early-phenotype CAR-T cells for the treatment of pediatric cancers. Ann Oncol. 2021;32:1366–80.
Liu Y, Sperling AS, Smith EL, Mooney DJ. Optimizing the manufacturing and antitumour response of CAR T therapy. Nat Rev Bioeng. 2023:1–15.
Lu J, Jiang G. The journey of CAR-T therapy in hematological malignancies. Mol Cancer. 2022;21:194.
Gardner RA, Finney O, Annesley C, Brakke H, Summers C, Leger K, et al. Intent-to-treat leukemia remission by CD19 CAR T cells of defined formulation and dose in children and young adults. Blood. 2017;129:3322–31.
Turtle CJ, Hanafi LA, Berger C, Gooley TA, Cherian S, Hudecek M, et al. CD19 CAR-T cells of defined CD4+:CD8 + composition in adult B cell ALL patients. J Clin Invest. 2016;126:2123–38.
Sommermeyer D, Hudecek M, Kosasih PL, Gogishvili T, Maloney DG, Turtle CJ, et al. Chimeric antigen receptor-modified T cells derived from defined CD8 + and CD4 + subsets confer superior antitumor reactivity in vivo. Leukemia. 2016;30:492–500.
Fraietta JA, Lacey SF, Orlando EJ, Pruteanu-Malinici I, Gohil M, Lundh S, et al. Determinants of response and resistance to CD19 chimeric antigen receptor (CAR) T cell therapy of chronic lymphocytic leukemia. Nat Med. 2018;24:563–71.
Chen GM, Chen C, Das RK, Gao P, Chen CH, Bandyopadhyay S, et al. Integrative bulk and single-cell profiling of premanufacture T-cell populations reveals factors mediating long-term persistence of CAR T-cell therapy. Cancer Discov. 2021;11:2186–99.
Gumber D, Wang LD. Improving CAR-T immunotherapy: overcoming the challenges of T cell exhaustion. EBioMedicine. 2022;77:103941.
Sheih A, Voillet V, Hanafi LA, DeBerg HA, Yajima M, Hawkins R, et al. Clonal kinetics and single-cell transcriptional profiling of CAR-T cells in patients undergoing CD19 CAR-T immunotherapy. Nat Commun. 2020;11:219.
Vormittag P, Gunn R, Ghorashian S, Veraitch FS. A guide to manufacturing CAR T cell therapies. Curr Opin Biotechnol. 2018;53:164–81.
Roddie C, O’Reilly M, Dias Alves Pinto J, Vispute K, Lowdell M. Manufacturing chimeric antigen receptor T cells: issues and challenges. Cytotherapy. 2019;21:327–40.
Depil S, Duchateau P, Grupp SA, Mufti G, Poirot L. ‘off-the-shelf’ allogeneic CAR T cells: development and challenges. Nat Rev Drug Discov. 2020;19:185–99.
Luecken MD, Theis FJ. Current best practices in single-cell RNA-seq analysis: a tutorial. Mol Syst Biol. 2019;15:e8746.
Andrews TS, Kiselev VY, McCarthy D, Hemberg M. Tutorial: guidelines for the computational analysis of single-cell RNA sequencing data. Nat Protoc. 2021;16:1–9.
Pai JA, Satpathy AT. High-throughput and single-cell T cell receptor sequencing technologies. Nat Methods. 2021;18:881–92.
Frank ML, Lu K, Erdogan C, Han Y, Hu J, Wang T, et al. T cell receptor repertoire sequencing in the era of cancer immunotherapy. Clin Cancer Res. 2023;14:994–1008.
Chiffelle J, Genolet R, Perez MA, Coukos G, Zoete V, Harari A. T-cell repertoire analysis and metrics of diversity and clonality. Curr Opin Biotechnol. 2020;65:284–95.
Buenrostro JD, Wu B, Litzenburger UM, Ruff D, Gonzales ML, Snyder MP, et al. Single-cell chromatin accessibility reveals principles of regulatory variation. Nature. 2015;523:486–90.
Gennert DG, Lynn RC, Granja JM, Weber EW, Mumbach MR, Zhao Y, et al. Dynamic chromatin regulatory landscape of human CAR T cell exhaustion. Proc Natl Acad Sci U S A. 2021;118:e2104758118.
Jiang P, Zhang Z, Hu Y, Liang Z, Han Y, Li X, et al. Single-cell ATAC-seq maps the comprehensive and dynamic chromatin accessibility landscape of CAR-T cell dysfunction. Leukemia. 2022;36:2656–68.
Bendall SC, Simonds EF, Qiu P, Amir el AD, Krutzik PO, Finck R, et al. Single-cell mass cytometry of differential immune and drug responses across a human hematopoietic continuum. Science. 2011;332:687–96.
Doerr A. Single-cell proteomics. Nat Methods. 2019;16:20.
Stoeckius M, Hafemeister C, Stephenson W, Houck-Loomis B, Chattopadhyay PK, Swerdlow H, et al. Simultaneous epitope and transcriptome measurement in single cells. Nat Methods. 2017;14:865–8.
Propper DJ, Balkwill FR. Harnessing cytokines and chemokines for cancer therapy. Nat Rev Clin Oncol. 2022;19:237–53.
Berraondo P, Sanmamed MF, Ochoa MC, Etxeberria I, Aznar MA, Pérez-Gracia JL, et al. Cytokines in clinical cancer immunotherapy. Br J Cancer. 2019;120:6–15.
Vistain LF, Tay S. Single-cell proteomics. Trends Biochem Sci. 2021;46:661–72.
Lu Y, Xue Q, Eisele MR, Sulistijo ES, Brower K, Han L, et al. Highly multiplexed profiling of single-cell effector functions reveals deep functional heterogeneity in response to pathogenic ligands. Proc Natl Acad Sci U S A. 2015;112:E607-E15.
Ma C, Cheung AF, Chodon T, Koya RC, Wu Z, Ng C, et al. Multifunctional T-cell analyses to study response and progression in adoptive cell transfer immunotherapy. Cancer Discov. 2013;3:418–29.
Xue Q, Bettini E, Paczkowski P, Ng C, Kaiser A, McConnell T, et al. Single-cell multiplexed cytokine profiling of CD19 CAR-T cells reveals a diverse landscape of polyfunctional antigen-specific response. J Immunother Cancer. 2017;5:85.
Zurko JC, Xu H, Chaney K, Schneider D, Szabo A, Hari P, et al. Bispecific targeting of CD20 and CD19 increases polyfunctionality of chimeric antigen receptor T-cell products in B-cell malignancies. Cytotherapy. 2022;24:767–73.
Rossi J, Paczkowski P, Shen Y-W, Morse K, Flynn B, Kaiser A, et al. Preinfusion polyfunctional anti-CD19 chimeric antigen receptor T cells are associated with clinical outcomes in NHL. Blood. 2018;132:804–14.
Boroughs AC, Larson RC, Marjanovic ND, Gosik K, Castano AP, Porter CBM, et al. A distinct transcriptional program in human CAR T cells bearing the 4-1BB signaling domain revealed by scRNA-Seq. Mol Ther. 2020;28:2577–92.
Wang Y, Buck A, Grimaud M, Culhane AC, Kodangattil S, Razimbaud C, et al. Anti-CAIX BBζ CAR4/8 T cells exhibit superior efficacy in a ccRCC mouse model. Mol Ther Oncolytics. 2022;24:385–99.
Bai Z, Lundh S, Kim D, Woodhouse S, Barrett DM, Myers RM, et al. Single-cell multiomics dissection of basal and antigen-specific activation states of CD19-targeted CAR T cells. J Immunother Cancer. 2021;9:e002328.
Zhang C, Wang XQ, Zhang RL, Liu F, Wang Y, Yan ZL, et al. Donor-derived CD19 CAR-T cell therapy of relapse of CD19-positive B-ALL post allotransplant. Leukemia. 2021;35:1563–70.
Zhang X, Yang J, Li W, Zhang G, Su Y, Shi Y, et al. Feasibility, and efficacy of Donor-Derived cd19-Targeted Car t-Cell therapy in Refractory/Relapsed (r/r) b-Cell Acute Lymphoblastic Leukemia (b-all) patients. Blood. 2020;136:4–6.
Chen S, Zhang Y, Fang C, Zhang N, Wang Y, Chen R, et al. Donor-derived and off-the-shelf allogeneic anti-CD19 CAR T-cell therapy for R/R ALL and NHL: a systematic review and meta-analysis. Crit Rev Oncol Hematol. 2022;179:103807.
Sachdeva M, Duchateau P, Depil S, Poirot L, Valton J. Granulocyte-macrophage colony-stimulating factor inactivation in CAR T-cells prevents monocyte-dependent release of key cytokine release syndrome mediators. J Biol Chem. 2019;294:5430–7.
Sterner RM, Sakemura R, Cox MJ, Yang N, Khadka RH, Forsman CL, et al. GM-CSF inhibition reduces cytokine release syndrome and neuroinflammation but enhances CAR-T cell function in xenografts. Blood. 2019;133:697–709.
Hu Y, Wang J, Wei G, Yu J, Luo Y, Shi J, et al. A retrospective comparison of allogenic and autologous chimeric antigen receptor T cell therapy targeting CD19 in patients with relapsed/refractory acute lymphoblastic leukemia. Bone Marrow Transplant. 2019;54:1208–17.
Panch SR, Srivastava SK, Elavia N, McManus A, Liu S, Jin P, et al. Effect of cryopreservation on autologous chimeric Antigen receptor T cell characteristics. Mol Ther. 2019;27:1275–85.
Palen K, Zurko J, Johnson BD, Hari P, Shah NN. Manufacturing chimeric antigen receptor T cells from cryopreserved peripheral blood cells: time for a collect-and-freeze model? Cytotherapy. 2021;23:985–90.
Haradhvala NJ, Leick MB, Maurer K, Gohil SH, Larson RC, Yao N, et al. Distinct cellular dynamics associated with response to CAR-T therapy for refractory B cell lymphoma. Nat Med. 2022;28:1848–59.
Rodriguez-Marquez P, Calleja-Cervantes ME, Serrano G, Oliver-Caldes A, Palacios-Berraquero ML, Martin-Mallo A, et al. CAR density influences antitumoral efficacy of BCMA CAR T cells and correlates with clinical outcome. Sci Adv. 2022;8:eabo0514.
Charitidis FT, Adabi E, Thalheimer FB, Clarke C, Buchholz CJ. Monitoring CAR T cell generation with a CD8-targeted lentiviral vector by single-cell transcriptomics. Mol Ther Methods Clin Dev. 2021;23:359–69.
Bai Z, Woodhouse S, Zhao Z, Arya R, Govek K, Kim D, et al. Single-cell antigen-specific landscape of CAR T infusion product identifies determinants of CD19-positive relapse in patients with ALL. Sci Adv. 2022;8:eabj2820.
Wang X, Peticone C, Kotsopoulou E, Göttgens B, Calero-Nieto FJ. Single-cell transcriptome analysis of CAR T-cell products reveals subpopulations, stimulation, and exhaustion signatures. Oncoimmunology. 2021;10:1866287.
Xhangolli I, Dura B, Lee G, Kim D, Xiao Y, Fan R. Single-cell analysis of CAR-T cell activation reveals a mixed T(H)1/T(H)2 response Independent of differentiation. Genomics Proteomics Bioinformatics. 2019;17:129–39.
Wilson TL, Kim H, Chou C-H, Langfitt D, Mettelman RC, Minervina AA, et al. Common trajectories of highly effective CD19-Specific CAR T cells identified by endogenous T-cell receptor lineages. Cancer Discov. 2022;12:2098–119.
Stein AM, Grupp SA, Levine JE, Laetsch TW, Pulsipher MA, Boyer MW, et al. Tisagenlecleucel Model-Based Cellular Kinetic Analysis of chimeric Antigen Receptor-T cells. CPT Pharmacometrics Syst Pharmacol. 2019;8:285–95.
Liu C, Ayyar VS, Zheng X, Chen W, Zheng S, Mody H, et al. Model-based Cellular Kinetic Analysis of chimeric Antigen Receptor-T cells in humans. Clin Pharmacol Ther. 2021;109:716–27.
Chaudhury A, Zhu X, Chu L, Goliaei A, June CH, Kearns JD, et al. Chimeric Antigen receptor T cell therapies: a review of Cellular Kinetic-Pharmacodynamic modeling approaches. J Clin Pharmacol. 2020;60 Suppl 1:S147-s59.
Li X, Guo X, Zhu Y, Wei G, Zhang Y, Li X, et al. Single-cell transcriptomic analysis reveals BCMA CAR-T Cell Dynamics in a patient with refractory primary plasma cell leukemia. Mol Ther. 2021;29:645–57.
Melenhorst JJ, Chen GM, Wang M, Porter DL, Chen C, Collins MA, et al. Decade-long leukaemia remissions with persistence of CD4(+) CAR T cells. Nature. 2022;602:503–9.
Korell F, Berger TR, Maus MV. Understanding CAR T cell-tumor interactions: paving the way for successful clinical outcomes. Med. 2022;3:538–64.
Ivashkiv LB. IFNγ: signalling, epigenetics and roles in immunity, metabolism, disease and cancer immunotherapy. Nat Rev Immunol. 2018;18:545–58.
Gocher AM, Workman CJ, Vignali DAA. Interferon-γ: teammate or opponent in the tumour microenvironment? Nat Rev Immunol. 2022;22:158–72.
Boulch M, Cazaux M, Loe-Mie Y, Thibaut R, Corre B, Lemaître F, et al. A cross-talk between CAR T cell subsets and the tumor microenvironment is essential for sustained cytotoxic activity. Sci Immunol. 2021;6:eabd4344.
Alizadeh D, Wong RA, Gholamin S, Maker M, Aftabizadeh M, Yang X, et al. IFNγ is critical for CAR T cell-mediated myeloid activation and induction of endogenous immunity. Cancer Discov. 2021;11:2248–65.
Good CR, Aznar MA, Kuramitsu S, Samareh P, Agarwal S, Donahue G, et al. An NK-like CAR T cell transition in CAR T cell dysfunction. Cell. 2021;184:6081 – 100.e26.
Deng Q, Han G, Puebla-Osorio N, Ma MCJ, Strati P, Chasen B, et al. Characteristics of anti-CD19 CAR T cell infusion products associated with efficacy and toxicity in patients with large B cell lymphomas. Nat Med. 2020;26:1878–87.
Jackson Z, Hong C, Schauner R, Dropulic B, Caimi PF, de Lima M, et al. Sequential single-cell transcriptional and protein marker profiling reveals TIGIT as a marker of CD19 CAR-T cell dysfunction in patients with Non-Hodgkin Lymphoma. Cancer Discov. 2022;12:1886–903.
Good Z, Spiegel JY, Sahaf B, Malipatlolla MB, Ehlinger ZJ, Kurra S, et al. Post-infusion CAR T cells identify patients resistant to CD19-CAR therapy. Nat Med. 2022;28:1860–71.
Singh N, Lee YG, Shestova O, Ravikumar P, Hayer KE, Hong SJ, et al. Impaired death receptor signaling in Leukemia causes Antigen-Independent resistance by inducing CAR T-cell dysfunction. Cancer Discov. 2020;10:552–67.
Lemoine J, Ruella M, Houot R. Overcoming intrinsic resistance of Cancer cells to CAR T-Cell killing. Clin Cancer Res. 2021;27:6298–306.
Xu X, Sun Q, Liang X, Chen Z, Zhang X, Zhou X, et al. Mechanisms of Relapse after CD19 CAR T-Cell Therapy for Acute Lymphoblastic Leukemia and its Prevention and Treatment strategies. Front Immunol. 2019;10:2664.
Nie Y, Lu W, Chen D, Tu H, Guo Z, Zhou X, et al. Mechanisms underlying CD19-positive ALL relapse after anti-CD19 CAR T cell therapy and associated strategies. Biomark Res. 2020;8:18.
Xu X, Huang S, Xiao X, Sun Q, Liang X, Chen S, et al. Challenges and clinical strategies of CAR T-Cell therapy for Acute Lymphoblastic Leukemia: overview and developments. Front Immunol. 2020;11:569117.
Dhodapkar KM, Cohen AD, Kaushal A, Garfall AL, Manalo RJ, Carr AR, et al. Changes in bone marrow tumor and Immune cells correlate with durability of remissions following BCMA CAR T therapy in Myeloma. Blood Cancer Discov. 2022:3:490–501.
Jonsson VD, Ng RH, Dullerud N, Wong RA, Hibbard J, Wang D, et al. CAR T cell therapy drives endogenous locoregional T cell dynamics in a responding patient with glioblastoma. bioRxiv. 2021:2021.09.22.460392.
Jiang VC, Hao D, Jain P, Li Y, Cai Q, Yao Y, et al. TIGIT is the central player in T-cell suppression associated with CAR T-cell relapse in mantle cell lymphoma. Mol Cancer. 2022;21:185.
Lemoine J, Ruella M, Houot R. Born to survive: how cancer cells resist CAR T cell therapy. J Hematol Oncol. 2021;14:199.
Bueno C, Barrera S, Bataller A, Ortiz-Maldonado V, Elliot N, O’Byrne S, et al. CD34 + CD19-CD22 + B-cell progenitors may underlie phenotypic escape in patients treated with CD19-directed therapies. Blood. 2022;140:38–44.
Rabilloud T, Potier D, Pankaew S, Nozais M, Loosveld M, Payet-Bornet D. Single-cell profiling identifies pre-existing CD19-negative subclones in a B-ALL patient with CD19-negative relapse after CAR-T therapy. Nat Commun. 2021;12:865.
Im NG, Guillaumet-Adkins A, Wal M, Rogers AJ, Frede J, Havig CC, et al. Regulatory programs of B-cell activation and germinal center reaction allow B-ALL escape from CD19 CAR T-cell therapy. Cancer Immunol Res. 2022;10:1055–1068.
Depoil D, Fleire S, Treanor BL, Weber M, Harwood NE, Marchbank KL, et al. CD19 is essential for B cell activation by promoting B cell receptor-antigen microcluster formation in response to membrane-bound ligand. Nat Immunol. 2008;9:63–72.
Xiao X, Huang S, Chen S, Wang Y, Sun Q, Xu X, et al. Mechanisms of cytokine release syndrome and neurotoxicity of CAR T-cell therapy and associated prevention and management strategies. J Exp Clin Cancer Res. 2021;40:367.
Morris EC, Neelapu SS, Giavridis T, Sadelain M. Cytokine release syndrome and associated neurotoxicity in cancer immunotherapy. Nat Rev Immunol. 2022;22:85–96.
Li X, Shao M, Zeng X, Qian P, Huang H. Signaling pathways in the regulation of cytokine release syndrome in human diseases and intervention therapy. Signal Transduct Target Ther. 2021;6:367.
Giavridis T, van der Stegen SJC, Eyquem J, Hamieh M, Piersigilli A, Sadelain M. CAR T cell-induced cytokine release syndrome is mediated by macrophages and abated by IL-1 blockade. Nat Med. 2018;24:731–8.
Norelli M, Camisa B, Barbiera G, Falcone L, Purevdorj A, Genua M, et al. Monocyte-derived IL-1 and IL-6 are differentially required for cytokine-release syndrome and neurotoxicity due to CAR T cells. Nat Med. 2018;24:739–48.
Fajgenbaum DC, June CH. Cytokine storm. N Engl J Med. 2020;383:2255–73.
Bickel M. The role of interleukin-8 in inflammation and mechanisms of regulation. J Periodontol. 1993;64:456–60.
Deshmane SL, Kremlev S, Amini S, Sawaya BE. Monocyte chemoattractant protein-1 (MCP-1): an overview. J Interferon Cytokine Res. 2009;29:313–26.
Holtzman NG, Xie H, Bentzen S, Kesari V, Bukhari A, El Chaer F, et al. Immune effector cell-associated neurotoxicity syndrome after chimeric antigen receptor T-cell therapy for lymphoma: predictive biomarkers and clinical outcomes. Neuro Oncol. 2021;23:112–21.
Parker KR, Migliorini D, Perkey E, Yost KE, Bhaduri A, Bagga P, et al. Single-cell analyses identify brain mural cells expressing CD19 as potential off-tumor targets for CAR-T immunotherapies. Cell. 2020;183.
Lareau CA, Yin Y, Maurer K, Sandor KD, Yagnik G, Peña J, et al. Latent human herpesvirus 6 is reactivated in chimeric antigen receptor T cells. bioRxiv. 2022:2022.08.12.503683.
Flugel CL, Majzner RG, Krenciute G, Dotti G, Riddell SR, Wagner DL, et al. Overcoming on-target, off-tumour toxicity of CAR T cell therapy for solid tumours. Nat Rev Clin Oncol. 2023;20:49–62.
Zhang Y, Li Y, Cao W, Wang F, Xie X, Li Y, et al. Single-cell analysis of Target Antigens of CAR-T reveals a potential Landscape of “On-Target, Off-Tumor Toxicity”. Front Immunol. 2021;12:799206.
Jing Y, Liu Y, Li Q, Ye Y, Diao L, Huang Y, et al. Expression of chimeric antigen receptor therapy targets detected by single-cell sequencing of normal cells may contribute to off-tumor toxicity. Cancer Cell. 2021;39:1558–9.
Xiao X, Wang Y, Zou Z, Yang Y, Wang X, Xin X, et al. Combination strategies to optimize the efficacy of chimeric antigen receptor T cell therapy in haematological malignancies. Front Immunol. 2022;13:954235.
Zhu S, Zhang T, Zheng L, Liu H, Song W, Liu D, et al. Combination strategies to maximize the benefits of cancer immunotherapy. J Hematol Oncol. 2021;14:156.
Srivastava S, Furlan SN, Jaeger-Ruckstuhl CA, Sarvothama M, Berger C, Smythe KS, et al. Immunogenic chemotherapy enhances recruitment of CAR-T cells to lung tumors and improves Antitumor Efficacy when combined with checkpoint blockade. Cancer Cell. 2021;39:193–208.e10.
Pfirschke C, Engblom C, Rickelt S, Cortez-Retamozo V, Garris C, Pucci F, et al. Immunogenic chemotherapy sensitizes tumors to checkpoint blockade therapy. Immunity. 2016;44:343–54.
Sen T, Rodriguez BL, Chen L, Corte CMD, Morikawa N, Fujimoto J, et al. Targeting DNA damage response promotes Antitumor immunity through STING-Mediated T-cell activation in small cell Lung Cancer. Cancer Discov. 2019;9:646–61.
Xu N, Palmer DC, Robeson AC, Shou P, Bommiasamy H, Laurie SJ, et al. STING agonist promotes CAR T cell trafficking and persistence in breast cancer. J Exp Med. 2021;218:e20200844.
Fassl A, Geng Y, Sicinski P. CDK4 and CDK6 kinases: from basic science to cancer therapy. Science. 2022;375:eabc1495.
Sherr CJ, Beach D, Shapiro GI. Targeting CDK4 and CDK6: from Discovery to Therapy. Cancer Discov. 2016;6:353–67.
Lelliott EJ, Kong IY, Zethoven M, Ramsbottom KM, Martelotto LG, Meyran D, et al. CDK4/6 inhibition promotes Antitumor Immunity through the induction of T-cell memory. Cancer Discov. 2021;11:2582–601.
Bashor CJ, Hilton IB, Bandukwala H, Smith DM, Veiseh O. Engineering the next generation of cell-based therapeutics. Nat Rev Drug Discov. 2022;21:655–75.
Irvine DJ, Maus MV, Mooney DJ, Wong WW. The future of engineered immune cell therapies. Science. 2022;378:853–8.
Huang R, Li X, He Y, Zhu W, Gao L, Liu Y, et al. Recent advances in CAR-T cell engineering. J Hematol Oncol. 2020;13:86.
Wang D, Prager BC, Gimple RC, Aguilar B, Alizadeh D, Tang H, et al. CRISPR Screening of CAR T cells and Cancer stem cells reveals critical dependencies for cell-based therapies. Cancer Discov. 2021;11:1192–211.
Zhang J, Hu Y, Yang J, Li W, Zhang M, Wang Q, et al. Non-viral, specifically targeted CAR-T cells achieve high safety and efficacy in B-NHL. Nature. 2022;609:369–374.
Mueller KP, Piscopo NJ, Forsberg MH, Saraspe LA, Das A, Russell B, et al. Production and characterization of virus-free, CRISPR-CAR T cells capable of inducing solid tumor regression. J Immunother Cancer. 2022;10:e004446.
Johnson LR, Lee DY, Eacret JS, Ye D, June CH, Minn AJ. The immunostimulatory RNA RN7SL1 enables CAR-T cells to enhance autonomous and endogenous immune function. Cell. 2021;184:4981–4995.e14.
Brog RA, Ferry SL, Schiebout CT, Messier CM, Cook WJ, Abdullah L, et al. Superkine IL-2 and IL-33 armored CAR T cells reshape the Tumor Microenvironment and reduce growth of multiple solid tumors. Cancer Immunol Res. 2022;10:962–77.
Cherkassky L, Hou Z, Amador-Molina A, Adusumilli PS. Regional CAR T cell therapy: an ignition key for systemic immunity in solid tumors. Cancer cell. 2022;40:569–74.
Nellan A, Rota C, Majzner R, Lester-McCully CM, Griesinger AM, Mulcahy Levy JM, et al. Durable regression of Medulloblastoma after regional and intravenous delivery of anti-HER2 chimeric antigen receptor T cells. J Immunother Cancer. 2018;6:30.
Donovan LK, Delaidelli A, Joseph SK, Bielamowicz K, Fousek K, Holgado BL, et al. Locoregional delivery of CAR T cells to the cerebrospinal fluid for treatment of metastatic medulloblastoma and ependymoma. Nat Med. 2020;26:720–31.
Wang X, Huynh C, Urak R, Weng L, Walter M, Lim L, et al. The Cerebroventricular Environment modifies CAR T cells for potent activity against both Central Nervous System and systemic lymphoma. Cancer Immunol Res. 2021;9:75–88.
Theruvath J, Sotillo E, Mount CW, Graef CM, Delaidelli A, Heitzeneder S, et al. Locoregionally administered B7-H3-targeted CAR T cells for treatment of atypical teratoid/rhabdoid tumors. Nat Med. 2020;26:712–9.
Goldberg L, Haas ER, Vyas V, Urak R, Forman SJ, Wang X. Single-cell analysis by mass cytometry reveals CD19 CAR T cell spatiotemporal plasticity in patients. Oncoimmunology. 2022;11:2040772.
Majzner RG, Ramakrishna S, Yeom KW, Patel S, Chinnasamy H, Schultz LM, et al. GD2-CAR T cell therapy for H3K27M-mutated diffuse midline gliomas. Nature. 2022;603:934–41.
Wei J, Han X, Bo J, Han W. Target selection for CAR-T therapy. J Hematol Oncol. 2019;12:62.
MacKay M, Afshinnekoo E, Rub J, Hassan C, Khunte M, Baskaran N, et al. The therapeutic landscape for cells engineered with chimeric antigen receptors. Nat Biotechnol. 2020;38:233–44.
Hu Z, Yuan J, Long M, Jiang J, Zhang Y, Zhang T, et al. The Cancer Surfaceome Atlas integrates genomic, functional and drug response data to identify actionable targets. Nat Cancer. 2021;2:1406–22.
Perna F, Berman SH, Soni RK, Mansilla-Soto J, Eyquem J, Hamieh M, et al. Integrating proteomics and transcriptomics for systematic combinatorial chimeric Antigen receptor therapy of AML. Cancer Cell. 2017;32:506 – 19.e5.
Bosse KR, Raman P, Zhu Z, Lane M, Martinez D, Heitzeneder S, et al. Identification of GPC2 as an oncoprotein and candidate immunotherapeutic target in high-risk neuroblastoma. Cancer Cell. 2017;32:295–309.e12.
Han X, Zhou Z, Fei L, Sun H, Wang R, Chen Y, et al. Construction of a human cell landscape at single-cell level. Nature. 2020;581:303–9.
Regev A, Teichmann SA, Lander ES, Amit I, Benoist C, Birney E, et al. The Human Cell Atlas. Elife. 2017;6.
He S, Wang LH, Liu Y, Li YQ, Chen HT, Xu JH, et al. Single-cell transcriptome profiling of an adult human cell atlas of 15 major organs. Genome Biol. 2020;21:294.
Jones RC, Karkanias J, Krasnow MA, Pisco AO, Quake SR, Salzman J, et al. The Tabula Sapiens: a multiple-organ, single-cell transcriptomic atlas of humans. Science. 2022;376:eabl4896.
Eraslan G, Drokhlyansky E, Anand S, Fiskin E, Subramanian A, Slyper M, et al. Single-nucleus cross-tissue molecular reference maps toward understanding disease gene function. Science. 2022;376:eabl4290.
Suo C, Dann E, Goh I, Jardine L, Kleshchevnikov V, Park JE, et al. Mapping the developing human immune system across organs. Science. 2022;376:eabo0510.
Domínguez Conde C, Xu C, Jarvis LB, Rainbow DB, Wells SB, Gomes T, et al. Cross-tissue immune cell analysis reveals tissue-specific features in humans. Science. 2022;376:eabl5197.
Kwon J, Kang J, Jo A, Seo K, An D, Baykan MY, et al. Single-cell mapping of combinatorial target antigens for CAR switches using logic gates. Nat Biotechnol. 2023.
Lähnemann D, Köster J, Szczurek E, McCarthy DJ, Hicks SC, Robinson MD, et al. Eleven grand challenges in single-cell data science. Genome Biol. 2020;21:31.
Lian TY, Yan Y, Ding D, Ma YJ, Zhang X, Jing ZC. Building a modern six-dimensional biobank fosters the future of precision medicine. Sci Bull (Beijing). 2022;67:2490–3.
Acknowledgements
XX, SH, XW, Yu Wang, Yajing Wang, CF, Yazhuo Wang, SC, TL, RC and YZ are the Small Small bird (SSb) group members and acknowledge the strong support from SSb. We also sincerely appreciate the continuous support from our best friend Xinyi Xiao.
Funding
This research was supported by grants from the National Key Research and Development Program of China (No.2017YFA0105503), Frontier Research Program of Guangzhou Regenerative Medicine and Health Guangdong Laboratory (No.2018GZR110105014), Basic and Applied Basic Research Fund of Guangdong Province (No. 2019A1515111113), Special Funds for the Cultivation of Guangdong College Students’ Scientific and Technological Innovation (No. pdjh2021b0101 and No. pdjh2022b0096), National Students’ Platform for Innovation and Entrepreneurship Training Program of China (No. 202212121023) and Guangdong Students’ Platform for Innovation and Entrepreneurship Training Program (No. S202212121074).
Author information
Authors and Affiliations
Contributions
XX contributed to conception and manuscript design.YL supervised the manuscript. SH, XW, Yu Wang, XX, Yajing Wang and CF drafted the manuscript. SH, XW, Yu Wang, Yajing Wang, Yazhuo Wang and CF prepared the tables and figures. XW and Yu Wang collected the related references. XX, SH, XW, Yu Wang, CF, SC, TL, RC, YZ participated in the revision of the manuscript. XX and YL were involved in funding acquisition. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Huang, S., Wang, X., Wang, Y. et al. Deciphering and advancing CAR T-cell therapy with single-cell sequencing technologies. Mol Cancer 22, 80 (2023). https://doi.org/10.1186/s12943-023-01783-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12943-023-01783-1