
Abstract
Background
Artesunate–amodiaquine (AS–AQ) and artemether–lumefantrine (AL) are the currently recommended first-and second-line therapies for uncomplicated Plasmodium falciparum infections in Chad. This study assessed the efficacy of these artemisinin-based combinations, proportion of day 3 positive patients, proportions of molecular markers associated with P. falciparum resistance to anti-malarial drugs and variable performance of HRP2-based malaria rapid diagnostic tests (RDTs).
Methods
A single-arm prospective study assessing the efficacy of AS–AQ and AL at three sites (Doba, Kelo and Koyom) was conducted between November 2020 to January 2021. Febrile children aged 6 to 59 months with confirmed uncomplicated P. falciparum infection were enrolled sequentially first to AS–AQ and then AL at each site and followed up for 28 days. The primary endpoint was PCR-adjusted adequate clinical and parasitological response (ACPR). Samples collected on day 0 were analysed for mutations in pfkelch13, pfcrt, pfmdr-1, pfdhfr, pfdhps genes and deletions in pfhrp2/pfhrp3 genes.
Results
By the end of 28-day follow-up, per-protocol PCR corrected ACPR of 97.8% (CI 95% 88.2–100) in Kelo and 100% in Doba and Kayoma were observed among AL treated patients. For ASAQ, 100% ACPR was found in all sites. All, but one patient, did not have parasites detected on day 3. Out of the 215 day 0 samples, 96.7% showed pfkelch13 wild type allele. Seven isolates carried nonsynonymous mutations not known to be associated artemisinin partial resistance (ART-R). Most of samples had a pfcrt wild type allele (79% to 89%). The most prevalent pfmdr-1 allele detected was the single mutant 184F (51.2%). For pfdhfr and pfdhps mutations, the quintuple mutant allele N51I/C59R/S108N + G437A/540E responsible for SP treatment failures in adults and children was not detected. Single deletion in the pfhrp2 and pfhrp3 gene were detected in 10/215 (4.7%) and 2/215 (0.9%), respectively. Dual pfhrp2/pfhrp3 deletions, potentially threatening the efficacy of HRP2-based RDTs, were observed in 5/215 (2.3%) isolates.
Conclusion
The results of this study confirm that AS–AQ and AL treatments are highly efficacious in study areas in Chad. The absence of known pfkelch13 mutations in the study sites and the high parasite clearance rate at day 3 suggest the absence of ART-R. The absence of pfdhfr/pfdhps quintuple or sextuple (quintuple + 581G) mutant supports the continued use of SP for IPTp during pregnancy. The presence of parasites with dual pfhrp2/pfhrp3 deletions, potentially threatening the efficacy of HRP2-based RDTs, warrants the continued surveillance.
Trial registration ACTRN12622001476729
Similar content being viewed by others
Background
Provision of effective treatment, using artemisinin-based combination therapy (ACT), fast-acting artemisinin derivatives partnered with longer-acting partner drugs, is a critical component of recommended malaria interventions [1]. The World Health Organization (WHO) recommends artemether–lumefantrine (AL), artesunate–amodiaquine (AS–AQ), artesunate–mefloquine (AS–MQ), dihydroartemisinin–piperaquine (DHA–PPQ), artesunate–sulfadoxine/pyrimethamine (AS–SP) and artesunate–pyronaridine (AS–PY) for the treatment of uncomplicated falciparum malaria infection [2]. The WHO also recommends intermittent preventive treatment with SP during pregnancy (IPTp-SP) and Seasonal Malaria Chemoprevention (SMC) using SP + AQ to protect children during the season of greatest risk in falciparum endemic areas in Africa [1].
Resistance of Plasmodium falciparum to anti-malarial drugs remains a threat to effective case management and undermines the global efforts to control and eliminate the burden of malaria. Artemisinin partial resistance (ART-R), defined as delayed clearance of parasitaemia (parasite half-life > 5 h or day 3 parasitaemia) following ACT or artemisinin monotherapy, emerged first in Southeast Asia [3, 4]. Non-synonymous mutations in the propeller region of the P. falciparum kelch13 (pfkelch13) gene have been documented to be associated with ART-R [5]. Recent emergence and expansion of validated pfkelch13 mutations in Rwanda (R561H), Uganda (A675V or C469Y) associated with delayed parasite clearance are of great concern [6, 7] and calls for frequent monitoring of ACT efficacy, including clearance of parasitaemia, and pfkelch13 mutations in Africa, as recommended by the WHO [8].
The SP combination inhibits the enzymes dihydrofolate reductase (DHFR) and dihydropteroate synthase (DHPS), which are involved in the folate pathway of nucleic acid synthesis of the parasite. Unfortunately, mutations in the parasite genes pfdhfr (codons 51, 59, 108 and 164) and pfdhps (codons 437, 540, 581 and 613) were found to confer resistance to pyrimethamine and sulfadoxine, respectively [9,10,11]. The pfdhfr triple mutant (N51I, C59R, S108N) combined with the pfdhps double mutant (A437G, K540E) have been associated with in vivo resistance to SP [12, 13] and the sextuple mutant (quintuple mutant with an additional mutation in codon 581 of the pfdhps gene) with clinical failure and loss of SP protection when the prevalence of this mutant exceeds 10% [14,15,16,17]. However, several studies suggest that although the use of IPTp-SP often results in the selection and increased prevalence of resistance-associated genetic mutations, this does not necessarily lead to a decrease in malaria prevention efficacy [18].
In Chad, malaria accounted for 43% of out-patient attendances, 44% of hospitalizations and 60% of hospital deaths among children in 2018 [19]. Since 2005, AS–AQ and AL are recommended first and second line treatments, respectively, for the treatment of uncomplicated P. falciparum malaria. To control the burden of malaria in the country, IPTp-SP is a part of the recommended interventions to control malaria among the pregnant women [20]. A study conducted in 2015 reported high cure rate (100%) with AS–AQ and absence of mutation in pfkelch13 gene among under-five children with uncomplicated falciparum malaria [21].
The aim of the current study was to monitor the therapeutic efficacy of AS–AQ and AL for the treatment of uncomplicated P. falciparum malaria in children and assess the proportion of day3 positive patients. Mutations in pfkelch13, pfcrt, pfmdr-1, pfdhfr, pfdhps associated with P. falciparum resistance to anti-malarial drugs along with deletions in pfhrp2/pfhrp3 involved in variable performance of HRP2-based malaria rapid diagnostic tests (RDTs) were also investigated in isolates collected prior anti-malarial treatment and those from patients experiencing recurrences.
Methods
Study design and areas
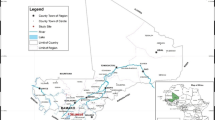
The study used one arm prospective design to assess the efficacy of AL and AS–AQ for the treatment of uncomplicated falciparum malaria among children aged 6 to 59 months from November 2020 to January 2021. The study sites were the (i) Gaki health center in Doba town (8° 40′ N, 16° 51′ E) in Pende department of Logone Oriental, (ii) Hindina health center in Kelo town in West Tandjile department of Tandjile province and (iii) Boubou health center in Koyom town in Mayo Boneye department of Mayo Kebbi Est province. Malaria transmission in the study areas occurs from June to October and P. falciparum is the predominant species [19] (Fig. 1).

Location of the study sites, Chad, 2022–2023
Sample size estimation
Sample size estimates assumed 5% of treatment failure with ASAQ or AL treatment in the study sites. At a confidence level of 95% and an estimate precision of 10%, a minimum sample size of 50 patients was required. With a 20% increase to allow for loss to follow-up and withdrawals during the 28-days of follow-up, 60 patients were targeted per drug and per site.
Recruitment and treatment procedures
Children attending the study health clinics between November 2020 and January 2021 were screened for eligibility criteria, and were enrolled, after obtaining consent from parents/guardians, if they met the study inclusion criteria: age 6 to 59 months, axillary temperature of ≥ 37.5 °C or history of fever during the past 24 h; P. falciparum mono-infection with parasitaemia of 2000 to 200,000 asexual parasites/µl by microscopy. Children presenting with signs or symptoms of severe falciparum malaria, non-falciparum species or severe malnutrition, third party administration of anti-malarial drugs and febrile conditions due to diseases other than malaria, were not recruited and received appropriate treatment and care according to national guidelines. Children who developed danger signs (convulsions, lethargy, inability to drink or breast feed, repeated vomiting, inability to stand/sit due to weakness) or severe malaria were referred to receive parenteral artesunate with dose regiments according to the national treatment guidelines.
Children were enrolled sequentially first to AS–AQ (Sanofi, lot number 8MA153) and then AL (Cipla, lot number ID01029) at each site. Children treated with AS–AQ received artesunate 4 mg/kg + amodiaquine 10 mg/kg once daily over 3 consecutive days. ASAQ tablets were given based on the following weight bands: one tablet of 25 artesunate + 67.5 amodiaquine for 4.5 to < 9 kg body weight, one table of 50 artesunate + 135 amodiaquine for 9–< 18 kg body weight, one tablet of 100 artesunate + 270 amodiaquine for 18 to < 36 kg body weight. The AL treated group received twice daily doses of the ACT for 3 days based on weight bands: one tablet for 5–14 kg and two tablets for 15–24 kg. Artemether–lumefantrine was administered with milk when possible. All treatment doses were given under direct observation by the study team and patients were observed for 30 min. If the first dose was vomited, treatment was re-administered. If vomited second time, the child was given parenteral artesunate according to the national treatment guidelines and the patient was withdrawn from the study. WHO/HQ provided the study drugs. Enrolled children were monitored for 28 days, and clinical and parasitological assessment were done at scheduled visits (days 1, 2, 3, 7, 14, 21 and 28) or unscheduled visits if children felt ill. Patients who did not return for follow-up were visited at home.
Malaria microscopy
Thick and thin blood smears were obtained through finger prick to detect the presence of P. falciparum and estimate parasite density at day 0 (before inclusion) and at each scheduled (days 1, 3, 7, 14, 21 and 28) or unscheduled visits. The blood smears were dried, Giemsa-stained and examined under light microscopy at 100× magnification. Parasite count and density (per μl blood) were determined using WHO procedure [22]. If the two readings were discordant, in terms of difference in parasite positivity, parasite species or parasite density above 50%, the slide was read by a third microscopist.
Treatment responses
The treatment outcomes by day 28 were classified, using on the WHO 2009 protocol [22], as adequate clinical and parasitological response (ACPR), early treatment failure (ETF), late parasitological failure (LPF), and late clinical failure (LCF). PCR analysis was performed to distinguish between a true recrudescence due to treatment failure and episodes of reinfection.
Parasite genotyping to differentiate recrudescence from re-infection
Filter paper blood samples collected from each patient on day 0 and on the day of parasite recurrence (from day 7 onward) were stored in individual plastic bags with desiccant and protected from light, moisture and extreme temperature until analysis. Each dried blood spot was cut out sterilely and placed in an Eppendorf tube. Parasite DNA was extracted by using the 96-well format protocol developed by Zainabadi et al. [23]. The eluted DNA was then quantified by fluorometric quantitation (Qubit, Thermo Fischer), adjusted to 20 ng/μl and stored at − 20 °C for later use. Paired DNA from patients with recurrent parasites (day-0 and day of recurrence) were genotyped using nested polymerase chain reaction (PCR) targeting the highly polymorphic genes msp1, msp2 and the microsatellite marker poly-α [24], according to the recent WHO recommendation [25]. All markers were assessed systematically. The fragment sizes were estimated by capillary electrophoresis (Fragment analyzer, Agilent) and the cut-off settings for PCR artefacts and stutter peaks was defined for peaks < 10% of the low and upper control bands. The bins used to define a match were ± 10 bp for msp1/msp2, and ± 5 bp for poly-α. Genotyping data were compared with the former genotyping approach assessing msp1, msp2 and glurp [26]. The bins used to define a match was ± 20 bp for glurp. The WHO/MMV decision algorithm was used to define PCR-adjusted clinical efficacy rates. Recrudescence was defined as a genotype that had already been detected in the blood sample taken before treatment (i.e., at least one allele is shared at day 0 and day of parasite recurrence at all three loci). A new infection was defined as the absence of a shared allele between day 0 and day of parasite recurrence at any of the three loci.
Molecular markers
Day 0 DNA were analysed for the presence of point mutations in the pfkelch13 gene (codons 430–720) associated with ART-R [5], the pfcrt (at codons 72–76, 93, 97, 145, 218, 343, 350 and 353) and pfmdr-1 (at codons 86, 184, 1034, 1042 and 1246) genes associated or suspected to be associated with 4-aminoquinolines and aminoalcohol resistance [27], the pfdhfr (at codons 51, 59, 108, 164) and pfdhps (at codons 431, 436, 437, 540, 581, 613) genes linked to pyrimethamine and sulfadoxine resistance [27]. hrp2 and hrp3 deletions that can cause false-negative results with HRP2-based rapid diagnostic tests (RDTs) were also screened [28].
Amplicons from targeted sequences were generated using multiplexing nested PCR assays using indexed primers that themselves contain specific tags (barcodes) consisting of individual 8-base indices, specific to the sample and adapter sequences (14 or 15 bases) that allow the final PCR product to bind to the sequencing flow cell (Table 1). A total of 4 μl of PCR reactions from each sample were mixed in one pool (96 samples) to increase the sample volume and minimize sample quantity for downstream protocol steps. For each pool, the amplicons were then purified with the AMPure XP beads (Beckman Coulter), according to manufacturer’s protocol to eliminate dNTPs, salts, primers and primer dimers. The quality of purified PCR products was assessed by analysing eluates containing the purified amplicons on a Fragment analyzer (Agilent). DNA concentration of pooled fragments was assessed by fluorometric quantitation (Qubit, Thermo Fischer). The pooled libraries were denatured with NaOH to a final concentration of 0.1 N, diluted with hybridization buffer before running the sequencing. Sequencing was performed using the MiSeq v2 reagents using the 300-cycle kit (Illumina) according to the manufacturer’s recommendations. The raw sequences were demultiplexed and quality trimmed at a phred score of 30. The primer sequences were trimmed from the 5′-end of the sequences, to avoid primer bias in the sequenced fragments. Base calling was performed by comparing reads with a custom database, consisting of the 3D7 reference sequence. Bioinformatic analysis were performed using the CLC Genomics Workbench 22 software (Qiagen). Laboratory reference parasite strains (Dd2, 7G8, HB3 and a Cambodian culture-adapted strain), with known alleles in each gene were used as controls.
Ethical considerations
Before study implementation, it obtained approval from the National Ethics Committee of Chad and the WHO ERC. The study objectives, its benefits and potential risks and its procedures were explained to the parents/guardians of the potential study children. Children were only included in the study if their parents or guardians gave written consent. If the parent or guardian cannot read or write, a witness chosen by the parent/guardian of the child consigned the consent form.
Statistical analysis
WHO excel software (WHO excel software programme (http://www.who.int/malaria/publications/atoz/9789241597531/en/) was used to double enter and analyse the data. Both per-protocol and Kaplan Meier analysis were used to evaluate the treatment outcomes. Patients who were lost or withdrawn or had new infection during the follow-up were excluded from the per-protocol analysis while these cases were censored last day the patient was follow-up, withdrawn or experienced new infection. Patients with indeterminate PCR results were excluded from both per-protocol and KM analysis. Percentages, mean, standard deviation and range were presented. Categorical variables were compared using Chi-square or Fisher exact test, and t-test for continuous variables. A p-value of < 0.05 was considered statistically significant.
Results
Clinical outcomes
Table 2 summarizes the baseline characteristics of the study children by site and drug. In total, 114 and 101 children were recruited for AL and AS–AQ, respectively. Due to the late start of the study in the transmission season, the targeted sample per drug (n = 60) in each site was not reached. There was no significant difference in the baseline patient profile between the sites and between treatment groups.
At the end of the 28 days follow up period, 111 (28 in Doba, 48 in Kelo and 35 in Kayoma) and 94 (30 in Doba, 41 in Kelo and 33 in Kayoma) patients in the AL and AS–AQ groups, respectively, reached the desired endpoints. Three patients in the AL and 7 in the AS–AQ groups were lost to follow-up or withdrawn, and consequently were excluded from the per-protocol analysis.
Per-protocol PCR uncorrected ACPR rate for AL were 100% (CI 95% 87.7–100) in Doba, 91.7% (CI 95% 80.0–97.7) in Kelo and 97.1% (CI 95% 85.1–99.9) in Kayoma, with an overall rate of 95.5% (CI 95% 89.8–98.5) (Table 3). For the AS–AQ treated groups, PCR uncorrected ACPR of 96.7% (CI 95% 82.8–99.9) in Doba, 100% in Kelo (CI 95% 88.8–100) and Kayoma (CI 95% 89.4–100) were observed with overall cure rate of 98.9% (CI 95% 94.2–100).
After PCR correction, ACPR rates of 97.8% (CI 95% 88.2–99.9) in Kelo and 100% in Doba (CI 95% 87.7–100) and in Kayoma (CI 95% 89.7–100), with an overall cure rate of 99.1% (CI 95% 94.9–100) were observed among AL treated patients (Table 4). For AS–AQ treated children, PCR corrected ACPR rate of 100% was observed in Doba (CI 95% 88.1–100), Kelo (CI 95% 88.8–100) and Kayoma (CI 95% 89.4–100), with overall cure rate of 100% (CI 95% 96.1–100). All, but one patient, were parasite free on day 3 (1/210, 0.5%).
Both msp1/mps2/polyα and msp1/msp2/glurp genotyping methods gave similar results, as reinfection in five recurrent infections and one recurrent case was classified as recrudescence. The unique recrudescence case (AS556) was observed at day 7. The patient was a female of 4 years aged treated with AL. The doses given were supervised and were fully administered without side effects.
Molecular markers
Out of the 215 day 0 samples analysed for the presence of polymorphism in pfkelch13 gene, majority (96.7%) showed wild type allele and seven carried nonsynonymous mutations: the A578S was observed in 3 isolates (once in each site) known to not confer in vitro ART-R and the N489Y that is a non-validated pfkelch13 mutation in 4 isolates (2 in Kelo and 2 in Koyom) (Table 5).
For pfcrt, eight alleles were detected, most of which were wild type (79% to 89%). The second most frequent alleles were alleles carrying the 74I/75E/76T mutations (4.7%) and the 74I/75E/76T/356T mutations (6.5%). The other mutations observed were at codons 145 (I> M or V). The most prevalent pfmdr-1 allele detect was the single mutant 184F (51.2%, ranging from 48.6 to 60.9%) followed by the wild type allele (32.1%), double mutants 86Y/184F (8.8%) and the 184F/1034C (3.7%). The frequency of the other alleles was < 2%.
The triple mutant 51I/59R/108N was the most frequent pfdhfr allele (81.4% ranging from 79.7 to 84.3%), followed by the double mutant 59R/108N (7.0%), the wild type allele (4.7%) and the double mutant 51I/108N (3.3%). The frequency of the other alleles was < 2% and the quadruple mutant 51I/59R/108N/164L was absent. For pfdhps, the most frequent allele was the double mutant 436A/437A (54.7%, ranging from 47.1 to 62.5%), followed by the single mutant 437K (16.8%), the triple mutant 431V/436A/437K (11.7%), the double mutant 436A/437K (9.8%) and the triple mutant 431V/436A/437A (2.8%). The frequency of the other alleles was < 2%. The wild type allele and the 581G mutation were not detected. A significant difference in proportion were observed between sites (p = 0.04).
The most frequent pfdhfr/pfdhps alleles were the quintuple 51I/59R/108N/436A/437A mutant (44.7%), the quadruple 51I/59R/108N/437K (12.6%) mutant, the sextuple 51I/59R/108N/431V/436A/437K and 51I/59R/108N/431V/436A/437A mutants (11.2% and 2.8%, respectively); Lower proportions were observed for the quadruple 59R/108N/436A/437A (4.2%) mutant and the double 436A/437A mutant (3.3%). The proportions of the other alleles were < 2%. The quintuple mutant alleles N51I/C59R/S108N + G437A/540E responsible for SP treatment failures in adults and children (but still effective for use in intermittent preventive treatment in pregnancy) was not detected (Table 6).
Genetic deletions in the hrp2 and hrp3 genes in Chadian P. falciparum population were also sought as there are two important factors responsible for variable performance of malaria Rapid Diagnostic Tests. For this analysis, 215 day 0 samples were tested. Single deletion in the hrp2 and hrp3 gene were detected in 10/215 (4.7%, CI 95% 2.2–8.5) and 2/215 (0.9%, CI 95% 0.1–3.3), respectively. Dual hrp2/hrp3 deletions, potentially threatening the efficacy of HRP2-based RDTs, were observed in 5/215 (2.3%, CI 95% 0.7–5.4) isolates in all sites: 2/64 (3.1%, CI 95% 0.4–1.1) in Doba, 2/81 (1.2%, CI 95% 0.03–6.8) in Kelo and 1/70 (1.4%, CI 95% 0.03–7.9) in Koyom.
Discussion
The study revealed high cure rate (100%) of first-line treatment (AS–AQ) for uncomplicated malaria in children, confirming that ACT remains highly efficacious after its introduction in 2005. A similar high cure rate with AS–AQ was observed in 2015 [21]. The study also showed high efficacy (> 98%) of the second-line treatment (AL), which was evaluated for the first time since its recommendation in 2005. These findings are similar to the efficacy rates observed in neighboring Sudan [29], Central African Republic [30, 31], Cameroon [31], Nigeria [32], Niger [33, 34], and other West African countries [35,36,37,38,39,40,41,42]. A study in Burkina Faso reported a PCR-corrected cure rate of AL and DHA–PPQ of less than 80% at two sites [43]. One of the limitations of the study was that children weighing 5–9 kg received only half the intended doses. Other recent studies also reported AL cure rates below 90% in Angola [44] and the Democratic Republic of Congo [45] and may raise concern about the efficacy of the partner drug, lumefantrine. However, these studies used microsatellite-based PCR correction and a Bayesian algorithm for analysis, a methodological deviation from the standard WHO genotyping protocol, that has been associated with higher treatment failure in areas of high transmission [25]. Until a robust and validated tool to distinguish recrudescence from new infection is developed, therapeutic efficacy studies based on the standard WHO protocol [25] should be used to capture temporal comparability of efficacy data within and across countries.
Molecular analysis for polymorphism in the pfkelch13 gene did not reveal any mutations known to be associated with delayed parasite clearance in Africa like the R561H, A675V or C469Y variants [6, 7]. Together with the high parasite clearance rate at day 3 (99.5%), this supports the absence of ART-R in the study areas, which is consistent with previous study [21]. However, the recent reports of validated pfkelch13 mutations with delayed parasite clearance after ACT treatment in Rwanda [7], and Uganda [6] underscore the importance of continuing to monitor anti-malarial drug efficacy and resistance of artemisinins and partner drugs.
Molecular analysis of the current study revealed a low frequency of the pfdhfr wild-type allele (4.7%) with a high frequency of the triple mutant (51I/59R/108N) (81.4%) and the absence of the quintuple mutant alleles (N51I/C59R/S108N + G437A/540E). In contrast, a pfdhfr wild type of 53.3% and a very low frequency (3.3%) of the triple mutant (51I/59R/108N) were detected in the Pala West region in 2015 [21]. This difference could be due to either spatial or temporal differences, as the two studies were conducted in different areas and at different times. Overall, the absence of the quintuple and quintuple + 581G sextuple mutations, which have been associated with reduced efficacy of SP [12, 13] and decreased effectiveness of IPT-SP, respectively, in pregnant women [17, 21, 46], argues for the continued use of IPTp-SP. Studies from other West African countries reported the absence or very low frequency of the quintuple mutation [47,48,49]. Notably, this study reports for the first time the pfdhps I431V mutation, which was observed in proportions ranging from 12.3% (Kelo) to 17.1% (Doba) in the three study sites. This confirms previous molecular studies showing high levels of this mutation in Nigeria, Cameroon and other parts of West Africa. It is not yet clear whether this mutation interferes with the binding of sulfadoxine to its active site, thereby reducing the susceptibility of P. falciparum to the drug. However, epidemiological data show that the molecular repertoire of pfdhps in West Africa is clearly different from that in East Africa, where the K540E mutation remains low, while the I431V mutation appears to be emerging. More information is needed to assess the impact of haplotypes containing the I431V mutation on the protective efficacy of IPTp.
Most of the parasites carried the pfcrt wild type allele (79% to 89%). The proportion of pfcrt variant associated with resistance to chloroquine (CVIET, 11.2%) remains similar to previous report in 2016 (8/191, 4.2%) [50] and relatively low compared to other regions such as Ethiopia [51], Democratic Republic of the Congo [52], Equatorial Guinea [53], and Liberia [38]. For pfmdr-1 gene, the single 184F mutation, which is suspected to be involved in recrudescent infections after AL treatment, was highly frequent in Chad as usually reported in numerous African settings such as Ghana [54], Uganda [55], Madagascar [56], Tanzania [57], and Equatorial Guinea [58].
Although, the study did not follow the recommended master protocol developed by WHO to guide surveillance and biobanking of pfhrp2/3 gene deletions in malaria endemic countries, this study provides, for the first time, the overall prevalence of hrp2, hrp3 and dual hrp2/3 gene deletions estimated in some sites in Chad and baseline data for future surveys aimed at determining the trend in the frequency of hrp2/3 gene deletions. The proportion of parasites with dual hrp2/3 deletions found here (2.3%) is below the WHO 5% prevalence criterion defined for switching away from HRP2-based RDTs for malaria case management [59]. This frequency remains well below the proportions detected in the Horn of Africa such as Eritrea, Ethiopia, Djibouti, Sudan and South Sudan [60,61,62,63] and similar to proportions usually observed in African countries.
A limitation of the study is that the target sample size per drug at each site to detect treatment failure of 5% at a 95% CI and power of 10% could not be achieved because the study was conducted late in the malaria transmission season. Consequently, this study was underpowered to rule out low efficacy of the drugs tested.”
Conclusion
The results of this study confirm that first-line (AS–AQ) and second line (AL) treatments are highly efficacious in the treatment of uncomplicated falciparum infections in Chad, with a cure rate of over 98%. The absence of known pfkelch13 mutations in the study sites and the high parasite clearance rate at day 3 confirm the absence of ART-R. The absence of pfdhfr/pfdhps quintuple and quintuple + 581G sextuple mutations supports the continued use of SP for IPTp during pregnancy. Routine monitoring of anti-malarial drug efficacy and resistance should continue in Chad to detect any change in the susceptibility of the parasite populations.
Availability of data and materials
The dataset used in this study is available and can be shared upon reasonable request to NMCP through the corresponding author.
Abbreviations
- ACPR:
-
Adequate clinical and parasitological response
- ACTs:
-
Artemisinin-based combination therapies
- AL:
-
Artemether–lumefantrine
- ART-R:
-
Artemisinin resistance
- AS–AQ:
-
Artesunate–amodiaquine
- AS–MQ:
-
Artesunate–mefloquine
- AS–PY:
-
Artesunate–pyronaridine
- AS–SP:
-
Artesunate–sulfadoxine/pyrimethamine
- pfdhps :
-
Plasmodium falciparum dihydropteroate synthetase
- DHA–PPQ:
-
Dihydroartemisinin–piperaquine
- ETF:
-
Early treatment failure
- LCF:
-
Late clinical failure
- LPF:
-
Late parasitological failure
- msp1 :
-
Merozoite surface proteins 1
- msp2 :
-
Merozoite surface proteins 2
- glurp :
-
Glutamate rich-protein
- PCR:
-
Polymerase chain reaction
- pfdhfr :
-
Plasmodium falciparum dihydrofolate reductase-thymidylate synthase
- pfmdr-1 :
-
Plasmodium falciparum multi drug resistance 1
- pfkelch13 :
-
Plasmodium falciparum kelch13
- WHO:
-
World Health Organization
References
WHO. Guidelines for malaria. Geneva: World Health Organization; 2023.
WHO. World malaria Report 2021. Geneva: World Health Organization; 2021.
Ashley EA, Dhorda M, Fairhurst RM, Amaratunga C, Lim P, Suon S, et al. Spread of artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med. 2014;371:411–23.
Dondorp AM, Nosten F, Yi P, Das D, Phyo AP, Tarning J, et al. Artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med. 2009;361:455–67.
Ariey F, Witkowski B, Amaratunga C, Beghain J, Langlois AC, Khim N, et al. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature. 2014;505:50–5.
Balikagala B, Fukuda N, Ikeda M, Katuro OT, Tachibana SI, Yamauchi M, et al. Evidence of artemisinin-resistant malaria in Africa. N Engl J Med. 2021;385:1163–71.
Uwimana A, Legrand E, Stokes BH, Ndikumana JM, Warsame M, Umulisa N, et al. Emergence and clonal expansion of in vitro artemisinin-resistant Plasmodium falciparum kelch13 R561H mutant parasites in Rwanda. Nat Med. 2020;26:1602–8.
WHO. Report on antimalarial drug efficacy, resistance and response: 10 years of surveillance (2010–2019). Geneva: World Health Organization; 2020.
Lozovsky ER, Chookajorn T, Brown KM, Imwong M, Shaw PJ, Kamchonwongpaisan S, et al. Stepwise acquisition of pyrimethamine resistance in the malaria parasite. Proc Natl Acad Sci USA. 2009;106:12025–30.
Plowe CV, Cortese JF, Djimde A, Nwanyanwu OC, Watkins WM, Winstanley PA, et al. Mutations in Plasmodium falciparum dihydrofolate reductase and dihydropteroate synthase and epidemiologic patterns of pyrimethamine-sulfadoxine use and resistance. J Infect Dis. 1997;176:1590–6.
Triglia T, Menting JG, Wilson C, Cowman AF. Mutations in dihydropteroate synthase are responsible for sulfone and sulfonamide resistance in Plasmodium falciparum. Proc Natl Acad Sci USA. 1997;94:13944–9.
Happi CT, Gbotosho GO, Folarin OA, Akinboye DO, Yusuf BO, Ebong OO, et al. Polymorphisms in Plasmodium falciparum dhfr and dhps genes and age related in vivo sulfadoxine-pyrimethamine resistance in malaria-infected patients from Nigeria. Acta Trop. 2005;95:183–93.
Kublin JG, Dzinjalamala FK, Kamwendo DD, Malkin EM, Cortese JF, Martino LM, et al. Molecular markers for failure of sulfadoxine-pyrimethamine and chlorproguanil-dapsone treatment of Plasmodium falciparum malaria. J Infect Dis. 2002;185:380–8.
Braun V, Rempis E, Schnack A, Decker S, Rubaihayo J, Tumwesigye NM, et al. Lack of effect of intermittent preventive treatment for malaria in pregnancy and intense drug resistance in western Uganda. Malar J. 2015;14:372.
Gutman J, Kalilani L, Taylor S, Zhou Z, Wiegand RE, Thwai KL, et al. The A581G mutation in the gene encoding Plasmodium falciparum dihydropteroate synthetase reduces the effectiveness of sulfadoxine-pyrimethamine preventive therapy in Malawian pregnant women. J Infect Dis. 2015;211:1997–2005.
Harrington WE, Mutabingwa TK, Kabyemela E, Fried M, Duffy PE. Intermittent treatment to prevent pregnancy malaria does not confer benefit in an area of widespread drug resistance. Clin Infect Dis. 2011;53:224–30.
Harrington WE, Mutabingwa TK, Muehlenbachs A, Sorensen B, Bolla MC, Fried M, et al. Competitive facilitation of drug-resistant Plasmodium falciparum malaria parasites in pregnant women who receive preventive treatment. Proc Natl Acad Sci USA. 2009;106:9027–32.
Plowe CV. Malaria chemoprevention and drug resistance: a review of the literature and policy implications. Malar J. 2022;21:104.
Chad National Strategic Plan for Malaria Control 2019–2023. https://scholar.google.com/citations?user=g5-0ZQwAAAAJ)
Ministère de la Sante Publique République du Tchad. Programme National de Lutte Contre le Paludisme. N’Djamena, Chad, 2014.
Issa MS, Ako AB, Kerah HC, Djimadoum M, Coulibaly B, Mbaitoloum MD, et al. Therapeutic efficacy of artesunate-amodiaquine and polymorphism of Plasmodium falciparum k13-propeller gene in Pala (Tchad). Int J Open Access Clin Trials 2017;1(1):1–6. https://symbiosisonlinepublishing.com/clinical-trials/clinical-trials04.pdf.
WHO. Methods for surveillance of antimalarial drug efficacy. Geneva: World Health Organisation; 2009.
Zainabadi K, Nyunt MM, Plowe CV. An improved nucleic acid extraction method from dried blood spots for amplification of Plasmodium falciparum kelch13 for detection of artemisinin resistance. Malar J. 2019;18:192.
Anderson TJ, Su XZ, Bockarie M, Lagog M, Day KP. Twelve microsatellite markers for characterization of Plasmodium falciparum from finger-prick blood samples. Parasitology. 1999;119:113–25.
World Health Organization. Informal consultation on methodology to distinguish reinfection from recrudescence in high malaria transmission areas, 7–18 May 2021. Geneva: World Health Organization; 2021.
WHO. Methods and techniques for clinical trials on antimalarial drug efficacy: genotyping to identify parasite populations: informal consultation organized by the Medicines for Malaria Venture and cosponsored by the World Health Organization, 29–31 May 2007, Amsterdam, The Netherlands. Geneva: World Health Organization; 2007.
Menard D, Dondorp A. Antimalarial drug resistance: a threat to malaria elimination. Cold Spring Harb Perspect Med. 2017;7:a025619.
WHO. False-negative RDT results and implications of new reports of P. falciparum histidine-rich protein 2/3 gene deletions. Geneva: World Health Organization; 2019.
Adeel AA, Elnour FA, Elmardi KA, Abd-Elmajid MB, Elhelo MM, Ali MS, et al. High efficacy of artemether–lumefantrine and declining efficacy of artesunate + sulfadoxine–pyrimethamine against Plasmodium falciparum in Sudan (2010–2015): evidence from in vivo and molecular marker studies. Malar J. 2016;15:285.
Nambei WS, Biago U, Balizou O, Pounguinza S, Moyen M, Ndoua C, et al. [Monitoring the efficacy of artemether–lumefantrine in the treatment of uncomplicated Plasmodium falciparum malaria by kelch 13 gene mutations in Bangui, Central African Republic]. Med Trop Sante Int. 2021;1:82 (in French).
Niba PTN, Nji AM, Ali IM, Akam LF, Dongmo CH, Chedjou JPK, et al. Effectiveness and safety of artesunate-amodiaquine versus artemether–lumefantrine for home-based treatment of uncomplicated Plasmodium falciparum malaria among children 6–120 months in Yaounde, Cameroon: a randomized trial. BMC Infect Dis. 2022;22:166.
Ebenebe JC, Ntadom G, Ambe J, Wammanda R, Jiya N, Finomo F, et al. Efficacy of artemisinin-based combination treatments of uncomplicated falciparum malaria in under-five-year-old Nigerian children ten years following adoption as first-line antimalarials. Am J Trop Med Hyg. 2018;99:649–64.
Grandesso F, Guindo O, Woi Messe L, Makarimi R, Traore A, Dama S, et al. Efficacy of artesunate-amodiaquine, dihydroartemisinin-piperaquine and artemether–lumefantrine for the treatment of uncomplicated Plasmodium falciparum malaria in Maradi, Niger. Malar J. 2018;17:52.
Ibrahima I, Laminou IM, Adehossi E, Maman D, Boureima S, Harouna HK, et al. Safety and efficacy of artemether–lumefantrine and artesunate-amodiaquine in Niger. Bull Soc Pathol Exot. 2020;113:17–23 (in French).
Abuaku B, Duah-Quashie NO, Quaye L, Matrevi SA, Quashie N, Gyasi A, et al. Therapeutic efficacy of artesunate-amodiaquine and artemether–lumefantrine combinations for uncomplicated malaria in 10 sentinel sites across Ghana: 2015–2017. Malar J. 2019;18:206.
Beavogui AH, Camara A, Delamou A, Diallo MS, Doumbouya A, Kourouma K, et al. Efficacy and safety of artesunate-amodiaquine and artemether–lumefantrine and prevalence of molecular markers associated with resistance, Guinea: an open-label two-arm randomised controlled trial. Malar J. 2020;19:223.
Diallo MA, Yade MS, Ndiaye YD, Diallo I, Diongue K, Sy SA, et al. Efficacy and safety of artemisinin-based combination therapy and the implications of Pfkelch13 and Pfcoronin molecular markers in treatment failure in Senegal. Sci Rep. 2020;10:8907.
Koko VS, Warsame M, Vonhm B, Jeuronlon MK, Menard D, Ma L, et al. Artesunate-amodiaquine and artemether–lumefantrine for the treatment of uncomplicated falciparum malaria in Liberia: in vivo efficacy and frequency of molecular markers. Malar J. 2022;21:134.
Konate A, Barro-Kiki PCM, Angora KE, Bedia-Tanoh AV, Djohan V, Kassi KF, et al. Efficacy and tolerability of artesunate-amodiaquine versus artemether–lumefantrine in the treatment of uncomplicated Plasmodium falciparum malaria at two sentinel sites across Cote d’Ivoire. Ann Parasitol. 2018;64:49–57.
Lingani M, Bonkian LN, Yerbanga I, Kazienga A, Valea I, Sorgho H, et al. In vivo/ex vivo efficacy of artemether–lumefantrine and artesunate-amodiaquine as first-line treatment for uncomplicated falciparum malaria in children: an open label randomized controlled trial in Burkina Faso. Malar J. 2020;19:8.
Riloha Rivas M, Warsame M, Mba Andeme R, Nsue Esidang S, Ncogo PR, Phiri WP, et al. Therapeutic efficacy of artesunate-amodiaquine and artemether–lumefantrine and polymorphism in Plasmodium falciparum kelch13-propeller gene in Equatorial Guinea. Malar J. 2021;20:275.
Smith SJ, Kamara ARY, Sahr F, Samai M, Swaray AS, Menard D, et al. Efficacy of artemisinin-based combination therapies and prevalence of molecular markers associated with artemisinin, piperaquine and sulfadoxine-pyrimethamine resistance in Sierra Leone. Acta Trop. 2018;185:363–70.
Gansane A, Moriarty LF, Menard D, Yerbanga I, Ouedraogo E, Sondo P, et al. Anti-malarial efficacy and resistance monitoring of artemether–lumefantrine and dihydroartemisinin–piperaquine shows inadequate efficacy in children in Burkina Faso, 2017–2018. Malar J. 2021;20:48.
Dimbu PR, Horth R, Candido ALM, Ferreira CM, Caquece F, Garcia LEA, et al. Continued low efficacy of artemether–lumefantrine in Angola in 2019. Antimicrob Agents Chemother. 2021;65:e01949-e2020.
Moriarty LF, Nkoli PM, Likwela JL, Mulopo PM, Sompwe EM, Rika JM, et al. Therapeutic efficacy of artemisinin-based combination therapies in Democratic Republic of the Congo and Investigation of molecular markers of antimalarial resistance. Am J Trop Med Hyg. 2021;105:1067–75.
Chico RM, Cano J, Ariti C, Collier TJ, Chandramohan D, Roper C, et al. Influence of malaria transmission intensity and the 581G mutation on the efficacy of intermittent preventive treatment in pregnancy: systematic review and meta-analysis. Trop Med Int Health. 2015;20:1621–33.
Afutu LL, Boampong JN, Quashie NB. High prevalence of molecular markers of Plasmodium falciparum resistance to sulphadoxine-pyrimethamine in parts of Ghana: a threat to ITPTp-SP? J Trop Pediatr. 2021;67:fmaa120.
Dicko A, Sagara I, Djimde AA, Toure SO, Traore M, Dama S, et al. Molecular markers of resistance to sulphadoxine-pyrimethamine one year after implementation of intermittent preventive treatment of malaria in infants in Mali. Malar J. 2010;9:9.
Naidoo I, Roper C. Mapping “partially resistant”, “fully resistant”, and “super resistant” malaria. Trends Parasitol. 2013;29:505–15.
Das S, Kerah-Hinzoumbe C, Kebfene M, Srisutham S, Nagorngar TY, Saralamba N, et al. Molecular surveillance for operationally relevant genetic polymorphisms in Plasmodium falciparum in Southern Chad, 2016–2017. Malar J. 2022;21:83.
Hassen J, Alemayehu GS, Dinka H, Golassa L. High prevalence of Pfcrt 76T and Pfmdr1 N86 genotypes in malaria infected patients attending health facilities in East Shewa zone, Oromia Regional State, Ethiopia. Malar J. 2022;21:286.
Yobi DM, Kayiba NK, Mvumbi DM, Boreux R, Kabututu PZ, Akilimali PZ, et al. Biennial surveillance of Plasmodium falciparum anti-malarial drug resistance markers in Democratic Republic of Congo, 2017 and 2019. BMC Infect Dis. 2022;22:145.
Berzosa P, Molina de la Fuente I, Ta-Tang TH, Gonzalez V, Garcia L, Rodriguez-Galet A, et al. Temporal evolution of the resistance genotypes of Plasmodium falciparum in isolates from Equatorial Guinea during 20 years (1999 to 2019). Malar J. 2021;20:463.
Mensah BA, Aydemir O, Myers-Hansen JL, Opoku M, Hathaway NJ, Marsh PW, et al. Antimalarial drug resistance profiling of Plasmodium falciparum infections in Ghana using molecular inversion probes and next-generation sequencing. Antimicrob Agents Chemother. 2020;64:e01423-e1519.
Tumwebaze PK, Katairo T, Okitwi M, Byaruhanga O, Orena S, Asua V, et al. Drug susceptibility of Plasmodium falciparum in eastern Uganda: a longitudinal phenotypic and genotypic study. Lancet Microbe. 2021;2:e441–9.
Dentinger CM, Rakotomanga TA, Rakotondrandriana A, Rakotoarisoa A, Rason MA, Moriarty LF, et al. Efficacy of artesunate-amodiaquine and artemether–lumefantrine for uncomplicated Plasmodium falciparum malaria in Madagascar, 2018. Malar J. 2021;20:432.
Bwire GM, Ngasala B, Mikomangwa WP, Kilonzi M, Kamuhabwa AAR. Detection of mutations associated with artemisinin resistance at k13-propeller gene and a near complete return of chloroquine susceptible falciparum malaria in Southeast of Tanzania. Sci Rep. 2020;10:3500.
Liu Y, Liang X, Li J, Chen J, Huang H, Zheng Y, et al. Molecular surveillance of artemisinin-based combination therapies resistance in Plasmodium falciparum parasites from Bioko Island, Equatorial Guinea. Microbiol Spectr. 2022;10:e0041322.
Menegon M, L’Episcopia M, Nurahmed AM, Talha AA, Nour BYM, Severini C. Identification of Plasmodium falciparum isolates lacking histidine-rich protein 2 and 3 in Eritrea. Infect Genet Evol. 2017;55:131–4.
Alemayehu GS, Messele A, Blackburn K, Lopez K, Lo E, Janies D, et al. Genetic variation of Plasmodium falciparum histidine-rich protein 2 and 3 in Assosa zone, Ethiopia: its impact on the performance of malaria rapid diagnostic tests. Malar J. 2021;20:394.
Golassa L, Messele A, Amambua-Ngwa A, Swedberg G. High prevalence and extended deletions in Plasmodium falciparum hrp2/3 genomic loci in Ethiopia. PLoS ONE. 2020;15:e0241807.
Iriart X, Menard S, Chauvin P, Mohamed HS, Charpentier E, Mohamed MA, et al. Misdiagnosis of imported falciparum malaria from African areas due to an increased prevalence of pfhrp2/pfhrp3 gene deletion: the Djibouti case. Emerg Microbes Infect. 2020;9:1984–7.
Prosser C, Gresty K, Ellis J, Meyer W, Anderson K, Lee R, et al. Plasmodium falciparum histidine-rich protein 2 and 3 gene deletions in strains from Nigeria, Sudan, and South Sudan. Emerg Infect Dis. 2021;27:471–9.
Acknowledgements
The authors thank the patients and parents or guardians of the study children who participated in the study as well as the health workers at the study health facilities for their support in conducting the studies. Sincere gratitude goes to Dr. Pascal Ringwald for his technical guidance and review of the manuscript.
Disclaimer
Charlotte Rasmussen, Jean Bosco Ndihiokubwayo and Djimrassengar Honoré are employees of the World Health Organization, and they alone are responsible for the views expressed in this publication, which do not necessarily represent the decisions, policies or views of the World Health Organization.
Funding
The work was supported by the World Health Organization. The study was also supported by the Institut Pasteur, Paris, the French government (Agence Nationale de la Recherche), Laboratoire d’Excellence (LabEx) “French Parasitology Alliance for Health Care” (ANR-11-15 LABX-0024-PARAFRAP), and the University of Strasbourg through the IdEX 2022 programme (DM).
Author information
Authors and Affiliations
Contributions
MSI and MHT led and coordinated the conduct of the study, analysis clinical data and contributed to writing of the manuscript. MW contributed to data analysis and led the preparation of the manuscript. IDMS, KB, and HD conceived and designed the study. AHI, OA, MH, and MD supervised the data collection and ensured data quality. CDL and LA performed laboratory work to genotype the parasites and molecular markers for artemisinin and partner drugs. CR provided technical support and reviewed the manuscript. DM led the parasite genotyping and drug resistance marker analysis and contributed to manuscript preparation. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study protocol was reviewed and approved by Ministry of Health, Public and Human Solidarity and the WHO Research Ethics Review Committee. Written informed consent was obtained from adult patient or parent/guardian of children before enrolling in the study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Raw data of msp-1, msp-2, glurp and poly α polymorphisms (band size in bp) detected on day0 and day of recurrence (dayX) in isolates from recurrent infections.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Issa, M.S., Warsame, M., Mahamat, M.H.T. et al. Therapeutic efficacy of artesunate–amodiaquine and artemether–lumefantrine for the treatment of uncomplicated falciparum malaria in Chad: clinical and genetic surveillance. Malar J 22, 240 (2023). https://doi.org/10.1186/s12936-023-04644-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-023-04644-w