
Abstract
Background
The widespread emergence of resistance to pyrethroids is a major threat to the gains made in malaria control. To monitor the presence and possible emergence of resistance against a variety of insecticides used for malaria control in Rwanda, nationwide insecticide resistance surveys were conducted in 2011 and 2013.
Methods
Larvae of Anopheles gambiae sensu lato mosquitoes were collected in 12 sentinel sites throughout Rwanda. These were reared to adults and analysed for knock-down and mortality using WHO insecticide test papers with standard diagnostic doses of the recommended insecticides. A sub-sample of tested specimens was analysed for the presence of knockdown resistance (kdr) mutations.
Results
A total of 14,311 mosquitoes were tested and from a sample of 1406 specimens, 1165 (82.9%) were identified as Anopheles arabiensis and 241 (17.1%) as Anopheles gambiae sensu stricto. Mortality results indicated a significant increase in resistance to lambda-cyhalothrin from 2011 to 2013 in 83% of the sites, permethrin in 25% of the sites, deltamethrin in 25% of the sites and DDT in 50% of the sites. Mosquitoes from 83% of the sites showed full susceptibility to bendiocarb and 17% of sites were suspected to harbour resistance that requires further confirmation. No resistance was observed to fenitrothion in all study sites during the entire survey. The kdr genotype results in An. gambiae s.s. showed that 67 (50%) possessed susceptibility (SS) alleles, while 35 (26.1%) and 32 (23.9%) mosquitoes had heterozygous (RS) and homozygous (RR) alleles, respectively. Of the 591 An. arabiensis genotyped, 425 (71.9%) possessed homozygous (SS) alleles while 158 (26.7%) and 8 (1.4%) had heterozygous (RS) and homozygous (RR) alleles, respectively. Metabolic resistance involving oxidase enzymes was also detected using the synergist PBO.
Conclusion
This is the first nationwide study of insecticide resistance in malaria vectors in Rwanda. It shows the gradual increase of insecticide resistance to pyrethroids (lambda-cyhalothrin, deltamethrin, permethrin) and organochlorines (DDT) and the large presence of target site insensitivity. The results demonstrate the need for Rwanda to expand monitoring for insecticide resistance including further metabolic resistance testing and implement an insecticide resistance management strategy to sustain the gains made in malaria control.
Similar content being viewed by others

Background
Most countries in Africa depend heavily on two vector control interventions in their battle against malaria: long-lasting insecticidal nets (LLINs) and indoor residual spraying (IRS). These tools use insecticides from four chemical classes: organochlorines, pyrethroids, carbamates and organophosphates. Whereas 14 formulations belonging to these classes are approved by the World Health Organization (WHO) for use in IRS [1], only pyrethroids are approved for use in LLINs [2] because of their low mammalian toxicity, excito-repellent properties and rapid knock-down and killing effect [3]. It has been estimated that since 2000 more than 670 million cases of malaria have been averted by combining IRS and LLINs with case management and community education [4, 5]. In Rwanda, the national scale-up of vector control interventions has contributed to a steady reduction of malaria cases from 1.6 million in 2005 to 472,000 cases in 2012 [6, 7]. This reduction has been attributed to the combined effects of universal coverage with LLINs [7] and targeted IRS operations in districts with the highest malaria endemicity (5 out of 30 districts) based on epidemiologic and entomologic data. The Rwanda Malaria Indicator Survey carried out in 2013 showed that overall LLIN coverage is high with 83% of the households owning at least one LLIN and 74% reported to have slept under a LLIN the previous night for pregnant women and children under five [8]. From 2005 to 2013, the National Malaria Control Programme (NMCP) of the Rwanda Ministry of Health has distributed approximately 11.2 million LLINs to a population of an estimated 11 million people [9].
From 2007 to 2012, nationwide distributions of LLIN have been conducted in conjunction with annual IRS applications of pyrethroids in high malaria transmission districts either in focal sectors or district-wide by blanket spraying, covering an estimated 98% of the targeted structures. In 2013, the NMCP shifted from pyrethroids to the use of carbamates (Bendiocarb 80% WP) for IRS as part of an insecticide resistance management strategy. This switch was because of confirmed pyrethroid resistance and following the WHO guidance of using active ingredients with different modes of action in rotation [6, 10].
The main mechanisms by which mosquitoes display resistance to insecticides are the expression of elevated levels of detoxifying enzymes (metabolic resistance) and target site insensitivity (knock-down mutations or altered acetylcholinesterase) [10, 11]. Two point mutations in the voltage-gated sodium channel are associated with knock down resistance (kdr) to DDT and pyrethroids in the malaria mosquito Anopheles gambiae s.s. [12]. One mutation involves a leucine (TTA) to phenylalanine (TTT) substitution at residue 1014 of the gene (L1014F). This mutation is mainly found in West Africa and hence named kdr-west [13]. The other mutation involves a leucine (TTA) to serine (TCA) substitution at the same residue (L1014S) and is mostly found in East Africa (kdr-east) [14], although both mutations co-occur in some parts of Africa [15].
In 2012, the WHO reported that insecticide resistance in malaria vectors had already been found in more than 64 malaria endemic countries worldwide, with the majority reporting resistance to pyrethroids [10]. This spread is alarming as it poses serious threats to the efficacy of vector control interventions and the gains made in malaria control over the last 10 years. Therefore, it is concerning that most national malaria control programmes (NMCPs) continue to use pyrethroid insecticides for vector control. The situation is also compounded by the extensive use of pyrethroids in agriculture, which poses an additional selection pressure on malaria vectors, for example via insecticide-contaminated ground water that permeates to mosquito larval habitats [14, 16, 17].
The WHO calls for all countries to develop and implement insecticide resistance management strategies in their malaria control programmes in order to curb the spread of resistance as well as preserve the effectiveness of LLINs [10]. Many African countries have now implemented entomological monitoring and susceptibility testing. To inform decisions in the control of malaria in Rwanda, the NMCP has been conducting entomological monitoring of malaria vectors in 12 sentinel sites throughout the country since 2010. Both An. gambiae s.l. (94.3%) and Anopheles funestus (5%) were the dominant Anopheles species. Anopheles arabiensis was the predominant sibling species of the An. gambiae complex (E.H., unpublished data). Mosquito susceptibility to the WHO recommended classes of insecticides are included annually in this routine survey to monitor resistance to organochlorines, organophosphates, pyrethroids and carbamates [18]. Here, findings of susceptibility tests conducted in 2011 and 2013 to detect knock-down mutations and mortality rates in female An. gambiae s.l. mosquitoes are described. This is the first report presenting nationwide results on the composition of the An. gambiae complex and its susceptibility to insecticides used for malaria control.
Methods
Study area
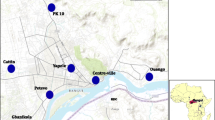
The study was carried out in 2011 and 2013 in 12 sentinel sites that are distributed over the three provinces of Rwanda (Eastern, Southern and Western province) and Kigali City, all having different levels of malaria endemicity. The 12 sentinel sites are: Busoro, Mbuga and Karambi in Southern Province; Rukara, Mimuri, Bukora and Mareba in Eastern Province; Mashesha, Kivumu, Mubuga and Kibogora in Western Province; and Kicukiro in Kigali City (Fig. 1).

Geographic location of the 12 sentinel sites used for insecticide resistance studies, Rwanda, 2011 and 2013
Mosquito collection
Mosquito larvae were collected in the 12 sentinel sites with standard dippers (350 ml) from stagnant water bodies particularly in rice paddies and other temporal breeding habitats typical for Anopheles mosquitoes. All mosquito larvae were collected between 8 and 11 a.m. and brought to a malaria entomology laboratory in the field site where they were reared to adults. Emerging adult Anopheles mosquitoes were put in holding cages and fed with 10% sugar solution from cotton wool pads. The adult holding room temperature was between 22 and 28 °C with a relative humidity of 70–80%. Approximately 15,000 An. gambiae s.l. mosquitoes were reared to the adult stage, of which 13,807 female mosquitoes were tested for insecticide susceptibility according to WHO protocols [18, 19]. A sample of 10% of these mosquitoes was sent to the International Centre of Insect Physiology and Ecology (icipe), Kenya for molecular identification and kdr genotyping [20, 21].
Susceptibility tests
Tests were conducted using kits and insecticide-impregnated filter papers supplied by the WHO collaborating center at the University Sains Malaysia [18]. A batch of 20–25 non blood-fed female mosquitoes that were between 2 and 3 days old were exposed for 1 h to the standard diagnostic concentrations of deltamethrin (0.05%), permethrin (0.75%), lambda-cyhalothrin (0.05%), DDT (4%), bendiocarb (0.1%) and fenitrothion (1%). Each test was run in four replicates and there was one control of 25 field-collected An. gambiae s.l. per test. The control mosquitoes were exposed to silicone oil impregnated paper for a similar period. The number of knocked down mosquitoes was recorded at 10, 15, 20, 30, 40, 50 and 60 min [18]. After 60 min, tested mosquitoes were transferred into the holding tube and supplied with 10% sugar solution for 24 h after which the final mortality was scored.
In order to explore metabolic resistance, a pre-exposure of adult female mosquitoes for 1 h to a synergist (piperonyl butoxide: PBO 4%) was carried out according to WHO protocols in six sites where resistance to pyrethroids was confirmed [19]. These tests were only carried out in 2013 and conducted alongside the susceptibility testing with the main insecticides used for malaria control, permethrin 0.75% and deltamethrin 0.05%. After 1 h pre-exposure to PBO, the mosquitoes were transferred to the tubes lined with the corresponding insecticide-impregnated paper. The counting of knockdown and mortality of mosquitoes pre-exposed to PBO was conducted as described.
Mosquito identification
All mosquitoes were identified to species based on morphological characteristics [22] and stored individually over silica gel awaiting molecular identification and detection of kdr mutations. From each sentinel site, 10% of female mosquitoes plus all mosquitoes classified as ‘resistant’ from each site were sent for molecular characterization to the icipe laboratory in Kenya where genomic DNA was extracted from the mosquitoes and amplified using specific diagnostic primers for An. gambiae [20, 21].
Mutations associated with knock-down resistance (kdr) genes were assayed [11] and the DNA products electrophoresed on a 2% agarose gel with ethidium bromide stain and visualized under UV light to identify the presence of susceptible and resistant alleles.
Interpretation of susceptibility test results
In all tests conducted, the observed mortality in the control tubes was less than 5% and therefore Abott’s correction was not applied [18, 19]. Mortality was calculated as the percentage of individuals that died within 24 h of exposure. Mosquito populations were considered ‘susceptible’ when mortality was between 98 and 100%. When mortality was below 90%, the mosquitoes were classified as ‘resistant’. According to the WHO protocol, an intermediate mortality of 90–97% is suggestive of the existence of resistance and indicates that further investigations are needed [19]. For the prior exposure of mosquitoes to the synergist, a return to full susceptibility to the insecticide in the WHO tube test compared to the WHO tube test with the insecticide-impregnated paper alone indicates that metabolic resistance plays a role in the insecticide resistance observed.
Statistical analysis
The mortality data were analysed and compared using a Generalized Linear Model with a binomial distribution (IBM SPSS statistics V.20, Chicago, IL, USA). The output provided estimated marginal means of mortality, 95% confidence intervals, standard errors and p values based on Chi square tests.
Results
Morphological and molecular identification
A total of 14,311 mosquitoes tested for insecticide resistance was morphologically identified as An. gambiae s.l. Of these, 1406 samples collected from 10 sentinel sites were identified by PCR of which 1165 (82.9%) were An. arabiensis and 241 (17.1%) An. gambiae s.s. (Table 1). Except for Mimuri, An. arabiensis was the dominant species in all sentinel sites.
Mortality rates
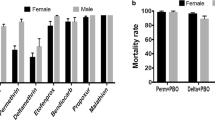
In 2011, mortality results showed that out of the 12 sentinel sites, mosquitoes were fully susceptible to lambda-cyhalothrin in all sites except for Mashesha, where resistance was potentially emerging with a mortality rate of 97.6%. Mosquitoes were fully susceptible to permethrin in 33% of the sites, were suggested to be resistant in 33% of sites (i.e. had an intermediate mortality of 90–97%) and were classified as resistant in the remaining 34% of sites. The permethrin resistant populations were from Bukora (84% mortality), Kibogora (89% mortality), Mimuri (86% mortality) and Rukara (84% mortality). For deltamethrin, mosquitoes were fully susceptible in 58% of sites. In 33% of sites, the existence of resistance was suggested. Resistance to deltamethrin was confirmed for one site: Bukora (85% mortality). For DDT, 28% of sites showed full susceptibility, 36% of sites had a suspicion of resistance and resistance was confirmed in another 36% of sites surveyed: Bukora (77% mortality), Kibogora (75% mortality), Kicukiro (52% mortality) and Mashesha (88% mortality). Mosquitoes were fully susceptible to bendiocarb in 92% of sites. In one site (Kibogora), the mortality of 96% is suggestive of the existence of resistance to bendiocarb. Fenitrothion was the only insecticide to which mosquitoes showed full susceptibility in all the sites tested with 100% mortality after 24 h (Table 2).
Two years later, in 2013, susceptibility to lambda-cyhalothrin was recorded in one site only, while recorded mortalities in 25% of sites were suggestive of the existence of resistance. Resistance was confirmed in 66% of sites. Anopheles gambiae s.l. was resistant to permethrin in 25% of sites, showed susceptibility to permethrin in 16% of sites, and mortalities from 58% of the sites were suggestive of the existence of resistance. For deltamethrin, the mosquitoes showed susceptibility in 25% of the sites, a suggestion of the existence of resistance in 25% of sites, and full resistance in 50% of sites. The test of resistance to DDT was carried out in 10 sites only, and susceptibility was recorded in 10% of sites, whereas 30% of sites suggested the existence of resistance, and full resistance was observed in 60% of sites. Mosquitoes were fully susceptible to bendiocarb in 83% of sites, and two sites (Bukora and Kivumu) suggested the existence of resistance. Mosquitoes were susceptible to fenitrothion in all sites.
Comparing the mortality rates of 2011 and 2013, there was a significant increase in resistance levels to lambda-cyhalothrin in 83% of the sites, to permethrin in 25% of sites, to deltamethrin in 25% of sites and to DDT in 50% of sites. A significant decrease of susceptibility of An. gambiae s.l. to bendiocarb was found in one site (Bukora) and, as in 2011, all mosquitoes tested remained 100% susceptible to fenitrothion in 2013 (Table 2).
kdr Genotypes
Seven hundred and twenty-five (134 An. gambiae s.s. and 591 An. arabiensis) mosquitoes collected in 2011 and 2013 were genotyped for kdr-east (L1014S). Of the 134 An. gambiae s.s. genotyped, 67 (50%) possessed susceptibility (SS) alleles, while 35 (26.1%) and 32 (23.9%) had heterozygous (RS) and homozygous (RR) alleles, respectively. Of the 591 An. arabiensis that were genotyped, 425 (71.9%) possessed homozygous (SS) alleles, while 158 (26.7%) and 8 (1.4%) had heterozygous (RS) and homozygous (RR) alleles, respectively. Anopheles gambiae s.s. and An. arabiensis from Mimuri (Eastern Province) contributed disproportionately to the total observed frequency of the homozygous resistance genotype (RR), with 36 and 12% for the two species, respectively (Table 3).
Pre-exposure of Anopheles gambiae s.l. to piperonyl butoxide
The pre-exposure of Anopheles gambiae s.l. to the synergist piperonyl butoxide (PBO) was conducted in six sites where resistance to pyrethroids was confirmed. The test was conducted only with permethrin 0.75% and deltamethrin 0.05%. In all sites, susceptibility was restored and ranged from 98 to 100% (Tables 4, 5).
Discussion
In Rwanda, integrated malaria control interventions (artemisinin-based combination therapy (ACT), LLINs and targeted IRS) have been in use since 2006. This has contributed to a significant reduction in clinical malaria cases in the country [7]. However, the gains made are fragile due to the decrease of efficacy of interventions, partially as a result of insecticide resistance development that has spread throughout Africa [4, 23, 24]. Rwanda achieved universal coverage with LLINs in 2011, but the major challenge is to maintain this coverage and use with effective mosquito nets, especially after it was recently reported for Rwanda that LLIN effectiveness lasts less than 3 years due to the rapid loss of insecticidal activity and physical deterioration in the field [25]. LLIN deterioration problems were also shown in recent findings from Senegal where damaged nets provided less protection from malaria compared to intact ones [26].
The fact that millions of nets may have lost their effectiveness and thus continue to expose mosquitoes to a sub-lethal dose of pyrethroids is of major concern, because this may contribute to the further development of resistance. Similarly, annual consecutive use of pyrethroids in IRS, combined with extensive use of pyrethroids in agriculture has also implications for emerging insecticide resistance [27]. Hence, Rwanda with the support of PMI and the Global Fund has been keen to monitor insecticide resistance so that it can take action to mitigate the emergence of resistance. In 2010, an initial susceptibility survey was conducted in eight sites in which mosquitoes were tested with deltamethrin, permethrin, bendiocarb, malathion and DDT using the CDC bottle assay [28, 29]. These mosquitoes were found to be fully susceptible, except to DDT in two sites (E.H., unpublished data).
The data collected in the current surveys (2011 and 2013) confirm that resistance to DDT has been on the rise since 2011. In addition, the results show resistance to the pyrethroids (deltamethrin, permethrin, and lambda-cyhalothrin) being present in 2011 and further increasing in 2013. Although DDT was banned for usage in Rwanda in 1989, high levels of resistance are of concern because resistance to DDT confers cross-resistance to pyrethroids [10]. Mosquitoes were fully susceptible to fenitrothion in all the sites, while possible emergence of resistance to bendiocarb was reported in the present study. Nevertheless, a switch to IRS with bendiocarb was made from September 2013 onwards so that the two main vector control interventions (LLINs and IRS) employed different classes of insecticide to delay resistance development.
The presence of kdr mutations recorded in some sites can be explained by intense indoor interventions with IRS and LLINs, as was reported in Burundi and Tanzania [30, 31]. However, it should be noted that the frequency of kdr in the vector population may not be a reliable marker for actual resistance [32] and that care should be taken when interpreting such data as other resistance mechanisms can play a role as well. In current study, metabolic resistance involving oxidases was proven by using piperonyl butoxide (PBO). Susceptibility was fully restored in the six selected sites where resistance to pyrethroids had been identified. This suggests that metabolic resistance may even account for all observed resistance in bioassays with malaria vectors from Rwanda.
The resistance level in Mimuri (Nyagatare District) could explain the high number of malaria cases occurring in this district: about 42% of all malaria cases of the 30 districts in 2011 were found here [33] before the introduction of bendiocarb for IRS. This site, as well as Mashesha, Mareba, and Kibogora, is characterized by rice growing in which agricultural pesticides, and pyrethroids in particular, are extensively used.
In response to these findings, Rwanda developed and implemented an insecticide resistance management (IRM) plan in 2013 that recommended transitioning to non-pyrethroid IRS to mitigate pyrethroid resistance and to lengthen the effectiveness of LLINs [6]. Rwanda transitioned to carbamate (bendiocarb) for IRS and the country will switch to a long-lasting organophosphate (Actellic–pirimiphos-methyl CS) in 2016 due to the reported trends of suggestive resistance to bendiocarb. The question remains, however, whether Rwanda should continue with blanket IRS treatment in an entire district when data on resistance is reported from only one sentinel site per district. To answer this question, it is recommended that additional sites should be identified in the targeted districts in order to determine whether or not the spread of resistance is homogeneous throughout the district. Similar to a recent report from Malawi, the use of long-lasting IRS formulations, such as pirimiphos-methyl, may be costly initially, but cost-effective in the longer term in managing insecticide resistance [34].
In this study, characterization of An. gambiae s.l. from 10 sentinel sites revealed that the predominant sibling species is An. arabiensis (83%). This is contrary to a study conducted in one site near Kigali City in 2007 by PMI-Rwanda, in which it was reported that An. gambiae s.s. accounted for 93.6% of the total 157 An. gambiae s.l. examined by PCR while An. arabiensis accounted for only 6.4% [35]. Although the earlier sampling was carried out in one site only, the results suggest that An. gambiae s.s. was the predominant species before the scale-up of interventions with LLINs and IRS. Such a shift in species composition has been reported in neighbouring countries, for instance in Kenya, Uganda and Tanzania [36–38]. This phenomenon has important implications for malaria epidemiology and control given that An. arabiensis is an opportunistic feeder which has a tendency to rest and feed on humans outdoors [39]. Outdoor-biting mosquitoes are less susceptible to indoor interventions and therefore outdoor interventions that supplement LLINs and IRS will need to be instituted in the context of an integrated approach to vector management [38, 40]. Although An. arabiensis is the dominant vector in Rwanda and resistance levels are high in An. gambiae s.l. populations, future studies should include testing of resistance by sibling species to assess differences in susceptibility levels.
With the goal of reaching the pre-elimination phase of malaria by 2018, the Ministry of Health developed an integrated vector management (IVM) strategy which aims at improving the efficiency, effectiveness, ecological soundness and sustainability of vector control interventions in Rwanda [41]. Entomological monitoring, including testing for insecticide resistance, is now a major part of the NMCP in Rwanda and a rotation strategy for management of insecticide resistance has been adopted in line with IVM.
Meanwhile, with spreading insecticide resistance and behavioural change of malaria vectors, there is an interest to integrate other innovative interventions which do not rely on insecticides [42, 43]. Larval control interventions have proven cost-effective across a range of different settings and include application of environmental management, insect growth regulators, and biological control [44]. Recently, successful field experiments have been carried out with microbial larvicides in Tanzania, Kenya, the Gambia and Benin. These showed a substantial impact on malaria disease [45–47]. It was demonstrated that for a sustainable solution, a horizontally organized community-based programme that takes the needs and wishes of people into account has to be established and technically empowered [48, 49]. Currently, Rwanda is supporting a research project in Ruhuha (South East Rwanda) that aims to involve communities in the application of the microbial larvicide Bacillus thuringiensis var. israelensis (Bti) (E.H., unpublished data). This may form the basis for the integration of alternative vector control interventions for the management of insecticide resistance [42].
Conclusion
The results of the study show that resistance to pyrethroids and organochlorines (DDT) was documented in 2011 and has been on the rise between 2011 and 2013. An emerging resistance to carbamates (bendiocarb) was found in few sites and gives reasons for concern of its efficacy in the near future. No resistance was recorded for organophosphates (fenitrothion). Probably due to the use of indoor insecticide-based interventions, the more opportunistic and outdoor feeding An. arabiensis was the dominant sibling species in the country. To curb further spread of pyrethroid resistance and to preserve the effectiveness of LLINs, Rwanda implemented an insecticide resistance management strategy in 2013 and switched to carbamates (bendiocarb) for IRS. It plans to implement a rotational strategy of insecticides, including organophosphates, every 2–3 years. The results reported here are the first results on the status of insecticide resistance in Rwanda and indicate that the NMCP should continue monitoring insecticide resistance for target site insensitivity and metabolic resistance so as to guide future decisions on insecticide use for public health.
Abbreviations
- ACT:
-
artemisinin-based combination therapy
- Bti :
-
Bacillus thuringiensis var. israelensis
- CDC:
-
Centers for Disease Control and Prevention
- DDT:
-
dichlorodiphenyltrichloroethane
- kdr :
-
knockdown resistance
- LLIN:
-
long-lasting insecticide-treated net
- IRS:
-
indoor residual spraying
- IVM:
-
integrated vector management
- NMCP:
-
National Malaria Control Programme
- PBO:
-
piperonyl butoxide
- PCR:
-
polymerase chain reaction
- PMI:
-
President’s Malaria Initiative
- WHO:
-
World Health Organization
References
WHO. Recommended insecticides for indoor residual spraying against malaria vectors. Geneva: World Health Organization; 2013.
WHO. Recommended long-lasting insecticidal mosquito nets. Geneva: World Health Organization; 2013.
Zaim M, Aitio A, Nakashima N. Safety of pyrethroids treated mosquito nets. Med Vet Entomol. 2000;14:1–5.
WHO. World malaria report 2014. Geneva: World Health Organization; 2014. p. 227.
Bhatt DJ, Weiss E, Cameron D, Bisanzio B, Mappin U, Dalrymple K, et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature. 2015;526:207–11.
MoH. National strategic plan for insecticide resistance management. Kigali: Ministry of Health; 2013. p. 35.
Karema C, Rukundo A, Kabayazi A, Mulindalhabi M, Fall I, Gausi K, et al. Trends in malaria cases, hospital admissions and deaths following scale-up of antimalarial interventions, 2001–2010, Rwanda. Malar J. 2012;11:236.
NISR. Rwanda malaria indicator survey. Kigali: National Institute of Statistics of Rwanda; 2013. p. 92.
NISR. Fourth population and housing census, Rwanda: thematic report on characteristics of households and housing. Kigali: National Institute of Statistics of Rwanda; 2012. p. 256.
WHO. Global Plan for insecticide resistance management in malaria vectors (GPIRM). Geneva: World Health Organization; 2012. p. 132.
Ranson H, Jensen B, Vulule JM, Wang X, Hemmingway J, Collins FH. Identification of a point mutation in the voltage-gated sodium channel gene of Kenyan Anopheles gambiae associated with presence of DDT and pyrethroids. Insect Mol Biol. 2000;9:491–7.
Santolamazza F, Etang J, Barrese E, Dia I, Donnelly MJ, Petrarca V, et al. Distribution of knock-down resistance mutations in Anopheles gambiae molecular forms in west and west-central Africa. Malar J. 2008;7:74.
Martinez-Torres D, Chandre F, Williamson MS, Darriet F, Berge JB, Devonshire AA, et al. Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae s.s. Insect Mol Biol. 1998;7:179–84.
Kabula B, Matowo J, Kitau J, Mweya M, Emidi B, Masue D, et al. Susceptibility status of malaria vectors to insecticides commonly used for malaria control in Tanzania. Trop Med Int Health. 2012;17:742–50.
Pinto J, Elissa N, Donnelly MJ, Costa C, Gentile G, Veder AC. Co-occurrence of East and West African kdr mutations suggests high levels of resistance to pyrethroid insecticides in Anopheles gambiae from Libreville, Gabon. Med Vet Entomol. 2006;20:27–32.
Yadouléton A, Allagbé H, Asidi A, Boko M, Osse R, Padonou G, et al. The impact of expansion of urban vegetable farming in malaria transmission in major cities of Benin. Parasit Vectors. 2014;3:118.
Ranson H, Badolo A, Guelbeogo WM, Hinzoumbé CK, Yangalbé-Kalnoné E, Sagnon NF, et al. Insecticide resistance in Anopheles gambiae: data from the first year of a multi-country study highlight the extent of the problem. Malar J. 2009;8:299.
WHO. Test procedure for insecticide resistance monitoring in malaria vectors: bio-efficacy and persistence of insecticide on treated surfaces. Geneva: World Health Organization; 1998. p. 43.
WHO. Test procedures for insecticide resistance monitoring for malaria vectors mosquitoes. Geneva: World Health Organization; 2013.
Collins F, Mendez MA, Rasmussen MO, Mehaffy PC, Besansky NJ, Funesty N. A ribosomal RNA gene probe differentiates member species of the Anopheles gambiae complex. Am J Trop Med Hyg. 1987;37:37–41.
Scott J, Brogden WG, Collins FH. Identification of single specimen of the Anopheles gambiae complex by the polymerase chain reaction. Am J Trop Med Hyg. 1993;49:520–9.
Gillies G, Coetzee M. A supplement to the Anophelinae of Africa south of the Sahara (Afrotropical region). Publ S Afr Inst Med Res. 1987;55:1–143.
Ranson H, Lines J, Moiroux N, Nkuni Z, Corbel V. Pyrethroid resistance in African anopheline mosquitoes: what are the implications for malaria control? Trends Parasitol. 2011;27:91–8.
Ranson H, Lissenden N. Insecticide resistance in African Anopheles: a worsening situation that needs urgent action to maintain malaria control. Trends Parasitol. 2016;32:187–96.
Hakizimana E, Cyubahiro B, Rukundo A, Kabayiza A, Mutabazi A, Beach R, et al. Monitoring long lasting insecticidal nets (LLINs) durability to validate net serviceable life assumptions in Rwanda. Malar J. 2014;13:344.
Gnanguenon V, Azondekon R, Oke-Agbo F, Sovi A, Ossè R, Padonou G, et al. Evidence of man-vector contact in torn long-lasting insecticide-treated nets. BMC Public Health. 2013;13:751.
Thomas M, Godfray HCJ, Read AF, van den Berg H, Tabashnik BE, van Lenteren JC, et al. Lessons from agriculture for the sustainable management of malaria vectors. PLoS Med. 2012;9:7.
Aïzoun N, Azondekon R, Alia R, Oussou O, Gnanguenon V, Aikpon R, et al. Comparison of the standard WHO susceptibility tests and the CDC bottle bioassay for the determination of insecticide susceptibility in malaria vectors and their correlation with biochemical and molecular biology assays in Benin, West Africa. Parasit Vectors. 2013;6:147.
Brogdon W, Mc Allister JC. Simplification of adult mosquito bioassays through use of time-mortality determination in glass bottles. J Am Mosq Control Assoc. 1998;14:159–64.
Protopopoff N, Verhaeghen K, van Bortel W, Roelants P, Marcotty T, Baza D, et al. A significant increase in kdr in Anopheles is associated with intensive vector control interventions in Burundi highlands. Trop Med Int Health. 2008;13:1479–87.
Kabula B, Tungu P, Malima R, Rowland M, Minja J, Wililo R, et al. Distribution and spread of pyrethroid and DDT resistance among the Anopheles gambiae complex in Tanzania. Med Vet Entomol. 2014;28:244–52.
Donnelly M, Corbel V, Weetman D, Wilding CS, Williamson MS, Black WC. Does kdr genotype predict insecticide-resistance phenotype in mosquitoes. Trends Parasitol. 2009;25:213–9.
MoH. Rwanda Health Management Information System (HMIS) report. Kigali: Ministry of Health; 2011.
Chanda E, Chipwanya J, Mulenga S, Ali D, Troell P, Dodoli W, et al. Preventing malaria transmission by indoor residual spraying in Malawi: grappling with the challenge of uncertain sustainability. Malar J. 2015;14:254.
Lansana K. Rwanda entomology monitoring: Technical Report. Kigali; 2008. p. 24.
Kabbale F, Kaddu JB, Onapa AW. Biting patterns and seasonality of Anopheles gambiae and Anopheles funestus mosquitoes in Kamuli District, Uganda. Parasit Vectors. 2014;7:262.
Mwangangi J, Mbogo CM, Orindi BO, Muturi EJ, Midega JT, Nzovu J, et al. Shifts in malaria vector species composition and transmission dynamics along the Kenyan coast over the past 20 years. Malar J. 2013;8:12.
Russell T, Govella NJ, Azizi S, Drakeley CJ, Kachur SP, Killeen GF. Increased proportion of outdoor feeding among residual malaria vector population following increased use of insecticide treated nets in rural Tanzania. Malar J. 2011;10:80.
Verhulst N, Takken W. Host preferences of blood-feeding mosquitoes. Annu Rev Entomol. 2013;58:433–53.
Tchouassi D, Quakyi IA, Addison EA, Bosompem KM, Wilson MD, Appawu MA, et al. Characterization of malaria transmission by vector populations for improved interventions during the dry season in the Kpone-on-Sea area of coastal Ghana. Parasit Vectors. 2012;5:212.
MoH. National strategic plan for integrated vector management. Kigali: Ministry of Health; 2013. p. 51.
Mukabana W, Kiama GM, Ijumba JN, Mathenge EM, Kiche I, Nkwengulila G, et al. Ecologists can enable communities to implement malaria vector control in Africa. Malar J. 2006;5:9.
WHO. Global strategic framework for integrated vector management. Geneva: World Health Organization; 2004. p. 12.
Beales P, Gilles HM. Rationale and technique of malaria control. In: Warrell DG, Gilles HM, editors. Essential malariology. 4th edn. Arnold edition. London; 2002. p. 107–190.
Geissbuhler Y, Kannady K, Chaki PP, Emidi B, Govella NJ, Mayagaya V, et al. Microbial larvicide application by a large-scale, community-based program reduces malaria infection prevalence in urban Dar es Salaam, Tanzania. PLoS ONE. 2009;4:e5107.
Worrall E, Fillinger U. Large-scale use of mosquito larval source management for malaria control in Africa: a cost analysis. Malar J. 2011;10:338.
Majambere S, Lindsay S, Green C, Kandeh B, Fillinger U. Microbial larvicides for malaria control in The Gambia. Malar J. 2007;6:76.
Chaki P, Shoo B, Hemed A, Tanner M, Fillinger U, Killeen GF. Achieving high coverage of larval-stage mosquito surveillance: challenges for a community-based mosquito control programme in urban Dar es Salaam, Tanzania. Malar J. 2009;8:311.
Chaki P, Mtasiwa D, Tanner M, Mshinda H, Kelly AH, Killeen GF. Institutional evolution of a community-based programme for malaria control through larval source management in Dar es Salaam, United Republic of Tanzania. Malar J. 2014;13:245.
Authors’ contributions
EH designed the study, trained and supervised technical staff, performed resistance and laboratory tests, compiled and analysed data and drafted the manuscript. DM trained and supervised technical staff involved in mosquito rearing and bio-assays, performed laboratory tests and reviewed the manuscript. GI supervised technical staff involved in mosquito rearing and bio-assays, performed bioassays tests and reviewed the manuscript. JET contributed to the data analysis and review of the manuscript. JG contributed to the data analysis, draft of the manuscript, technical guidance and review of the manuscript. CK contributed to the design and implementation of the study, mobilized funds for monitoring activities and reviewed the manuscript. WT contributed to the design of manuscript outlines, data analysis and critical review of the manuscript. AB contributed to the design of the study, and review of the manuscript. CJMK contributed to the data analysis, writing and review of manuscript. All authors read and approved the final manuscript.
Acknowledgements
Great appreciation is extended to Mr. Alphonse Mutabazi, and Alphonse Rukundo who contributed to the mapping of study sites, and data analysis, respectively. We express our sincere gratitude to the entomology technicians who conducted mosquito larval collections and mosquito rearing, as well as the district and community leadership for their cooperation and support during the study. Staff from icipe, Kenya, is acknowledged for their help with carrying out the molecular analyses.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets during and/or analysed during the current study are available from the corresponding author on reasonable request.
Funding
The President’s Malaria Initiative of the US Government (PMI) and the Global Fund Malaria SSF Project provided the financial and technical support to strengthen insecticide resistance monitoring, as well as malaria control and prevention in Rwanda. EH and CJMK are supported by a Grant from the Global Health Systems and Health Policy Research program of the Dutch NWO-WOTRO Science for Global Development.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Hakizimana, E., Karema, C., Munyakanage, D. et al. Susceptibility of Anopheles gambiae to insecticides used for malaria vector control in Rwanda. Malar J 15, 582 (2016). https://doi.org/10.1186/s12936-016-1618-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-016-1618-6