
Abstract
Background
In Comoros, the widespread of chloroquine (CQ)-resistant Plasmodium falciparum populations was a major obstacle to malaria control, which led to the official withdrawal of CQ in 2004. Continuous monitoring of CQ-resistant markers of the P. falciparum CQ resistant transporter (pfcrt) and the P. falciparum multiple drug resistance 1 (pfmdr-1) is necessary inder to obtain first-hand information on CQ susceptibility of parasite populations in the field. The objective of this study is to assess the prevalence and evolution of CQ-resistance in the P. falciparum populations on the Comoros’ Grande Comore island after withdrawal of CQ.
Methods
A total of 207 P. falciparum clinical isolates were collected from the island, including 118 samples from 2006 to 2007 and 89 samples from 2013 to 2014. Nucleotide substitutions in the pfcrt and pfmdr-1 genes linked to CQ response in parasite isolates were assessed using nested PCR and DNA sequencing.
Results
From the pfcrt gene segment sequenced, we detected C72S, M74I, N75E, and K76T substitutions in the parasite isolates collected from both 2006–2007 to 2013–2014 periods. Significant decline of pfcrt resistant alleles at C72S (42.6 to 6.9 %), M74I (39.1 to 14.9 %), N75E (63.5 to 18.3 %), and K76T (72.2 to 19.5 %) from 2006–2007 to 2013–2014 were observed, and the frequency of pfcrt wild type allele was significantly increased from 19.1 % in 2006–2007 to 75.8 % in 2013–2014. Sequence analysis of pfmdr-1 also detected point mutations at codons N86Y, Y184F, and D1246Y, but not S1034C and N1042D, in the isolates collected from both examined periods. An increasing trend in the prevalence of the pfmdr-1 wild type allele (NYD, 4.3 % in 2006–2007; and 28.7 % in 2013–2014), and a decreasing trend for pfmdr-1 N86Y mutation (87.0 % in 2006–2007; and 40.2 % in 2013–2014) were observed in our samples.
Conclusions
The present data indicate that the prevalence and patterns of mutant pfcrt and pfmdr-1 dramatically decreased in the Grande Comore isolates from 2006 to 2014, suggesting that the CQ-sensitive P. falciparum strains have returned after the withdrawal of CQ. The data also suggests that the parasites with wild type pfcrt/pfdmr-1 genes may have growth and/or transmission advantages over the mutant parasites. The information obtained from this study will be useful for developing and updating anti-malarial treatment policy in Grande Comore island.
Similar content being viewed by others


Background
Despite being a readily preventable and treatable disease, malaria still infects more than 210 million people and kills approximately 438,000 individuals annually [1]. In the Union of Comoros (including Moheli, Anjouan Island, and Grande Comore islands), Plasmodium falciparum infection was one of the most serious public health problems until 2013, and malaria made up 15–30 % of the hospitalization cases and contributed 15–20 % of registered deaths in the pediatric services [2]. One of the main factors contributing to the disease burden is the emergence and spread of parasites resistant to anti-malarial drugs in malaria-endemic areas of the world [3]. Chloroquine (CQ) has been the first-line treatment of acute uncomplicated falciparum malaria in this island nation since the 1950s. Unfortunately, the first case of CQ-resistance (CQR) P. falciparum malaria was reported in Comoros in 1980 [4]. Since the first report, various studies have subsequently confirmed that the emergence and spread of CQR parasite strains [5–7], leading to the replacement of CQ with artemisinin-based combined therapy (ACT), including artemether-lumefantrine (AL), as the first-line therapy for uncomplicated P. falciparum malaria in 2004. However, it should be noted that there was period (between 2004 and 2007) with overlapping CQ and AL treatments due to the unavailability of AL treatment in some health facilities in Comoros. Additionally, massive application of long-lasting insecticide-treated nets and indoor residual sprayings had been implemented in Comoros since 2007. Furthermore, mass drug administration (MDA) with a therapeutic dose of artemisinin-piperaquine (AP) plus a low-dose of primaquine (APP, Artepharm Co. Ltd, PR China) was launched in 2007, 2012, and 2013 on Moheli, Anjouan, and Grande Comore islands, respectively. According to a report from the Ministry of Health, the numbers of annual malaria cases have been dramatically reduced after MDA, from 108,260 in 2006–2154 in 2014 in Union of Comoros (a 97.7 % reduction) and from 92,480 in 2006–2142 in 2014 in Grande Comore (a 98.0 % reduction). The dramatic reduction in annual malaria cases in Grande Comore could be mainly attributed to ACT-based MDA regimens in synergy with other malaria control measures. Currently, delayed parasite clearance (DPC) after ACT treatment has been reported in countries of Southeast Asia, including Cambodia, Thailand, Myanmar, Vietnam, and Laos [8–10]. The increased K13-propeller gene mutations previously associated with DPC among P. falciparum populations from 2013 to 2014 in Grande Comore (a ~20 % increment) may present new challenges in the ACT efficacy in the future [11, 12]. To achieve the ambitious goal to completely eliminate malaria by 2020 in Comoros, as well as to preserve the high efficacy of ACT, there is an urgent need to develop and update anti-malarial guidance in Comoros.
Resistance to CQ in P. falciparum parasites is mainly linked to mutations in the P. falciparum CQR transporter gene (pfcrt), which encodes a protein localized at the membrane of parasite digestive vacuole (DV). A second gene called P. falciparum multidrug resistance gene 1 (pfmdr-1) that encodes a P-glycoprotein homologue and is also located at the membrane of the parasite DV may modulate the level of resistance. Specific mutations in pfcrt (K76T) and pfmdr-1 (N86Y) have been used as molecular markers for monitoring CQR in the field parasite populations [13, 14]. The continuous monitoring of molecular markers is a very useful tool for malaria control in endemic areas, since it may detect changes in parasite susceptibility to anti-malarial drugs and provide guidance to therapeutic policies. Several studies have assessed allele frequencies in different time periods in various malaria-endemic areas. For example, in Malawi over 8 years [15], in Papua New Guinea over 12 years [16], in Gabon over 14 and 25 years [17, 18], and in Eastern Kenya over 14 years [19]. In Malawi [15, 20, 21], Gambia [18], Kenya [19], Ethiopia [22], and Tanzania [23], the withdrawal of CQ chemotherapy led to a decline in the frequency of mutant pfcrt and pfmdr-1 alleles and the return of CQ sensitive (CQS) malaria. In the contrary, high frequencies of CQR strains were still detected in some endemic regions despite the withdrawal of CQ from national treatment guidelines [17, 24, 25].
Before the change of the drug policy in Comoros, clinical failure rates of CQ therapy ranged between 31 and 90 % [7, 26, 27]. Subsequent genotyping showed that the prevalence of the pfcrt K76T and the pfmdr-1 N86Y mutations ranged from 62 to 98 %, and from 90 to 100 %, respectively [7, 26, 28]. After the change in the drug policy, the frequency of the 76T and 86Y mutant alleles were 43–80 and 97–99 %, respectively, in 2006 [29–31]. Currently, the status of P. falciparum susceptibility to CQ in Comoros is unknown. Monitoring the parasite’s CQ susceptibility is crucial for considering the possibility of reintroducing this safe and affordable drug in Comoros. Thus, the aim of this study is to assess the prevalence and evolution of CQR by investigating the temporal variations of pfcrt and pfmdr-1 gene polymorphisms in P. falciparum isolates collected from Grande Comore during the periods of 2006–2007 and 2013–2014.
Methods
Study sites
This study was conducted in three malaria endemic sites of the Grande Comore Island (Mitsoudje Centre Hospital, National Malaria Centre, and Mitsamiouli Centre Hospital), Union of Comoros (Fig. 1). The island is located in the Indian Ocean between Madagascar and the eastern coast of Africa, with 11°00′–12°00′S latitude and 43°10′–43°35′E longitude. The population of this island was estimated to be 420,000 in 2012. The island has a tropical climate with temperature ranging from 11 to 35 °C and rainfall of 1000–3000 mm per year (rainy season between November and April and dry season between May and October). Malaria is transmitted principally by Anopheles gambiae s.l. and Anopheles funestus [2]. Malaria transmission is year-round with a peak of transmission during the rainy season. P. falciparum and Plasmodium malariae malaria are present on the island, with P. falciparum being the predominant species (>95.5 %).

Map of Grande Comore island, Union of Comoros showing the locations of Mitsoudje Center Hospital (open triangle), National Malaria Center (open square), and Mitsamiouli Center Hospital (open circle) where P. falciparum isolates were collected
Study samples
Ethical approval for this study was obtained from the Ethics Committees of Comoros Ministry of Health (No. 07-123/VP-MSSPG/DNS) and Guangzhou University of Chinese Medicine (No. 2012L0816). Blood samples were collected in two different periods (March 2006–October 2007, and March 2013–December 2014) from patients with symptomatic malaria admitted to Mitsoudje Center Hospital, National Malaria Center, and Mitsamiouli Center Hospital for anti-malarial drug treatment. Written informed consent from all adults or legal guardians of children was also obtained. Inclusion criteria were patients infected with P. falciparum, but not other human malaria species, as confirmed by peripheral thick and thin blood smear examination after Giemsa staining. A 1.0 ml of whole blood was collected from P. falciparum patients in an EDTA tube and stored at −20 °C. A total of 207 P. falciparum clinical blood samples were collected, including 118 samples from March 2006 to October 2007 and 89 from March 2013 to December 2014.
PCR amplification and sequence analysis of P. falciparum crt and mdr-1 genes
Parasite DNA was isolated from 100 μl of blood sample using Takara DNA Blood Mini Kit following the manufacturer’s instructions (Takara, Kyoto, Japan). The extracted DNA was dissolved in TE buffer (10 mM Tris–HCl, 0.1 M EDTA, pH 8.0) and stored at −20 °C until use. Segments of the pfcrt and pfmdr-1 genes spanning codons 72–76 (pfcrt) and codons 86, 184, 1034, 1042, and 1246 (pfmdr-1) were amplified as described protocol previously [22]. For pfcrt and pfmdr-1 both the primary and nested amplifications were carried out in 25 μl of final volume with 10.0 μl of dH2O, 12.5 μl of Taq PCR Mast Mix (2.5 U), and 0.4 µM of forward and reverse primers, following the manufacturer’s instructions (Sangon Bio Inc., Shanghai, China) on a S1000 Thermal cycler (Bio-Rad, Hercules, USA). Primary amplification reactions were initiated with the addition of 2.0 μl of template genomic DNA prepared from the blood samples. For the nested PCR, 0.5 μl of primary PCR productions was used as a template. The amplified PCR products were separated on a 2.0 % agarose gel run with a 100 bp DNA ladder (Sangon Bio Inc., Shanghai, China). The nested PCR products were directly sequenced in both directions using an ABI PRISM3730 DNA sequencer (Sangon Bio Inc., Shanghai, China). The nucleotide and amino acid sequences of the pfcrt and pfmdr-1 genes were compared with those of wild-type sequences of pfcrt (GenBank accession number KM288867) and pfmdr-1 (GenBank accession number XM_001351751) using Clustal W of the BioEdit 7.0 and MEGA 4.0 programs.
Statistical analysis
Statistical significance was determined using SPSS software (version 13.0). Mann–Whitney U test was used to compare in the frequencies of the mutations and alleles of the pfcrt and pfmdr-1 in isolates collected between 2006–2007 and 2013–2014. P < 0.05 was considered statistically significant.
Results
Prevalence and patterns of mutant pfcrt gene in P. falciparum isolates
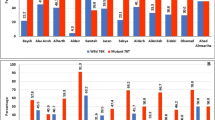
All blood samples (n = 207) were identified as P. falciparum mono-species infections after the thin blood smear examination. Because of low parasitaemia and/or poor DNA quality, some samples were not successfully amplified the pfcrt gene. Only 98 % of the P. falciparum isolates from Grande Comore Island (n = 202) could be amplified, including samples collected during 2006–2007 (n = 115) and 2013–2014 (n = 87). Mutations at codons C72S, M74I, N75E, and K76T of the pfcrt gene were detected in isolated from both 2006–2007 and 2013–2014 groups (Table 1). Mutant codon K76T was the most prevalent in both 2006–2007 and 2013–2014 groups, accounting for 72.3 and 19.5 % of the isolates examined, respectively. Over the course of 8 years (2006–2014), the number of non-synonymous mutations significantly decreased from 42.6 to 6.7 % at codon C72S (P < 0.01), from 39.1 to 14.9 % at codon M74I (P < 0.01), from 63.5 to 18.3 % at codon N75E (P < 0.01), and from 72.2 to 19.5 % at codon K76T (P < 0.01).
Haplotype analysis of the pfcrt gene revealed that nine and six distinct allelic forms were detected in isolates from the 2006–2007 to 2013–2014 groups, respectively (Table 2). Of the nine allelic variants, the most prevalent were the quadruple-mutant allele (72S/74I/75E/76T) (26.1 %, 30/115) among 2006–2007 group isolates, followed by the WT allele (19.1 %, 22/115), triple mutant allele (74I/75E/76T) (13.0 %, 15/115), single mutant allele 76T (11.3 %, 13/115), and another triple mutant allele (72S/75E/76T) (10.4 %, 12/115), and a double mutant allele (75E/76T) (8.7 %, 10/115). The remaining three allelic variants were only detected in 13 (11.3 %) of P. falciparum isolates in the 2006–2007 group. Among the six allelic variants in the 2013–2014 group, WT allele was the most prevalent, accounting for 75.9 % (66/87). Except for the triple mutant allele (74I/75E/76T) with the frequency of 12.6 %, the remaining allelic variants were evenly distributed at low frequency (2.2 to 4.5 %) among isolates in the 2013–2014 group. Compared with the 2006–2007 group, the frequency of WT allele was significantly (P < 0.05) increased in those from 2013 to 2014 group, whereas the frequencies of single mutant allele 75E (P < 0.05) and 76T (P < 0.01), triple mutant allele 72S/75E/76T (P < 0.01), and quadruple-mutant allele 72S/74I/75E/76T (P < 0.01) were significantly decreased.
Prevalence and patterns of mutant pfmdr-1 gene in P. falciparum isolates
The pfmdr-1 segment was successfully amplified from 115 of 118 (97 %) and 87 of 89 (98 %) of the P. falciparum isolates collected from 2006–2007 to 2013–2014, respectively. Sequence analysis revealed point mutations at codons N86Y, Y184F, and D1246Y of the pfmdr-1 gene in isolates collected in both 2006–2007 and 2013–2014 groups (Table 1). None of the investigated P. falciparum clinical isolates carried S1034C or N1042D substitution. Among the above three mutants, codon N86Y was the most prevalent, accounting for 87.0 %, followed by codons Y184F and D1246Y with frequency of 52.2 and 26.1 % in isolates from 2006 to 2007 group, respectively. Similarly, the N86Y was the most prevalent in the 2013–2014 group, accounting for 40.2 %, followed by codons Y184F and D1246Y with frequency of 29.8 and 13.8 %, respectively. The frequencies of mutations at codons N86Y (P < 0.01), Y184F (P < 0.01), and D1246Y (P < 0.05) were significantly decreased in the 2013–2014 group compared with those in the 2006–2007 group, respectively.
Haplotype analysis of pfmdr-1 gene of the samples revealed eight distinct allelic forms (Table 2), including the wild type allele, three single-mutant alleles (86Y, 184F, and 1246Y), three double-mutant alleles (86Y/184F, 86Y/1246Y, and 184F/1246Y), and one triple mutant allele (86Y/1246Y/1246Y). Of the eight allelic variants, the most prevalent allelic variant was single-mutant allele 86Y in isolates from both 2006–2007 (34.8 %, 40/115) and 2013–2014 (33.3 %, 29/87) groups, followed by double-mutant allele 86Y/184F in isolates from 2006 to 2007 (30.4 %, 35/115) and wild type allele in isolates from 2013 to 2014 group (28.7 %, 25/115). Compared with the 2006–2007 group, the frequency of WT allele (changed from 4.3 to 28.7 %) was significantly (P < 0.01) increased in those from 2013 to 2014 group, whereas the frequency of double-mutant allele (86Y/184F) was significantly decreased (from 30.4 to 14.9 %; P < 0.01).
Discussion
In Comoros, with the official ban of CQ in 2004, CQ has now been out of use for almost 12 years although self-medication might have continued for a few years after the ban. The dramatic reduction of annual malaria cases in Grande Comore may be mainly due to ACT-based MDA regimens and other malaria control interventions. However, the emergence and spread of artemisinin-resistant P. falciparum parasites in Southeast Asia [8–10] and the increased K13-propeller gene diversity among P. falciparum populations from 2006 to 2014 in Grande Comore (a ~20 % increment) [11, 12] may open up a new challenge in the ACT efficacy in the future and call for monitoring the changes in parasite susceptibility to CQ and other anti-malarial drugs. The issue as to whether or not CQ-susceptible strains may be returned in malaria-endemic regions after the withdrawal of CQ selective pressure is of great interest for malaria control. This study assessed the evolution of CQR by investigating the temporal variations of pfcrt and pfmdr-1 gene polymorphisms in P. falciparum isolates collected from Grande Comore for two different periods (2006–2007 and 2013–2014). Due to the dramatic reduction of annual malaria cases in Grande Comore since 2013, the number of P. falciparum isolates collected from 2013 to 2014 (n = 89) was smaller than those from 2006 to 2007 (n = 118). Our data showed that the frequencies of mutant alleles pfcrt and pfmdr-1 significantly decreased in isolates from 2013 to 2014 when compared with those from 2006 to 2007, suggesting that CQS P. falciparum populations have returned to Grande Comore after the withdrawal of CQ.
The long history of CQ use has exposed P. falciparum to the drug pressure continually in Comoros. Clinical failure after standard CQ treatment was initially reported in a case of P. falciparum malaria in Comoros in 1980 [4]. A trend of gradual increment of CQ clinical failure rates (from 31 to 90 %) had been documented in this malaria-endemic area between 1990 and 2004 subsequently [7, 26, 27]. Sets of SNPs in the pfcrt codons 72, 74, 75, and 76 were associated with CQR in P. falciparum from Southeast Asia, Africa and South America [14, 32]. The pfcrt K76T mutation has been considered as the most reliable molecular marker of CQR among the various mutations identified [14, 32]. In the present study, the high frequency (72.2 %) of pfcrt K76T was observed in P. falciparum isolates from 2006 to 2007. Our data are consistent with those in other reports, where 45–80 % of Comoros P. falciparum isolates collected in 2006–2007 had the pfcrt K76T mutation [29–31]. We observed an increasing trend in the prevalence of the pfcrt K76 wild type allele (19.1 % in 2006–2007; and 75.9 % in 2013–2014) and a decreasing trend for pfcrt K76T mutation (72.2 % in 2006–2007; and 19.5 % in 2013–2014). The decreasing trend for pfcrt K76T mutation in our study is consistent with several previous reports from other malaria endemic regions. In Ghana, a decrease in frequency of the pfcrt K76T mutation (from 88 to 56 % for 2005–2010) was observed [33]. In Malawi [15] and Kenya [19], decreases from 85 to 13 % between the years of 1993 and 2000 and from 95 to 60 % between 1993 and 2006 were reported, respectively. Similar decreases in mutant pfcrt were reported in many other regions, including Senegal [34], Mozambique [35], Tanzania [23], and Madagascar [36, 37]. In another in vivo study, P. falciparum parasites carrying the CQS pfcrt K76 allele was selected after treatment with AL [38]. The observation of increased CQS pfcrt K76 allele after AL use is consistent with our results; discontinue of CQ use (or CQ related drugs such as amodiaquine) allows the return of parasites with wild type pfcrt allele. Thus, the decreasing trend for pfcrt K76T mutation observed in our study may highlight a benefit of using AP or AL in an area with a high prevalence of CQ-resistant P. falciparum malaria such as Grande Comoros, but not artemether-lumefantrine-amodiaquine (AQ) because AQ (with amodiaquine) was shown to select for pfcrt 76T allele and pfmdr1 YYY haplotype [39]. In contrast, high frequencies of the pfcrt K76T mutant were detected in some countries despite the withdrawal of CQ for many years, including Gabon [17] and Benin [24]. The decreases in mutant pfcrt alleles after withdraw of CQ use is likely due to fitness cost of the pfcrt mutations, and the persistence of high frequencies of the mutant alleles in some parasite populations is because of compensatory changes in the parasite genome that allow the parasite to overcome the negative effects of drug resistant mutations. For example, in a study of gene expression in pfcrt mutants, CQ treated pfcrt mutants were found to have significant enrichment in glycerol and polyol metabolic processes and iron/cation transport activities [40]. Compensatory mutations have been widely reported in other microorganisms [41].
The mutant pfcrt SVMNT haplotype, which is mostly associated with amodiaquine resistance and lower level of CQR compared to CVIET [42], had been reported mostly in Southeast Asia (India and Laos) [43–45] and South America [46, 47], but is still very rare in Africa [48]. In the present study, the mutant pfcrt SVMNT (2.6 %, 3/195) observed in the P. falciparum isolates examined was identical to that of CQR isolates identified in Ethiopia [22] and Tanzania [48]. Therefore, results from the current study suggest that P. falciparum populations with the mutant pfcrt SVMNT allele may have a global distribution now.
Although pfmdr-1 haplotype alone does not determine the level of CQ resistance, the mutations at codons N86Y, S1034C, N1042D, and D1246Y in pfmdr-1 gene are related to reduced sensitivity to CQ [49]. It was reported that CQR P. falciparum populations were introduced into Madagascar from Comoros islands [29], and in vivo resistance to CQ in Madagascar that was not associated with the pfcrt 76T mutation, but with the mutation pfmdr-1 86Y and perhaps with other yet unknown mechanisms [49, 50]. In the present study, none of the P. falciparum clinical isolates collected in 2006–2007 and 2013–2014 carried S1034C and N1042D substitutions, which was similar to those reported from Madagascar [30]. In the present study, three mutations at codons N86Y (87.0 %), Y184F (52.2 %), and D1246Y (26.1 %) were detected in the 2006–2007 group isolates. The results from this study are similar to other previous reports in Grande Comore, where 97.4 and 77.5 % of parasite isolates collected in 2006 had mutations at codons N86Y and D1246Y, respectively, followed by only 1.8 % of isolates with pfmdr-1 wild type gene [29, 30]. In the present study, over the course of 8 years (2006–2014), the frequencies of mutations in pfmdr-1 gene dramatically changed from 87.0 to 40.2 % (at codon N86Y, P < 0.01) and from 26.1 to 13.8 % (at codon D1246Y, P < 0.05). Our data are in line with those of a report describing the change in frequency of pfmdr-1 gene mutations in Madagascar isolates [30, 37], in which the pfmdr-1mutation N86Y was reduced from 50 to 11 % 6 years after the withdrawal of CQ. Similarly, with the prevalence of pfmdr-1 N86Y reaching a peak in 2000 (78 %), there was a highly significant decline in prevalence of the pfmdr-1 N86Y allele in 2008 in Gambia [18]. Therefore, the observations from this study suggested that an increasing trend in P. falciparum susceptibility to CQ may exist in Grande Comore currently. However, this observation showed that over the course of 8 years (2006–2014), the frequencies of mutations in pfmdr-1 gene at codon Y184F dramatically changed from 52.2 to 30.0 % (P < 0.01) in the present study. The observation was in contrast to other reports from Kenya [39], Uganda [51], and Sudan [52] where the pfmdr-1 184F allele has been previously associated with in vivo selection by AQ or AL.
In the present study, the decline in prevalence of pfmdr-1 gene mutations (~24 %) was lower than those in pfcrt gene mutations (~67 %) among Grande Comore isolates over the course of 8 years (2006–2014). The observation is line with other reports describing mdr-1 and crt polymorphisms in P. falciparum isolates collected from Malawi, in which ~30 or ~70 % decline was observed in the change of prevalence of mutant pfmdr-1 or pfcrt among P. falciparum isolates collected from 1992 to 2000, respectively [15]. These observations suggest that pfmdr-1 mutations may be less deleterious to parasite fitness than the pfcrt mutations [13, 14], even though pfmdr-1 mutations can modulate the level of CQ resistance.
In the present study, the rapid shift in P. falciparum from CQR to CQS suggests that the replacement of CQ with ACT for the treatment of P. falciparum may eventually result in the significant decline of pfcrt and pfmdr-1 mutations or CQ-resistant strains in Grande Comore isolates. It was reported that the return of CQS was the result of re-expansion of the susceptible parasites, but not back mutations in a formerly resistant parasite or a new selective sweep in Africa [20] where the malaria transmission rates are higher and naturally immune individuals are more common compared with those in Southeast Asia. Whether or not the CQS resurgence is due to the expansion of surviving CQS reservoir populations or back-mutations in the CQR allele in Grande Comore can not be inferred from our results.
Conclusions
Results from the current study showed that the prevalence and patterns of mutant pfcrt and pfmdr-1 dramatically decreased in Grande Comore isolates over the course of 8 years (2006–2014). Although the data related to the in vivo or in vitro efficacy of CQ of the parasites was not collected in this study, the present data suggest that the CQS P. falciparum parasites have returned in Grande Comore to some extent after the official withdrawal of CQ. The data presented here will be useful for developing and updating anti-malarial policies in Grande Comore.
Abbreviations
- CQ:
-
chloroquine
- CQS:
-
CQ-sensitive
- CQR:
-
CQ-resistance
- Pfcrt :
-
P. falciparum CQ resistant transporter
- Pfmdr-1 :
-
P. falciparum multiple drug resistance 1
- ACT:
-
artemisinin-based combined therapy
- AL:
-
artemether-lumefantrine
- AP:
-
artemisinin-piperaquine
- AQ:
-
artemether-amodiaquine
References
WHO. World malaria report 2015. Geneva: World Health Organization; 2015.
Ouledi A. Epidemiology and control of malaria in the Federal Islamic Republic of Comoros. Sante. 1995;5:368–71.
WHO. Global report on antimalarial drug efficacy and drug resistance 2000–2010. Geneva: World Health Organization; 2010.
Eichenlaub D, Pohle H. A case of chloroquine-resistant (R1) falciparum malaria from the East African Comoros Islands (in German. Infection. 1980;8:90–2.
Feillet N, Agnamey P, Brasseur P, Druilhe P. In vivo resistance of Plasmodium falciparum to chloroquine in Anjouan (Comoros) (in French). Bull Soc Pathol Exot. 1993;86:48–51.
Blanchy S, Julvez J, Mouchet J. Epidemiological stratification of malaria in the Comoro archipelago (in French). Bull Soc Pathol Exot. 1999;92:177–84.
Ariey F, Randrianarivelojosia M, Duchemin J, Rakotondramarina D, Ouledi A, Robert V, et al. Mapping of a Plasmodium falciparum Pfcrt K76T mutation: a useful strategy for controlling chloroquine resistance in Madagascar. J Infect Dis. 2002;185:710–2.
Phyo A, Nkhoma S, Stepniewska K, Ashley E, Nair S, McGready R, et al. Emergence of artemisinin-resistant malaria on the western border of Thailand: a longitudinal study. Lancet. 2012;379:1960–6.
Ariey F, Witkowski B, Amaratunga C, Beghain J, Langlois A, Khim N, et al. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature. 2014;505:50–5.
Ashley E, Dhorda M, Fairhurst R, Amaratunga C, Lim P, Suon S, et al. Spread of artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med. 2014;371:411–23.
Huang B, Deng C, Yang T, Xue L, Wang Q, Huang S, et al. Polymorphisms of the artemisinin resistant (K13) in Plasmodium falciparum parasite populations of Grande Comore island 10 years after artemisinin combination therapy. Parasite Vectors. 2015;8:634.
Torrentino-Madamet M, Collet L, Lepère J, Benoit N, Amalvict R, Ménard D, et al. K13-propeller polymorphisms in Plasmodium falciparum isolates from patients in Mayotte in 2013 and 2014. Antimicrob Agents Chemother. 2015;59:7878–81.
Reed M, Saliba K, Caruana S, Kirk K, Cowman A. Pgh1 modulates sensitivity and resistance to multiple antimalarials in Plasmodium falciparum. Nature. 2000;403:906–9.
Djimde A, Doumbo O, Cortese J, Kayentao K, Doumbo S, Diourte Y, et al. A molecular marker for chloroquine-resistant falciparum malaria. N Eng J Med. 2001;344:257–63.
Kublin J, Cortese J, Njunju E, Mukadam R, Wirima J, Kazembe P, et al. Reemergence of chloroquine sensitive Plasmodium falciparum malaria after cessation of chloroquine use in Malawi. J Infect Dis. 2003;187:1870–5.
Nsanzabana C, Hastings I, Marfurt J, Müller I, Baea K, Rare L, et al. Quantifying the evolution and impact of antimalarial drug resistance: drug use, spread of resistance, and drug failure over a 12-year period in Papua New Guinea. J Infect Dis. 2010;201:435–43.
Frank M, Lehners N, Mayengue P, Gabor J, Dal-Bianco M, Kombila D, et al. A thirteen-year analysis of Plasmodium falciparum populations reveals high conservation of the mutant pfcrt haplotype despite the withdrawal of chloroquine from national treatment guidelines in Gabon. Malar J. 2011;10:304.
Nwakanma D, Duffy C, Amambua-Ngwa A, Oriero E, Bojang K, Pinder M, et al. Changes in malaria parasite drug resistance in an endemic population over a 25-year period with resulting genomic evidence of selection. J Infect Dis. 2014;209:1126–35.
Mwai L, Ochong E, Abdirahman A, Kiara S, Ward S, Kokwaro G, et al. Chloroquine resistance before and after its withdrawal in Kenya. Malar J. 2009;8:106.
Laufer M, Takala-Harrison S, Dzinjalamala F, Stine O, Taylor T, Plowe C. Return of chloroquine-susceptible falciparum malaria in Malawi was a reexpansion of diverse susceptible parasites. J Infect Dis. 2010;202:801–8.
Laufer M, Thesing P, Eddington N, Masonga R, Dzinjalamala F, Takala S, et al. Return of chloroquine antimalarial efficacy in Malawi. N Engl J Med. 2006;355:1959–66.
Mekonnen S, Aseffa A, Berhe N, Teklehaymanot T, Clouse R, Gebru T, et al. Return of chloroquine-sensitive Plasmodium falciparum parasites and emergence of chloroquine-resistant Plasmodium vivax in Ethiopia. Malar J. 2014;13:244.
Mohammed A, Ndaro A, Kalinga A, Manjurano A, Mosha J, Mosha D, et al. Trends in chloroquine resistance marker, Pfcrt-K76T mutation 10 years after chloroquine withdrawal in Tanzania. Malar J. 2013;12:415.
Ogouyèmi-Hounto A, Ndam N, Kinde Gazard D, d’Almeida S, Koussihoude L, Ollo E, et al. Prevalence of the molecular marker of Plasmodium falciparum resistance to chloroquine and sulphadoxine/pyrimethamine in Benin 7 years after the change of malaria treatment policy. Malar J. 2013;12:147.
Inoue J, Lopes D, Rosario V, Machado M, Hristov A, Lima G, et al. Analysis of polymorphisms in Plasmodium falciparum genes related to drug resistance: a survey over four decades under different treatment policies in Brazil. Malar J. 2014;13:372.
Randrianarivelojosia M, Raherinjafy R, Migliani R, Mercereau-Puijalon O, Ariey F, Bedja S. Plasmodium falciparum resistant to chloroquine and to pyrimethamine in Comoros. Parasite. 2004;11:419–23.
Silai R, Moussa M, Abdalli M, Astafieva-Djaza M, Hafidhou M, Oumadi A, et al. Surveillance of falciparum malaria susceptibility to antimalarial drugs and policy change in the Comoros (in French). Bull Soc Pathol Exot. 2007;100:6–9.
Parola P, Pradines B, Gazin P, Keundjian A, Silai R, Parzy D. Chemosusceptibility analysis of Plasmodium falciparum imported from Comoros to Marseilles, France in 2001–2003. Med Mal Infect. 2005;35:489–91.
Menard D, Randrianarivo-Solofoniaina A, Ahmed B, Jahevitra M, Andriantsoanirina V, Rasolofomanana R, et al. Drug-resistant malaria parasites introduced into Madagascar from Comoros islands. Emerg Infect Dis. 2007;13:1759–62.
Andriantsoanirina V, Ratsimbasoa A, Bouchier C, Jahevitra M, Rabearimanana A, Radrianjafy R, et al. Plasmodium falciparum drug resistance in Madagascar: facing the spread of unusual pfdhfr and pfmdr-1 haplotypes and the decrease of dihydroartemisinin susceptibility. Antimicrob Agents Chemother. 2009;53:4588–97.
Rebaudet S, Bogreau H, Silaï R, Lepère J, Bertaux L, Pradines B, et al. Genetic structure of Plasmodium falciparum and elimination of malaria, Comoros archipelago. Emerg Infect Dis. 2010;16:1686–94.
Plowe C, Wellems T. Detection of mutations in a putative Plasmodium falciparum transporter linked to chloroquine resistance. Report for the WHO workshop on markers of antimalarial drug resistance; 1999.
Duah N, Matrevi S, de Souza D, Binnah D, Tamakloe M, Opoku V, et al. Increased pfmdr1 gene copy number and the decline in pfcrt and pfmdr1 resistance alleles in Ghanaian Plasmodium falciparum isolates after the change of anti-malarial drug treatment policy. Malar J. 2013;12:377.
Wurtz N, Fall B, Pascual A, Diawara S, Sow K, Baret E, et al. Prevalence of molecular markers of Plasmodium falciparum drug resistance in Dakar, Senegal. Malar J. 2012;11:197.
Raman J, Mauff K, Muianga P, Mussa A, Maharaj R, Barnes K. Five years of antimalarial resistance marker surveillance in Gaza Province, Mozambique, following artemisinin-based combination therapy roll out. PLoS ONE. 2011;6:e25992.
Deloron P, Le Bras J, Ramanamirija J, Coulanges P. Plasmodium falciparum in Madagascar: in vivo and in vitro sensitivity to seven drugs. Ann Trop Med Parasitol. 1985;79:357–65.
Maïga-Ascofaré O, Rakotozandrindrainy R, Girmann M, Hahn A, Randriamampionona N, Poppert S, et al. Molecular epidemiology and seroprevalence in asymptomatic Plasmodium falciparum infections of Malagasy pregnant women in the highlands. Malar J. 2015;4:188.
Sisowath C, Petersen I, Veiga M, Martensson A, Premji Z, Bjorkman A, et al. In vivo selection of Plasmodium falciparum parasites carrying the chloroquine-susceptible pfcrt K76 allele after treatment with artemether-lumefantrine in Africa. J Infect Dis. 2009;199:750–7.
Henriques G, Hallett R, Beshir K, Gadalla N, Johnson R, Burrow R, et al. Directional selection at the pfmdr1, pfcrt, pfubp1, and pfap2mu loci of Plasmodium falciparum in Kenyan children treated with ACT. J Infect Dis. 2014;210:2001–8.
Jiang H, Patel J, Yi M, Mu J, Ding J, Stephens R, et al. Genome-wide compensatory changes accompany drug-selected mutations in the Plasmodium falciparum crt gene. PLoS ONE. 2008;3:e2484.
Levin B, Perrot V, Walker N. Compensatory mutations, antibiotic resistance and the population genetics of adaptive evolution in bacteria. Genetics. 2000;154:985–97.
Sa J, Twu O. Protecting the malaria drug arsenal: halting the rise and spread of amodiaquine resistance by monitoring the Pfcrt SVMNT type. Malar J. 2010;9:374.
Vathsala P, Pramanik A, Dhanasekaran S, Devi C, Pillai C, Subbarao S, et al. Widespread occurrence of the Plasmodium falciparum chloroquine resistance transporter (Pfcrt) gene haplotype SVMNT in P. falciparum malaria in India. Am J Trop Med Hyg. 2004;70:256–9.
Awasthi G, Prasad G, Das A. Population genetic analyses of Plasmodium falciparum chloroquine receptor transporter gene haplotypes reveal the evolutionary history of chloroquine-resistant malaria in India. Int J Parasitol. 2011;41:705–9.
Dittrich S, Alifrangis M, Stohrer J, Thongpaseuth V, Vanisaveth V, Phetsouvanh R, et al. Falciparum malaria in the north of Laos: the occurrence and implications of the Plasmodium falciparum chloroquine resistance transporter (pfcrt) gene haplotype SVMNT. Trop Med Int Health. 2005;10:1267–70.
Fidock A, Nomura T, Talley K, Cooper A, Dzekunov M, Ferdig T, et al. Mutations in the P. falciparum digestive vacuole transmembrane protein Pfcrt and evidence for their role in chloroquine resistance. Mol Cell. 2000;6:861–71.
Cooper R, Hartwig C, Ferdig M. pfcrt is more than the Plasmodium falciparum chloroquine resistance gene: a functional and evolutionary perspective. Acta Trop. 2005;94:170–80.
Alifrangis M, Dalgaard M, Lusingu J, Vestergaard L, Staalsoe T, Jensen A, et al. Occurrence of the Southeast Asian/South American SVMNT haplotype of the chloroquine-resistance transporter gene in Plasmodium falciparum in Tanzania. J Infect Dis. 2006;193:1738–41.
Andriantsoanirina V, Ratsimbasoa A, Bouchier C, Tichit M, Jahevitra M, Rabearimanana S, et al. Chloroquine clinical failures in P. falciparum malaria are associated with mutant Pfmdr-1, not Pfcrt in Madagascar. PLoS ONE. 2010;5:e13281.
Rason M, Andrianantenaina H, Ariey F, Raveloson A, Domarle O, Randrianarivelojosia M. Prevalent pfmdr1 n86y mutant Plasmodium falciparum in Madagascar despite absence of pfcrt mutant strains. Am J Trop Med Hyg. 2007;76:1079–83.
Dokomajilar C, Nsobya S, Greenhouse B, Rosenthal P, Dorsey G. Selection of Plasmodium falciparum pfmdr1 alleles following therapy with artemether-lumefantrine in an area of Uganda where malaria is highly endemic. Antimicrob Agents Chemother. 2006;50:1893–5.
Gadalla N, Adam I, Elzaki S, Bashir S, Mukhtar I, Oguike M, et al. Increased pfmdr1 copy number and sequence polymorphisms in Plasmodium falciparum isolates from Sudanese malaria patients treated with artemether-lumefantrine. Antimicrob Agents Chemother. 2011;55:5408–11.
Authors’ contributions
JS and BH designed, organized, and supervised the programs. BH, YG, QW, CD, AB, KA, RA, AM, LP, GL, DL, and HZ carried out the field work. BH and XzS analyzed data, wrote, and revised the manuscript. TY, JW, SH, YL, SZ, QX, LX, and FL participated in field work and preliminary data analysis. All authors read and approved the final manuscript.
Acknowledgements
We thank all doctors and nurses at the field sites for their extensive help in this study. We also thank all patients and their relatives for their participation. We would like to thank Dera Tompkins, NIH Library Editing Service, for reviewing the manuscript.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Data supporting the conclusion are included in the main text.
Consent for publication
All authors have approved the final draft, contributed to it significantly and commensurate with their status as co-authors, and agreed with the decision to seek publication. Each participant consented to the publication of data contained in this manuscript.
Ethics approval and consent to participate
The study was approved by the Ethics Committees of Comoros Ministry of Health (No. 07-123/VP-MSSPG/DNS) and Guangzhou University of Chinese Medicine (No. 2012L0816). Blood samples from patients with symptomatic malaria admitted to hospitals were obtained upon signed acceptance of informed consent terms and conditions by the participant.
Funding
This work was supported in part by grants from Natural Science Foundation of China [Grant Number 81273643] and Guangdong Provincial Science and Technology Program [Grant Number 2014B050502013] to JS, China Postdoctoral Science Foundation [Grant Number 2015M570699 and 2016T90773], Guangdong Provincial Science Foundation [Grant Number 2015A030310107], and Guangdong Provincial Medicine Science Foundation [Grant Number A2016315] to BH, Foundation of Traditional Chinese Medicine Bureau of Guangdong Province [Grant Number 20141074] to QW, Science and Technology Program of Guangzhou [2014J4500037] to YG, and by the Division of Intramural Research, National Institute of Allergy and Infectious Diseases, National Institutes of Health (XzS). The funds had no role in the study design, data collection and analysis, decision to publish, or the preparation of the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Bo Huang and Qi Wang contributed equally to this work
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Huang, B., Wang, Q., Deng, C. et al. Prevalence of crt and mdr-1 mutations in Plasmodium falciparum isolates from Grande Comore island after withdrawal of chloroquine. Malar J 15, 414 (2016). https://doi.org/10.1186/s12936-016-1474-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-016-1474-4