
Abstract
Background
Acute myeloid leukemia (AML), a malignancy Often resistant to common chemotherapy regimens (Cytarabine (Ara-c) + Daunorubicin (DNR)), is accompanied by frequent relapses. Many factors are involved in causing chemoresistance. Heme Oxygenase-1 (HO-1) and Hypoxia-Inducible Factor 1-alpha (HIF-1α) are two of the most well-known genes, reported to be overexpressed in AML and promote resistance against chemotherapy according to several studies. The main chemotherapy agent used for AML treatment is Ara-c. We hypothesized that simultaneous targeting of HO-1 and HIF-1α could sensitize AML cells to Ara-c.
Method
In this study, we used our recently developed, Trans-Activator of Transcription (TAT) - Chitosan-Carboxymethyl Dextran (CCMD) - Poly Ethylene Glycol (PEG) - Nanoparticles (NPs), to deliver Ara-c along with siRNA molecules against the HO-1 and HIF-1α genes to AML primary cells (ex vivo) and cell lines including THP-1, KG-1, and HL-60 (in vitro). Subsequently, the effect of the single or combinational treatment on the growth, proliferation, apoptosis, and Reactive Oxygen Species (ROS) formation was evaluated.
Results
The designed NPs had a high potential in transfecting cells with siRNAs and drug. The results demonstrated that treatment of cells with Ara-c elevated the generation of ROS in the cells while decreasing the proliferation potential. Following the silencing of HO-1, the rate of apoptosis and ROS generation in response to Ara-c increased significantly. While proliferation and growth inhibition were considerably evident in HIF-1α-siRNA-transfected-AML cells compared to cells treated with free Ara-c. We found that the co-inhibition of genes could further sensitize AML cells to Ara-c treatment.
Conclusions
As far as we are aware, this study is the first to simultaneously inhibit the HO-1 and HIF-1α genes in AML using NPs. It can be concluded that HO-1 causes chemoresistance by protecting cells from ROS damage. Whereas, HIF-1α mostly exerts prolific and direct anti-apoptotic effects. These findings imply that simultaneous inhibition of HO-1 and HIF-1α can overcome Ara-c resistance and help improve the prognosis of AML patients.
Similar content being viewed by others

Introduction
Acute myeloid leukemia (AML) is a hematological neoplasm, resulting from the clonal proliferation of malignant myeloid progenitors [1, 2]. AML is distinguished by the repletion of myeloblasts in the Bone Marrow (BM) and the Peripheral Blood (PB) [3]. The treatment may include chemotherapy and Hematopoietic Stem Cell Transplantation (HSCT) [2]. The main chemotherapy regimens for AML consist of Cytarabine (Ara-c) plus Daunorubicin (DNR) [4]. Despite these intensive treatment regimens, overall and disease-free survival rates of patients remain low due to frequent relapse and chemo-resistance, in particular for the elderly [2, 5, 6]. Thus, identifying new therapeutic targets for AML is of great importance [2].
Ara-c, a pyrimidine nucleoside analog, is one of the major AML treatment regimens, which is injected intravenously. Following cellular uptake, Ara-c is metabolized into nucleotide triphosphate by deoxycytidine kinase and is incorporated into the DNA strand, resulting in cell cycle arrest in the S phase [7, 8]. Moreover, Ara-c is reported to induce cell death by inducing the formation of Reactive Oxygen Species (ROS). ROS generation subsequently upregulates antioxidant enzymes such as Heme oxygenase 1 (HO-1). Therefore, silencing HO-1 can be beneficial by further sensitizing AML cells to Ara-c [9].
HO-1 or Heat shock protein 32 (Hsp32) is an inducible enzyme coded by the HMOX1 gene on the 22q13.1 chromosome [10]. HO-1 catabolizes heme (a prosthetic group of hemoproteins, and an oxidant) into biliverdin, iron, and carbon monoxide [11]. According to several studies, HO-1 is overexpressed in different malignancies [2]. HO-1 protects leukemic cells by various mechanisms such as inducing autophagy [12], proliferation, inhibiting inflammation [13], and apoptosis in response to chemotherapeutics [1]. HO-1 utilizes various pathways and interacts with several molecules to protect leukemic cells, but not all of them are known.
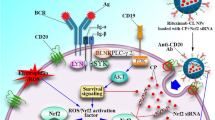
Hypoxia-Inducible Factors (HIF) are super-regulators in several processes such as angiogenesis and proliferation [14]. Our recent experiences have shown that targeting HIF-1 has a significant effect in inhibiting the growth and spread of cancer [15, 16]. HIF-1α is a ubiquitously expressed subunit and is reported to be overexpressed in AML cells [17]. HIF-1α induces Glucose Transporter-1 (GLUT-1), and hexokinase expression. Therefore, resulting in glycolysis induction, rather than in oxidative phosphorylation, thus decreasing ROS generation [1, 18]. HIF-1α also upregulates Vascular Endothelial Growth Factor (VEGF) as well as CXC chemokine type 4 Receptors (CXCR4) [18]. It has been reported that following HO-1 inhibition by siRNA, HIF-1α expression was also decreased [1]. Thus, it seems that HO-1 influences HIF-1α expression as well. Resistant leukemic cells are also reported to be adapted to hypoxic conditions as well [7, 19]. Thus, HIF-1α, as a major response element to hypoxia, might be a suitable target. HO-1 and HIF-1α are reported to be overexpressed in AML cells [2, 20]. Various roles of HIF-1α and HO-1 in AML resistance to therapy are demonstrated in Fig. 1.

The co-inhibition of HO-1 and HIF-1α synergizes with Ara-c treatment in AML. HIF complex is made of two subunits: HIF-1α and HIF-1β. Under atmospheric conditions, HIF-1α is hydroxylated on proline residues by Prolyl hydroxylases (PHDs), followed by binding to Von Hippel-Lindau (VHL), resulting in HIF-1α ubiquitination and degradation by the proteasome. While in hypoxic conditions, HIF-1α is preserved from degradation. In AML cells, HIF-1α is aberrantly overexpressed, despite the absence of hypoxia. Several molecular mechanisms have been proposed to explain this phenomenon (described in the discussion). Following the accumulation of HIF-1α and HIF-1β in the nucleus, the HIF-1 complex is formed which then binds to the HRE regions on the promotors of HIF-1 target genes. These genes include HO-1, VEGF, ENO, GAPDH, ALD, PYK, GLUT-1, LDH, etc. involved in the chemoresistance of AML cells. The induced expression of GLUT-1 by HIF-1 is reported to induce glycolysis and hypoxia resistance resulting in AML chemoresistance. The upregulated ALD, ENO, LDH, PYK, and GAPDH elevate glucose metabolism and induce chemoresistance as well. HIF-1 complex is also reported to induce the production of VEGF, which in turn induces proliferation and inhibits leukemic cell apoptosis by binding to its receptor. Also, this binding induces IL-6, GM-CSF, and G-CSF production resulting in further proliferation. Another target gene of the HIF-1 complex is HO-1. HO-1 exerts direct anti-apoptotic effects on the cell by upregulating Bcl-XL, and Bcl-2, and downregulating BAX and BAK. It is also reported that this molecule increases cell survival by inducing the JNK-c-Jun signaling pathway. However, almost all of the anti-apoptotic actions of HO-1 are derived from its products. HO-1 is responsible for the degradation of free heme into Fe2+, CO, and biliverdin. By degrading free heme molecules of the cell, HO-1 decreases ROS production and cell damage, as free heme is considered a crucial factor leading to the production and accumulation of ROS in the cell. Released CO molecule binds to Cyt C Oxidase which in turn activates the p38 MAPK signaling pathway, resulting in cell survival. Biliverdin is converted into bilirubin (a strong anti-oxidant) and protects the cell from damage caused by ROS. Also, bilirubin possesses anti-inflammatory properties as well, due to its ability to inhibit the presence of P and E selectins on the surface of the cell. The released Fe2+ atom induces ferritin production as well as the activity of a Fe2+ efflux pump called FPN1, both of which result in decreased ROS production and improved cell viability. Therefore, by simultaneous inhibition of these genes combined with a more efficient Ara-c delivery into the cells, the pro-apoptotic activity of Ara-c is improved and the amount of ROS in the cell is predicted to be elevated, resulting in more apoptosis
In the current study, siRNA molecules were used to silence genes and study their functions. Although siRNAs can inhibit gene expression, they are poorly unstable due to degradation by phosphatases and RNases [21]. To efficiently transfect cells with siRNA and increase the stability of siRNAs, we used our recently developed nanoparticles (NPs) [22]. NPs increment the solvency of medications and diminish the necessary successful dose [23, 24]. We used Trans-Activator of Transcription (TAT)-Chitosan-Carboxymethyl Dextran (CCMD)-Poly Ethylene Glycol (PEG) NPs designed in our previous study. TAT usage increases the efficiency of transfection. All the necessary tests on NPs to ensure the efficient delivery of the therapeutics to the leukemic cells were performed in our previous study [22].
As far as we are aware, there is no similar research targeting both HO-1 and HIF-1α simultaneously in AML using NPs. In the following study, the effect of Ara-c on ROS generation as well as the proliferation of AML cell lines was studied. In addition, HO-1 and HIF-1 inhibition effect combined with Ara-c was investigated on the growth of AML cells.
Methods
Reagents
Ara-c was obtained from Cayman Chemical Company. NaOH was purchased from Sigma-Aldrich (Manheim, Germany). Human HIF-1α targeting siRNA (catalog number: sc-35,561), and human HO-1 targeting siRNA (catalog number: sc-35,554), were bought from Santa Cruz Biotechnology, Inc. The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2 H-tetrazolium bromide (MTT) cell proliferation assay kit was purchased from Sigma Aldrich and was used according to the directions given by the company. Cell Proliferation Enzyme-Linked ImmunoAssay (ELISA) bromodeoxyuridine (BrdU) (colorimetric) was supplied by (Roche Molecular Biochemicals, Indianapolis, IN).
Clinical samples and cell lines
After obtaining informed consent from patients, heparinized PB and BM samples were collected from 11 Untreated/ relapsed AML patients from Shahid Ghazi hospital in Tabriz according to the declaration of Helsinki. A brief description of patients’ characteristics is presented in the supplementary Table S1. Peripheral blood mononuclear cells (PBMCs) and bone marrow mononuclear cells (BMMCs) were isolated from the samples via Ficol paque Plus (Sigma Aldrich). HL-60 (AML-M2), THP-1 (AML-M5), and KG-1 (AML-M6) cell lines were supplied by the national cell bank of Iran (Pasteur Institute of Iran). Cells were cultivated in RPMI-1640 medium, enriched with 20% FBS, 100 unit/mL streptomycin, and 100 unit/mL penicillin, and cultured in a standard incubator with 95% humidity, 37 ̊C temperature, and 5% CO2.
Transfection of cells with NPs loaded with siRNA and Ara-c
In the current study, CCMD-PEG-TAT NPs were used to deliver Ara-c and siRNAs against HO-1 and HIF-1 into the leukemic cells. As demonstrated by our previous study, these NPs are highly effective in transfecting hematological malignant cells [22]. The presence of TAT increases the efficiency of this transfection [22]. The cells were cultivated and then treated with one of the following pharmacological compounds for 24 (for qRT-PCR, MTT test), 48 h (for MTT test), and 72 h (for protein analysis); Untreated, NP-HIF-1α siRNA, NP-HO-1 siRNA, NP-HO-1 + HIF-1α siRNA, Ara-c, NP-Ara-c, NP-HO-1 siRNA-Ara-c, NP- HIF-1α siRNA-Ara-c, NP- HIF-1α + HO-1 siRNA + Ara-c. For each treatment group, 50 nM of each siRNA was used. The amount of drug was used according to the measured IC50 value for each cell (Supplementary Figure S1 and Table 1).
Gene expression analysis
The total RNA of cells was extracted by TRIzol Reagent (Invitrogen). RNA was then transcribed into cDNA and stored at -20 ̊C. Using SYBER Green PCR Master Mix (Thermo Fisher, US) a µl of cDNA was used for qRT-PCR by a light-cycler 480 qRT-PCR system (Roche). ∆∆CT method was utilized for quantification relative to β-actin gene expression. The used primer sequences are available in Table 2.
ROS detection
ROS generation was assessed with 5,6-chloromethyl-2’,7’-dichlorodihydrofluorescein diacetate (CM-H2DCF-DA, Invitrogen). After treating cells with the different treatment groups for 24 h, cells were washed and incubated with 10 µM CM-H2DCF-DA (100 µL) for 30 min at 37 °C. Subsequently, excess CM-H2DCF-DA was washed, and finally, ROS was detected by a plate reader at 485 nm (excitation) and 530 nm (emission).
Cell line proliferation assay by BrdU incorporation
For quantification of cell proliferation, BrdU (Roche Molecular Biochemicals) was used. Pre-performing the proliferation assay, cell lines were cultured and incubated with the various pharmacological compounds for 24 h in a 96-well plate at the concentration of 20,000 cells/well. Subsequently, 20 µl of BrdU labeling solution was added (final concentration 10 µM BrdU) and cells were incubated for 24 h at 37 ̊C. Next, cells were centrifuged at 300 ×g for 10 min, and the upper solution was removed. Cells were then dried by a hair dryer for 15 min. After that, 200 µl/well of FixDenant was added (incubation for 30 min at 20 ̊C). FixDenant solution was then removed and 100 µl of Anti-BrdU-POX solution was added and cells were incubated for 90 min at 20 ̊C. Next, the washing solution was used to remove the antibody, and thereafter, washing was removed by flicking it off. Eventually, 100 µl/well of substrate solution was added and the plate was incubated at 20 ̊C for 20 min. The blank was 100 µl of culture medium.
Gene knockdown verification at the protein level by colorimetry
To evaluate the protein levels of target genes after 72 h of treatment with the various pharmacological groups, a QuantiSir™ Gene Knockdown Quantification Kit (EPIGENTEK, US) was used. The protocol provided by the manufacturer was followed. Horseradish peroxidase was used as the color developing agent. GAPDH was used as control. The absorbance values were read at 450 nm on a microplate reader [30].
Cytotoxicity and cell death analysis
Post-treatment with various combinational compounds to assess cell growth, an MTT test was carried out. Primary cells and cell lines (2 × 104 cells/well) were inoculated in a 96-well plate and incubated at 37 ̊C, 5% CO2 for a day. Then, the medium was replaced with 100 µL of medium enriched with various pharmacological mixtures and was incubated for 24–48 h. Next, the medium was replaced with 200 µL of medium containing 10 µL of MTT solution (incubation for 4 h). Finally, 200 µL of DMSO replaced the medium (mixture for 10 min). Eventually, a microplate reader was utilized to assess the OD of each well at 570 nm. The following equation was used to assess the viability percentage of cells [31]:
Evaluation of intrinsic apoptosis pathway by Caspase-9 activity assessment
To assess the activity of Caspase-9, a Caspase-9 Colorimetric Assay (R&D Systems) was used. Following the treatment of cells with various pharmacological groups for 24 h, cells were washed with PBS. Subsequently, cells were lysed with lysis buffer and the cell lysate was used to assess the activity of Caspase-9 according to the directions of the manufacturer. The system measures the level of caspase-9 enzymatic activity in the cell lysate by measuring the color reaction following the addition of a caspase-specific peptide conjugated to a color reporter molecule p-nitroalanine (pNA), which is then cleaved by caspase-9 activity. The OD of the plates was read using a microplate reader at 405 nm.
Statistical analysis
GraphPad Prism V9 software was used to statistically analyze the data. A one-way ANOVA test was used to evaluate the differences between groups. The linear regression between the parameters was determined by Pearson’s correlation assay. A significance level was considered less than 0.05. Each assay was carried out in triplicate.
Results
NPs loaded with Ara-c and siRNAs significantly suppressed HO-1 and HIF-1α expression in AML primary cells and cell lines
qRT-PCR and protein analytic tests were utilized to evaluate the effect of NPs loaded with Ara-c and siRNA molecules on HO-1 and HIF-1α expression in AML PBMCs, BMMCs, and cell lines. Following 24 h treatment of the cell lines and primary cells with NPs loaded with HO-1, HIF-1α, and Ara-c, cellular mRNA was extracted and analyzed. The results demonstrated that following HO-1 silencing by siRNA-loaded NPs, HO-1 mRNA levels were highly reduced in the AML-related cell lines (Fig. 2a) and patient-derived BMMCs and PBMCs (Fig. 2b). Similar results were observed regarding the expression level of HIF-1α in cell lines (Fig. 2c) and primary cells (Fig. 2d) of AML patients following treatment with therapeutics-loaded NPs.

NPs efficiently decreased HO-1 and HIF-1α mRNA levels in treated cells, measured by qRT-PCR. mRNA expression levels of HO-1 after treatment with various pharmacological treatments in (a) HL-60, KG-1, and THP-1 cell lines and patients-derived PBMCs and BMMCs (b). mRNA expression levels of HIF-1α after treatment with various pharmacological treatments in (c) HL-60, KG-1, and THP-1 cell lines and patients-derived PBMCs and BMMCs (d). The values are normalized to the β-actin mRNA expression level, and then relatively targeted gene mRNA levels were determined by taking the ratio to the value of the untreated cell as a calibrator based on the ΔΔCT method. The cell line is presented as means ± SD of triplicate experiments. Primary AML patient PBMCs and BMMCs sample data are presented as means of individual patient experiments (n = 11). P-values < 0.05 (*), P-values < 0.01 (**), and P-values < 0.001 (***)
Moreover, the decrease in the expression of each of the factors could have a slight decreasing effect on the expression of the other factor. This result suggests that probably HIF-1α and HO-1 influence each other’s expression. Interestingly, treatment of cells with Ara-c and especially NPs loaded with Ara-c considerably elevated HO-1 and HIF-1α expression. But as expected, treating cells with NPs loaded with siRNAs against HO-1 and HIF-1α could neutralize the inducing effect of Ara-c.
In addition to qRT-PCR, following 72 h of cell incubation with any of the treatments, protein levels were analyzed to evaluate the efficacy of gene silencing (Fig. 3). It was discovered that the level of the HO-1 protein level considerably decreased after exposure of AML cell lines (Fig. 3a) and primary cells (Fig. 3b) to NPs loaded with HO-1 siRNA. Silencing HO-1 also decreased the expression of HIF-1α very slightly. In addition, similar to what was observed in mRNA level expression, treatment of cells with Ara-c and especially NPs loaded with Ara-c significantly increased the expression of HO-1, which was neutralized by the addition of anti-HO-1 siRNA.

Treatment with NP-bound siRNA inhibited the protein expression of HO-1, and HIF-1α, in AML cells, determined by colorimetry. The protein expression level of HO-1 based on the colorimetric test after treatment with various compounds in (a) HL-60, KG-1, and THP-1 cell lines and (b) in AML patients derived PBMCs and BMMCs. The protein expression levels of HIF-1α based on the colorimetric test after treatment with various compounds in (c) HL-60, KG-1, and THP-1 cell lines and (d) in AML patients derived PBMCs and BMMCs. The values are normalized to the GAPDH protein expression level, then relative targeted genes’ protein levels were determined by taking the ratio to the value of the untreated cell as a calibrator. The cell line is presented as means ± SD of triplicate experiments. Primary AML patient PBMCs and BMMCs sample data are presented as means of individual patient experiments (n = 11). P values < 0.05 (*), P values < 0.01 (**), and P values < 0.001 (***)
Evaluation of HIF-1α expression after treatment with NPs had similar results to HO-1. NPs loaded with anti-HIF-1α siRNA significantly reduced the expression of this factor (this effect was also observed to some extent on HO-1 expression as well) in both cell lines and primary cells of AML patients. Also, Ara-c considerably increased the expression of HIF-1α, which was reversed by adding anti-HIF-1α. (Figs. 3d, e). An interesting point was that the simultaneous addition of both siRNAs caused the greatest silencing effect on both HO-1 and HIF-1α expression in the cells.
Inhibition of HO-1 and HIF-1α sensitized AML cells to Ara-c in
To determine the IC50 value of Ara-c in cell lines and primary cells of AML patients, an MTT test was performed following 24 h incubation with different concentrations of Ara-c. IC50 values are presented in Supplementary Figure S1 and Table 1.
Subsequently, based on IC50 results, another MTT test was carried out to study the effect of combinational treatments of HO-1/HIF-1α siRNA with Ara-c on the growth of AML cells. After 24 h of cell treatment with the mentioned compounds, we found that inhibition of HO-1 and HIF-1α in combination with Ara-c had an almost similar effect and increased cell sensitivity to Ara-c. Notably, the effect of HIF-1α inhibition was slightly higher when compared to HO-1 silencing. Moreover, the co-inhibition of HO-1 and HIF-1α, induced almost the same apoptosis rates as free Ara-c treatment did. However, the highest rate of cell death was observed in cells that received NPs loaded with siRNAs against HIF-1α, HO-1, and Ara-c treatment. This indicates that inhibition of these two genes could eliminate resistance to Ara-c chemotherapy. Also, it was evident that Ara-c-loaded NPs exerted more apoptosis induction compared to any siRNA-carrying NPs and even higher than free Ara-c. In addition, the growth inhibition was slightly higher in HL-60 cells, compared to KG-1, and THP-1 (Fig. 4a). Also, the effect of treatments increased over the next 24 h incubation, indicating the time-dependency of the treatment (Fig. 4b). The same results were observed in patient-derived AML cells as well (Fig. 4c, d).

The viability of (a, b) HL-60, KG-1, and THP-1 cell lines and patient-derived PBMCs and BMMCs (c, d) was decreased following 24 and 48 h treatment with different pharmacological groups. A Caspase-9 assay kit was used to evaluate the activity of Caspase-9 in HL-60, KG-1, and THP-1 cell lines (e) and Patient-derived AML cells (f) following the treatment with various combination treatments. Cell line data are presented as means ± SD of triplicate experiments. Primary AML patient PBMCs and BMMCs samples data are presented as means of individual patients experiments (n = 11). P-values < 0.05 (*), P-values < 0.01 (**), and P-values < 0.001 (***)
Simultaneous HO-1 and HIF-1α silencing in combination with Ara-c-induced intrinsic apoptosis pathway
To investigate the impact of combination therapy on the intrinsic apoptosis pathway, the activity of Caspase-9 was evaluated in both cell lines and primary cells following 24 h incubation with the treatments (Fig. 4e, f). In cells with silenced HO-1 or HIF-1α genes, the activity of caspase-9 was significantly elevated compared to the controls while, co-inhibition of HO-1 and HIF-1α increased caspase-9 activity almost as free Ara-c, but not as much as NP-Ara-c. Moreover, silencing each gene seemed to form a synergism with Ara-c in inducing Caspase-9 activity. The effect of HIF-1α silencing was slightly higher than HO-1 inhibition. Moreover, it was evident that the co-inhibition of HO-1 and HIF-1α combined with Ara-c treatment was associated with the highest activity of Caspase-9 compared to other cell groups.
HO-1 and HIF-1α co-inhibition affects the expression level of apoptosis-associated proteins
To study the impact of combination therapy on apoptosis-involved factors, we next evaluated the effect of various treatments on the mRNA levels of apoptosis-related proteins including B-cell lymphoma-2 (Bcl-2) and Bcl-2 Associated X protein (BAX).
Treating cell lines with HIF-1α siRNA-NP significantly decreased Bcl-2 mRNA expression and elevated the expression of BAX mRNA as for the treatment with HO-1 siRNA-NP but with slighter amounts (Fig. 5a, b). The same results were observed with primary cells as well (Fig. 5c, d). Co-inhibition of HIF-1α and HO-1 exerted a more significant effect which was even comparable with free Ara-c treatment. On the other hand, the effect of NP-Ara-c was much more considerable. The results demonstrated a notable increase in changes of Bcl-2 and BAX mRNAs following the treatment with NPs loaded with HO-1 and HIF-1α siRNAs and Ara-c. Thus, silencing these two genes directly induced apoptosis in AML cells.

Treating cells with different mixtures of drugs and siRNAs, affected the mRNA expression levels of Bcl-2 and BAX, achieved by qRT-PCR. mRNA expression levels of BAX and Bcl-2 gene analyzed by RT-PCR after 24 h treatment with various combinations in (a, b) HL-60, KG-1, and THP-1 cell lines and patient-derived PBMCs and BMMCs (c, d). The values are normalized to the β-actin mRNA expression level, and then the relatively targeted genes’ mRNA levels are determined by taking the ratio to the value of the untreated cell as a calibrator based on the ΔΔCTT method. Cell lines data are presented as means ± SD of triplicate experiments. Primary AML patient PBMCs and BMMCs samples data are presented as means of individual patients experiments (n = 11). P-values < 0.05 (*), P-values < 0.01 (**), and P-values < 0.001 (***)
Codelivery of anti-HIF-1a/HO-1 siRNAs and Ara-c induced oxidative stress in leukemic cells
We also evaluated the effect of treatments on the production of ROS in cell lines (Fig. 6a) and primary cells (Fig. 6b), as Ara-c is a strong inducer of ROS generation in AML cells leading to cell apoptosis.

Treating cells with different pharmacological groups affects ROS formation as well as the proliferation potential to. The effect of 24-hour treatment with various treatment groups on ROS production was measured by DCF-DA in HL-60, KG-1, and THP-1 cells (a) and patient-derived PBMCs and BMMCs (b). Correlations between caspase-9 activity and the ROS levels in BMMCs (c) and PBMCs (d) upon exposure to various treatments. Blue dots are the means of 11 replicates. Pearson’s correlation coefficients are indicated on each plot. The solid line illustrates the regression line. (e) The result of single and combinational treatments on the proliferation of AML cell lines following 24 h incubation. Data were means ± SD (n = 3); *P < 0.05, **P < 0.01, ***P < 0.001, compared to untreated cells
The results showed that NP-Ara-c treatment induced higher levels of intercellular ROS compared to free Ara-c treatment. In addition, inhibition of HO-1 and HIF-1α increased the generation of ROS as well. The effect of HO-1 inhibition was slightly higher compared to HIF-1α silencing. Silencing of each gene synergistically induced ROS generation with Ara-c treatment. The highest levels of ROS generation were observed in cells treated with NPs loaded with siRNAs against HIF-1α and HO-1 and Ara-c.
Regarding the association of ROS production and apoptosis, we then evaluated the correlation between ROS generation and caspase-9 activity in AML primary cells (Fig. 6c and d).
As demonstrated by the graphs, a very strong correlation (r = 93, r = 94) exists between ROS production and caspase-9 activity in AML PBMCs as well as BMMCs, indicating that ROS generation is a major contributor to programmed cell death of AML cells.
The combinational therapy inhibited the proliferation of AML cells
The proliferation of AML cell lines in response to the various combinational treatments was also assessed (Fig. 6e). Treating cells with each siRNA exerted a slight effect on the proliferation of cells, while treatment with both siRNAs inhibited proliferation considerably, which was comparable with free Ara-c treatment. Another finding was that the effect of HIF-1α inhibition on proliferation was slightly higher compared to HO-1 inhibition. Moreover, compared to treatment with free Ara-c, cells treated with NP-Ara-c demonstrated a more significant inability to proliferate. The inhibition of each gene seemed to form a synergism with Ara-c to inhibit proliferation. The highest level of proliferation inhibition was noted in the group receiving both siRNAs and Ara-c-loaded NPs.
Discussion
AML as one of the main hematological malignancies is associated with frequent relapses and poor prognosis, especially in the elderly despite the use of intensive chemotherapies [5].
In addition to relapse, chemoresistance is another clinical issue emerging post-AML treatment [32].
Moreover, Ara-c/DNR (the main chemotherapy regimen for AML) doses are highly toxic and have plenty of side effects [33]. Thus, the need for novel therapeutic approaches to reverse this drug resistance and reduce the required dose of chemotherapy to efficiently kill tumor cells is always felt.
Nowadays, combination therapy and simultaneous manipulation of two genes is a very common method in such studies. A large part of the studies investigate the effect of silencing several genes together and alone, to compare the effect of simultaneous inhibition of the genes with inhibition alone [34]. Combination gene therapy cooperates with chemotherapy to ensure the maximum death of tumor cells and decrease the possibility of relapse. Combination gene therapy overcomes the limitations of chemotherapy, including multidrug resistance, high drug dose, side effects, etc. To be able to successfully use combination gene therapy in clinics, certain issues need to be solved. First, the long-term effects of this treatment must be studied thoroughly to ensure its safety. Second, safe vectors are required to efficiently deliver the cargo to the target cells. This makes nanoparticles a new promising tool for this aim [35]. Therefore, combined gene therapy with chemotherapy strategy was investigated on AML cells in this study.
HO-1 is an inducible enzyme, that catabolizes heme into biliverdin, iron, and carbon monoxide [36].
According to several studies, HO-1 is reported to be upregulated in AML cell lines and primary cells of AML patients. The overexpression of HO-1 in AML patients is associated with poor prognosis [6, 37, 38]. HO-1 promotes resistance to chemotherapy using a variety of pathways. Neutralizing ROS [6], induction of the JNK/c-jun pathway to inhibit apoptosis [38], and many other mechanisms are examples of HO-1 anti-apoptotic functions. Moreover, HO-1 can exert cytoprotective activities by its products [39], including the induction of ferritin heavy chain transcription and Fe efflux pump function, induced by Fe2+ atom released from catabolized heme by HO-1, resulting in decreased ROS formation [40,41,42,43]. In addition to Fe, biliverdin, which is then converted to bilirubin ( a strong antioxidant molecule) can protect tumor cells from ROS, and also the anti-inflammatory effects of bilirubin (due to its effect on inhibiting the expression of P and E selectins) [44,45,46] are several mechanisms to name a few. In addition to leukemic cells, HO-1 is reported to be overexpressed in CD34+CD38- as well as CD34+CD38- cells (both stem cells and progenitor cells) of AML patients compared to normal CD34+ cells, which brings to mind the idea of possible benefit of HO-1 silencing in eliminating the Minimal Residual Disease (MRD) [3].
HIF-1α is a well-known regulator of angiogenesis and proliferation. HIF-1α is degraded in normoxic conditions by the ubiquitin proteasomal system due to proline residue hydroxylation, while preserved from degradation in hypoxic conditions [47, 48]. However, this is not completely certain. Since it has been shown that in addition to hypoxia, other molecular mechanisms can regulate the levels of this protein in even normal cells [49]. In normal cells, HIF-1α is proven to be activated by the signaling of Phosphoinositide 3-kinase activity/ protein kinase B (PI3K/AKT) [50, 51], and CDK2 activity, through blockade of lysosomal HIF-1α degradation in normoxia [52]. Additionally, CDK5 activity has proven to elevate HIF-1α protein levels in cells by stabilizing phosphorylation of HIF-1α at Ser687 mediated mechanism [53]. Also, it has been reported that HIF-1α can be stabilized by a ROS-mediated mechanism in normoxic conditions [54]. Besides oxygen-independent HIF-1α stabilization in normal cells, under oncogenic mutation or inactivation of anti-oncogenes, HIF-1α can be activated under normoxic conditions as well [55,56,57]. Mutations of oncogenic genes including P53, RB, Ras, ARF, and Myc are proven to stabilize HIF-1α in cancer cells [58].
In AML in particular, several other mechanisms have also been proposed to explain the reason behind the accumulation of HIF-1α in cells in normoxia. In 10–15% of AML cases, the mutations of Isocitrate Dehydrogenase (IDH) 1 and 2 result in PHD inactivation, therefore the suppression of HIF-1α hydroxylation [59]. Moreover, FMS-Like Tyrosine kinase 3 (FLT3) mutations which are found at a frequency of 25–355 in AML, activate PI3K/AKT in the cells [60, 61], resulting in the activation of Mammalian Target Of Rapamycin (mTOR) and subsequently the accumulation of HIF-1α in the cells [62, 63]. Additionally, Nucleophosmin (NPM1) mutations (observed in 30–40% of AML cases) can induce P14ARF degradation, which in turn stimulates HIF-1α activity [59]. Finally, the accumulation of HIF-1α in AML cells could be due to the P53 loss of function (observed in 10% of AML) resulting in decreased expression of Mouse double minute 2 homolog (MDM2 which is an alternative E3 ubiquitinylation ligase for HIF-1α [59].
Therefore, HIF-1α is overexpressed in many cancers and AML [17, 64,65,66]. HIF-1α induces the expression of many genes related to chemoresistance, including GLUT-1, HO-1, VEGF, etc [1, 18]. , making HIF-α an ideal target for cancer therapy [67].
Based on several studies, HO-1 and HIF-1α are overexpressed in AML cells. This overexpression is associated with resistance to DNR, and Ara-c [1, 2, 17, 20]. Based on another study, HIF-1α is overexpressed in AML cells that possess mutant TP53 gene (an indicator for poor prognosis in AML, and poor response to chemotherapy) compared to AML cells with wild TP53 gene (an indicator of ideal prognosis) [68].
For the reasons stated above, as well as the proven role of both molecules in various cancers, we selected these two genes to assess the effect of their simultaneous inhibition in response to Ara-c treatment, a main chemotherapy regimen used for AML treatment. To silence the expression of these genes, we used siRNA molecules. Due to the instability of siRNA in the extracellular environment, we used NPs to transport siRNA into cells. The usage of NPs to deliver drugs is very efficient in chemotherapy [69].
In this study, TAT-CCMD-PEG-NPs were utilized to transport siRNA and Ara-c to the cells. The usage of CCMD improves the loading and delivery of NPs, while binding NPs via PEG to TAT, increases the cellular uptake by cells. We designed these NPs and performed all the required tests to characterize TAT-CCMD-PEG-NPs (including transfection and cargo delivery efficiency and other required tests as well) in our recent research [22].
Based on the MTT results, it can be seen that following the silencing of HO-1, the rate of growth in response to Ara-c treatment was greatly decreased. The reason behind this sensitivity following the inhibition of HO-1 may be due to the fact that Ara-c induces ROS production, which is lethal to the cell. But this ROS is neutralized by the overexpressed HO-1 (induced by high ROS levels), which is why cell treatment with Ara-c induces HO-1 expression, and HL-60R cells that are resistant to Ara-c have higher levels of HO-1 expression [1]. Thus, we evaluated the level of ROS generated following the inhibition of each gene as well as the treatment with Ara-c. We found that HO-1 inhibition induced the repletion of ROS in AML cells following Ara-c treatment.
HIF-1α inhibition also induced ROS generation, (probably because HIF-1α regulates HO-1 expression as well as other stress responder genes, and its silencing can affect the expression of these genes) but to a lesser amount. The highest level of ROS was observed following the inhibition of both genes and treatment with Ara-c.
Following the inhibition of the HIF-1α alone, the rate of growth inhibition was higher than in the case in which the HO-1 gene was silenced. This is probably due to the fact that HIF-1α controls more genes that are involved in Ara-c resistance. However, the highest rate of apoptosis was seen when the two genes were inhibited simultaneously by NPs and cells were treated via Ara-c. We further studied the role of these genes in preventing cell apoptosis by studying the effect of HO-1 and HIF-1α silencing on apoptosis-related genes including Caspase-9, Bcl-2, and BAX. Inhibition of these two genes decreased Bcl-2 and increased BAX expression. This indicates the role of HO-1 and HIF-1α in preventing apoptosis. Following the inhibition of either of the genes, the activity of Caspase-9 increased. Therefore, the co-inhibition of HIF-1α and HO-1 synergizes with Ara-c treatment and activates the intrinsic apoptotic pathway. The effect of HIF-1α inhibition was slightly higher than HO-1 inhibition, this may be also due to the regulation of several anti-apoptotic genes by HIF-1α. Our study indicates that HO-1 and HIF-1α contribute to Ara-c resistance in AML, and silencing them could overcome this resistance possibly by increasing ROS levels in AML cells, thus resulting in apoptosis.
Based on Pearson’s correlation results, a positive connection was noted between caspase-9 concentration and ROS levels in the cells, following the treatment with various combinational treatment groups, indicating that the elevated levels of ROS in cells induced by HO-1 and HIF-1α silencing as well as treatment with Ara-c, contribute in the induction of caspase-9-dependant apoptosis. Therefore, a major mechanism synergistically being used to induce apoptosis by HO-1, HIF-1α silencing, and Ara-c treatment is the induction of ROS generation which in turn results in caspase-dependent cell death. However, several other mechanisms may be of importance as well.
We next evaluated the effect of treatment on the proliferation of cell lines and found that HIF-1α and HO-1 co-silencing could exert almost the same effect as free Ara-c did. Moreover, the effect of HIF-1α inhibition was slightly higher compared to the HO-1 inhibition result. However, cells with silenced HO-1 and HIF-1α combined with Ara-c treatment demonstrated the lowest level of proliferation.
Silencing both genes significantly improved apoptosis induction. Silencing HO-1 induced higher levels of ROS compared to HIF-1α silencing, while the data regarding the proliferation of AML cells was vice-versa.
Our results are in line with a study indicating that HO-1 inhibition sensitizes cells to Ara-c treatment. It is suggested that HO-1 silencing could sensitize AML cells to Ara-c by increasing intercellular ROS levels [1]. Based on the results of another study, transfection of AML-M2 cells with HO-1 siRNA decreased the viability of AML cells and improved the survival of AML mice models in response to DNR treatment [2]. Zhe et al. also reported that following the inhibition of HIF-1α by 2-methoxyestradiol, the survival, and proliferation of AML cells decreased more significantly compared to cells treated with Ara-c. They also showed that HIF-1α inhibition by this agent eliminated AML stem cells more efficiently than Ara-c. They found that this HIF-1α inhibitor induces ROS generation, which then triggers the mitochondrial apoptotic pathway [17].
Wang et al. also reported that treatment of TP53 mutated AML cells with echinomycin (a HIF-1α inhibitor) eliminates AML stem cells more efficiently than DNR + Ara-c treatment. Inhibition of HIF-1α also improved the survival of TP53-mutated AML murine models as well. Thus, HIF-1α inhibition could be beneficial even in eliminating MRD, due to its potential for selective elimination of leukemic stem cells [68].
Future studies can evaluate the effect of this combinational therapy with a higher number of patients, and compare the results in T-ALL and B-ALL groups, which was not possible in the current study due to the scarcity of samples. Moreover, the effect of this treatment on other cellular processes for example cell cycle, metabolism, etc. should be further studied as well. In the second step, the treatment could be used in AML xenograft models to help provide a more practical result and also evaluate. Finally, clinical trials could prove the applicability of the treatment in practice as well.
Conclusions
As far as we know, this was the first time that HO-1 and HIF-1α were simultaneously silenced using NPs loaded with siRNA to evaluate its effect on Ara-c chemoresistance. To sum up, while HO-1 mostly induces resistance to ROS, HIF-1α is involved in the induction of cell proliferation. Therefore, both genes contribute to chemoresistance and cosilencing of HO-1 and HIF-1α eliminates chemoresistance, proliferation potential, induces ROS generation, caspase-9 activity, and thus apoptosis. This study presents new hope for individuals afflicted with AML.
Data availability
No datasets were generated or analysed during the current study.
Abbreviations
- AML:
-
Acute Myeloid Leukemia
- Ara-c:
-
Cytarabine
- BAX:
-
Bcl-2 Associated X protein
- Bcl-2:
-
B-cell lymphoma-2
- BM:
-
Bone Marrow
- BMMC:
-
Mone Marrow Mononuclear Cells
- BrdU:
-
Bromodeoxyuridine
- CCMD:
-
Chitosan-Carboxymethyl Dextran
- CMDFA:
-
5,6-chloromethyl-2’,7’-dichlorodihydrofluorescein diacetate
- CXCR4:
-
CXC chemokine type 4 Receptor
- DNR:
-
Daunorubicin
- ELISA:
-
Enzyme-Linked ImmunoAssay
- GLUT − 1:
-
Glucose Transporter-1
- HIF-1α:
-
Hypoxia-Inducible Factor-1 alpha
- HO-1:
-
Heme Oxygenase-1
- HSCT:
-
Hematopoietic Stem Cell Transplantation
- Hsp32:
-
Heat shock protein 32
- MRD:
-
Minimal Residual Disease
- MTT:
-
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2 H-tetrazolium bromide
- NP:
-
NanoParticles
- PB:
-
Peripheral Blood
- PBMC:
-
Peripheral Blood Mononuclear Cells
- PEG:
-
Poly Ethylene Glycol
- ROS:
-
Reactive Oxygen Species
- SiRNA:
-
Small interfering RNA
- TAT:
-
Trans-Activator of Transcription
- VEGF:
-
Vascular Endothelial Growth Factor
References
Zhe N, Wang J, Chen S, Lin X, Chai Q, Zhang Y, Zhao J, Fang Q. Heme oxygenase-1 plays a crucial role in chemoresistance in acute myeloid leukemia. Hematology. 2015;20(7):384–91.
Wei S, Wang Y, Chai Q, Fang Q, Zhang Y, Wang J. In vivo and in vitro effects of heme oxygenase-1 silencing on the survival of acute myelocytic leukemia-M2 cells. Exp Ther Med. 2015;9(3):931–40.
Herrmann H, Kneidinger M, Cerny-Reiterer S, Rülicke T, Willmann M, Gleixner KV, Blatt K, Hörmann G, Peter B, Samorapoompichit P, et al. The Hsp32 inhibitors SMA-ZnPP and PEG-ZnPP exert major growth-inhibitory effects on D34+/CD38 + and CD34+/CD38- AML progenitor cells. Curr Cancer Drug Targets. 2012;12(1):51–63.
Tallman MS, Gilliland DG, Rowe JM. Drug therapy for acute myeloid leukemia. Blood. 2005;106(4):1154–63.
Walter RB, Estey EH. Management of older or unfit patients with acute myeloid leukemia. Leukemia. 2015;29(4):770–5.
Heasman SA, Zaitseva L, Bowles KM, Rushworth SA, Macewan DJ. Protection of acute myeloid leukaemia cells from apoptosis induced by front-line chemotherapeutics is mediated by haem oxygenase-1. Oncotarget. 2011;2(9):658–68.
Farge T, Saland E, de Toni F, Aroua N, Hosseini M, Perry R, Bosc C, Sugita M, Stuani L, Fraisse M, et al. Chemotherapy-resistant human acute myeloid leukemia cells are not enriched for leukemic stem cells but require oxidative metabolism. Cancer Discov. 2017;7(7):716–35.
Murphy T, Yee KWL. Cytarabine and daunorubicin for the treatment of acute myeloid leukemia. Expert Opin Pharmacother. 2017;18(16):1765–80.
Kanno S, Higurashi A, Watanabe Y, Shouji A, Asou K, Ishikawa M. Susceptibility to cytosine arabinoside (Ara-C)-induced cytotoxicity in human leukemia cell lines. Toxicol Lett. 2004;152(2):149–58.
Agúndez JA, García-Martín E, Martínez C, Benito-León J, Millán-Pascual J, Díaz-Sánchez M, Calleja P, Pisa D, Turpín-Fenoll L, Alonso-Navarro H, et al. Heme Oxygenase-1 and 2 common genetic variants and risk for multiple sclerosis. Sci Rep. 2016;6:20830.
Yang YC, Huang YT, Hsieh CW, Yang PM, Wung BS. Carbon monoxide induces heme oxygenase-1 to modulate STAT3 activation in endothelial cells via S-glutathionylation. PLoS ONE. 2014;9(7):e100677.
Cao L, Wang J, Ma D, Wang P, Zhang Y, Fang Q. Heme oxygenase-1 contributes to imatinib resistance by promoting autophagy in chronic myeloid leukemia through disrupting the mTOR signaling pathway. Biomed Pharmacother. 2016;78:30–8.
Chen S, Wang J, Fang Q, Gao R, Shi Q, Zhang H, Zhao J. Upregulated heme oxygenase-1 expression of mouse mesenchymal stem cells resists to chemotherapy-induced bone marrow suppression. Chin Med J (Engl). 2014;127(7):1310–6.
Hutt DM, Roth DM, Vignaud H, Cullin C, Bouchecareilh M. The histone deacetylase inhibitor, Vorinostat, represses hypoxia inducible factor 1 alpha expression through translational inhibition. PLoS ONE. 2014;9(8):e106224.
Fathi M, Bahmanpour S, Barshidi A, Rasouli H, Karoon Kiani F, Mahmoud Salehi Khesht A, Izadi S, Rashidi B, Kermanpour S, Mokhtarian R et al. Simultaneous blockade of TIGIT and HIF-1α induces synergistic anti-tumor effect and decreases the growth and development of cancer cells. Int Immunopharmacol 2021;101(Pt A):108288.
Budi HS, Izadi S, Timoshin A, Asl SH, Beyzai B, Ghaderpour A, Alian F, Eshaghi FS, Mousavi SM, Rafiee B, et al. Blockade of HIF-1α and STAT3 by hyaluronate-conjugated TAT-chitosan-SPION nanoparticles loaded with siRNA molecules prevents tumor growth. Nanomedicine. 2021;34:102373.
Zhe N, Chen S, Zhou Z, Liu P, Lin X, Yu M, Cheng B, Zhang Y, Wang J. HIF-1α inhibition by 2-methoxyestradiol induces cell death via activation of the mitochondrial apoptotic pathway in acute myeloid leukemia. Cancer Biol Ther. 2016;17(6):625–34.
Spégel P, Malmgren S, Sharoyko VV, Newsholme P, Koeck T, Mulder H. Metabolomic analyses reveal profound differences in glycolytic and tricarboxylic acid cycle metabolism in glucose-responsive and -unresponsive clonal β-cell lines. Biochem J. 2011;435(1):277–84.
Wilson WR, Hay MP. Targeting hypoxia in cancer therapy. Nat Rev Cancer. 2011;11(6):393–410.
Song K, Li M, Xu XJ, Xuan L, Huang GN, Song XL, Liu QF. HIF-1α and GLUT1 gene expression is associated with chemoresistance of acute myeloid leukemia. Asian Pac J Cancer Prev. 2014;15(4):1823–9.
Rozema DB, Lewis DL, Wakefield DH, Wong SC, Klein JJ, Roesch PL, Bertin SL, Reppen TW, Chu Q, Blokhin AV, et al. Dynamic PolyConjugates for targeted in vivo delivery of siRNA to hepatocytes. Proc Natl Acad Sci U S A. 2007;104(32):12982–7.
Hosseinpour Feizi AA, Vakili-Samiani S, Karpisheh V, Masjedi A, Izadi S, Adibfar S, Nikkhoo A, Hojjat-Farsangi M, Atyabi F, Joodi khanghah O, et al. Increased susceptibility to doxorubicin-induced cell death in acute lymphocytic leukemia cells by inhibiting serine/threonine WEE1 kinase expression using the chitosan-carboxymethyl dextran-polyethylene glycol-TAT nanoparticles. J Drug Deliv Sci Technol. 2022;77:103868.
Gao S, Dagnaes-Hansen F, Nielsen EJ, Wengel J, Besenbacher F, Howard KA, Kjems J. The effect of chemical modification and nanoparticle formulation on stability and biodistribution of siRNA in mice. Mol Ther. 2009;17(7):1225–33.
Young SW, Stenzel M, Yang JL. Nanoparticle-siRNA: a potential cancer therapy? Crit Rev Oncol Hematol. 2016;98:159–69.
Feng C, Zhang T, Pan C, Kang Q, Wang L, Liu X, Shang Q, Chen S, Hu T, Wang J. Heme oxygenase-1 inhibits the cytotoxicity of natural killer cells to acute myeloid leukemia by downregulating human leukocyte antigen-C. Cytotherapy 2023.
Huang Y, Zhang Y, Ding X, Liu S, Sun T. Osmolarity influences chondrocyte repair after injury in human articular cartilage. J Orthop Surg Res. 2015;10:19.
Pan W, Yang J, Wei J, Chen H, Ge Y, Zhang J, Wang Z, Zhou C, Yuan Q, Zhou L, et al. Functional BCL-2 regulatory genetic variants contribute to susceptibility of esophageal squamous cell carcinoma. Sci Rep. 2015;5:11833.
Shen T, Lyu D, Zhang M, Shang H, Lu Q. Dioscin alleviates Cardiac Dysfunction in Acute myocardial infarction via rescuing mitochondrial malfunction. Front Cardiovasc Med. 2022;9:783426.
Huang Y, Zhang Y, Ding X, Liu S, Sun T. Osmolarity influences chondrocyte repair after injury in human articular cartilage. J Orthop Surg Res. 2015;10(1):19.
Martini D, Trirè A, Breschi L, Mazzoni A, Teti G, Falconi M, Ruggeri A Jr. Dentin matrix protein 1 and dentin sialophosphoprotein in human sound and carious teeth: an immunohistochemical and colorimetric assay. Eur J Histochem. 2013;57(4):e32.
Joshi N, Hajizadeh F, Ansari Dezfouli E, Zekiy AO, Nabi Afjadi M, Mousavi SM, Hojjat-Farsangi M, Karpisheh V, Mahmoodpoor A, Hassannia H, et al. Silencing STAT3 enhances sensitivity of cancer cells to doxorubicin and inhibits tumor progression. Life Sci. 2021;275:119369.
Polednak AP. Recent improvement in completeness of incidence data on acute myeloid leukemia in US cancer registries. J Registry Manag. 2014;41(2):77–84.
Baker WJ, Royer GL Jr., Weiss RB. Cytarabine and neurologic toxicity. J Clin Oncol. 1991;9(4):679–93.
Malik MA, Wani MY, Hashmi AA. Chap. 1 - Combination therapy: Current status and future perspectives. In: Combination Therapy Against Multidrug Resistance Edited by Wani MY, Ahmad A: Academic Press; 2020:1–38.
Lin J, Wang X, Ni D, Chen Y, Chen C, Liu Y. Combinational Gene Therapy toward Cancer with Nanoplatform: strategies and principles. ACS Mater Au. 2023;3(6):584–99.
Willis D, Moore AR, Frederick R, Willoughby DA. Heme oxygenase: a novel target for the modulation of the inflammatory response. Nat Med. 1996;2(1):87–90.
Yu M, Wang J, Ma D, Chen S, Lin X, Fang Q, Zhe N. HO-1, RET and PML as possible markers for risk stratification of acute myelocytic leukemia and prognostic evaluation. Oncol Lett. 2015;10(5):3137–44.
Lin X, Fang Q, Chen S, Zhe N, Chai Q, Yu M, Zhang Y, Wang Z, Wang J. Heme oxygenase-1 suppresses the apoptosis of acute myeloid leukemia cells via the JNK/c-JUN signaling pathway. Leuk Res. 2015;39(5):544–52.
Brouard S, Otterbein LE, Anrather J, Tobiasch E, Bach FH, Choi AM, Soares MP. Carbon monoxide generated by heme oxygenase 1 suppresses endothelial cell apoptosis. J Exp Med. 2000;192(7):1015–26.
Gozzelino R, Jeney V, Soares MP. Mechanisms of cell protection by heme oxygenase-1. Annu Rev Pharmacol Toxicol. 2010;50:323–54.
Barañano DE, Wolosker H, Bae BI, Barrow RK, Snyder SH, Ferris CD. A mammalian iron ATPase induced by iron. J Biol Chem. 2000;275(20):15166–73.
Baker HM, Anderson BF, Baker EN. Dealing with iron: common structural principles in proteins that transport iron and heme. Proc Natl Acad Sci U S A. 2003;100(7):3579–83.
Balla G, Jacob HS, Balla J, Rosenberg M, Nath K, Apple F, Eaton JW, Vercellotti GM. Ferritin: a cytoprotective antioxidant strategem of endothelium. J Biol Chem. 1992;267(25):18148–53.
Singleton JW, Laster L. Biliverdin reductase of guinea pig liver. J Biol Chem. 1965;240(12):4780–9.
Dudnik LB, Khrapova NG. Characterization of bilirubin inhibitory properties in free radical oxidation reactions. Membr Cell Biol. 1998;12(2):233–40.
Stagg J, Galipeau J. Mechanisms of immune modulation by mesenchymal stromal cells and clinical translation. Curr Mol Med. 2013;13(5):856–67.
Ke Q, Costa M. Hypoxia-inducible factor-1 (HIF-1). Mol Pharmacol. 2006;70(5):1469–80.
Powis G, Kirkpatrick L. Hypoxia inducible factor-1alpha as a cancer drug target. Mol Cancer Ther. 2004;3(5):647–54.
Jun JC, Rathore A, Younas H, Gilkes D, Polotsky VY. Hypoxia-inducible factors and Cancer. Curr Sleep Med Rep. 2017;3(1):1–10.
He Q, Gao Z, Yin J, Zhang J, Yun Z, Ye J. Regulation of HIF-1{alpha} activity in adipose tissue by obesity-associated factors: adipogenesis, insulin, and hypoxia. Am J Physiol Endocrinol Metab. 2011;300(5):E877–885.
Déry MA, Michaud MD, Richard DE. Hypoxia-inducible factor 1: regulation by hypoxic and non-hypoxic activators. Int J Biochem Cell Biol. 2005;37(3):535–40.
Hubbi ME, Gilkes DM, Hu H, Kshitiz, Ahmed I, Semenza GL. Cyclin-dependent kinases regulate lysosomal degradation of hypoxia-inducible factor 1α to promote cell-cycle progression. Proc Natl Acad Sci U S A. 2014;111(32):E3325–3334.
Herzog J, Ehrlich SM, Pfitzer L, Liebl J, Fröhlich T, Arnold GJ, Mikulits W, Haider C, Vollmar AM, Zahler S. Cyclin-dependent kinase 5 stabilizes hypoxia-inducible factor-1α: a novel approach for inhibiting angiogenesis in hepatocellular carcinoma. Oncotarget. 2016;7(19):27108–21.
Movafagh S, Crook S, Vo K. Regulation of hypoxia-inducible factor-1a by reactive oxygen species: new developments in an old debate. J Cell Biochem. 2015;116(5):696–703.
Thornton RD, Lane P, Borghaei RC, Pease EA, Caro J, Mochan E. Interleukin 1 induces hypoxia-inducible factor 1 in human gingival and synovial fibroblasts. Biochem J. 2000;350(Pt 1):307–12.
Fukuda R, Hirota K, Fan F, Jung YD, Ellis LM, Semenza GL. Insulin-like growth factor 1 induces hypoxia-inducible factor 1-mediated vascular endothelial growth factor expression, which is dependent on MAP kinase and phosphatidylinositol 3-kinase signaling in colon cancer cells. J Biol Chem. 2002;277(41):38205–11.
Zundel W, Schindler C, Haas-Kogan D, Koong A, Kaper F, Chen E, Gottschalk AR, Ryan HE, Johnson RS, Jefferson AB, et al. Loss of PTEN facilitates HIF-1-mediated gene expression. Genes Dev. 2000;14(4):391–6.
Evan GI, Vousden KH. Proliferation, cell cycle and apoptosis in cancer. Nature. 2001;411(6835):342–8.
Peng G, Liu Y. Hypoxia-inducible factors in cancer stem cells and inflammation. Trends Pharmacol Sci. 2015;36(6):374–83.
Chen W, Drakos E, Grammatikakis I, Schlette EJ, Li J, Leventaki V, Staikou-Drakopoulou E, Patsouris E, Panayiotidis P, Medeiros LJ, et al. mTOR signaling is activated by FLT3 kinase and promotes survival of FLT3-mutated acute myeloid leukemia cells. Mol Cancer. 2010;9:292.
Sathaliyawala T, O’Gorman WE, Greter M, Bogunovic M, Konjufca V, Hou ZE, Nolan GP, Miller MJ, Merad M, Reizis B. Mammalian target of rapamycin controls dendritic cell development downstream of Flt3 ligand signaling. Immunity. 2010;33(4):597–606.
Brugarolas J, Lei K, Hurley RL, Manning BD, Reiling JH, Hafen E, Witters LA, Ellisen LW, Kaelin WG Jr. Regulation of mTOR function in response to hypoxia by REDD1 and the TSC1/TSC2 tumor suppressor complex. Genes Dev. 2004;18(23):2893–904.
Van de Velde S, Hogan MF, Montminy M. mTOR links incretin signaling to HIF induction in pancreatic beta cells. Proc Natl Acad Sci U S A. 2011;108(41):16876–82.
Semenza GL. Hypoxia-inducible factors: mediators of cancer progression and targets for cancer therapy. Trends Pharmacol Sci. 2012;33(4):207–14.
Schmid T, Zhou J, Brüne B. HIF-1 and p53: communication of transcription factors under hypoxia. J Cell Mol Med. 2004;8(4):423–31.
Stiehl DP, Wirthner R, Köditz J, Spielmann P, Camenisch G, Wenger RH. Increased prolyl 4-hydroxylase domain proteins compensate for decreased oxygen levels. Evidence for an autoregulatory oxygen-sensing system. J Biol Chem. 2006;281(33):23482–91.
Semenza GL. Targeting HIF-1 for cancer therapy. Nat Rev Cancer. 2003;3(10):721–32.
Wang Y, Liu Y, Bailey C, Zhang H, He M, Sun D, Zhang P, Parkin B, Baer MR, Zheng P, et al. Therapeutic targeting of TP53-mutated acute myeloid leukemia by inhibiting HIF-1α with echinomycin. Oncogene. 2020;39(14):3015–27.
Wang AZ, Langer R, Farokhzad OC. Nanoparticle delivery of cancer drugs. Annu Rev Med. 2012;63:185–98.
Acknowledgements
We appreciate the monetary support of the following research provided by the Tabriz University of Medical Sciences.
Funding
The study was funded by the Tabriz University of Medical Sciences (grant number: 69339 and 70310).
Author information
Authors and Affiliations
Contributions
M.S. contributed to the conceptualization, methodology, data analysis, and writing the original draft. AA.MA. administered the project and contributed to the conceptualization, methodology, and data analysis. F.JN. reviewed and edited the manuscript, analyzed the data, and participated in the material collection. V.K. prepared the figures and contributed to the software process. A.K. contributed to the methodology and software process and was a major contributor to data analysis. M.R. contributed to sample collection and data analysis and reviewed the manuscript. A.Z. contributed to performing the experiments and the review and editing of the manuscript. B.R. and A.M. contributed to Formal analysis, Validation, Writing - review & editing. MH.F., H.K., A.M., F.KK., and M.Y reviewed and edited the manuscript and analyzed the data. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The following study was approved by the ethics code number of IR.TBZMED.VCR.REC.1401.029 by the Tabriz University of Medical Sciences ethical committee. Informed consent was obtained prior to sampling.
Competing interests
The authors declare no competing interests.
Conflict of interest
The authors declare that there are no conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Sadeghi, M., Moslehi, A., Kheiry, H. et al. The sensitivity of acute myeloid leukemia cells to cytarabine is increased by suppressing the expression of Heme oxygenase-1 and hypoxia-inducible factor 1-alpha. Cancer Cell Int 24, 217 (2024). https://doi.org/10.1186/s12935-024-03393-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-024-03393-3