
Abstract
Gastric cancer (GC) is the fifth most common malignancy and the third leading cause of cancer-related deaths worldwide. Similar to other types of tumors, GC cells undergo metabolic reprogramming and switch to a “predominantly glycolytic” metabolic pattern to promote its survival and metastasis, also known as “the Warburg effect”, which is characterized by enhanced glucose uptake and lactate production. A large number of studies have shown that targeting cancer cells to enhanced glycolysis is a promising strategy, that can make cancer cells more susceptible to other conventional treatment methods of treatment, including chemotherapy, radiotherapy and immunotherapy, and so on. Therefore, this review summarizes the metabolic characteristics of glycolysis in GC cells and focuses on how abnormal lactate concentration can lead to immunosuppression through its effects on the differentiation, metabolism, and function of infiltrating immune cells, and how targeting this phenomenon may be a potential strategy to improve the therapeutic efficacy of GC.
Similar content being viewed by others

Introduction
In the 1920s, Otto Warburg first showed that, unlike normal cells, which catabolize glucose by oxidative phosphorylation (OXPHOS) in the mitochondria, tumor cells tend to convert glucose to lactate even in conditions of sufficient oxygen [1]. This phenomenon was termed aerobic glycolysis or the Warburg effect and is characterized by enhanced glucose uptake and lactate production. Although the adenosine triphosphate (ATP) production efficiency was low during aerobic glycolysis, it still took up to 50–70% of the ATP supply in different tumors [2]. Furthermore, the metabolic intermediates generated during aerobic glycolysis can be used for the biosynthesis of biomacromolecules used by the tumor to meet the demands for rapid growth [3]. The production of lactate also provided an acidic environment to aid the invasion and metastasis of cancer [4].
Gastric cancer (GC) is one of the most malignant tumors worldwide and remains a major health threat in Asia-Pacific regions presently, while its pathological mechanism is generally unknown [5]. Furthermore, despite advances in diagnosis and treatment, prognosis has not improved significantly in the past decade. The five-year survival rate in China is only 35.9%, and the mortality rate remains very high [6]. The occurrence, development, and metastasis of GC are inseparable from its survival environment—tumor microenvironment (TME). A compelling body of research has demonstrated that the stomach has a strongly acidic environment and a unique endocrine system, which is very suitable for promoting tumor progression and metastasis [7]. GC originates within gastric epithelial cells, and similar to other tumor types, it demonstrates a Warburg effect. This involves heightened glucose uptake, intensified glycolysis, and the conversion of substantial pyruvate to lactic acid instead of OXPHOS for energy provision under aerobic conditions [1]. The metabolic alterations primarily driven by the Warburg effect have been termed metabolic reprogramming, enriching our comprehension of tumor cell metabolism. Concurrently, tumor cell glycolysis yields significant lactic acid, resulting in an acidic immunosuppressive microenvironment. This, in turn, exerts metabolic stress on infiltrating immune cells, fostering the development of immunosuppression and immune evasion [8, 9]. Further understanding of strategies to confine these adverse metabolites’ impact on immune cell function could facilitate the adept application of suitable immunotherapies, culminating in optimal therapeutic efficacy.
Glycolysis in GC
Cancer cells have a significant metabolic difference from normal cells due to the Warburg effect. Song et al. [10] used gas chromatography/mass spectrometry (GC/MS) to analyze the tissue metabolites of GC patients and healthy controls. GC/MS analysis revealed that several intermediates of aerobic glycolysis pathways, such as fumaric acid and alpha-ketoglutaric acid, showed a significant increase in cancer tissue compared to normal mucosa. This suggests that the modified glucose metabolism might serve as a pivotal parameter in delineating GC cells from their normal counterparts. In another investigation, Ikeda and colleagues [11] revealed elevated serum levels of 3-hydroxypropionic acid and pyruvic acid in GC patients. Likewise, abnormal glucose metabolism has been observed in GC tissue by various other researchers [2,3,4, 12]. Bhattacharya et al. [13] provided evidence that hypoglycemia and heightened glycolysis contribute to increased drug resistance in GC when exposed to chemotherapy. Lin et al [14] further confirmed that TIIA therapy inhibited cell growth and proliferation of GC by inhibiting glucose metabolism of cancer cells.
The association between cancer onset and the activation of proto-oncogenes along with the deactivation of tumor suppressor genes, intricately linked to glucose metabolism, is widely recognized. As a proto-oncogene, Myc plays a significant role in regulating glucose metabolism by increasing the expression of glycolytic enzymes, including glucose transporter 1 (GLUT1) [15], lactate dehydrogenase A (LDHA) [16, 17], and pyruvate Kinase M2 (PKM2) [18]. Simultaneously, the inhibition of p53, a well-known tumor suppressor, directly contributes to the Warburg effect. In various cancer types, the loss of p53 has been observed to promote the flow of glucose through the glycolytic pathway while reducing OXPHOS [18]. The p53 protein enhances OXPHOS while restraining glycolysis by suppressing the expression of GLUT1, GLUT3, and GLUT4 [19], as well as deactivating glycolytic enzymes like phosphoglycerate mutase (PGM) [20]. Compared to normal cells, which generate energy primarily through mitochondrial OXPHOS, cancer cells predominantly obtain energy through increased glycolysis even under aerobic conditions. Conversion of glucose to lactate via glycolysis is inefficient in ATP generation but produces a large number of intermediates that drive cell proliferation. Therefore, increased glucose consumption leading to anaerobic glycolysis is thought to provide an evolutionary advantage for cancer cells [21].
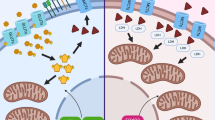
Moreover, several studies have indicated that glycolytic cancer cells offer further advantages for tumor growth by adapting selectively to the tumor TME. For example, extracellular accumulation of lactate from glycolysis alters the TME by creating an acidic pH that is harmful to normal cells. In general, lactate concentrations in normal serum are between 1.5 and 3 mM [22], while in tumor patients they increase to 10-30 mM and can even reach incredibly high levels (50 mM) in the inner tumor cores [23]. Numerous studies have illustrated that elevated lactate concentrations can be absorbed by various cell types and utilized as a metabolic resource within the TME [24,25,26]. Interestingly, cancer cells, encompassing both glycolytic and oxidative phenotypes, exhibit metabolic heterogeneity contingent upon their spatial distribution within the tumor [27]. Specifically, glycolytic cancer cells are located far from the vasculature, while oxidative cancer cells are located near the vasculature. Lactic acid and protons are co-exported through monocarboxylic transporters (MCTS) [24] to produce extracellular lactic acid. Cancer cells far away from blood vessels are in a state of hypoxia, which mainly obtains energy through glycolysis, and at the same time produces excessive lactic acid, which is exported to TME through MCT4 [28]. Cancer cells situated near blood vessels experience normal oxygen levels and can utilize lactate through MCT1 for ATP synthesis. This symbiotic lactate metabolism is not limited to cancer cells alone; it also involves other cell types like cancer-associated fibroblasts (CAFs) and tumor-associated endothelial cells [29,30,31]. Additionally, Gladden LB and colleagues demonstrated that inhibiting MCT1 disrupts glycolysis and hampers glutathione synthesis in tumor cells. This disruption elevates intracellular hydrogen peroxide levels, causing mitochondrial damage and eventually cell death [32]. Therefore, the transport of lactate between different cell populations emerges as a significant aspect of the TME, playing a crucial role in tumor initiation, progression, and metastasis (Fig. 1). Excessive lactate fosters the creation of an immunosuppressive environment that nurtures cancer cell growth and significantly influences immune cell functionality [33]. A growing body of research is directed toward unraveling the interplay between lactate and various immune cells within TME, to enhance the effectiveness of ongoing antitumor immunotherapies.

Regulation of the progress of lactate metabolism in cancer cells. Cancer cells, under hypoxic conditions, primarily derive energy through glycolysis while producing excessive lactate, which is exported to the TME through MCT4. The accumulation of lactate allows cancer cells to maintain an acidic microenvironment, shielding them from the effect of therapeutic agents. Additionally, cancer cells near blood vessels are normoxic and can oxidize lactate for ATP synthesis via MCT1. This lactate metabolic symbiosis occurs not only in cancer cells but also in other cells(e.g. immune cells)
Glycolysis mediates immunosuppression
Although lactate generated by glycolytic cancer cells has been linked to the suppression of anticancer immune cells, the precise mechanism underlying this inhibition has remained elusive. This lack of clarity primarily stems from the fact that immune cells themselves shift to glycolysis and generate lactate during their growth or maturation/activation processes [34, 35]. Consequently, exposure to lactate isn’t entirely novel to immune cells and, in itself, might not exhibit cytotoxic effects on them. Here, we will specifically discuss lactic-mediated antitumor immunity and focus on T cells, NK cells, Treg cells, and tumor-associated macrophages (TAMs). (Fig. 2)

Impact of lactate accumulation on immune cells within the TME. Within the TME, tumor cells predominantly consume nutrients while concurrently releasing excessive lactate, which leads to acidosis and immune suppression. Lactate further influences the metabolic activities of both innate and adaptive immune cells, impeding the functions of T cells, NK cells, and macrophages. Conversely, lactate promotes the sustenance of Treg cells, allowing them to maintain their immunosuppressive functions in the acidic milieu. Combining immunotherapy with medications targeting lactate production and lactate transporters can enhance the therapeutic efficacy of immune checkpoint inhibitors
T cells
T cell-mediated immunity constitutes the central component of the immune response against cancer. Within this category, various T cell subtypes exist, including CD4, CD8, helper T cells, memory T cells, and Tregs. Each subset of T cells possesses distinct functions, encompassing both anti-tumor activities and mechanisms of immune evasion. Among these, CD8+ T cells and Tregs are two specific lymphocyte types recognized for their roles in cancer immunity. T cells can perceive extracellular lactate levels, and this sensing process influences intracellular signaling, cellular function, and overall homeostasis. Elevated levels of lactic acid can hinder T cell-mediated immune responses. Within the TME, when the pH falls within the range of 6.0 to 6.5, activated effector CD8+ and CD4+ T cells typically lose their responsiveness. This results in a reduction in their cytolytic activity and cytokine production [30, 36, 37]. Another study further validated that lactate inhibited the production of interferon (IFN), tumor necrosis factor (TNF), and IL-2 triggered by the T cell receptor (TCR). It also impairs the function of cytotoxic T lymphocytes by suppressing the phosphorylation of the p38 signaling protein [38]. The CD155 molecule present on the surface of GC cells can interact with the immune checkpoint molecule TIGIT, which is found on the surface of T cells. This interaction hampers the glucose metabolism of T cells, diminishes the production of IFN-γ, and suppresses the cytotoxic activity of CD8+ T cells [39].
Understanding the regulatory mechanisms of T-cell immunity is crucial for immunotherapy against cancer cells. However, there are still many unknown factors in this complex microenvironment.
NK cells
As important effectors of host immunity, NK cells induce apoptosis of cancer cells by secreting IFN-γ [40], and TNF-α [41] or forming the Fas/FasL and TRAIL/TRAILR [31] complexes. NKG2D is an important receptor for NK activation, and MICA and MICB are known NKG2D inhibitory ligands that inhibit NK function [42, 43]. Numerous investigations have reported that GC cells can diminish the expression of NKG2D, thereby dampening the activity of NK cells. This is achieved by releasing soluble MICA and MICB molecules. For instance, Midkine has been shown to elevate CHOP expression and create complexes with the transcription factor AP-1. This, in turn, leads to increased MICA/B expression while concurrently inhibiting NK cytotoxicity [44]. STA21 was found to enhance the expression and release of MICB by inhibiting the STAT3 signaling pathway. Consequently, this suppression of STAT3 led to a reduction in NKG2D expression, impairing the function of NK cells [45]. On the other hand, inhibiting matrix metallopeptidase increased the expression of NKG2D ligand, consequently boosting NK activity in GC [46].
High concentrations of lactate have been reported to affect the cytotoxic activity of NK cells by decreasing intracellular pH and inducing apoptosis of NK cells [47,48,49]. Furthermore, lactate inhibits the activation of nuclear factor-activated T cells (NFAT) in NK cells, thereby inhibiting the production of IFN-γ [50]. Elevated lactate levels not only directly limit the cytolytic functions of NK cells but also indirectly inhibit NK cells by increasing the number of MDSCs [51]. Restoring NK cell function and cytotoxicity is the key to effective treatment of GC, which should be carefully considered in the combined treatment strategy.
Treg cells
However, not all immune cells in TME react negatively to lactate. Tregs make up about 5 to 10% of the peripheral blood and lymphocytes of humans and mice. Tregs are a potent immune system inhibitor responsible for maintaining immune homeostasis and preventing autoimmunity. The TME actively recruits and promotes the differentiation of Tregs by increasing the expression of forkhead box protein P3 (FOXP3) and MCT1 [52, 53]. The elevated presence of FOXP3 can suppress c-Myc and glycolysis, elevate OXPHOS, enhance NAD(+) oxidation, and reconfigure the metabolic profile of Treg cells. This adaptation makes Treg cells more resilient in low glucose and high lactic acid TME [37]. Research has indicated that FOXO4 expression is diminished in GC cells. Reinstating FOXO4 expression notably decreased the glycolysis rate in GC cells, whereas inhibiting FOXO4 expression led to an increase in the glycolysis rate [54]. Furthermore, MCT1-mediated lactating flux and intracellular lactate metabolism are crucial for tumor-infiltrating Treg cells to maintain their suppressive activity, while high glucose levels dampen their function and stability [30]. Furthermore, lactic acid is crucial for tumor-infiltrating Treg proliferation and function [30]. It was found that as the disease progressed, the accumulation of Treg cells in TME gradually increased, resulting in Treg cell imbalance in patients with GC. Furthermore, Treg cells help tumor cells escape host immune surveillance by secreting TGF, which promotes tumor progression [55].
TAMs
Cancer cells not only evade immune surveillance but also influence the physiology of selective immune cells to promote the rapid growth of tumors. Macrophages are the main component of tumor-infiltrating lymphocytes. TAMs play important roles in angiogenesis, metastasis, and immunosuppression. TAMs can be categorized into two subtypes: M1, also known as classical activated macrophages, and M2, referred to as activated macrophages. M1 macrophages exhibit anti-tumor activity, whereas M2 macrophages promote cancer development. In GC samples, there is a notable abundance of M2-TAMs, and they are strongly linked to invasion, metastasis, peritoneal dissemination, and unfavorable prognostic outcomes [56, 57]. They can change their character according to their environment. Tumor-derived lactate is reported to directly induce cancer-associated macrophages to become M2-like polarized cells, promoting tumor growth in the TME [58]. Mechanistically, the activation of extracellular signal-regulated kinase (ERK)/transcription 3 (STAT3), the activator of the STAT3 signaling pathway and the stimulated expression of vascular endothelial growth factor (VEGF) and arginase-1 (ARG1), as well as the stabilization of the hypoxic inducible factor 1 (HIF-1) contribute to lactate-induced polarization of M2 macrophages and its protumorigenic effects in breast cancer [59, 60]. Furthermore, tumor-derived lactate can induce TAM polarization to the immunosuppressive M2 phenotype by binding to the lactate-sensitive receptor-G protein-coupled receptor 132 (GPR132) [58, 59]. Overall, it is conceivable that the lactate-enriched environment within TME drives the re-education of TAMs to an M2 phenotype. Lactate significantly influences the metabolic reprogramming and immunomodulatory effects of macrophages, chiefly by promoting polarization shifts that detrimentally impact tumor immune responses. In contrast to the glycolytic metabolism that characterizes M1 macrophages, M2 TAMs primarily rely on OXPHOS to fulfill their bioenergetic requirements. Investigators have identified that Yes-associated protein 1 (YAP1) exhibits heightened expression in 5-FU-resistant GC tissues when compared to 5-FU-sensitive GC tissues [61]. Additionally, in GC with elevated YAP1 expression, IL-3 secretion has been observed to induce the polarization of macrophages toward an M2-like phenotype, prompting a GLUT3-dependent glycolytic program. Simultaneously, these polarized M2 macrophages contribute to increased resistance of tumor cells to 5-FU (an antimetabolite known as 5-fluorouracil) by secreting CCL8 and activating phosphorylation of the JAK1/STAT3 signaling pathway [61].
Endothelial cells and dendritic cells
Endothelial cells (ECs) reside in the innermost layer of blood vessels, regulating local vascular tension and permeability while coordinating with neighboring cells to modulate immune/inflammatory responses and blood supply. Tumor angiogenesis involves the proliferation of a vascular network that supports tumors with oxygen and nutrients. Research indicated that distinct metabolic features of ECs in cancer, and the function of vascular ECs can be modulated by metabolites [62]. ECs primarily generate ATP through glycolysis and rely on glucose for proliferation. This phenotype is driven by signaling pathways adjacent to tumors. For instance, conditioned media from hypoxic glioma cells induced the upregulation of surface GLUT1 in ECs, enhancing glucose uptake [63]. Additionally, signals from tumor-delivered VEGF led to the upregulation of 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3 (PFKFB3) in endothelial cells, activating PFK-1, further intensifying the glycolytic phenotype [64]. Lactate was also abundant in the TME, where it triggered tube formation in ECs through HIF-1α-dependent NF-κB activation [65].
Dendritic cells (DCs) are pivotal antigen-presenting cells capable of efficiently capturing, processing, and conveying antigenic information to CD8+ T cells. The activation of DCs involves metabolic reprogramming, transitioning from OXPHOS to aerobic glycolysis. Lactate inhibits the differentiation of DCs by inducing IL-10 production and leads to the loss of IL-12 in response to Toll-like receptor (TLR) stimulation [66]. Studies have revealed that lactate enhanced tryptophan breakdown metabolism and kynurenine generation in plasmacytoid DCs and immunosuppressive FoxP3+CD4+ regulatory T cells, resulting in immune suppression within the TME [67]. Simultaneously, high extracellular lactate concentrations inhibited lactate export by DCs, causing intracellular lactate accumulation, which affects the glycolytic process [68].
Therefore, the accumulation of lactic acid in tumors can inhibit the immune behavior of immune cells through a variety of ways and accelerate the immune escape of tumors.
Targeting glycolysis combined with immunotherapy in GC
Within tumors, the metabolites present in the TME exert a notable suppressive impact on immune cells, particularly anti-tumor T cells. Consequently, treatments focused on metabolic interventions that regulate glucose metabolism to enhance the TME’s conditions could serve as appealing adjunct therapies in combination with immune checkpoint inhibitors (ICIs). Gaining a deeper understanding of strategies to restrict the influence of these harmful metabolites on immune cell function will aid in optimizing the selection of immunotherapies to achieve maximal efficacy.
Recently, a large number of emerging research has emphasized the importance of neutralization in TME and its influence on the treatment outcome. Systematic and bicarbonate-buffered pH values have been shown to improve TME [69, 70]. More importantly, neutralizing of TME-pH through systemic buffering can enhance the outcome of anti-tumor immunotherapy. [71]. Recently, such neutralization efforts to reverse tumor acidosis have shown improved results in NK cell-mediated cancer therapy [72]. In melanoma patients, high LDH levels indicate a poor response to anti-PD-1 immunotherapy [73]. In a mouse melanoma model, an increase in IFN-γ and granzyme B production in NK cells and CD8+ T cells as well as an increase in PD-1 antitumor immune responses to immune checkpoint inhibitors have been reported by blocking LDHA can be observed [74]. Another study indicated that the use of the inhibitor GSK2837808A to suppress LDHA in both patient-derived and B16 melanoma cells can enhance the functionality of T cells both in vitro and in vivo [75]. This improvement contributed to the increased efficacy of immune cells, thereby enhancing adoptive cell therapy. In addition, lactate can also be reduced within the TME through targeted export. MCT 1 has the highest affinity for lactate and can import and export lactate based on a substrate concentration gradient, and the proton cancer MCT4 is expressed at higher levels by highly glycolytic tissues, including tumor cells [76]. In preclinical research, many MCT1 and MCT4 small molecule inhibitors, such as 7ACC2, AR-C155858, syrosingopine, and AZD3965, have been developed. However, only AstraZeneca’s AZD 3965 compound is currently being tested in humans (ClinicalTrials.gov NCT 01791595) [76]. Preclinical trials have shown that the combination of MCT1 inhibitor AZD3965 and anti-PD-1 therapy can reduce lactate secretion into the TME, reduce the infiltration of depleted PD-1+ Tim-3+ T cells in solid tumors, and improve antitumor immunity [77]. In a mouse model of hepatocellular carcinoma (HCC), overexpression of MCT4 leads to inhibition of CD 8+ T cell recruitment and reduced activity [78]. The utilization of an MCT4 inhibitor can efficiently halt the acidification process within the TME and stimulate the expression of chemokine (CXC motif) ligands, specifically CXCL9 and CXCL10, through the ROS/NF-κB signaling pathway. Additionally, in murine models of tumors, the inhibition of MCT4 substantially amplifies the therapeutic efficacy of PD-1 immune checkpoint inhibitors [72]. This suggests that the concurrent administration of MCT4 inhibitors could present a potential avenue for immunotherapy-resistant patients with HCC. Similarly, LDHA and MCT in GC cells express abnormally elevated and poor prognosis in patients with significant correlation [79]. Therefore, we speculate whether inhibition of LDHA and targeting MCT have similar effects on gastric tumors as described above. The research on this aspect, however, the current is less and deep enough.
GC has long been histologically classified into diffuse and intestinal types [80]. Recent studies have suggested potential metabolic differences between diffuse and intestinal subtypes of GC [81, 82]. Compared to the diffuse subtype, the intestinal subtype showed upregulation in glycolysis, which is potentially related to the lower expression of GLUT1 in diffuse gastric cancer [82, 83]. The proteome of diffuse gastric cancer was more enriched in metabolic pathways such as fatty acid metabolism, OXPHOS, and amino acid metabolism [81]. However, these differences are not fully elucidated at present, and further in-depth research is required to validate these potential metabolic disparities and determine their significance in the treatment of GC.
To date, glycolysis inhibitors are still in the preclinical stage, so their effect in humans is unclear. Currently, only a few therapeutic studies target GC metabolism. Further research is expected to investigate the role of cancer-specific glycolytic inhibitors in the development of effective therapeutic regimens for GC.
Conclusion
Modified glucose metabolism stands as a hallmark of GC, offering novel perspectives into GC development and the recognition of biomarkers targeting distinct metabolic facets of the condition. This review primarily focuses on how aberrant lactate concentrations influence tumor infiltration via diverse pathways of immune cell differentiation, metabolism, and function. The discovery of targets related to lactic acid metabolism in cancer immunomodulation has opened up new avenues for immunotherapy. A comprehensive understanding of how lactic acid metabolites influence the functionality of immune cells holds the potential to significantly enhance the effectiveness of immunotherapy. Despite some current obstacles in the clinical application of changes in lactic acid metabolism in immunotherapy for GC, it is anticipated that further in-depth research in these areas will facilitate the clinical utilization of its metabolites in GC treatment in the near future.
Data Availability
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
References
Warburg O. On the origin of Cancer cells. Science. 1956;123(3191):309–14.
Cai Z, Zhao J, Li J, Peng D, Wang X, Chen T, Qiu Y, Chen P, Li W, Xu L, et al. A combined proteomics and Metabolomics profiling of gastric Cardia Cancer reveals characteristic dysregulations in glucose Metabolism*. MOL CELL PROTEOMICS. 2010;9(12):2617–28.
Hirayama A, Kami K, Sugimoto M, Sugawara M, Toki N, Onozuka H, Kinoshita T, Saito N, Ochiai A, Tomita M, et al. Quantitative metabolome profiling of colon and Stomach cancer microenvironment by capillary electrophoresis time-of-flight mass spectrometry. CANCER RES. 2009;69(11):4918–25.
Hur H, Paik MJ, Xuan Y, Nguyen DT, Ham IH, Yun J, Cho YK, Lee G, Han SU. Quantitative measurement of organic acids in tissues from gastric cancer patients indicates increased glucose metabolism in gastric cancer. PLoS ONE. 2014;9(6):e98581.
Xiao S, Zhou L. Gastric cancer: metabolic and metabolomics perspectives (review). INT J ONCOL. 2017;51(1):5–17.
Glimelius B, Ekström K, Hoffman K, Graf W, Sjödén PO, Haglund U, Svensson C, Enander LK, Linné T, Sellsröm H, et al. Randomized comparison between chemotherapy plus best supportive care with best supportive care in advanced gastric cancer. ANN ONCOL. 1997;8(2):163–8.
Liu Y, Li C, Lu Y, Liu C, Yang W. Tumor microenvironment-mediated immune tolerance in development and treatment of gastric cancer. FRONT IMMUNOL. 2022;13:1016817.
Wang JX, Choi SYC, Niu X, Kang N, Xue H, Killam J, Wang Y. Lactic acid and an acidic Tumor Microenvironment suppress Anticancer Immunity. INT J MOL SCI. 2020;21(21):8363.
Wang Y, Wang D, Yang L, Zhang Y. Metabolic reprogramming in the immunosuppression of tumor-associated macrophages. Chin MED J-PEKING. 2022;135(20):2405–16.
Song H, Wang L, Liu HL, Wu XB, Wang HS, Liu ZH, Li Y, Diao DC, Chen HL, Peng JS. Tissue metabolomic fingerprinting reveals metabolic disorders associated with human gastric cancer morbidity. ONCOL REP. 2011;26(2):431–8.
Ikeda A, Nishiumi S, Shinohara M, Yoshie T, Hatano N, Okuno T, Bamba T, Fukusaki E, Takenawa T, Azuma T, et al. Serum metabolomics as a novel diagnostic approach for gastrointestinal cancer. BIOMED CHROMATOGR. 2012;26(5):548–58.
Wu H, Xue R, Tang Z, Deng C, Liu T, Zeng H, Sun Y, Shen X. Metabolomic investigation of gastric cancer tissue using gas chromatography/mass spectrometry. ANAL BIOANAL CHEM. 2010;396(4):1385–95.
Bhattacharya B, Low SHH, Soh C, Kamal Mustapa N, Beloueche Babari M, Koh KX, Loh J, Soong R. Increased drug resistance is associated with reduced glucose levels and an enhanced glycolysis phenotype. BRIT J PHARMACOL. 2014;171(13):3255–67.
Lin LL, Hsia CR, Hsu CL, Huang HC, Juan HF. Integrating transcriptomics and proteomics to show that tanshinone IIA suppresses cell growth by blocking glucose metabolism in gastric cancer cells. BMC Genomics. 2015;16(1):41.
Buller CL, Loberg RD, Fan M, Zhu Q, Park JL, Vesely E, Inoki K, Guan K, Brosius FC. A GSK-3/TSC2/mTOR pathway regulates glucose uptake and GLUT1 glucose transporter expression. AM J PHYSIOL-CELL PH. 2008;295(3):C836–43.
Ma J, Liu W, Guo H, Li S, Cao W, Du X, Lei S, Hou W, Xiong L, Yao L, et al. N-myc downstream-regulated gene 2 expression is associated with glucose transport and correlated with prognosis in breast carcinoma. BREAST CANCER RES. 2014;16(2):R27.
Qiu H, Jackson AL, Kilgore JE, Zhong Y, Chan LL, Gehrig PA, Zhou C, Bae-Jump VL. JQ1 suppresses Tumor growth through downregulating LDHA in Ovarian cancer*. Oncotarget. 2015;6(9):6915–30.
Wu H, Li Z, Yang P, Zhang L, Fan Y, Li Z. PKM2 depletion induces the compensation of glutaminolysis through β-catenin/c-Myc pathway in Tumor cells. CELL SIGNAL. 2014;26(11):2397–405.
Watanabe M, Naraba H, Sakyo T, Kitagawa T. DNA damage–Induced Modulation ofGLUT3 expression is mediated through p53-Independent Extracellular Signal-regulated kinase signaling in HeLa cells. MOL CANCER RES. 2010;8(11):1547–57.
RUIZ-LOZANO P, HIXON ML, WAGNER MW, FLORES AI, BALDWIN IKAWAS. CHIEN KR, GUALBERTO A: p53 is a transcriptional activator of the muscle-specific phosphoglycerate mutase gene and contributes in vivo to the control of its cardiac expression. Cell Growth Differ. 1999;10(5):295–306.
Cairns RA, Harris IS, Mak TW. Regulation of cancer cell metabolism. NAT REV CANCER. 2011;11(2):85–95.
Walenta S, Wetterling M, Lehrke M, Schwickert G, Sundfor K, Rofstad EK, Mueller-Klieser W. High lactate levels predict likelihood of metastases, Tumor recurrence, and restricted patient survival in human cervical cancers. CANCER RES. 2000;60(4):916–21.
Brizel DM, Schroeder T, Scher RL, Walenta S, Clough RW, Dewhirst MW, Mueller-Klieser W. Elevated Tumor lactate concentrations predict for an increased risk of metastases in head-and-neck cancer. INT J RADIAT ONCOL. 2001;51(2):349–53.
Roland CL, Arumugam T, Deng D, Liu SH, Philip B, Gomez S, Burns WR, Ramachandran V, Wang H, Cruz-Monserrate Z, et al. Cell surface lactate receptor GPR81 is crucial for Cancer Cell Survival. CANCER RES. 2014;74(18):5301–10.
Sonveaux P, Végran F, Schroeder T, Wergin MC, Verrax J, Rabbani ZN, De Saedeleer CJ, Kennedy KM, Diepart C, Jordan BF, et al. Targeting lactate-fueled respiration selectively kills hypoxic Tumor cells in mice. J CLIN INVEST. 2008;118(12):3930–42.
Draoui N, Feron O. Lactate shuttles at a glance: from physiological paradigms to anti-cancer treatments. DIS MODEL MECH. 2011;4(6):727–32.
Brown TP, Bhattacharjee P, Ramachandran S, Sivaprakasam S, Ristic B, Sikder MOF, Ganapathy V. The lactate receptor GPR81 promotes Breast cancer growth via a paracrine mechanism involving antigen-presenting cells in the Tumor microenvironment. Oncogene. 2020;39(16):3292–304.
Wilde L, Roche M, Domingo-Vidal M, Tanson K, Philp N, Curry J, Martinez-Outschoorn U. Metabolic coupling and the Reverse Warburg Effect in cancer: implications for novel biomarker and anticancer agent development. SEMIN ONCOL. 2017;44(3):198–203.
Payen VL, Mina E, Van Hée VF, Porporato PE, Sonveaux P. Monocarboxylate transporters in cancer. MOL METAB. 2020;33:48–66.
Watson MJ, Vignali P, Mullett SJ, Overacre-Delgoffe AE, Peralta RM, Grebinoski S, Menk AV, Rittenhouse NL, DePeaux K, Whetstone RD, et al. Metabolic support of tumour-infiltrating regulatory T cells by lactic acid. Nature. 2021;591(7851):645–51.
Kes MMG, Van den Bossche J, Griffioen AW, Huijbers EJM. Oncometabolites lactate and succinate drive pro-angiogenic macrophage response in tumors. Biochim et Biophys Acta (BBA) - Reviews Cancer. 2020;1874(2):188427.
GLADDEN LB. A lactatic perspective on metabolism. Med Sci Sports Exerc. 2008;40(3):477–85.
Ngwa VM, Edwards DN, Philip M, Chen J. Microenvironmental Metabolism regulates Antitumor Immunity. CANCER RES. 2019;79(16):4003–8.
Frauwirth KA, Riley JL, Harris MH, Parry RV, Rathmell JC, Plas DR, Elstrom RL, June CH, Thompson CB. The CD28 Signaling Pathway Regulates Glucose Metabolism. IMMUNITY 2002, 16(6):769–777.
Tan Z, Xie N, Banerjee S, Cui H, Fu M, Thannickal VJ, Liu G. The Monocarboxylate transporter 4 is required for Glycolytic Reprogramming and inflammatory response in Macrophages*. J BIOL CHEM. 2015;290(1):46–55.
Tu VY, Ayari A, O’Connor RS. Beyond the Lactate Paradox: How Lactate and Acidity Impact T Cell Therapies against Cancer. ANTIBODIES 2021, 10(3).
Angelin A, Gil-de-Gómez L, Dahiya S, Jiao J, Guo L, Levine MH, Wang Z, Quinn WJ, Kopinski PK, Wang L, et al. Foxp3 reprograms T cell metabolism to function in Low-Glucose, high-lactate environments. CELL METAB. 2017;25(6):1282–93.
Mendler AN, Hu B, Prinz PU, Kreutz M, Gottfried E, Noessner E. Tumor lactic acidosis suppresses CTL function by inhibition of p38 and JNK/c-Jun activation. INT J CANCER. 2012;131(3):633–40.
He W, Zhang H, Han F, Chen X, Lin R, Wang W, Qiu H, Zhuang Z, Liao Q, Zhang W, et al. CD155T/TIGIT signaling regulates CD8 + T-cell metabolism and promotes Tumor Progression in Human gastric Cancer. CANCER RES. 2017;77(22):6375–88.
Stetson DB, Mohrs M, Reinhardt RL, Baron JL, Wang Z, Gapin L, Kronenberg M, Locksley RM. Constitutive cytokine mRNAs Mark Natural Killer (NK) and NK T cells poised for Rapid Effector function. J Exp Med. 2003;198(7):1069–76.
Wajant H, Pfizenmaier K, Scheurich P. Tumor necrosis factor signaling. Cell Death & Differentiation. 2003;10(1):45–65.
Martín-Antonio B, Suñe G, Perez-Amill L, Castella M, Urbano-Ispizua A. Natural killer cells: angels and devils for Immunotherapy. INT J MOL SCI. 2017;18(9):1868.
Saito H, Osaki T, Ikeguchi M. Decreased NKG2D expression on NK cells correlates with impaired NK cell function in patients with gastric cancer. Gastric Cancer. 2012;15(1):27–33.
Zhao S, Wang H, Nie Y, Mi Q, Chen X, Hou Y. Midkine upregulates MICA/B expression in human gastric cancer cells and decreases natural killer cell cytotoxicity. Cancer Immunol Immunother. 2012;61(10):1745–53.
Garrido-Tapia M, Hernández CJ, Ascui G, Kramm K, Morales M, Gárate V, Zúñiga R, Bustamante M, Aguillón JC, Catalán D, et al. STAT3 inhibition by STA21 increases cell surface expression of MICB and the release of soluble MICB by gastric adenocarcinoma cells. Immunobiology. 2017;222(11):1043–51.
Shiraishi K, Mimura K, Kua LF, Koh V, Siang LK, Nakajima S, Fujii H, Shabbir A, Yong WP, So J, et al. Inhibition of MMP activity can restore NKG2D ligand expression in gastric cancer, leading to improved NK cell susceptibility. J GASTROENTEROL. 2016;51(12):1101–11.
Habif G, Crinier A, André P, Vivier E, Narni-Mancinelli E. Targeting natural killer cells in solid tumors. CELL MOL IMMUNOL. 2019;16(5):415–22.
Terrén I, Orrantia A, Vitallé J, Zenarruzabeitia O, Borrego F. NK Cell Metabolism and Tumor Microenvironment. FRONT IMMUNOL. 2019;10:2278.
Renner K, Singer K, Koehl GE, Geissler EK, Peter K, Siska PJ, Kreutz M. Metabolic hallmarks of Tumor and Immune cells in the Tumor Microenvironment. FRONT IMMUNOL. 2017;8:248.
Brand A, Singer K, Koehl GE, Kolitzus M, Schoenhammer G, Thiel A, Matos C, Bruss C, Klobuch S, Peter K, et al. LDHA-Associated Lactic Acid Production blunts Tumor Immunosurveillance by T and NK Cells. CELL METAB. 2016;24(5):657–71.
Husain Z, Seth P, Sukhatme VP. Tumor-derived lactate and myeloid-derived suppressor cells: linking metabolism to cancer immunology. ONCOIMMUNOLOGY. 2013;2(11):e26383.
Wang H, Franco F, Ho P. Metabolic regulation of Tregs in Cancer: opportunities for Immunotherapy. TRENDS CANCER. 2017;3(8):583–92.
Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and Immune Tolerance. Cell. 2008;133(5):775–87.
Wang XH, Jiang ZH, Yang HM, Zhang Y, Xu LH. Hypoxia-induced FOXO4/LDHA axis modulates gastric cancer cell glycolysis and progression. CLIN TRANSL MED 2021, 11(1).
Li Q, Li Q, Chen J, Liu Y, Zhao X, Tan B, Ai J, Zhang Z, Song J, Shan B. Prevalence of Th17 and Treg cells in gastric cancer patients and its correlation with clinical parameters. ONCOL REP. 2013;30(3):1215–22.
Wang H, Chen C, Yang C, Tsai I, Hou Y, Chen C, Shan Y. Tumor-Associated macrophages promote epigenetic silencing of Gelsolin through DNA methyltransferase 1 in gastric Cancer cells. CANCER IMMUNOL RES. 2017;5(10):885–97.
Yin S, Huang J, Li Z, Zhang J, Luo J, Lu C, Xu H, Xu H. The prognostic and clinicopathological significance of Tumor-Associated macrophages in patients with gastric Cancer: a Meta-analysis. PLoS ONE. 2017;12(1):e170042.
Colegio OR, Chu NQ, Szabo AL, Chu T, Rhebergen AM, Jairam V, Cyrus N, Brokowski CE, Eisenbarth SC, Phillips GM, et al. Functional polarization of tumour-associated macrophages by tumour-derived lactic acid. Nature. 2014;513(7519):559–63.
Chen P, Zuo H, Xiong H, Kolar MJ, Chu Q, Saghatelian A, Siegwart DJ, Wan Y. Gpr132 sensing of lactate mediates tumor-macrophage interplay to promote Breast cancer Metastasis. P NATL ACAD SCI USA. 2017;114(3):580–5.
Yang K, Xu J, Fan M, Tu F, Wang X, Ha T, Williams DL, Li C. Lactate suppresses macrophage pro-inflammatory response to LPS stimulation by inhibition of YAP and NF-κB activation via GPR81-Mediated signaling. FRONT IMMUNOL 2020, 11.
He Z, Chen D, Wu J, Sui C, Deng X, Zhang P, Chen Z, Liu D, Yu J, Shi J, et al. Yes associated protein 1 promotes resistance to 5-fluorouracil in gastric cancer by regulating GLUT3-dependent glycometabolism reprogramming of tumor-associated macrophages. ARCH BIOCHEM BIOPHYS. 2021;702:108838.
Zecchin A, Kalucka J, Dubois C, Carmeliet P. How endothelial cells adapt their metabolism to form vessels in tumors. FRONT IMMUNOL. 2017;8:1750.
Yeh W, Lin C, Fu W. Enhancement of glucose transporter expression of Brain endothelial cells by vascular endothelial growth factor derived from Glioma exposed to Hypoxia. MOL PHARMACOL. 2008;73(1):170–7.
Cantelmo AR, Conradi L, Brajic A, Goveia J, Kalucka J, Pircher A, Chaturvedi P, Hol J, Thienpont B, Teuwen L, et al. Inhibition of the glycolytic activator PFKFB3 in Endothelium induces Tumor Vessel normalization, Impairs Metastasis, and improves chemotherapy. Cancer Cell. 2016;30(6):968–85.
Carmona-Fontaine C, Deforet M, Akkari L, Thompson CB, Joyce JA, Xavier JB. Metabolic origins of spatial organization in the Tumor microenvironment. P NATL ACAD SCI USA. 2017;114(11):2934–9.
Nasi A, Fekete T, Krishnamurthy A, Snowden S, Rajnavolgyi E, Catrina AI, Wheelock CE, Vivar N, Rethi B. Dendritic cell reprogramming by endogenously produced lactic acid. J IMMUNOL. 2013;191(6):3090–9.
Raychaudhuri D, Bhattacharya R, Sinha BP, Liu C, Ghosh AR, Rahaman O, Bandopadhyay P, Sarif J, D’Rozario R, Paul S, et al. Lactate induces pro-tumor reprogramming in Intratumoral Plasmacytoid dendritic cells. FRONT IMMUNOL. 2019;10:1878.
Dong H, Bullock TNJ. Metabolic influences that regulate dendritic cell function in tumors. FRONT IMMUNOL. 2014;5:24.
Robey IF, Baggett BK, Kirkpatrick ND, Roe DJ, Dosescu J, Sloane BF, Hashim AI, Morse DL, Raghunand N, Gatenby RA, et al. Bicarbonate increases Tumor pH and inhibits spontaneous metastases. CANCER RES. 2009;69(6):2260–8.
SILVA AS, YUNES JA, GILLIES RJ. The potential role of systemic buffers in reducing Intratumoral Extracellular pH and acid-mediated Invasion. Cancer Res (Chicago Ill). 2009;69(6):2677–84.
Pilon-Thomas S, Kodumudi KN, El-Kenawi AE, Russell S, Weber AM, Luddy K, Damaghi M, Wojtkowiak JW, Mulé JJ, Ibrahim-Hashim A, et al. Neutralization of Tumor Acidity improves antitumor responses to Immunotherapy. CANCER RES. 2016;76(6):1381–90.
Potzl J, Roser D, Bankel L, Homberg N, Geishauser A, Brenner CD, Weigand M, Rocken M, Mocikat R. Reversal of Tumor acidosis by systemic buffering reactivates NK cells to express IFN-gamma and induces NK cell-dependent Lymphoma control without other immunotherapies. INT J CANCER. 2017;140(9):2125–33.
Daneshmandi S, Wegiel B, Seth P. Blockade of Lactate Dehydrogenase-A (LDH-A) Improves Efficacy of Anti-Programmed Cell Death-1 (PD-1) Therapy in Melanoma. CANCERS 2019, 11(4):450.
Cascone T, McKenzie JA, Mbofung RM, Punt S, Wang Z, Xu C, Williams LJ, Wang Z, Bristow CA, Carugo A, et al. Increased Tumor glycolysis characterizes Immune Resistance to adoptive T cell therapy. CELL METAB. 2018;27(5):977–87.
Gu J, Zhou J, Chen Q, Xu X, Gao J, Li X, Shao Q, Zhou B, Zhou H, Wei S, et al. Tumor metabolite lactate promotes tumorigenesis by modulating MOESIN lactylation and enhancing TGF-β signaling in regulatory T cells. CELL REP. 2022;39(12):110986.
Beloueche-Babari M, Casals GT, Delgado-Goni T, Wantuch S, Parkes HG, Tandy D, Harker JA, Leach MO. Monocarboxylate transporter 1 blockade with AZD3965 inhibits lipid biosynthesis and increases tumour immune cell infiltration. BRIT J CANCER. 2020;122(6):895–903.
Fang Y, Liu W, Tang Z, Ji X, Zhou Y, Song S, Tian M, Tao C, Huang R, Zhu G, et al. Monocarboxylate transporter 4 inhibition potentiates hepatocellular carcinoma immunotherapy through enhancing T cell infiltration and immune Attack. Hepatology. 2023;77(1):109–23.
Yang M, Zhong X, Yuan Y. Does Baking Soda function as a magic bullet for patients with Cancer? A Mini Review. Volume 19. Los Angeles, CA: SAGE Publications; 2020. p. 1872141011.
Zhao Z, Han F, He Y, Yang S, Hua L, Wu J, Zhan W. Stromal-epithelial metabolic coupling in gastric cancer: stromal MCT4 and mitochondrial TOMM20 as poor prognostic factors. Eur J Surg Oncol (EJSO). 2014;40(10):1361–8.
LAURÉN P: THE TWO HISTOLOGICAL MAIN TYPES OF GASTRIC CARCINOMA. DIFFUSE AND SO-CALLED INTESTINAL‐TYPE CARCINOMA. Acta Pathologica Microbiologica Scandinavica. 1965;64(1):31–49.
Ge S, Xia X, Ding C, Zhen B, Zhou Q, Feng J, Yuan J, Chen R, Li Y, Ge Z et al. A proteomic landscape of diffuse-type gastric cancer. NAT COMMUN 2018, 9(1).
Nam S, Lee Y. Genome-scale metabolic model analysis of metabolic differences between Lauren Diffuse and intestinal subtypes in gastric Cancer. CANCERS 2022, 14(9).
Yuan LW, Yamashita H, Seto Y. Glucose metabolism in gastric cancer: the cutting-edge. WORLD J GASTROENTERO. 2016;22(6):2046–59.
Acknowledgements
None.
Funding
This study was supported by the National Natural Science Foundation of China (No. 82174197, 81973609, 81973782,); Natural Science Foundation of Jiangsu Province, China (No. BK20211392); “333” Project of Jiangsu Province (LGY2018065); A Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD); Jiangsu Provincial Hospital of Traditional Chinese Medicine Academic Talent Program (Y2021RC29, Y2021RC45).
Author information
Authors and Affiliations
Contributions
S.QM. and W.J. created the original concept of this paper; Z.SS. and L.HZ. wrote the manuscript; L.YQ. and C.X. helped to write the manuscript; S.JY. and C.ML. create the figures; Z.CC. helped to revised it. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zheng, S., Li, H., Li, Y. et al. The emerging role of glycolysis and immune evasion in gastric cancer. Cancer Cell Int 23, 317 (2023). https://doi.org/10.1186/s12935-023-03169-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-023-03169-1