
Abstract
Matrix stiffness is a mechanical characteristic of the extracellular matrix (ECM) that increases from the tumor core to the tumor periphery in a gradient pattern in a variety of solid tumors and can promote proliferation, invasion, metastasis, drug resistance, and recurrence. Cancer stem cells (CSCs) are a rare subpopulation of tumor cells with self-renewal, asymmetric cell division, and differentiation capabilities. CSCs are thought to be responsible for metastasis, tumor recurrence, chemotherapy resistance, and consequently poor clinical outcomes. Evidence suggests that matrix stiffness can activate receptors and mechanosensor/mechanoregulator proteins such as integrin, FAK, and YAP, modulating the characteristics of tumor cells as well as CSCs through different molecular signaling pathways. A deeper understanding of the effect of matrix stiffness on CSCs characteristics could lead to development of innovative cancer therapies. In this review, we discuss how the stiffness of the ECM is sensed by the cells and how the cells respond to this environmental change as well as the effect of matrix stiffness on CSCs characteristics and also the key malignant processes such as proliferation and EMT. Then, we specifically focus on how increased matrix stiffness affects CSCs in breast, lung, liver, pancreatic, and colorectal cancers. We also discuss how the molecules responsible for increased matrix stiffness and the signaling pathways activated by the enhanced stiffness can be manipulated as a therapeutic strategy for cancer.
Similar content being viewed by others

Introduction
Solid tumors, as abnormal organs, are complex entities composed of heterogeneous populations of tumor cells and various types of stromal cells that produce soluble factors, signaling molecules, and extracellular matrix (ECM) components; which altogether can regulate tumor growth and progression and affect the response to treatment [1, 2]. The ECM is a three-dimensional network that mostly consists of macromolecules such as collagen, fibronectin, laminin, elastin, proteoglycans, and glycoproteins that provide structural and biochemical support to the cell [3]. During the progression of several solid tumors, deposition, remodeling, and crosslinking of the ECM composition alter and induce stiffening of the stroma from the tumor periphery to the tumor core in a gradient pattern [4, 5]. It has been demonstrated that high-grade invasive ductal carcinoma is 13-fold stiffer compared to normal human breast tissue [6]. Studies show that the stiffness of the ECM can effectively alter cell behavior at the cellular and molecular level through mechanosensing pathways [7]. Increased matrix stiffness appears to be a hallmark of solid tumor progression and metastasis. Considering the important function of the matrix stiffness in tumors, targeting the matrix stiffness has emerged as one of the next-generation therapies for cancer treatment [8, 9].
Cancer cells exhibit considerable heterogeneity in a variety of phenotypic and functional aspects [10]. Cancer stem cells (CSCs) are subpopulations of cancer cells that have similar characteristics to normal stem cells or progenitor cells. Recent findings suggest that cancer stem cells play a pivotal role in tumor initiation, progression, development, metastasis, resistance to treatment, and recurrence of cancer [11]. Also, there is evidence that the plasticity of tumors can lead to a dynamic variation in the relative abundance of CSCs and non-CSCs [12, 13]. Studies have shown that plasticity has significant implications for cancer therapies and cancers with a higher ratio of CSCs to non-CSCs are more resistant to chemotherapy [14]. On the other hand, after successful tumor resection, the remnant CSCs can lead to recurrence and be the culprit in certain forms of cancer cell dormancy, i.e. the state that the cells can remain dormant for many years, and suddenly awaking and causing overt recurrence and metastasis [15]. The interaction of non-CSCs with their surrounding microenvironmental niche contributes to their transformation into CSCs [16, 17].Among various components and signals of tumor stroma, matrix stiffness, arising from increased levels of collagen and enhanced crosslinking can have an impact on the formation, maintenance, and characteristics of CSCs[18, 19]. Higher matrix stiffness in tumors is associated with increased invasion and metastasis at least partly due to the increase in CSC population and markers [20, 21].
In this review, the effect of tumor matrix stiffness on some specific characteristics of CSCs including cell membrane CSC markers and tumor sphere formation (also used to enrich CSC/CSC like population) will be discussed. First, the molecular mechanisms by which stiffness of the matrix affects tumor cells are explored. Then effect of increased matrix stiffness on the progression and development of the tumor will be reviewed; and next, role of CSC in tumor progression, metastasis, drug resistance and recurrence will be explained. Following that, alteration in matrix stiffness in several solid tumors, including liver, breast, colorectal, lung and pancreas, and the effect of these changes on the special characteristics of CSCs will be reviewed. Finally, CSCs characteristics mediated by matrix stiffness alterations useful for obtaining novel insights into cancer biology will be discussed. Understanding how matrix stiffness regulates CSCs features and its functional consequences in cancer processes can represent a new perspective on cancer treatment.
Molecular mechanisms by which increased matrix stiffness influences cell characteristics
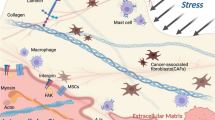
In many solid tumors, concurrent with the progression of the tumor, the accumulation of several ECM proteins leads to a gradual increase in matrix stiffness and ECM rearrangement [4]. Tumor cells and other tumor microenvironment (TME) cells, especially cancer-associated fibroblasts (CAF), produce collagen, which forms the majority of the tumor matrix and enhance the production of Lysyl oxidase (LOX), leading to collagen crosslinking, ECM rearrangement, and higher stiffness [22, 23]. Also, increased stiffness within tumors contributes to the incremental and continuous activation of CAFs, establishing a feed-forward loop that aids to the development of a permanent stiff tumor niche [24] (Fig. 1). In advanced stages of breast and colon cancers, the expression of collagen I, LOX has been found to be significantly higher in the TME, resulting in increased stiffness [8, 25]. In normal breast tissue, collagen fibrils are relaxed and non-oriented, whereas in breast cancer these fibers are usually thicker and aligned [26]. Stiffness is defined as the resistance of a material to deformation when a force is applied [27]. Several techniques have been used to measure the stiffness of tissue, and stiffness values can differ significantly between methods (Table 1). In atomic force microscopy (AFM), a tip enters the specimen and the cantilever beam flexes in response to the sample's stiffness. By combining the tip position, cantilever spring constant, and piezoelectric sensor measurements, the stiffness of the tissue can be determined at a microscale [28]. A compression test based on specimen indentation and rheometry can measure macroscale (mm) stiffness [29]. Shear wave elastography (SWE), as a higher accuracy method, uses acoustic radiation to induce mechanical vibrations and measures the stiffness of a tissue by capturing propagating shear waves [30].

Alterations in tumor matrix stiffness: normal organs are surrounded by irregularly thin collagen, which forms an ECM that is compliant and soft. In several solid tumors, the accumulation of ECM proteins causes a gradual rise in matrix stiffness parallel with the tumor’s growth. Tumor cells and other TME cells, particularly cancer-associated fibroblasts (CAF), produce collagen and Lysyl oxidase (LOX), resulting in collagen crosslinking, ECM rearrangement, and increased stiffness. In addition, increasing stiffness within tumors contributes to the continuous activation of CAFs, establishing a feed-forward loop that aids in the formation of a permanently stiff tumor niche. It is important to note that CSCs are not distributed uniformly across cancerous tissues. More CSCs are distributed in invasive areas to facilitate metastasis. The invasive tumor front (ITF) is stiffer than the tumor’s core
Through a process known as mechanosensing and mechanotransduction, tumor cells detect the stiffness index of the ECM and molecular effectors respond and transmit this signal, and then transform this information into biochemical signals that alter cellular behavior [31]. Integrin receptors respond to forces caused by increase in matrix stiffness. Each subunit of an integrin has a particular specificity for a particular ECM ligand, which can transmit the stiffness signal of the ECM into the cells via distinct mechanisms. Adapter molecules, such as focal kinase adhesion (FAK), accumulate in response to ligand binding to integrin receptors. The degree of matrix stiffness can regulate FAK activity and, consequently, the activation rate of several signaling pathways [32]. In these pathways, FAK can activate phosphatidylinositol-3-kinase (PI3K), serine/threonine-protein kinase (AKT), β-catenin, ERK, JNK, and other molecules, while inhibiting tumor suppressor genes such as phosphatase and tensin homolog (PTEN) and glycogen synthase kinase 3α/β (GSK3α/β) [33, 34]. Moreover, the cell can convey mechanical cues through the RhoA/Rho-associated protein kinase (ROCK) pathway [35]. Additionally, tumor stiffness influences tumor and stromal cells through the transcriptional activators yes-associated protein 1 (YAP1) and WW domain-containing transcription regulator 1 (WWDR1) (TAZ) [36]. In stiff environments, YAP and TAZ are activated and accumulate in the nucleus, whereas they are suppressed and localized in the cytoplasm in physiological stiffness [37]. YAP and TAZ are transcriptional coactivators lacking DNA-binding domain. Hence, these molecules must interact with DNA-binding transcription factors to regulate the expression of target genes. From there, based on the DNA-binding partner, tumorigenic and tumor suppressor genes can be expressed [38] (Fig. 2).

Matrix stiffness signaling pathways. Stiffness activates integrin-focal adhesion kinase (FAK) and activated FAK regulate several downstream mechanoresponsive signaling pathways. Pathways such as ERK, AKT, β-catenin, RhoA-ROCK, YAP/TAZ play major roles in stiffness mediated characteristics. In stiff ECM, YAP and TAZ are activated and accumulate in the nucleus, whereas in physiological stiffness, they are suppressed and localized in the cytoplasm. The transcriptional coactivators YAP and TAZ lack a DNA-binding domain. Hence, nuclear YAP/TAZ binds to TEAD and regulates the activation of several target genes involved in cell migration, proliferation, anti-apoptotic processes, and stemness
The importance of matrix stiffness in cancer progression, development, recurrence and treatment
Multiple studies have demonstrated the importance of matrix stiffness in physiological and pathological states [19, 39]. Cell adhesion, migration, proliferation, and differentiation can be regulated by stiffness [39,40,41]. Matrix stiffness is important in embryonic morphogenesis and development [42]. During tissue injury and aging, alterations in matrix stiffness also play a special role [43] and significantly contributes to tissue homeostasis and function [39]. If the homeostasis of stiffness is disturbed for any reasons, it may lead to tissue dysfunction and the associated to pathologic conditions such as cancer [44].
In a number of solid tumors, increased matrix stiffness can affect tumor progression, metastasis, and therapeutic response [31, 45] (Fig. 3). It’s important to note that the effect of tumor matrix stiffness on tumor progression and metastasis is a multifaceted phenomenon. Matrix stiffness in the primary tumor promotes the formation of CAFs, which in turn increases collagen and LOX synthesis, generating a positive feedback loop that promotes tumor progression and metastasis [45]. Interestingly, in terms of stiffness, tumors are heterogenous; for instance, in breast cancer, stiffness gradually increases from the core to the periphery [4, 46]. Different local maps of ECM stiffness could dictate distinct cancer cell functions during tumor progression. Breast cancer cells grown in a matrix similar to tumor's core stiffness exhibit increased proliferation, rise in glycolysis rate, and the high tumor formation potential; whereas, tumor cells grown in a matrix with variable stiffness that matched the peripheral zones of breast tumors show increased fibronectin 1 (FN1) and matrix metalloproteinase 9 (MMP9) expression, migration, oxidative phosphorylation (OXPHOS) and fatty acid (FA) metabolism processes, and also angiogenesis [4].

Several functions of increased matrix stiffness in cancer. Increased matrix stiffness can have an effect on tumor cell proliferation, vascular permeability, invasion and migration, metastasis, angiogenesis, immune evasion, treatment resistance, and recurrence
Tumorigenesis in breast is associated with ECM stiffening, stiffness induces the formation of integrin-FAK, leading to ROCK-generated contractility and promoting a malignant phenotype [47]. When cell–cell adhesions decreases due to the increased stiffness of the matrix, nuclear-activated YAP/TAZ binds to the TEAD and regulate the activation of several target genes involved in cell migration, proliferation, anti-apoptotic processes, and stemness [38, 48]. Most of the functions of YAP and TAZ include promoting sustained proliferation by expression of proto-oncogenes such as MYC and AP-1 family, transcriptionally upregulating the enzymes involved in metabolic requirements to support proliferating cancer cells [42], controlling the expression of cell cycle regulators, DNA replication and repair, and mitosis [49, 50]. YAP/TAZ can reprogram non-CSCs into CSCs [51]. This indicates that YAP/TAZ can modify the proportion of CSCs present in tumor tissues [52]. In addition, YAP is a critical molecule in the maintenance of CSCs in a variety of tumor types [53]. YAP/TAZ activation contributes to the induction of resistance to MAPK pathway–targeted therapies (RAF and MEK inhibitors) [54]. YAP/TAZ are involved in metastasis through several mechanisms, one of which is responsible for anoikis- resistance of circulating tumor cells (CTCs) [55, 56].The nuclear accumulation of YAP/TAZ can modify E/N cadherin and vimentin expression in response to stiffness, inducing epithelial–mesenchymal transition (EMT), a prerequisite for invasion and metastasis [57,58,59]. It is well known that the induction of EMT in carcinoma cells, produce stem cell-like cells [60], and would increase stem-like cell features [61]. Also, the stiffened ECM can exert a physical force on basement membrane to generate permeable pores, facilitating the invasion of CSCs [62].
Collagen fiber properties and organization (length, alignment, etc.), as stiffness characteristics can also be used as a prognostic marker and innovative paradigm for cancer metastasis and survival prediction [63, 64]. Enhanced stiffness -through a variety of mechanisms- can result in chemotherapeutic drug resistance [65,66,67]. Increased stiffness can establish a barrier and increase the interstitial fluid pressure within the tumor; thereby, it limits access, impairs perfusion and prevents drug delivery [68]. Matrix stiffness can induce EMT in pancreatic cancer cell lines and contribute to the role of EMT in chemotherapeutic drug resistance [41]. In a variety of cancers, stiffness serves as a predictor of chemotherapy response, so that the softer tumors are more drug-sensitive [69,70,71]. Measuring the stiffness of the liver prior to curative resection could be predictive of hepatocellular carcinoma (HCC) recurrence [72]. A stiffer matrix can upregulate osteopontin (OPN) expression in HCC cells through a Wnt-independent-β-catenin pathway (OPN is a molecule strongly associated with metastasis, early recurrence, and poor prognosis) [73]. EMT induction in immortalized human mammary epithelial cells results in the expression of stem cell markers [60]. This raises the possibility that the EMT-mediated stiffness established by surgery and scar formation may confer self-renewal capacity to epithelial cells, hence promoting cancer recurrence. Also, immune modulation is significantly affected by the matrix stiffness and the expression of programmed cell death-ligand 1 (PD-L1) [74]. The orientation, spacing, and density of collagen fibrils in the stroma can also affect the distribution and migration of CD8 T cells [75]. In addition to its effects on the primary tumor, matrix stiffness can influence the formation of the metastatic niche and the rate of metastasis [76,77,78]. Chu et al. revealed that matrix stiffness can regulate cellular adhesion and promote breast cancer cell homing in premetastatic niches [79] (Fig. 3).
Moreover, matrix stiffness indirectly affects tumor cell behavior through exosomes. Using stiffness-tunable scaffolds, Patwardhan et al. found that stiff ECMs promote exosome secretion in a YAP/TAZ pathway-dependent manner. Stiffness-mediated secreted exosomes promote cell motility and invasion. Based on genomic and proteomic profiling of secreted exosomes, thrombospondin-1 (THBS1) was identified as a regulator of tumor invasion, dependent on the stiffness of the tumor. THBS1 levels per exosome were significantly higher in stiff ECMs secreted exosomes, which were amplified by the greater total number of exosomes in stiff scaffolds. According to knockdown experiments, the pro-invasive effects of stiffness-tuned exosomes are driven by exosomal THBS1; MMP-9 and FAK are engaged by exosomal THBS1 in order to promote cancer invasiveness [80]. Therefore, stiffness-mediated secreted exosomes and their components can be used as potential therapeutics. As the effects of matrix stiffness on cancer processes is well known, comprehensive investigation of the effect of matrix stiffness on CSCs, one of the most crucial and challenging therapeutic targets, can lead to the development of novel cancer therapeutic strategies.
The significance of cancer stem cells (CSCs)
Tumor cells are heterogeneous populations with significant differences in cell surface markers, gene expression, proliferation, invasiveness, and therapeutic response [81]. Two theories explain the heterogeneity of tumor cells: in the stochastic model, a unique population of tumor cells acquires mutations and develops the ability to metastasize. In the hierarchical model, a small subpopulation of cancer cells in a tumor adheres to a functional hierarchy, allowing for self-renewal and differentiation. In this model, CSCs are responsible for developing initial tumors and metastasis [82, 83]. Bonnet et al. reported in 1997 that acute myeloid leukemia (AML) contains a stem cell hierarchy that mimics the normal hematopoietic stem cell hierarchy. They revealed that serial transplantation of a rare population of CD34+ CD38− leukemia cells can repopulate the tumor in its entirety; indicating that this population of cells possesses stem cell-like properties, such as the potential to proliferate. This research formed the basis for subsequent CSC studies [84]. CSCs are identified by stem cell markers such as CD44 + and CD133 + , and their characteristics are maintained via the expression of pluripotency factors such as Nanog and Oct-4 [11, 85, 86].
CSCs show a variety of characteristics, including self-renewal, proliferation, and differentiation into several cancer cell lineages through symmetric and asymmetric cell division, migration capacity, and specific surface markers. These cells are widely believed to play a critical role in tumor initiation, progression, development, metastasis, drug resistance, and recurrence [87]. Due to their increased chemoresistance and quiescence, CSCs are one of the most important cancer recurrence drivers [88]. The frequency of CSCs varies significantly between tumor types, ranging from < 1% in liver cancer to 82% in acute lymphoblastic leukemia (ALL) [89]. The proportion of CSCs in tumors is influenced by various parameters, including the host environmental conditions. The TME, especially the tumor stroma, is one of the parameters involved in the maintenance of CSC populations [90]. Moreover, changes in TME components can result in the dedifferentiation of mesenchymal or epithelial cells into CSCs [15]. Stem cell lineage commitment and differentiation can be affected by the stiffness of the matrix. In a soft matrix, mesenchymal stem cells (MSCs) differentiate into the neurogenic lineage, whereas in a stiff matrix, MSCs differentiate into the osteogenic lineage [91, 92]. Deregulation of ECM dynamics is essential for the formation of the niche for tumor stem cells and the generation of CSCs [93], and matrix stiffness as an important ECM characteristic can induce stemness [20]. Based on the CSC hierarchy/heterogeneity model and the importance of CSC, targeting all tumor cells with a similar approach would be ineffective, while eliminating the CSCs will eradicate the tumor and prevent recurrence. It is important to understand the matrix stiffness-mediated effects on CSCs in order to find targetable pathways that may be clinically advantageous. The next section discusses the effect of matrix stiffness on CSC in various cancers as one of the characteristics of the tumor stroma.
Effect of matrix stiffness on the CSC population, characteristics, and functions various malignancies
Effect of matrix stiffness on breast CSCs (BCSCs)
Breast cancer (BC) is the most common cancer in women and the fifth leading cause of cancer-related deaths worldwide [94]. Twelve percent of all women in the United States will be diagnosed with BC in their lifetime, according to the American Cancer Society [95]. Abnormal modifications in the quantity and organization of ECM components, such as collagen, occur during the progression of BC, leading to an increase in matrix stiffness that promotes tumor development and metastasis [32, 96]. Matrix stiffness is one of the most well-known risk factors for BC development [97]. Collagen fibers in grade 3 mammary carcinoma are thicker, longer, and straighter than those in grades I and II [98]. In BC patients, tumor stiffness can predict prognosis and classify treatment response [99]. Specific tumor-associated collagen signatures (TACS), which represent the density and organization of collagen fibers, can predict recurrence, therapeutic response, and clinical outcomes in BC [63, 97, 100]. These data highlight the importance of collagen organization and stiffness in breast cancer.
In the study by Samani et al. that evaluate the stiffness of normal and pathological human breast tissues, the elastic moduli of normal, low-grade invasive ductal carcinoma (IDC), ductal carcinoma in Situ (DCIS), and high-grade IDC were 3.25, 10.40, 16.38, and 42.52 kilopascal (kPa), respectively [6] (Table 1). The mean values of stiffness measured by SWE correlate with subtypes and histological characteristics. The ECM stiffness of ER-positive tumor cells was 136 kPa, HER2-positive 160 kPa, triple-negative BC (TNBC) 169 kPa, and the stiffnesses of grades I, II, and III were 117, 132, and 165 kPa, respectively [101].
Using the cell surface markers Epithelial Specific Antigen (ESA+), CD44+, and CD24−, populations of stem-cell-like cells in breast cancer can be identified [11, 102]. It is shown that in the xenograft mouse model, as few as 200 cells of these cells can form tumors [11]. In addition, the expression and activity of aldehyde dehydrogenase (ALDH) has been applied to isolate and detect human breast CSC populations. ALDH1+ CSCs are significantly aggressive; patients with ALDH1 positive tumor cells are more resistant to treatment, and prognosis is poor. ALDH1high cancer cells generate more colonies and mammospheres than ALDH1low cancer cells [103]. The stiffness of the matrix has a significant effect on the maintenance of BCSC phenotypes [19]. The translocation of YAP and TAZ to nuclear promotes the BCSC phenotype [52, 104], and TNBC has the highest proportion of cells expressing BCSC markers compared to other BC subtypes [105].
Increasing stiffness can lead to an increase in BCSC as shown in both in-vitro and in-vivo studies. Application of mechanical forces can increase the CSC populations in MCF-7 breast cancer cells [106]. In 3D scaffolds with the same stiffness as breast tumor tissue, stemness markers (Nanog, Sox2, and Oct4) and CD44 were found to be expressed at a higher level than in 2D cultures. Also, sphere formation was higher in the scaffold than in 2D culture, indicating an increase in stemness and metastatic potential [107]. In aligned collagen matrices, the motility and contact-guided migration of BCSCs were significantly enhanced [108]. Using polyacrylamide (PA) substrates to mimic the stiff (4020 pa) and soft (120 pa) microenvironments of breast tumors and normal tissues, respectively; in the stiffer matrix, 4T1 and MDA-MB-231 cells expressed higher CSC markers, including CD44, Nanog, CD49, and ALDH and in regions of human breast cancer with low collagen levels (soft region), only 4% of tumor cells expressed CD44 and Integrin-linked kinase (ILK), a crucial mediator used by cells to sense their surroundings as opposed to the regions of BC with high collagen levels, where more than 25% of cells expressed CD44 and ILK. These results indicate that breast CSCs are frequently found in the dense regions. In 4T1 and MDA-MB-231 cells cultured on stiff substrata, ILK knockdown reduced CSC markers and decreased the tumorigenic and metastatic potential of tumors [19]. Following culturing human MCF7 and MDA-MB-231 breast cancer cells on Polyethylene Glycol Diacrylate (PEGDA) gels with moduli ranging from 2 to 70 kPa, it was found that 5 kPa was the optimal stiffness for maintaining the population of BCSCs [109]. When 4T1 breast cancer cells were grown on PEGDA hydrogels, increasing matrix stiffness from 2.5 kPa to 5.3, 26.1, and 47.1 kPa, resulted in tumor sphere size increasing from 37 to 57 µm, 20 µm, and 12 µm, respectively, and CD44 expression increased from 17-fold to 38-fold, threefold, and twofold, respectively, compared to the baseline levels. Additionally, MCF7 human breast cancer cells had similar results, and cells cultured in gel with modulus of 5.3 kPa showed the highest CD44 expression and the largest tumor spheres [110] (Table 2).
Following culturing the MDA-MB 231 breast tumor spheroids on 3D hydrogels with 2 kPa and 12 kPa stiffness, in cells cultured in a stiff environment, the expression of Mammalian-enabled (MENA), an invadopodia protein associated with breast cancer metastasis, was observed [111] (Fig. 4). Overexpression of MENA in cancer cells could increase CSC production and EMT markers expression [112]. Accumulation of Collagen I in tumor of Col1a1tmJae/+ mice and higher CSC activity due to AKT-mTOR and YAP activation is shown, and that these mice have more and larger lung metastases. Rapamycin, an inhibitor of mTOR, decreased mammary tumor size and CSC activity. However, in contrast to primary tumor, inhibition of mTOR signaling did not inhibit lung metastases due to the lower activity of mTOR and proliferation activity in lung cells compared to mammary tumors; as a result, Col1a1tmJae/+ mice continued to sustain higher metastatic burdens. These findings shed light on the association between stiffness and CSC activity and metastatic behavior; it also emphasizes the different therapeutic responses of local versus distant breast cancer lesions [113]. As such, increasing stiffness in breast cancer leads to higher BCSC rates and modified CSC features, which promote aggressive behaviors and metastasis in BC.

The effect of matrix stiffness on the CSC population in several malignancies. Molecular expression and activation were altered when breast, lung, liver, pancreatic, and colon cancer cell lines were cultured on a scaffold with gradient stiffness. These molecules enhance CSC population and stemness characteristics
Effect of matrix stiffness on lung CSCs (LuCSCs)
Lung cancer is the leading cause of cancer-related death (18% of mortality) and the second most commonly diagnosed cancer [94]. According to one study,lung tumors are more stiff (20–30 kPa) than normal lung parenchyma (0.5–5 kPa) [74] (Table 1). In another study, the matrix stiffness of healthy lung parenchyma was reported to be 0.15 to 0.2 kPa, whereas fibrotic lung parenchyma had a matrix stiffness of 15 kPa [114]. Injury-repair and tumorigenesis are associated, and injury-induced inflammation can result in lung fibrosis, and a stiff tissue matrix increases the risk of carcinogenesis [114]. Non-small-cell lung cancer (NSCLC) patients with lung fibrosis had a worse prognosis and treatment response rate than NSCLC patients without lung fibrosis [114]. Increased collagen expression in NSCLCs activates FAK and ERK signaling pathways and promotes cytokine production such a IL-23, hence promoting lung cancer progression [115]. Moreover, primary tumor resection develops hypoxic areas that are a source of LOX, which enters the circulation and ultimately reaches to the lungs, resulting in a stiffer environment that promotes lung metastasis through FAK activation [116]. Due to FAK activation and collagen-dependent metastasis, FAK inhibitors may improve survival [116, 117]. Downregulation of miR-29a is associated with posttranslational overexpression of LOXL2 in lung cancer, promoting tumor progression through modulating ECM stiffness [118].
In addition, the increased stiffness of lung can affect immunomodulation. Expression of programmed cell death-ligand1 (PD-L1) on cancer cells is important for immune evasion. and it is also positively correlated with EMT, cell migration, and invasion [119]. Multiple mechanisms, including matrix stiffness, regulate PD-L1 expression. When HCC827 lung cancer cells were grown on 2 and 25 kPa PA hydrogels; on substrates with higher stiffness, PD-L1 protein expression was higher than in the 2 kPa gel [74]. The YAP/TEAD complex regulates PD-L1 transcription by binding to the PD-L1 promoter. Due to YAP overexpression, PC9 adenocarcinoma cells express PD-L1 at a higher level [120]. Interestingly, CSCs and PD-L1 are correlated. A positive association between CD44 and PD-L1 expression in lung adenocarcinoma patients is shown [121]. In the context of lung tumors, stiffness can regulate the expression of PD-L1 in CSCs, hence facilitating immune evasion and tumor growth.
Several markers, including ALDH, ATP binding cassette subfamily G member 2 (ABCG2), CD44, CD117/KIT, CD133, and stem markers Nanog and OCT3/4, are overexpressed in lung cancer and have been used to identify CSC populations [122]. Malignant lung tissues with stiffer matrix are more favorable for CSC formation and maintenance via YAP/TAZ signaling pathways [20, 123].
Following is a series of studies that evaluate the effect of matrix stiffness on LuCSCs. A549 cells were seeded on matrices with stiffnesses of 0.2, 2, and 25 kPa to mimic physiological, fibrotic, and severe fibrosis tissues, respectively. Increasing stiffness elevated the expression of epidermal growth factor receptor (EGFR) and hepatic growth factor receptor (c-Met), leading to an increase in tumor cell proliferation and EMT [124].Khater et al. found that c-Met signal transduction increases bulk tumor CSC enrichment and self-renewal potential [125]. In another study, several lung cancer cell lines were cultured on 2D, very stiff scaffolds (ABS, HIPS, and PLA) and stiffness comparable to lung cancer (GelMA-PEGDA). The levels of CSC markers expression in cells grown on scaffolds were significantly higher than in 2D culture. However, the expression levels of cells seeded on very stiff scaffolds (ABS, HIPS, and PLA) were lower than those on GelMA-PEG-based scaffolds [126].
In an elegant study, Pankova et al. used two cells: 1- H1299 cell line that is highly methylated and lacks the expression of RASSF1A as control and 2- H1299 that continuously express RASSF1A. Both cells are seeded on collagen-coated matrigel with defined stiffness. Soft ECM (0.5 kPa) induces the reprogramming of H1299control cells to a cancer stem cell-like state and NANOG expression, but not in H1299RASSF1A cells. Increasing the ECM stiffness (4 kPa) enhanced NANOG expression in H1299RASSF1A cells. These data support the correlation between cancer stemness and ECM stiffness and suggest that RASSF1A suppresses stemness in soft ECM. Surprisingly, the expression of NANOG and CD133 was not increased in H1299control and H1299RASSF1A cells grown on a very stiff (25 kPa) scaffold [20]. They concluded that high matrix stiffness may lock the ECM conformation, preventing the exposure of binding sites such as integrins and so decreasing the capacity to respond to ECM [127] (Table 2). Moreover, IHC staining of H1299control primary tumors showed NANOG expression and significant levels of nuclear YAP1, whereas in H1299RASSF1A tumors, the majority of YAP1 was localized in the cytoplasm and with is no detectable NANOG staining [20]. These results support the stiffness-mediated activation of NANOG and cancer stemness. Accordingly, increased lung cancer stiffness correlates with higher lung CSC fraction and modified CSC characteristics, which promotes aggressive behaviors and metastasis.
Effect of matrix stiffness on liver CSCs (LCSCs)
Liver cancer is the third leading cause of cancer-related deaths worldwide [94]. Approximately 90% of liver cancers are HCC and 10% cholangiocellular carcinoma (CCC) [128]. Liver stiffness increases in primary and metastatic cancers and promotes proliferation and cancer development [129]. Over 80% of patients with HCC have a background of cirrhosis or severe liver fibrosis [130, 131]. There is a two-to fivefold increase in total collagen content in a cirrhotic liver, and an increase in type I collagen is the primary distinguishing hallmark of liver fibrosis [132]. Importantly, fibrosis precedes the development of HCC, making it an important characteristic of the premalignant hepatic milieu. It is estimated that approximately one-third of cirrhotic patients will eventually develop HCC [24]. Patients with chronic hepatitis B with liver stiffness greater than 13 kPa had a 4-to 13-fold increased risk of HCC. In one study, it was shown that no patient with liver stiffness < 12 kPa had HCC within 21.8 months of follow-up, whereas 26% of those with liver stiffness > 12 kPa developed HCC. [133]. Thus, increased matrix stiffness in HCC promotes tumor progression and metastasis. Research on HCC indicates that matrix stiffness can modulate cell proliferation, angiogenesis, metastasis, and drug resistance [24].
Using a shear elasticity probe, the elastic modulus of a healthy liver ranged between 1.5 and 5 kPa, but it ranged between 5 and 69 kPa for fibrosis grades 1 to 4 [134]. According to one report, the tissue stiffness for HCC, CCC and metastatic tumors are 55, 75 and, 66.5 kPa respectively [129]. AFM analysis of HCC tissue stiffness classified patients into low degree (8–15 kPa) and high degree (14–18 kPa) malignant groups [135] (Table 1). The liver stiffness, as measured by Two-Dimensional Shear-Wave Elastography, could be an effective predictor of overall survival (OS) following radiofrequency ablation (RFA) for HCC. Patients with stiffness ≥ 13.3 kPa had a 3-year OS of 76.8%, whereas patients with stiffness < 13.3 kPa had a 3-year OS of 96.3% [136], and a later study confirmed these results [72]. In addition, liver cancer displays mechanical heterogeneity, with the invasive tumor front (ITF) becoming stiffer than the tumor's core. Intriguingly, the distribution of LCSCs correlates with the stiffness of the tumor, with the highest proportion of these cells be observed at the ITF [5].
LCSCs are identified by surface markers including CD133, epithelial cell adhesion molecule (EpCAM), CD90, CD44, CD24, CD13, OV6, and ALDH activity. LCSCs are associated with increased proliferation, tumorigenicity, metastasis, radiation or chemotherapy resistance, recurrence, and poor prognosis [137, 138]. EpCAM-positive HCC cells show highly tumorigenic capacity and CD90-positive HCC cells are highly metastatic [139]. It has been shown that chronic inflammation-induced stiffness increases the overall population of HCC stem cells [140]. Tumor-associated macrophages (TAM) are the cells involved in this phenomenon. Both M1 and M2 macrophages TAMs contribute to the synthesis of ECM molecules [141], and M2 can support stem cells and regulate their behavior as a part of the niche. Considering the increased stiffness of the matrix in HCC and the significance of CSCs, here we review a number of studies on the impact of matrix stiffness on LCSCs.
Culturing SMMC-7721 cells on stiff PA hydrogels increased the expression of stemness genes [135]. The addition of matrigel, collagen 1, or methyl cellulose to the sphere-forming culture medium significantly increased the initial oncosphere formation and the expression of pluripotent and stemness markers in LCSCs in response to increased stiffness [142]. Huh7 and Hep3B cells were cultured on COLI-coated PA gel substrates with tunable stiffness (6, 10, and 16 kPa). In the stiffer matrix, cells display increased stemness-related gene expression, including SOX2 and Nanog, as well as with high self-renewal capacity and a high proportion of CD133(+)/EpCAM cells. In addition, the phosphorylation levels of AKT and mTOR are increased in cells on the stiffer matrix. The knockdown of integrin beta 1 reduces the phosphorylation of AKT and mTOR molecules, hence decreasing the cellular response to stiffness. Moreover, mTOR inhibitors decrease SOX2 expression; thus, stiffness may exert its effect on cells through the integrin beta 1 molecule, and with the cascade of events such as phosphorylation and activation of the molecular pathways of AKT and mTOR, ultimately leads to increased expression of the stemness genes [143]. In a related study, several human HCC cell lines, including MHCC97H, Hep3B, HepG2, and Huh7, were cultured on gels of three distinct stiffnesses (from 1 to 40 kPa). In medium stiffness-cultured cells, EpCAM and cholangiocyte markers, including cytokeratin7 (CK7) and CK19, were considerably increased. The formation of tumors and the expression levels of EpCAM, CK7, and CK19 were also elevated in mice injected with cells derived from medium-stiffness gels [144]. The presence of CK, a stem cell marker, is associated with a poor prognosis [145, 146]. Also, miR-3682-3p was significantly up-regulated in stiffness-cultured MHCC-97H [147]. In HCC cells, the upregulation of miR-3682-3p improved the spheroid forming capacity, the side population cell fractions, the expression of CSC factors [148], and the poor prognosis in HCC patients [149]. Contrary to previous studies, the culture of Huh7 and HepG2 cells on a PA coated with collagen-I showed that soft substrate cells expressed higher levels of CSCs markers, including OCT4, Nanog, CD44, CD133, and CXCR4 as a chemokine receptor. Due to the use of a soft matrix with a stiffness of 1 kPa, which does not accurately represent hepatic stiffness in normal or pathological livers, the results of this study may be contradictory [150] (Table 2).
In a recent study, Yang et al. [151] found that as matrix stiffness increased, CXCR4 expression in HCC cells increased significantly, promoting EMT and stemness. According to the known role of CXCR4 on CSCs function, it was determined that increased expression of CXCR4 correlates with chemotaxis, invasion, and CSC characteristics in a variety of solid tumor malignancies, and that treatment with miR-139, which directly targets CXCR4, inhibited mesenchymal traits of CSCs [152, 153]. Matrix stiffness acts through CXCR4 to decrease the levels of ubiquitin domain-containing protein) UBTD1(, which is involved in the degradation of YAP, hence activating YAP-targeted genes and YAP downstream signaling [151]. In addition, activated metastasis-associated fibroblasts increase liver stiffness and promote angiogenesis, thereby providing sufficient nutrients for CSCs. Due to ECM stiffness, colorectal cancer patients with liver metastases are resistant to anti-angiogenic therapy. Inhibitors of fibroblast contraction reduce metastatic liver stiffness and increase bevacizumab's antiangiogenic effects [154]. In conclusion, recent research indicates that increasing stiffness can elevate the population of LCSCs and amplify their characteristics.
Effect of matrix stiffness on pancreatic CSCs (PaCSCs)
Pancreatic cancer is an aggressive malignancy that counts as the seventh leading cause of cancer-related death worldwide [155]. It has one of the worst prognoses among solid tumors, with a 5-year survival rate of less than 10% [155, 156]. Despite increased understanding of pancreatic cancer risk factors and the development of new diagnostic techniques, the incidence of pancreatic cancer is still increasing. Pancreatic ductal adenocarcinoma (PDAC) is estimated to become the second leading cause of death from cancer by 2030 [157]. TME is very prominent in PDAC, and approximately 90% of the tumor volume is composed of stromal cells and extensive ECM deposition [158]. The elastic modulus of the non-neoplastic adjacent pancreas was less than 15 kPa, whereas PDAC tumors was over 40 kPa, measured by Harmonic Motion Elastography (HME) [159]. Also, AFM analysis revealed that the stiffness of normal pancreatic tissue was 0.4 kPa and that of pancreatic cancer tissue was 1.2 kPa [41] (Table 1). Pancreatic cancer tissues had higher amounts of collagen, hyaluronan, and the CD44 receptor. In addition, the transition from a healthy pancreas to invasive pancreatic ductal adenocarcinoma is accompanied by an increase in the thickness of collagen fibers, which is associated with a poor prognosis [160]. A high strain ratio (SR), which reflects the stiffness of pancreatic tissue, has prognostic value, and as higher SR predicts poor overall survival [161].
Increased collagen I expression in PDAC is associated with higher invadopodia formation in invading cancer cells, increased metastasis, and poor prognosis [162]. Also, stiffness can affect the sensitivity of pancreatic cancer cells to chemotherapy [68]. More than 90% of PDAC patients have mutant oncogenic KRAS, which is activated and converges with downstream signaling pathways such as YAP/TAZ [163]. Zhang et al. found that YAP is essential for tumorigenesis and the development of invasive PDAC in mice inoculated with KRAS mutant neoplastic pancreatic ductal cells [163]. Also, activation of the YAP1/TEAD complex cooperatively acts to promote PDAC recurrence in the absence of oncogenic KRAS, implying a novel mechanism for PDAC recurrence independent of the KRAS mutation. This suggests that YAP1/TAZ-dependent signaling may be essential for the early development and recurrence of PDAC [164].
Less than 1% of pancreatic tumor cells are CSCs, and the elimination of PaCSCs is a necessity for any PDAC therapeutic treatment [165]. Several cell-surface markers are used to detecting pancreatic CSCs. CD133, CD24, CD44, EPCAM, ESA, c-Met, Aldh1, ABCG2, and more recently, DclK1 and Lgr5 have been identified as markers of PaCSCs [166]. In pancreatic cancer, stiffness-sensing receptors activate Ras, Rac, MAPK, and PI3K signaling pathways, resulting in increased cell proliferation and stem cell characteristics [167]
In a recent study, 1, 4, and 25 kPa of acrylamide/bisacrylamide were utilized to determine the impact of stiffness on pancreatic cancer cells. The results demonstrated that BxPC-3 cells are more resistant to chemotherapy when cultured on matrix with 4 and 25 kPa stiffness. In a mouse model, stiff pancreatic cancer tissues led to EMT, increased vimentin expression, decreased E-cadherin expression, treatment resistance, and increased β-catenin and YAP nucleus localization [41]. In response to the increased matrix stiffness, YAP increases CD133 expression, which leads to an increase in cell proliferation and metastasis [168, 169]. Moreover, it has been shown that pancreatic tumors with a high proportion of tumor-associated fibroblasts (TAFs) have more drug-resistant and stem-like cells due to the fact that TAFs increase ECM synthesis in response to inflammation [170].
PDAC organoids were expanded on the HELP (Hyaluronan and elastin-like protein) Low and HELP High matrices with stiffnesses of 279, 1253, and 3040 Pa, respectively, to determine the effect of matrix stiffness on CSCs. In comparison to HELP Low, CSC markers such as CD44, ABCG2, and CD24 increased in HELP High matrices. CSCs frequently exhibit chemo resistance through the altered expression of drug transporters. The drug efflux transporters (ABCG2, ABCC3/4/5) associated with PDAC chemoresistance are increased in organoids grown on stiff matrices [171]. Also, the side population (SP), which has become an important hallmark for defining the stem-cell population [172], was significantly larger in organoids grown in the high stiff matrix (3.74%) than in the low stiff matrix (0.79%). Intriguingly, PDAC organoids that expanded in the stiff matrix were not drug-sensitized, but when switched to a matrix with low stiffness, they became drug-sensitized. Also, following multiple passages, the expression of CD44 and ABCG2 decreased in the soft matrix [171] (Table 2). These findings indicate that stiffness can affect PaCSCs and suggest that treatment of PDAC tumors with drugs that target matrix stiffness in combination with anti-cancer agents may improve therapeutic sensitivity of tumors, reduce the aggressive behavior of CSCs, and improve patient outcomes.
Effect of matrix stiffness on colorectal CSCs (CCSCs)
Colorectal cancer (CRC) is the second leading cause of cancer-related deaths and ranks third in incidence worldwide [94]. In 2021, it is estimated that there were approximately 149,500 new cases and 52,980 deaths in the United States due to CRC [173]. CRC tissue is stiffer than normal tissue and promotes the proliferation, invasion, and metastasis of CRC cells [174, 175]. Recent research found that in the regions 10 to 20 cm away from the tumor, the ECM of uninvolved rectal mucosa is remodeled and stiffness is increased. Hence, the fact that increased matrix stiffness in CRC is not restricted to the primary lesion shows that the effect of increased matrix stiffness in CRC is very complicated [176]. Positive correlation between YAP/TAZ expression and poor prognosis in CRC patients emphasizes the carcinogenic properties of mechanoregulators in CRCs [177, 178]. YAP also promotes CRC chemotherapy resistance and cancer recurrence [179]. YAP inhibition reduced CRC cell lines proliferation and metastasis considerably, whereas YAP overexpression enhanced the rate of cell proliferation [180].
Correlation of CRC tissue stiffness with the clinicopathological characteristics of patients were evaluated by Kawano et al. The median elastic modulus (EM) of normal colorectal tissue was 0.90 kPa, which is considerably lower than the median EM of CRC tissue (7.5 kPa; min = 1.1 kPa, max = 68 kPa). Increasing stiffness correlates with the pathological T, N, and M stages of cancer as well as with survival. T1, T2, T3, and T4 had respective median EM values of 2.8, 3.5, 8.8, and 13.8 kPa. In addition, the median EM of patients without distant metastases was 7 kPa, whereas it was 13.6 kPa in patients with metastasis (Table 1). Patients with stiffer tumors also had a shorter disease-free survival than those with less stiff tumors [181].
CD44, ALDH1, ALCAM, and CD133 have been identified as CRC stem cell markers. CD133+ colon cancer cells are highly tumorigenic, self-renewing, and capable of tumor formation, whereas CD133− cells are unable to do so [182,183,184]. Overexpression of Collagen 1 promotes expression of CD133 and BMI1 stem cell markers in CRC [185], and high stiffness enhances the expression of CCSCs markers and is critical for the maintenance of the CSC phenotype [186].
HCT-8 colon cancer cells were cultured on PA gels with different stiffnesses (1, 21, 47 kPa) and on a polystyrene surface with a stiffness of 3.6 GPa. At 21 kPa, a higher proportion of metastatic-like R-cells (rounded, separated, metastatic -like phenotype; more aggressive) was observed in comparison to E cells (cells with an epithelial-like phenotype) [187]. In addition to confirming the E-R transition of HCT-8 cells on a PA scaffold with 20 kPa [174], R cells exhibited higher ALDH3A1 activity as a CSC marker for colon carcinoma and other cancer tissues [188, 189]. Upregulation of TNS4, CLDN2, and AKR1B10 in cells cultured on scaffold with 20 kPa; all of these molecules play key roles in cancer cell migration, invasion, proliferation, and apoptosis [174]. The optimum stiffness for HCT-116 tumor stem cell proliferation and marker expression in PEGDA gels was determined to be 25 kPa (2–70 kPa). HCT-116 cells grown on PEGDA gels (2–70 kPa) showed the highest level of tumor stem cell proliferation and marker expression at 25 kPa [109]. HCT-116 cells were grown on matrices with different stiffness (2–20 kPa); at high matrix stiffness, YAP activation in CRC stem cells was considerably elevated, leading to an increase in stemness marker expression (CD133, ALDH1, and Lgr5). Also, due to collagen deposition, CD133 expression was higher in the ITF of CRC tissue samples. Matrix stiffness regulates and maintains CCSC characteristics via the integrin 1/FAK/YAP pathway [186]. CCD18 cells (colon fibroblast cells) were grown on fibronectin-functionalized PA substrates of 2, 10, 40, 95, and 120 kPa. Concentration of Activin A [190], the molecule that regulate self-renewal, plasticity, differentiation and metastatic potential of CSCs [191], was elevated and reached a plateau at 40 kPa in the conditioned medium of CCD18 cells. CRC epithelial FET cells were treated with conditioned medium from CCD18 cultured on increasing stiffness substrates in order to investigate the functional effects of activin A. The highest migration in cancer cells was detected using conditioned medium of the 40 kPa substrate. CRC cells migrated less after the addition of follistatin, a ligand trap for activin A [190] (Table 2). The elevated Activin A level enhanced invasive ALDHhi CSC-like phenotypes and cancer cell plasticity and metastatic potential [192, 193]. As such, studies have shown that the stiffness of the ECM modulates the frequency and characteristics of CCSCs.
Clinical implications
Considering the significant role of CSCs in the progression of cancer, therapeutic approaches that fail to eliminate CSCs are likely ineffective [194]. Therefore, it is important to develop novel anticancer strategies that directly target CSC populations [195, 196] or components of the TME that cause CSCs to proliferate, make them more aggressive, and sustain their population. Over the course of previous years, there is a significant body of knowledge addressing the mechanisms and players of TME involved in the maintenance of CSCs. Based on the findings reported in the preceding sections, we conclude that matrix stiffness, a mechanical characteristic of TME, plays a key role in CSCs function. Inhibition of stiffness as a supportive niche for CSCs appears to be one of the most effective cancer treatments and provides a novel therapeutic approach to enhance patient outcomes.
Also, cancer stem cell numbers and functional populations are distinct concepts; in fact, stem cell identity and functionality are different [197, 198]. For instance, each homeostatic mouse colonic crypt includes 5 to 7 functional stem cells [198]. However, the number of cells that express stem cell markers such as Lgr5 is ~ 16 per crypt [199]. The position of CSCs in the TME affects its functionality [200]. Functional CSCs that drive tumor progression mainly reside at the tumor's edge, close to CAFs [201]. Also, functional CSC are not necessarily the same cells that express known-CSC markers. In addition, the TME defines the enrichment of functional CSC cells in response to chemotherapy [201]. Therefore, the TME is dominant over cell-autonomous features in defining stem cell functionality. Hence, cancer therapies could be improved by strategies that particularly target the TME compartment, including its stiffness. This approach can block access to the activating signals that provide the soil for differentiated cells to become clonogens and restrict CSCs from entering their favorable niche.
Several approaches for targeting ECM stiffness in cancer have been explored. Targeted therapies against the factors that contribute to establishment of stiffness or against the stiffness-induced activated signaling pathways could potentially modulate and control the effects of stiffness on CSCs. In the mice treated with collagozome, a liposome encapsulating collagenase, malignant tumors were reduced in size by 87% [202]. Collagenases degrade collagen, allowing for improved drug delivery to tumor sites [203]. Inhibition of LOX activity, one of the well-known molecules for matrix stiffness, reduced tumor progression and metastasis in mice [204]. The drug losartan efficiently suppresses lung tumor metastasis by decreasing the level of LOX [205], and inhibiting collagen I synthesis and deposition (NCT01821729 and NCT04106856). In a model of collagen-dependent lung cancer metastasis, trihydroxyphenolics blocked collagen deposition by inhibiting LOXL2 [206].
After resection of the primary tumor, stiffness-affecting components may cause recurrence and distant metastases [207, 208]. Peritoneal surgery in mice creates hypoxic areas at the surgical site and increases LOX expression, which enters the circulation and lungs. The presence and activity of LOX as well as the expression of fibrillar collagen were considerably elevated in the lungs of surgically treated mice, which led to tumor cell seeding and lung metastasis. Notably, LOX inhibition following surgery reduces metastasis and improves survival [116].
Patients with metastatic CRC are widely treated with anti-VEGF in combination with chemotherapy, but the survival benefit is modest due to acquired resistance [209]. Anti-VEGF therapy, such as bevacizumab, enhanced hyaluronic acid (HA) deposition and stiffness of metastatic liver. The remodeling of the ECM and increased stiffness appears to be driven by treatment-induced hypoxia in the tumor. Stiff ECM decreases blood perfusion, which is a key factor in determining the treatment outcome. In preclinical models, they showed that enzymatic depletion of HA, partially restored perfusion in the liver in metastatic colorectal cancer following chemotherapy and anti-VEGF therapy leading to prolonged survival [210]. These results suggest that factors causing stiffness such as HA could be a potential therapeutic target for reducing physical barriers to systemic treatments in cancer patients receiving anti-VEGF therapy.
Volociximab is a monoclonal antibody used to target integrin α5β1 and reduce ECM stiffness in several tumors [211]. Cilengitide, as an α6β5 integrin inhibitor, reduced the progression of malignancies in a variety of preclinical studies, leading to its investigation in clinical trials [212]. Focal adhesion kinase (FAK) inhibitor defactinib suppressed tumor growth and metastatic ability and increased the overall survival of xenografted animals [213]. YAP plays a significant role in the formation and maintenance of CSCs characteristics as well as the promotion of tumorigenesis, metastasis, and recurrence, several attempts have been conducted to investigate its therapeutic potential [179, 214, 215]. The pro-oncogenic property of the YAP pathway, requires YAP/TEAD binding to activate YAP-dependent downstream signaling [38]. The molecules that impair this binding could be potential therapeutic agents. Verteporfin is an inhibitor of YAP/TEAD interaction that suppressed the CSC-associated characteristics of gastric cancer cell line and inhibited tumor growth in a xenograft model [216]. In addition, VGLL4 competes with YAP for binding with TEAD, which suppresses cancer [217]. These results showed the therapeutic potential of YAP for modulating CSC characteristics. Also, the activity of TAZ, another key mechanotransducer, can affect CSCs [218, 219]. Mechanical cues can lead to the formation of a transcriptional complex of TAZ and TEAD4 and the expression of SOX2, which modulates the maintenance and self-renewal of CSCs [218]. These findings suggest that targeting the TAZ-SOX2 axis could be a potential treatment for cancer. The activation of TAZ is required for breast CSCs to maintain their self-renewal and tumor-initiation capacities [52]. In addition, TAZ activation could drive non-CSCs into cells with tumor initiating and self-renewal potential [52], and loss of TAZ impairs the invasiveness, self-renewal, and tumorigenic capacity [220]. YAP/TAZ promotes autophagy through modulating TBC1D2 gene transcription. Autophagy is crucial for the maintenance of oncogenic characteristics and the acquisition of CSC properties, as well as the promotion of cell plasticity and self-renewal of somatic stem cells via YAP/TAZ [214]. Thus, direct or indirect targeting of YAP/TAZ mechanotransduction may block autophagy and, as a result, reducing CSC populations and rendering these cells less aggressive (Table 3). Furthermore, as a result of enhanced exosome secretion in stiff ECM, stiffness-mediated secreted exosomes and their contents could be potential therapeutic agents [80]. Collectively, in order to increase cancer patients’ survival, it is suggested that novel therapeutic agents, such as those that target stiffness, be used in combination with standard cancer treatments, such as chemotherapy and immunotherapy. Future clinical trials should focus on novel therapeutic agents that target the stiffness of the CSC niche or critical molecules that activate signaling pathways mediated by changes in stiffness.
Future directions
We have progressed in our understanding of the complex molecular mechanisms whereby matrix stiffness influences CSCs, but important questions remain.
How long does the tumor cell's mechanical memory persist?
Over time, cells store information on past mechanical cues, and this mechanical memory can influence the initiation of metastasis. The transfer of mesenchymal stem cells from a stiff matrix to a soft matrix blocked the re-localization of YAP from the nucleus to the cytoplasm for up to 10 days [221]. This mechanical memory effect suggests that the optimum time to initiate treatment with stiffness-reducing medications must be considered and determined.
Treatment based on heterogeneity of stiffness
As mentioned above section, some tumors display mechanical heterogeneity, and ITF is stiffer than the tumor's core. This heterogeneity can affect tumor cell activity in various ways. Cell in core have higher proliferation, glycolytic metabolism, whereas cell in peripheral zones have increased MMP9, and OXPHOS and FA metabolism [4]. Also, the distribution of CSCs correlates with this mechanical heterogeneity, such that ITF has the highest proportion of CSCs [5]. As such, the therapeutic strategy could be based on the local maps of ECM stiffness and its function.
And from a different perspective, may the stiffness of the cancer tissues serve as a platform for CSC-targeted therapy?
The stiffness index can serve as a platform to convert the pro-drug into a drug molecule for the targeted therapy of cancer cells and probably CSCs. Liu et al. designed a mechanoresponsive cell system (MRCS) that uses the specific stiffness index in the TME to target and treat cancer metastases selectively [222]. It is known that infused mesenchymal stem cells (MSC), selectively home to tumors and metastatic sites, in response to increased matrix stiffness [223]. Hence, MSCs can be utilized to designed a MRCS that have a mechanosensitive promoter–driven -based vectors. In stiff matrix, YAP of MRCS localizes to the nucleus, and cytosine deaminase (CD) is expressed, CD converts the prodrug 5-fluorocytosine (5-FC) to the active drug 5-fluorouracil (5-FU) at the metastatic site, which leads to the death of cancer cells [222]. In soft matrix, MSC YAP localizes to the cytoplasm and inhibits CD transcription. This shows that the stiffness of the matrix can serve as a platform for targeted therapies and enables the efficient delivery of drugs to the target site (Fig. 5).

The matrix’s stiffness as a platform for targeted therapy. Mesenchymal stem cells that have a mechanosensitive promoter- driven -based vectors (MSCs) are used to develop a mechanoresponsive cell system (MRCS). In response to increased matrix stiffness, engineered MSCs, selectively home to and target cancer metastases. In stiff ECM, YAP of MRCS localizes to the nucleus, and cytosine deaminase (CD) is expressed. In the tumor microenvironment (TME), CD converts the prodrug 5-fluorocytosine (5-FC) to the active drug 5-fluorouracil (5-FU), which leads to the death of cancer cells. In soft ECM, MSC YAP localizes to the cytoplasm and inhibits CD transcription
The surgical removal of primary tumors is associated with the formation of scars, which are stiffer than healthy tissue [116]. Is there a possibility that surgery-induced stiffness contributes to the tumor recurrence through inducing stem cell features? Is the stiffness of the matrix formed during tumorigenesis and the stiffness caused by surgery a more suitable niche for CSCs to enter a dormant state? If so, does stiffness affect the period of dormancy for these CSCs?
Conclusions
A plenty of evidence suggest that the stiffness of tumor matrix is significantly higher than that of normal tissues and strongly correlates with disease progression, metastasis and clinical outcomes in a range of cancers, including BC, CRC, HCC, and PDAC [9, 224]. Matrix stiffness not only plays a role in the transformation of tumor cells into CSCs but can serve as a means of sustainment of the CSC niches, hence promoting and maintaining particular CSC characteristics. Recent progress in understanding the molecular biology of tumor stiffness, particularly its effect on CSC biology, has provided an alternative explanation for tumor development, metastasis, and prospective therapies. Herein, we propose that the importance of tumor matrix stiffness in CSCs can provide insight into novel cancer therapy strategies. All factors that create matrix stiffness, such as LOX, and signaling pathways that are modulated by matrix stiffness, such as YAP, seem to be viable candidates for therapeutic approaches.
Availability of data and materials
Not applicable.
Abbreviations
- AFM:
-
Atomic force microscopy
- ALDH:
-
Aldehyde dehydrogenase
- ABCG2:
-
ATP binding cassette subfamily G member 2
- ALL:
-
Acute lymphoblastic leukemia
- α-SMA:
-
Alpha-smooth muscle actin
- AKT:
-
Serine/threonine-protein kinase
- BC:
-
Breast cancer
- BCSC:
-
Breast CSC
- CSC:
-
Cancer stem cell
- CAF:
-
Cancer-associated fibroblast
- CCC:
-
Cholangiocellular carcinoma
- CCSC:
-
Colorectal CSC
- CRC:
-
Colorectal cancer
- CTC:
-
Circulating tumor cell
- DCIS:
-
Ductal carcinoma in situ
- ECM:
-
Extracellular matrix
- EMT:
-
Epithelial-mesenchymal transition
- EpCAM:
-
Epithelial cell adhesion molecule
- EM:
-
Elastic modulus
- EGFR:
-
Epidermal growth factor receptor
- FAK:
-
Focal kinase adhesion
- FN1:
-
Fibronectin 1
- FA:
-
Fatty acid
- GSK3α/β:
-
Glycogen synthase kinase 3α/β
- HCC:
-
Hepatocellular carcinoma
- HA:
-
Hyaluronic acid
- HME:
-
Harmonic motion elastography
- ITF:
-
Invasive tumor front
- IBC:
-
Invasive breast cancer
- IDC:
-
Invasive ductal breast carcinoma
- LOX:
-
Lysyl oxidase
- LCSC:
-
Liver CSC
- MMP:
-
Matrix metalloproteinase
- MSC:
-
Mesenchymal stem cell
- NSCLC:
-
Non-small-cell lung cancer
- OPN:
-
Osteopontin
- OXPHOS:
-
Oxidative phosphorylation
- PDAC:
-
Pancreatic ductal adenocarcinoma
- PI3K:
-
Phosphatidylinositol-3-kinase
- PTEN:
-
Phosphatase and tensin homolog
- PA:
-
Polyacrylamide
- PaCSC:
-
Pancreatic CSC
- ROCK:
-
RhoA/Rho-associated protein kinase
- TAM:
-
Tumor associated macrophages
- TACS:
-
Tumor-associated collagen signatures
- TNBC:
-
Triple-negative BC
- 3D:
-
Three-dimensional
- THBS1:
-
Thrombospondin-1
- TME:
-
Tumor microenvironment
- TAZ:
-
WW domain-containing transcription regulator 1 (WWDR1)
- YAP1:
-
Yes-associated protein 1
References
Egeblad M, Nakasone ES, Werb Z. Tumors as organs: complex tissues that interface with the entire organism. Dev Cell. 2010;18(6):884–901.
Bissell MJ, Radisky D. Putting tumours in context. Nat Rev Cancer. 2001;1(1):46.
Mouw JK, Ou G, Weaver VM. Extracellular matrix assembly: a multiscale deconstruction. Nat Rev Mol Cell Biol. 2014;15(12):771–85.
Liu C, Li M, Dong Z-X, Jiang D, Li X, Lin S, Chen D, Zou X, Zhang X-D, Luker GD. Heterogeneous microenvironmental stiffness regulates pro-metastatic functions of breast cancer cells. Acta Biomater. 2021;131:326–40.
Sun Y, Li H, Chen Q, Luo Q, Song G. The distribution of liver cancer stem cells correlates with the mechanical heterogeneity of liver cancer tissue. Histochem Cell Biol. 2021;156(1):47–58.
Samani A, Zubovits J, Plewes D. Elastic moduli of normal and pathological human breast tissues: an inversion-technique-based investigation of 169 samples. Phys Med Biol. 2007;52(6):1565.
Dupont S, Morsut L, Aragona M, Enzo E, Giulitti S, Cordenonsi M, Zanconato F, Le Digabel J, Forcato M, Bicciato S. Role of YAP/TAZ in mechanotransduction. Nature. 2011;474(7350):179–83.
Acerbi I, Cassereau L, Dean I, Shi Q, Au A, Park C, Chen Y, Liphardt J, Hwang E, Weaver V. Human breast cancer invasion and aggression correlates with ECM stiffening and immune cell infiltration. Integr Biol. 2015;7(10):1120–34.
Reid SE, Kay EJ, Neilson LJ, Henze AT, Serneels J, McGhee EJ, Dhayade S, Nixon C, Mackey JB, Santi A. Tumor matrix stiffness promotes metastatic cancer cell interaction with the endothelium. EMBO J. 2017;36(16):2373–89.
Marusyk A, Polyak K. Tumor heterogeneity: causes and consequences. Biochim Biophys Acta (BBA) Rev Cancer. 2010;1805(1):105–17.
Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci. 2003;100(7):3983–8.
Chaffer CL, Marjanovic ND, Lee T, Bell G, Kleer CG, Reinhardt F, D’Alessio AC, Young RA, Weinberg RA. Poised chromatin at the ZEB1 promoter enables breast cancer cell plasticity and enhances tumorigenicity. Cell. 2013;154(1):61–74.
Christin JR, Wang C, Chung C-Y, Liu Y, Dravis C, Tang W, Oktay MH, Wahl GM, Guo W. Stem cell determinant SOX9 promotes lineage plasticity and progression in basal-like breast cancer. Cell Rep. 2020;31(10):107742.
Creighton CJ, Li X, Landis M, Dixon JM, Neumeister VM, Sjolund A, Rimm DL, Wong H, Rodriguez A, Herschkowitz JI. Residual breast cancers after conventional therapy display mesenchymal as well as tumor-initiating features. Proc Natl Acad Sci. 2009;106(33):13820–5.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–74.
Gupta PB, Fillmore CM, Jiang G, Shapira SD, Tao K, Kuperwasser C, Lander ES. Stochastic state transitions give rise to phenotypic equilibrium in populations of cancer cells. Cell. 2011;146(4):633–44.
Chaffer CL, Brueckmann I, Scheel C, Kaestli AJ, Wiggins PA, Rodrigues LO, Brooks M, Reinhardt F, Su Y, Polyak K. Normal and neoplastic nonstem cells can spontaneously convert to a stem-like state. Proc Natl Acad Sci. 2011;108(19):7950–5.
Wei J, Yao J, Yang C, Mao Y, Zhu D, Xie Y, Liu P, Yan M, Ren L, Lin Y. Heterogeneous matrix stiffness regulates the cancer stem-like cell phenotype in hepatocellular carcinoma. J Transl Med. 2022;20(1):555.
Pang M-F, Siedlik MJ, Han S, Stallings-Mann M, Radisky DC, Nelson CM. Tissue stiffness and hypoxia modulate the integrin-linked kinase ILK to control breast cancer stem-like cells. Can Res. 2016;76(18):5277–87.
Pankova D, Jiang Y, Chatzifrangkeskou M, Vendrell I, Buzzelli J, Ryan A, Brown C, O’Neill E. RASSF 1A controls tissue stiffness and cancer stem-like cells in lung adenocarcinoma. EMBO J. 2019;38(13):e100532.
Hui L, Zhang J, Ding X, Guo X, Jiang X. Matrix stiffness regulates the proliferation, stemness and chemoresistance of laryngeal squamous cancer cells. Int J Oncol. 2017;50(4):1439–47.
Pickup MW, Mouw JK, Weaver VM. The extracellular matrix modulates the hallmarks of cancer. EMBO Rep. 2014;15(12):1243–53.
Wright K, Ly T, Kriet M, Czirok A, Thomas SM. Cancer-associated fibroblasts: master tumor microenvironment modifiers. Cancers. 2023;15(6):1899.
Affo S, Yu L-X, Schwabe RF. The role of cancer-associated fibroblasts and fibrosis in liver cancer. Annu Rev Pathol. 2017;12:153–86.
Wei B, Zhou X, Liang C, Zheng X, Lei P, Fang J, Han X, Wang L, Qi C, Wei H. Human colorectal cancer progression correlates with LOX-induced ECM stiffening. Int J Biol Sci. 2017;13(11):1450.
Koorman T, Jansen KA, Khalil A, Haughton PD, Visser D, Rätze MA, Haakma WE, Sakalauskaitè G, van Diest PJ, de Rooij J. Spatial collagen stiffening promotes collective breast cancer cell invasion by reinforcing extracellular matrix alignment. Oncogene. 2022;41(17):2458–69.
Baumgart E. Stiffness—an unknown world of mechanical science? Injury. 2000;31(Suppl 2):s-b14-23.
Stylianou A, Lekka M, Stylianopoulos T. AFM assessing of nanomechanical fingerprints for cancer early diagnosis and classification: from single cell to tissue level. Nanoscale. 2018;10(45):20930–45.
Chokhandre S, Erdemir A. A comprehensive testing protocol for macro-scale mechanical characterization of knee articular cartilage with documented experimental repeatability. J Mech Behav Biomed Mater. 2020;112: 104025.
Song EJ, Sohn Y-M, Seo M. Tumor stiffness measured by quantitative and qualitative shear wave elastography of breast cancer. Br J Radiol. 2018;91(1086):20170830.
Broders-Bondon F, Nguyen Ho-Bouldoires TH, Fernandez-Sanchez M-E, Farge E. Mechanotransduction in tumor progression: the dark side of the force. J Cell Biol. 2018;217(5):1571–87.
Levental KR, Yu H, Kass L, Lakins JN, Egeblad M, Erler JT, Fong SF, Csiszar K, Giaccia A, Weninger W. Matrix crosslinking forces tumor progression by enhancing integrin signaling. Cell. 2009;139(5):891–906.
Mouw JK, Yui Y, Damiano L, Bainer RO, Lakins JN, Acerbi I, Ou G, Wijekoon AC, Levental KR, Gilbert PM. Tissue mechanics modulate microRNA-dependent PTEN expression to regulate malignant progression. Nat Med. 2014;20(4):360.
Wu X, Cai J, Zuo Z, Li J. Collagen facilitates the colorectal cancer stemness and metastasis through an integrin/PI3K/AKT/Snail signaling pathway. Biomed Pharmacother. 2019;114:108708.
Xu B, Song G, Ju Y, Li X, Song Y, Watanabe S. RhoA/ROCK, cytoskeletal dynamics, and focal adhesion kinase are required for mechanical stretch-induced tenogenic differentiation of human mesenchymal stem cells. J Cell Physiol. 2012;227(6):2722–9.
Calvo F, Ege N, Grande-Garcia A, Hooper S, Jenkins RP, Chaudhry SI, Harrington K, Williamson P, Moeendarbary E, Charras G. Mechanotransduction and YAP-dependent matrix remodelling is required for the generation and maintenance of cancer-associated fibroblasts. Nat Cell Biol. 2013;15(6):637.
Panciera T, Azzolin L, Cordenonsi M, Piccolo S. Mechanobiology of YAP and TAZ in physiology and disease. Nat Rev Mol Cell Biol. 2017;18(12):758.
Kim M-K, Jang J-W, Bae S-C. DNA binding partners of YAP/TAZ. BMB Rep. 2018;51(3):126.
Chen WLK, Simmons CA. Lessons from (patho) physiological tissue stiffness and their implications for drug screening, drug delivery and regenerative medicine. Adv Drug Deliv Rev. 2011;63(4–5):269–76.
Zhao D, Xue C, Li Q, Liu M, Ma W, Zhou T, Lin Y. Substrate stiffness regulated migration and angiogenesis potential of A549 cells and HUVECs. J Cell Physiol. 2018;233(4):3407–17.
Rice A, Cortes E, Lachowski D, Cheung B, Karim S, Morton J, Del Rio HA. Matrix stiffness induces epithelial–mesenchymal transition and promotes chemoresistance in pancreatic cancer cells. Oncogenesis. 2017;6(7):e352.
Fu J, Wang Y-K, Yang MT, Desai RA, Yu X, Liu Z, Chen CS. Mechanical regulation of cell function with geometrically modulated elastomeric substrates. Nat Methods. 2010;7(9):733–6.
Mathew-Steiner SS, Roy S, Sen CK. Collagen in wound healing. Bioengineering. 2021;8(5):63.
Cox TR, Erler JT. Remodeling and homeostasis of the extracellular matrix: implications for fibrotic diseases and cancer. Dis Model Mech. 2011;4(2):165–78.
Chaudhuri PK, Low BC, Lim CT. Mechanobiology of tumor growth. Chem Rev. 2018;118(14):6499–515.
Plodinec M, Loparic M, Monnier CA, Obermann EC, Zanetti-Dallenbach R, Oertle P, Hyotyla JT, Aebi U, Bentires-Alj M, Lim RY. The nanomechanical signature of breast cancer. Nat Nanotechnol. 2012;7(11):757–65.
Paszek MJ, Zahir N, Johnson KR, Lakins JN, Rozenberg GI, Gefen A, Reinhart-King CA, Margulies SS, Dembo M, Boettiger D. Tensional homeostasis and the malignant phenotype. Cancer Cell. 2005;8(3):241–54.
LeBlanc L, Ramirez N, Kim J. Context-dependent roles of YAP/TAZ in stem cell fates and cancer. Cell Mol Life Sci. 2021;78(9):4201–19.
Zanconato F, Forcato M, Battilana G, Azzolin L, Quaranta E, Bodega B, Rosato A, Bicciato S, Cordenonsi M, Piccolo S. Genome-wide association between YAP/TAZ/TEAD and AP-1 at enhancers drives oncogenic growth. Nat Cell Biol. 2015;17(9):1218.
Jang W, Kim T, Koo JS, Kim SK, Lim DS. Mechanical cue-induced YAP instructs Skp2-dependent cell cycle exit and oncogenic signaling. EMBO J. 2017;36(17):2510–28.
Panciera T, Azzolin L, Fujimura A, Di Biagio D, Frasson C, Bresolin S, Soligo S, Basso G, Bicciato S, Rosato A. Induction of expandable tissue-specific stem/progenitor cells through transient expression of YAP/TAZ. Cell Stem Cell. 2016;19(6):725–37.
Cordenonsi M, Zanconato F, Azzolin L, Forcato M, Rosato A, Frasson C, Inui M, Montagner M, Parenti AR, Poletti A. The Hippo transducer TAZ confers cancer stem cell-related traits on breast cancer cells. Cell. 2011;147(4):759–72.
Basu-Roy U, Bayin NS, Rattanakorn K, Han E, Placantonakis DG, Mansukhani A, Basilico C. Sox2 antagonizes the Hippo pathway to maintain stemness in cancer cells. Nat Commun. 2015;6(1):1–14.
Lin L, Sabnis AJ, Chan E, Olivas V, Cade L, Pazarentzos E, Asthana S, Neel D, Yan JJ, Lu X. The Hippo effector YAP promotes resistance to RAF-and MEK-targeted cancer therapies. Nat Genet. 2015;47(3):250.
Zanconato F, Cordenonsi M, Piccolo S. YAP/TAZ at the roots of cancer. Cancer Cell. 2016;29(6):783–803.
Zhao B, Li L, Wang L, Wang C-Y, Yu J, Guan K-L. Cell detachment activates the Hippo pathway via cytoskeleton reorganization to induce anoikis. Genes Dev. 2012;26(1):54–68.
Wei SC, Fattet L, Tsai JH, Guo Y, Pai VH, Majeski HE, Chen AC, Sah RL, Taylor SS, Engler AJ. Matrix stiffness drives epithelial–mesenchymal transition and tumour metastasis through a TWIST1–G3BP2 mechanotransduction pathway. Nat Cell Biol. 2015;17(5):678–88.
Ros M, Sala M, Saltel F. Linking matrix rigidity with EMT and cancer invasion. Dev Cell. 2020;54(3):293–5.
Ghasemi H, Mousavibahar SH, Hashemnia M, Karimi J, Khodadadi I, Mirzaei F, Tavilani H. Tissue stiffness contributes to YAP activation in bladder cancer patients undergoing transurethral resection. Ann N Y Acad Sci. 2020;1473(1):48–61.
Mani SA, Guo W, Liao M-J, Eaton EN, Ayyanan A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. 2008;133(4):704–15.
Sarrio D, Franklin CK, Mackay A, Reis-Filho JS, Isacke CM. Epithelial and mesenchymal subpopulations within normal basal breast cell lines exhibit distinct stem cell/progenitor properties. Stem cells. 2012;30(2):292–303.
Najafi M, Mortezaee K, Ahadi R. Cancer stem cell (a) symmetry & plasticity: tumorigenesis and therapy relevance. Life Sci. 2019;231:116520.
Conklin MW, Eickhoff JC, Riching KM, Pehlke CA, Eliceiri KW, Provenzano PP, Friedl A, Keely PJ. Aligned collagen is a prognostic signature for survival in human breast carcinoma. Am J Pathol. 2011;178(3):1221–32.
Fenner J, Stacer AC, Winterroth F, Johnson TD, Luker KE, Luker GD. Macroscopic stiffness of breast tumors predicts metastasis. Sci Rep. 2014;4(1):1–8.
Singh A, Brito I, Lammerding J. Beyond tissue stiffness and bioadhesivity: advanced biomaterials to model tumor microenvironments and drug resistance. Trends in cancer. 2018;4(4):281–91.
Chang F-C, Tsao C-T, Lin A, Zhang M, Levengood SL, Zhang M. PEG-chitosan hydrogel with tunable stiffness for study of drug response of breast cancer cells. Polymers. 2016;8(4):112.
Qin X, Lv X, Li P, Yang R, Xia Q, Chen Y, Peng Y, Li L, Li S, Li T. Matrix stiffness modulates ILK-mediated YAP activation to control the drug resistance of breast cancer cells. Biochim Biophys Acta (BBA) Mol Basis Dis. 2020;1866(3):165625.
Provenzano PP, Cuevas C, Chang AE, Goel VK, Von Hoff DD, Hingorani SR. Enzymatic targeting of the stroma ablates physical barriers to treatment of pancreatic ductal adenocarcinoma. Cancer Cell. 2012;21(3):418–29.
Hayashi M, Yamamoto Y, Ibusuki M, Fujiwara S, Yamamoto S, Tomita S, Nakano M, Murakami K, Iyama K-I, Iwase H. Evaluation of tumor stiffness by elastography is predictive for pathologic complete response to neoadjuvant chemotherapy in patients with breast cancer. Annal Surg Oncol. 2012;19(9):3042–9.
Jing H, Cheng W, Li Z-Y, Ying L, Wang Q-C, Wu T, Tian J-W. Early evaluation of relative changes in tumor stiffness by shear wave elastography predicts the response to neoadjuvant chemotherapy in patients with breast cancer. J Ultrasound Med. 2016;35(8):1619–27.
Kim B, Kim SS, Cho SW, Cheong JY, Huh J, Kim JK, Lee JH, Ahn HR, Cho HJ. Liver stiffness in magnetic resonance elastography is prognostic for sorafenib-treated advanced hepatocellular carcinoma. Eur Radiol. 2021;31(4):2507–17.
Lau RS-T, Ip P, Wong GL-H, Wong VW-S, Lo EJ-Y, Wong KK-C, Fung AK-Y, Wong J, Lee K-F, Ng KK-C. Liver stiffness measurement predicts short-term and long-term outcomes in patients with hepatocellular carcinoma after curative liver resection. Surgeon. 2022;20(2):78–84.
You Y, Zheng Q, Dong Y, Wang Y, Zhang L, Xue T, Xie X, Hu C, Wang Z, Chen R. Higher matrix stiffness upregulates osteopontin expression in hepatocellular carcinoma cells mediated by integrin β1/GSK3β/β-catenin signaling pathway. PLoS ONE. 2015;10(8):e0134243.
Miyazawa A, Ito S, Asano S, Tanaka I, Sato M, Kondo M, Hasegawa Y. Regulation of PD-L1 expression by matrix stiffness in lung cancer cells. Biochem Biophys Res Commun. 2018;495(3):2344–9.
Bougherara H, Mansuet-Lupo A, Alifano M, Ngô C, Damotte D, Le Frère-Belda M-A, Donnadieu E, Peranzoni E. Real-time imaging of resident T cells in human lung and ovarian carcinomas reveals how different tumor microenvironments control T lymphocyte migration. Front Immunol. 2015;6:500.
Barenholz-Cohen T, Merkher Y, Haj J, Shechter D, Kirchmeier D, Shaked Y, Weihs D. Lung mechanics modifications facilitating metastasis are mediated in part by breast cancer-derived extracellular vesicles. Int J Cancer. 2020;147(10):2924–33.
Wu S, Zheng Q, Xing X, Dong Y, Wang Y, You Y, Chen R, Hu C, Chen J, Gao D. Matrix stiffness-upregulated LOXL2 promotes fibronectin production, MMP9 and CXCL12 expression and BMDCs recruitment to assist pre-metastatic niche formation. J Exp Clin Cancer Res. 2018;37(1):1–12.
Wong CCL, Tse APW, Huang YP, Zhu YT, Chiu DKC, Lai RKH, Au SLK, Kai AKL, Lee JMF, Wei LL. Lysyl oxidase-like 2 is critical to tumor microenvironment and metastatic niche formation in hepatocellular carcinoma. Hepatology. 2014;60(5):1645–58.
Chu H-Y, Chen Y-J, Hsu C-J, Liu Y-W, Chiou J-F, Lu L-S, Tseng F-G. Physical cues in the microenvironment regulate stemness-dependent homing of breast cancer cells. Cancers. 2020;12(8):2176.
Patwardhan S, Mahadik P, Shetty O, Sen S. ECM stiffness-tuned exosomes drive breast cancer motility through thrombospondin-1. Biomaterials. 2021;279:121185.
Jacquemin V, Antoine M, Dom G, Detours V, Maenhaut C, Dumont JE. Dynamic cancer cell heterogeneity: diagnostic and therapeutic implications. Cancers. 2022;14(2):280.
Tirino V, Desiderio V, Paino F, De Rosa A, Papaccio F, La Noce M, Laino L, De Francesco F, Papaccio G. Cancer stem cells in solid tumors: an overview and new approaches for their isolation and characterization. FASEB J. 2013;27(1):13–24.
Dang HT, Budhu A, Wang XW. The origin of cancer stem cells. J Hepatol. 2014;60(6):1304–5.
Bonnet D, Dick JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med. 1997;3(7):730–7.
Beltran AS, Rivenbark AG, Richardson BT, Yuan X, Quian H, Hunt JP, Zimmerman E, Graves LM, Blancafort P. Generation of tumor-initiating cells by exogenous delivery of OCT4transcription factor. Breast Cancer Res. 2011;13(5):1–21.
Jeter CR, Badeaux M, Choy G, Chandra D, Patrawala L, Liu C, Calhoun-Davis T, Zaehres H, Daley GQ, Tang DG. Functional evidence that the self-renewal gene NANOG regulates human tumor development. Stem cells. 2009;27(5):993–1005.
Medema JP. Cancer stem cells: the challenges ahead. Nat Cell Biol. 2013;15(4):338–44.
Xie XP, Laks DR, Sun D, Ganbold M, Wang Z, Pedraza AM, Bale T, Tabar V, Brennan C, Zhou X. Quiescent human glioblastoma cancer stem cells drive tumor initiation, expansion, and recurrence following chemotherapy. Dev Cell. 2022;57(1):32-46.e38.
Enderling H, Hlatky L, Hahnfeldt P. Cancer stem cells: a minor cancer subpopulation that redefines global cancer features. Front Oncol. 2013;3:76.
Zheng X, Yu C, Xu M. Linking tumor microenvironment to plasticity of cancer stem cells: mechanisms and application in cancer therapy. Front Oncol. 2021;11:2552.
Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix elasticity directs stem cell lineage specification. Cell. 2006;126(4):677–89.
Pek YS, Wan AC, Ying JY. The effect of matrix stiffness on mesenchymal stem cell differentiation in a 3D thixotropic gel. Biomaterials. 2010;31(3):385–91.
Plaks V, Kong N, Werb Z. The cancer stem cell niche: how essential is the niche in regulating stemness of tumor cells? Cell Stem Cell. 2015;16(3):225–38.
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–49.
DeSantis CE, Ma J, Gaudet MM, Newman LA, Miller KD, Goding Sauer A, Jemal A, Siegel RL. Breast cancer statistics, 2019. CA Cancer J Clin. 2019;69(6):438–51.
McKnight AL, Kugel JL, Rossman PJ, Manduca A, Hartmann LC, Ehman RL. MR elastography of breast cancer: preliminary results. Am J Roentgenol. 2002;178(6):1411–7.
Boyd NF, Martin LJ, Yaffe MJ, Minkin S. Mammographic density and breast cancer risk: current understanding and future prospects. Breast Cancer Res. 2011;13(6):223.
Case A, Brisson BK, Durham AC, Rosen S, Monslow J, Buza E, Salah P, Gillem J, Ruthel G, Veluvolu S. Identification of prognostic collagen signatures and potential therapeutic stromal targets in canine mammary gland carcinoma. PLoS ONE. 2017;12(7):e0180448.
Yoo J, Seo BK, Park EK, Kwon M, Jeong H, Cho KR, Woo OH, Song SE, Cha J. Tumor stiffness measured by shear wave elastography correlates with tumor hypoxia as well as histologic biomarkers in breast cancer. Cancer Imaging. 2020;20(1):1–10.
Xi G, Qiu L, Xu S, Guo W, Fu F, Kang D, Zheng L, He J, Zhang Q, Li L. Computer-assisted quantification of tumor-associated collagen signatures to improve the prognosis prediction of breast cancer. BMC Med. 2021;19:1–16.
Chang JM, Park I, Lee SH, Kim WH, Bae MS, Koo HR, Yi A, Kim SJ, Cho N, Moon WK. Stiffness of tumours measured by shear-wave elastography correlated with subtypes of breast cancer. Eur Radiol. 2013;23(9):2450–8.
Bauerschmitz GJ, Ranki T, Kangasniemi L, Ribacka C, Eriksson M, Porten M, Herrmann I, Ristimäki A, Virkkunen P, Tarkkanen M. Tissue-specific promoters active in CD44+ CD24−/low breast cancer cells. Can Res. 2008;68(14):5533–9.
Ginestier C, Hur MH, Charafe-Jauffret E, Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG, Liu S. ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell Stem Cell. 2007;1(5):555–67.
Kim T, Yang S-J, Hwang D, Song J, Kim M, Kyum Kim S, Kang K, Ahn J, Lee D, Kim M-Y. A basal-like breast cancer-specific role for SRF–IL6 in YAP-induced cancer stemness. Nat Commun. 2015;6(1):1–15.
Idowu MO, Kmieciak M, Dumur C, Burton RS, Grimes MM, Powers CN, Manjili MH. CD44+/CD24−/low cancer stem/progenitor cells are more abundant in triple-negative invasive breast carcinoma phenotype and are associated with poor outcome. Hum Pathol. 2012;43(3):364–73.
Triantafillu UL, Park S, Klaassen NL, Raddatz AD, Kim Y. Fluid shear stress induces cancer stem cell-like phenotype in MCF7 breast cancer cell line without inducing epithelial to mesenchymal transition. Int J Oncol. 2017;50(3):993–1001.
Balachander GM, Balaji SA, Rangarajan A, Chatterjee K. Enhanced metastatic potential in a 3D tissue scaffold toward a comprehensive in vitro model for breast cancer metastasis. ACS Appl Mater Interfaces. 2015;7(50):27810–22.
Ray A, Slama ZM, Morford RK, Madden SA, Provenzano PP. Enhanced directional migration of cancer stem cells in 3D aligned collagen matrices. Biophys J. 2017;112(5):1023–36.
Jabbari E, Sarvestani SK, Daneshian L, Moeinzadeh S. Optimum 3D matrix stiffness for maintenance of cancer stem cells is dependent on tissue origin of cancer cells. PLoS ONE. 2015;10(7):e0132377.
Yang X, Sarvestani SK, Moeinzadeh S, He X, Jabbari E. Three-dimensional-engineered matrix to study cancer stem cells and tumorsphere formation: effect of matrix modulus. Tissue Eng Part A. 2013;19(5–6):669–84.
Berger AJ, Renner CM, Hale I, Yang X, Ponik SM, Weisman PS, Masters KS, Kreeger PK. Scaffold stiffness influences breast cancer cell invasion via EGFR-linked Mena upregulation and matrix remodeling. Matrix Biol. 2020;85:80–93.
Hu K, Huang P, Luo H, Yao Z, Wang Q, Xiong Z, Lin J, Huang H, Xu S, Zhang P. Mammalian-enabled (MENA) protein enhances oncogenic potential and cancer stem cell-like phenotype in hepatocellular carcinoma cells. FEBS Open Bio. 2017;7(8):1144–53.
Shea MP, O’Leary KA, Wegner KA, Vezina CM, Schuler LA. High collagen density augments mTOR-dependent cancer stem cells in ERα+ mammary carcinomas, and increases mTOR-independent lung metastases. Cancer Lett. 2018;433:1–9.
Kumar P, Goldstraw P, Yamada K, Nicholson A, Wells A, Hansell D, Ladas G. Pulmonary fibrosis and lung cancer: risk and benefit analysis of pulmonary resection. J Thorac Cardiovasc Surg. 2003;125(6):1321–7.
Voiles L, Lewis DE, Han L, Lupov IP, Lin T-L, Robertson MJ, Petrache I, Chang H-C. Overexpression of type VI collagen in neoplastic lung tissues. Oncol Rep. 2014;32(5):1897–904.
Rachman-Tzemah C, Zaffryar-Eilot S, Grossman M, Ribero D, Timaner M, Mäki JM, Myllyharju J, Bertolini F, Hershkovitz D, Sagi I. Blocking surgically induced lysyl oxidase activity reduces the risk of lung metastases. Cell Rep. 2017;19(4):774–84.
Burns TF, Borghaei H, Ramalingam SS, Mok TS, Peters S. Targeting KRAS-mutant non–small-cell lung cancer: one mutation at a time, with a focus on KRAS G12C mutations. J Clin Oncol. 2020;38(35):4208.
Kamikawaji K, Seki N, Watanabe M, Mataki H, Kumamoto T, Takagi K, Mizuno K, Inoue H. Regulation of LOXL2 and SERPINH1 by antitumor microRNA-29a in lung cancer with idiopathic pulmonary fibrosis. J Hum Genet. 2016;61(12):985–93.
Eichberger J, Schulz D, Pscheidl K, Fiedler M, Reichert TE, Bauer RJ, Ettl T. PD-L1 influences cell spreading, migration and invasion in head and neck cancer cells. Int J Mol Sci. 2020;21(21):8089.
Lee BS, Park DI, Lee DH, Lee JE, Yeo M-K, Park YH, Lim DS, Choi W, Yoo G, Kim H-B. Hippo effector YAP directly regulates the expression of PD-L1 transcripts in EGFR-TKI-resistant lung adenocarcinoma. Biochem Biophys Res Commun. 2017;491(2):493–9.
Zhang C, Wang H, Wang X, Zhao C, Wang H. CD44, a marker of cancer stem cells, is positively correlated with PD-L1 expression and immune cells infiltration in lung adenocarcinoma. Cancer Cell Int. 2020;20(1):1–8.
Heng WS, Gosens R, Kruyt FA. Lung cancer stem cells: origin, features, maintenance mechanisms and therapeutic targeting. Biochem Pharmacol. 2019;160:121–33.
Tremblay AM, Camargo FD. Hippo signaling in mammalian stem cells. Semin Cell Dev Biol. 2012;23:818–26.
Chang C-C, Hsieh T-L, Tiong T-Y, Hsiao C-H, Ji AT-Q, Hsu W-T, Lee OK, Ho JH. Regulation of metastatic ability and drug resistance in pulmonary adenocarcinoma by matrix rigidity via activating c-Met and EGFR. Biomaterials. 2015;60:141–50.
Khater AR, Abou-Antoun T. Mesenchymal epithelial transition factor signaling in pediatric nervous system tumors: implications for malignancy and cancer stem cell enrichment. Front Cell Dev Biol. 2021;9:654103.
Herreros-Pomares A, Zhou X, Calabuig-Fariñas S, Lee S-J, Torres S, Esworthy T, Hann SY, Jantus-Lewintre E, Camps C, Zhang LG. 3D printing novel in vitro cancer cell culture model systems for lung cancer stem cell study. Mater Sci Eng C. 2021;122:111914.
Doyle AD, Yamada KM. Mechanosensing via cell-matrix adhesions in 3D microenvironments. Exp Cell Res. 2016;343(1):60–6.
Nault JC, Bioulac-Sage P, Zucman-Rossi J. Hepatocellular benign tumors—from molecular classification to personalized clinical care. Gastroenterology. 2013;144(5):888–902.
Masuzaki R, Tateishi R, Yoshida H, Sato T, Ohki T, Goto T, Yoshida H, Sato S, Sugioka Y, Ikeda H. Assessing liver tumor stiffness by transient elastography. Hep Intl. 2007;1(3):394–7.
Singal AG, El-Serag HB. Hepatocellular carcinoma from epidemiology to prevention: translating knowledge into practice. Clin Gastroenterol Hepatol. 2015;13(12):2140–51.
Nordenstedt H, White DL, El-Serag HB. The changing pattern of epidemiology in hepatocellular carcinoma. Dig Liver Dis. 2010;42:S206–14.
Rojkind M, Giambrone M-A, Biempica L. Collagen types in normal and cirrhotic liver. Gastroenterology. 1979;76(4):710–9.
Wang HM, Hung CH, Lu SN, Chen CH, Lee CM, Hu TH, Wang JH. Liver stiffness measurement as an alternative to fibrotic stage in risk assessment of hepatocellular carcinoma incidence for chronic hepatitis C patients. Liver Int. 2013;33(5):756–61.
Sandrin L, Fourquet B, Hasquenoph J-M, Yon S, Fournier C, Mal F, Christidis C, Ziol M, Poulet B, Kazemi F. Transient elastography: a new noninvasive method for assessment of hepatic fibrosis. Ultrasound Med Biol. 2003;29(12):1705–13.
Wang C, Jiang X, Huang B, Zhou W, Cui X, Zheng C, Liu F, Bi J, Zhang Y, Luo H. Inhibition of matrix stiffness relating integrin β1 signaling pathway inhibits tumor growth in vitro and in hepatocellular cancer xenografts. BMC Cancer. 2021;21(1):1–11.
Lee DH, Lee JM, Yoon J-H, Kim YJ, Lee J-H, Yu SJ, Han JK. Liver stiffness measured by two-dimensional shear-wave elastography: prognostic value after radiofrequency ablation for hepatocellular carcinoma. Liver cancer. 2018;7(1):65–75.
Nio K, Yamashita T, Kaneko S. The evolving concept of liver cancer stem cells. Mol Cancer. 2017;16(1):1–12.
Liu Y-C, Yeh C-T, Lin K-H. Cancer stem cell functions in hepatocellular carcinoma and comprehensive therapeutic strategies. Cells. 2020;9(6):1331.
Yamashita T, Honda M, Nakamoto Y, Baba M, Nio K, Hara Y, Zeng SS, Hayashi T, Kondo M, Takatori H. Discrete nature of EpCAM+ and CD90+ cancer stem cells in human hepatocellular carcinoma. Hepatology. 2013;57(4):1484–97.
Chan LH, Luk ST, Ma S. Turning hepatic cancer stem cells inside out–a deeper understanding through multiple perspectives. Mol Cells. 2015;38(3):202–9.
Yang J, Liao D, Chen C, Liu Y, Chuang T-H, Xiang R, Markowitz D, Reisfeld RA, Luo Y. Tumor-associated macrophages regulate murine breast cancer stem cells through a novel paracrine EGFR/Stat3/Sox-2 signaling pathway. Stem cells. 2013;31(2):248–58.
Zhu P, Wang Y, Wu J, Huang G, Liu B, Ye B, Du Y, Gao G, Tian Y, He L, et al. LncBRM initiates YAP1 signalling activation to drive self-renewal of liver cancer stem cells. Nat Commun. 2016;7:13608.
You Y, Zheng Q, Dong Y, Xie X, Wang Y, Wu S, Zhang L, Wang Y, Xue T, Wang Z, et al. Matrix stiffness-mediated effects on stemness characteristics occurring in HCC cells. Oncotarget. 2016;7(22):32221–31.
Zhang R, Ma M, Dong G, Yao RR, Li JH, Zheng QD, Dong YY, Ma H, Gao DM, Cui JF. Increased matrix stiffness promotes tumor progression of residual hepatocellular carcinoma after insufficient heat treatment. Cancer Sci. 2017;108(9):1778–86.
Lee C-W, Kuo W-L, Yu M-C, Chen T-C, Tsai C-N, Lee W-C, Chen M-F. The expression of cytokeratin 19 in lymph nodes was a poor prognostic factor for hepatocellular carcinoma after hepatic resection. World J Surg Oncol. 2013;11(1):1–11.
Kitamura R, Toyoshima T, Tanaka H, Kawano S, Matsubara R, Goto Y, Jinno T, Maruse Y, Oobu K, Nakamura S. Cytokeratin 17 mRNA as a prognostic marker of oral squamous cell carcinoma. Oncol Lett. 2017;14(6):6735–43.
Yao B, Niu Y, Li Y, Chen T, Wei X, Liu Q. High-matrix-stiffness induces promotion of hepatocellular carcinoma proliferation and suppression of apoptosis via miR-3682-3p-PHLDA1-FAS pathway. J Cancer. 2020;11(21):6188.
Chen Q, Yang S-B, Zhang Y-W, Han S-Y, Jia L, Li B, Zhang Y, Zuo S. miR-3682-3p directly targets FOXO3 and stimulates tumor stemness in hepatocellular carcinoma via a positive feedback loop involving FOXO3/PI3K/AKT/c-Myc. World J Stem Cells. 2022;14(7):539–55.
Zhang Y, Cai H, Wu M-H, Zhu D-H, Wang X-Y, Chen Z-Y, Yang L, Liu P, Liu Z. miR-3682-3p activated by c-Myc aggravates the migration and stemness in hepatocellular carcinoma cells by regulating PTEN/PI3K/AKT/β-catenin signaling. Dig Dis. 2023;41(3):447–57.
Schrader J, Gordon-Walker TT, Aucott RL, van Deemter M, Quaas A, Walsh S, Benten D, Forbes SJ, Wells RG, Iredale JP. Matrix stiffness modulates proliferation, chemotherapeutic response, and dormancy in hepatocellular carcinoma cells. Hepatology. 2011;53(4):1192–205.
Yang N, Chen T, Wang L, Liu R, Niu Y, Sun L, Yao B, Wang Y, Yang W, Liu Q. CXCR4 mediates matrix stiffness-induced downregulation of UBTD1 driving hepatocellular carcinoma progression via YAP signaling pathway. Theranostics. 2020;10(13):5790–801.
Cheng C-W, Liao W-L, Chen P-M, Yu J-C, Shiau H-P, Hsieh Y-H, Lee H-J, Cheng Y-C, Wu P-E, Shen C-Y. miR-139 modulates cancer stem cell function of human breast cancer through targeting CXCR4. Cancers. 2021;13(11):2582.
Gassenmaier M, Chen D, Buchner A, Henkel L, Schiemann M, Mack B, Schendel DJ, Zimmermann W, Pohla H. CXC chemokine receptor 4 is essential for maintenance of renal cell carcinoma-initiating cells and predicts metastasis. Stem Cells. 2013;31(8):1467–76.
Shen Y, Wang X, Lu J, Salfenmoser M, Wirsik NM, Schleussner N, Imle A, Valls AF, Radhakrishnan P, Liang J. Reduction of liver metastasis stiffness improves response to bevacizumab in metastatic colorectal cancer. Cancer Cell. 2020;37(6):800-817.e807.
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424.
Hosein AN, Brekken RA, Maitra A. Pancreatic cancer stroma: an update on therapeutic targeting strategies. Nat Rev Gastroenterol Hepatol. 2020;17(8):487–505.
Rahib L, Smith BD, Aizenberg R, Rosenzweig AB, Fleshman JM, Matrisian LM. Projecting cancer incidence and deaths to 2030: the unexpected burden of thyroid, liver, and pancreas cancers in the United States. Can Res. 2014;74(11):2913–21.
Neesse A, Bauer CA, Öhlund D, Lauth M, Buchholz M, Michl P, Tuveson DA, Gress TM. Stromal biology and therapy in pancreatic cancer: ready for clinical translation? Gut. 2019;68(1):159–71.
Nabavizadeh A, Payen T, Iuga AC, Sagalovskiy IR, Desrouilleres D, Saharkhiz N, Palermo CF, Sastra SA, Oberstein PE, Rosario V. Noninvasive Young’s modulus visualization of fibrosis progression and delineation of pancreatic ductal adenocarcinoma (PDAC) tumors using harmonic motion elastography (HME) in vivo. Theranostics. 2020;10(10):4614.
Laklai H, Miroshnikova YA, Pickup MW, Collisson EA, Kim GE, Barrett AS, Hill RC, Lakins JN, Schlaepfer DD, Mouw JK. Genotype tunes pancreatic ductal adenocarcinoma tissue tension to induce matricellular fibrosis and tumor progression. Nat Med. 2016;22(5):497–505.
Shi S, Liang C, Xu J, Meng Q, Hua J, Yang X, Ni Q, Yu X. The strain ratio as obtained by endoscopic ultrasonography elastography correlates with the stroma proportion and the prognosis of local pancreatic cancer. Ann Surg. 2020;271(3):559–65.
Shields MA, Dangi-Garimella S, Krantz SB, Bentrem DJ, Munshi HG. Pancreatic cancer cells respond to type I collagen by inducing snail expression to promote membrane type 1 matrix metalloproteinase-dependent collagen invasion. J Biol Chem. 2011;286(12):10495–504.
Zhang W, Nandakumar N, Shi Y, Manzano M, Smith A, Graham G, Gupta S, Vietsch EE, Laughlin SZ, Wadhwa M. Downstream of mutant KRAS, the transcription regulator YAP is essential for neoplastic progression to pancreatic ductal adenocarcinoma. Sci Signal. 2014;7(324):ra42.
Kapoor A, Yao W, Ying H, Hua S, Liewen A, Wang Q, Zhong Y, Wu C-J, Sadanandam A, Hu B. Yap1 activation enables bypass of oncogenic Kras addiction in pancreatic cancer. Cell. 2014;158(1):185–97.
Hermann PC, Sainz B Jr. Pancreatic cancer stem cells: a state or an entity? Semin Cancer Biol. 2018;53:223–31.
Takao S, Ding Q, Matsubara S. Pancreatic cancer stem cells: regulatory networks in the tumor microenvironment and targeted therapy. J Hepatobiliary Pancreat Sci. 2012;19(6):614–20.
Kanat O, Ertas H. Shattering the castle walls: Anti-stromal therapy for pancreatic cancer. World J Gastrointest Oncol. 2018;10(8):202–10.
Zhang L, Shi H, Chen H, Gong A, Liu Y, Song L, Xu X, You T, Fan X, Wang D. Dedifferentiation process driven by radiotherapy-induced HMGB1/TLR2/YAP/HIF-1α signaling enhances pancreatic cancer stemness. Cell Death Dis. 2019;10(10):1–16.
Chen W, Wang H, Liu Y, Xu W, Ling C, Li Y, Liu J, Chen M, Zhang Y, Chen B. Linc-RoR promotes proliferation, migration, and invasion via the Hippo/YAP pathway in pancreatic cancer cells. J Cell Biochem. 2020;121(1):632–41.
Su S, Chen J, Yao H, Liu J, Yu S, Lao L, Wang M, Luo M, Xing Y, Chen F. CD10+ GPR77+ cancer-associated fibroblasts promote cancer formation and chemoresistance by sustaining cancer stemness. Cell. 2018;172(4):841-856.e816.
LeSavage BL, Gilchrist AE, Krajina BA, Karlsson K, Smith AR, Karagyozova K, Klett KC, Curtis C, Kuo CJ, Heilshorn SC. Engineered extracellular matrices reveal stiffness-mediated chemoresistance in patient-derived pancreatic cancer organoids. bioRxiv. 2022. https://doi.org/10.1101/2022.04.22.488943.
Petriz J. Flow cytometry of the side population (SP). Curr Protoc Cytom. 2013;64(1):9.23.21-29.23.20.
Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer statistics, 2021. CA Cancer J Clin. 2021;71(1):7–33.
Tang X, Kuhlenschmidt TB, Li Q, Ali S, Lezmi S, Chen H, Pires-Alves M, Laegreid WW, Saif TA, Kuhlenschmidt MS. A mechanically-induced colon cancer cell population shows increased metastatic potential. Mol Cancer. 2014;13(1):1–15.
Palmieri V, Lucchetti D, Maiorana A, Papi M, Maulucci G, Calapà F, Ciasca G, Giordano R, Sgambato A, De Spirito M. Mechanical and structural comparison between primary tumor and lymph node metastasis cells in colorectal cancer. Soft Matter. 2015;11(28):5719–26.
Despotović SZ, Milićević NM, Milošević DP, Despotović N, Erceg P, Svorcan P, Schumacher U, Ullrich S, Mihajlović G, Kalem D. Remodeling of extracellular matrix of the lamina propria in the uninvolved human rectal mucosa 10 and 20 cm away from the malignant tumor. Tumor Biology. 2017;39(7):1010428317711654.
Yuen H-F, McCrudden CM, Huang Y-H, Tham JM, Zhang X, Zeng Q, Zhang S-D, Hong W. TAZ expression as a prognostic indicator in colorectal cancer. PLoS ONE. 2013;8(1):e54211.
Lee K-W, Lee SS, Kim S-B, Sohn BH, Lee H-S, Jang H-J, Park Y-Y, Kopetz S, Kim SS, Oh SC. Significant association of oncogene YAP1 with poor prognosis and cetuximab resistance in colorectal cancer patients. Clin Cancer Res. 2015;21(2):357–64.
Touil Y, Igoudjil W, Corvaisier M, Dessein A-F, Vandomme J, Monté D, Stechly L, Skrypek N, Langlois C, Grard G. Colon cancer cells escape 5FU chemotherapy-induced cell death by entering stemness and quiescence associated with the c-Yes/YAP axis. Clin Cancer Res. 2014;20(4):837–46.
Zhou D, Zhang Y, Wu H, Barry E, Yin Y, Lawrence E, Dawson D, Willis JE, Markowitz SD, Camargo FD. Mst1 and Mst2 protein kinases restrain intestinal stem cell proliferation and colonic tumorigenesis by inhibition of Yes-associated protein (Yap) overabundance. Proc Natl Acad Sci. 2011;108(49):E1312–20.
Kawano S, Kojima M, Higuchi Y, Sugimoto M, Ikeda K, Sakuyama N, Takahashi S, Hayashi R, Ochiai A, Saito N. Assessment of elasticity of colorectal cancer tissue, clinical utility, pathological and phenotypical relevance. Cancer Sci. 2015;106(9):1232–9.
Ricci-Vitiani L, Lombardi DG, Pilozzi E, Biffoni M, Todaro M, Peschle C, De Maria R. Identification and expansion of human colon-cancer-initiating cells. Nature. 2007;445(7123):111–5.
Huang EH, Hynes MJ, Zhang T, Ginestier C, Dontu G, Appelman H, Fields JZ, Wicha MS, Boman BM. Aldehyde dehydrogenase 1 is a marker for normal and malignant human colonic stem cells (SC) and tracks SC overpopulation during colon tumorigenesis. Can Res. 2009;69(8):3382–9.
Vermeulen L, Todaro M, de Sousa MF, Sprick MR, Kemper K, Perez Alea M, Richel DJ, Stassi G, Medema JP. Single-cell cloning of colon cancer stem cells reveals a multi-lineage differentiation capacity. Proc Natl Acad Sci. 2008;105(36):13427–32.
Kirkland S. Type I collagen inhibits differentiation and promotes a stem cell-like phenotype in human colorectal carcinoma cells. Br J Cancer. 2009;101(2):320–6.
Tan F, Huang Y, Pei Q, Liu H, Pei H, Zhu H. Matrix stiffness mediates stemness characteristics via activating the Yes-associated protein in colorectal cancer cells. J Cell Biochem. 2019;120(2):2213–25.
Tang X, Kuhlenschmidt TB, Zhou J, Bell P, Wang F, Kuhlenschmidt MS, Saif TA. Mechanical force affects expression of an in vitro metastasis-like phenotype in HCT-8 cells. Biophys J. 2010;99(8):2460–9.
Deng S, Yang X, Lassus H, Liang S, Kaur S, Ye Q, Li C, Wang L-P, Roby KF, Orsulic S. Distinct expression levels and patterns of stem cell marker, aldehyde dehydrogenase isoform 1 (ALDH1), in human epithelial cancers. PLoS ONE. 2010;5(4):e10277.
Sullivan JP, Spinola M, Dodge M, Raso MG, Behrens C, Gao B, Schuster K, Shao C, Larsen JE, Sullivan LA. Aldehyde dehydrogenase activity selects for lung adenocarcinoma stem cells dependent on notch signaling. Can Res. 2010;70(23):9937–48.
Bauer J, Emon MAB, Staudacher JJ, Thomas AL, Zessner-Spitzenberg J, Mancinelli G, Krett N, Saif MT, Jung B. Increased stiffness of the tumor microenvironment in colon cancer stimulates cancer associated fibroblast-mediated prometastatic activin A signaling. Sci Rep. 2020;10(1):1–11.
Pauklin S, Vallier L. Activin/Nodal signalling in stem cells. Development. 2015;142(4):607–19.
Lonardo E, Hermann PC, Mueller M-T, Huber S, Balic A, Miranda-Lorenzo I, Zagorac S, Alcala S, Rodriguez-Arabaolaza I, Ramirez JC. Nodal/Activin signaling drives self-renewal and tumorigenicity of pancreatic cancer stem cells and provides a target for combined drug therapy. Cell Stem Cell. 2011;9(5):433–46.
Chen L, De Menna M, Groenewoud A, Thalmann GN, Kruithof-de Julio M, Snaar-Jagalska BE. A NF-ĸB-Activin A signaling axis enhances prostate cancer metastasis. Oncogene. 2020;39(8):1634–51.
Donnenberg VS, Donnenberg AD. Multiple drug resistance in cancer revisited: the cancer stem cell hypothesis. J Clin Pharmacol. 2005;45(8):872–7.
Hashemi F, Razmi M, Tajik F, Zöller M, Dehghan Manshadi M, Mahdavinezhad F, Tiyuri A, Ghods R, Madjd Z. Efficacy of whole cancer stem cell-based vaccines: a systematic review of preclinical and clinical studies. Stem Cells 2023; 41(3):207–232
Li Y, Rogoff HA, Keates S, Gao Y, Murikipudi S, Mikule K, Leggett D, Li W, Pardee AB, Li CJ. Suppression of cancer relapse and metastasis by inhibiting cancer stemness. Proc Natl Acad Sci. 2015;112(6):1839–44.
Vermeulen L, Snippert HJ. Stem cell dynamics in homeostasis and cancer of the intestine. Nat Rev Cancer. 2014;14(7):468–80.
Kozar S, Morrissey E, Nicholson AM, van der Heijden M, Zecchini HI, Kemp R, Tavare S, Vermeulen L, Winton DJ. Continuous clonal labeling reveals small numbers of functional stem cells in intestinal crypts and adenomas. Cell Stem Cell. 2013;13(5):626–33.
Snippert HJ, Van Der Flier LG, Sato T, Van Es JH, Van Den Born M, Kroon-Veenboer C, Barker N, Klein AM, Van Rheenen J, Simons BD. Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lgr5 stem cells. Cell. 2010;143(1):134–44.
Vermeulen L, De Melo FSE, Van Der Heijden M, Cameron K, De Jong JH, Borovski T, Tuynman JB, Todaro M, Merz C, Rodermond H. Wnt activity defines colon cancer stem cells and is regulated by the microenvironment. Nat Cell Biol. 2010;12(5):468–76.
Lenos KJ, Miedema DM, Lodestijn SC, Nijman LE, van den Bosch T, Romero Ros X, Lourenço FC, Lecca MC, van der Heijden M, van Neerven SM. Stem cell functionality is microenvironmentally defined during tumour expansion and therapy response in colon cancer. Nat Cell Biol. 2018;20(10):1193–202.
Zinger A, Koren L, Adir O, Poley M, Alyan M, Yaari Z, Noor N, Krinsky N, Simon A, Gibori H. Collagenase nanoparticles enhance the penetration of drugs into pancreatic tumors. ACS Nano. 2019;13(10):11008–21.
Dolor A, Szoka FC Jr. Digesting a path forward: the utility of collagenase tumor treatment for improved drug delivery. Mol Pharm. 2018;15(6):2069–83.
Miller BW, Morton JP, Pinese M, Saturno G, Jamieson NB, McGhee E, Timpson P, Leach J, McGarry L, Shanks E. Targeting the LOX/hypoxia axis reverses many of the features that make pancreatic cancer deadly: inhibition of LOX abrogates metastasis and enhances drug efficacy. EMBO Mol Med. 2015;7(8):1063–76.
Zhang L, Wang Y, Xia T, Yu Q, Zhang Q, Yang Y, Cun X, Lu L, Gao H, Zhang Z. Suppression for lung metastasis by depletion of collagen I and lysyl oxidase via losartan assisted with paclitaxel-loaded pH-sensitive liposomes in breast cancer. Drug Delivery. 2016;23(8):2970–9.
Wei Y, Kim TJ, Peng DH, Duan D, Gibbons DL, Yamauchi M, Jackson JR, Le Saux CJ, Calhoun C, Peters J. Fibroblast-specific inhibition of TGF-β1 signaling attenuates lung and tumor fibrosis. J Clin Investig. 2017;127(10):3675–88.
Jensen BV, Johansen JS, Skovsgaard T, Brandt J, Teisner B. Extracellular matrix building marked by the N-terminal propeptide of procollagen type I reflect aggressiveness of recurrent breast cancer. Int J Cancer. 2002;98(4):582–9.
Barkan D, El Touny LH, Michalowski AM, Smith JA, Chu I, Davis AS, Webster JD, Hoover S, Simpson RM, Gauldie J. Metastatic growth from dormant cells induced by a Col-I–Enriched fibrotic environmentmetastatic outgrowth from dormant tumor cells. Can Res. 2010;70(14):5706–16.
Fakih MG. Metastatic colorectal cancer: current state and future directions. J Clin Oncol. 2015;33(16):1809–24.
Rahbari NN, Kedrin D, Incio J, Liu H, Ho WW, Nia HT, Edrich CM, Jung K, Daubriac J, Chen I. Anti-VEGF therapy induces ECM remodeling and mechanical barriers to therapy in colorectal cancer liver metastases. Sci Transl Med. 2016;8(360):360ra135-360ra135.
Evans T, Ramanathan R, Yazji S, Glynne-Jones R, Anthoney A, Berlin J, Valle J. Final results from cohort 1 of a phase II study of volociximab, an anti-α5β1 integrin antibody, in combination with gemcitabine (GEM) in patients (pts) with metastatic pancreatic cancer (MPC). J Clin Oncol. 2007;25(18_Suppl):4549–4549.
Haddad T, Qin R, Lupu R, Satele D, Eadens M, Goetz MP, Erlichman C, Molina J. A phase I study of cilengitide and paclitaxel in patients with advanced solid tumors. Cancer Chemother Pharmacol. 2017;79:1221–7.
Zhang L, Zhao D, Wang Y, Zhang W, Zhang J, Fan J, Zhan Q, Chen J. Focal adhesion kinase (FAK) inhibitor-defactinib suppresses the malignant progression of human esophageal squamous cell carcinoma (ESCC) cells via effective blockade of PI3K/AKT axis and downstream molecular network. Mol Carcinog. 2021;60(2):113–24.
Totaro A, Zhuang Q, Panciera T, Battilana G, Azzolin L, Brumana G, Gandin A, Brusatin G, Cordenonsi M, Piccolo S. Cell phenotypic plasticity requires autophagic flux driven by YAP/TAZ mechanotransduction. Proc Natl Acad Sci. 2019;116(36):17848–57.
Liu Q, Xia H, Zhou S, Tang Q, Zhou J, Ren M, Bi F. Simvastatin inhibits the malignant behaviors of gastric cancer cells by simultaneously suppressing YAP and β-catenin signaling. Onco Targets Ther. 2020;13:2057.
Giraud J, Molina-Castro S, Seeneevassen L, Sifré E, Izotte J, Tiffon C, Staedel C, Boeuf H, Fernandez S, Barthelemy P. Verteporfin targeting YAP1/TAZ-TEAD transcriptional activity inhibits the tumorigenic properties of gastric cancer stem cells. Int J Cancer. 2020;146(8):2255–67.
Jiao S, Wang H, Shi Z, Dong A, Zhang W, Song X, He F, Wang Y, Zhang Z, Wang W. A peptide mimicking VGLL4 function acts as a YAP antagonist therapy against gastric cancer. Cancer Cell. 2014;25(2):166–80.
Li J, Li Z, Wu Y, Wang Y, Wang D, Zhang W, Yuan H, Ye J, Song X, Yang J. The Hippo effector TAZ promotes cancer stemness by transcriptional activation of SOX2 in head neck squamous cell carcinoma. Cell Death Dis. 2019;10(8):1–15.
Liu X, Ye Y, Zhu L, Xiao X, Zhou B, Gu Y, Si H, Liang H, Liu M, Li J. Niche stiffness sustains cancer stemness via TAZ and NANOG phase separation. Nat Commun. 2023;14(1):238.
Bhat KP, Salazar KL, Balasubramaniyan V, Wani K, Heathcock L, Hollingsworth F, James JD, Gumin J, Diefes KL, Kim SH. The transcriptional coactivator TAZ regulates mesenchymal differentiation in malignant glioma. Genes Dev. 2011;25(24):2594–609.
Yang C, Tibbitt MW, Basta L, Anseth KS. Mechanical memory and dosing influence stem cell fate. Nat Mater. 2014;13(6):645–52.
Liu L, Zhang SX, Liao W, Farhoodi HP, Wong CW, Chen CC, Ségaliny AI, Chacko JV, Nguyen LP, Lu M. Mechanoresponsive stem cells to target cancer metastases through biophysical cues. Sci Transl Med. 2017;9(400):eaan2966.
Reagan MR, Kaplan DL. Concise review: mesenchymal stem cell tumor-homing: detection methods in disease model systems. Stem cells. 2011;29(6):920–7.
Kalli M, Stylianopoulos T. Defining the role of solid stress and matrix stiffness in cancer cell proliferation and metastasis. Front Oncol. 2018;8:55.
Acknowledgements
The authors would like to thank the Iran University of Medical Sciences for supporting this research.
Funding
This work was supported by a grant from Iran University of Medical Sciences (No: 96-04-87-32781) and Iran National Science Foundation (INSF) (No: 96015369).
Author information
Authors and Affiliations
Contributions
SS: conceptualization, writing—original draft preparation, designing figures and preparing tables. RS: Writing—original draft preparation. AS: critically revised the manuscript, improve English writing. SD: designing figures and preparing tables. LSZ: writing—reviewing and editing. MDM: writing—reviewing and editing, validation. ZM: conceptualization, writing—reviewing and editing, supervision. RG: conceptualization, writing—original draft preparation, supervision. The author(s) read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Safaei, S., Sajed, R., Shariftabrizi, A. et al. Tumor matrix stiffness provides fertile soil for cancer stem cells. Cancer Cell Int 23, 143 (2023). https://doi.org/10.1186/s12935-023-02992-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-023-02992-w