
Abstract
Background and purpose
Dysregulated miRNAs play an important role in many malignant tumors. However, elucidating the roles of miRNAs in cancer biology, especially in epithelial cancers, remains an ongoing process. In this study, we identified the differentially expressed miR-645 in the progressing of head and neck squamous cell carcinoma (HNSCC) and investigated its biological function.
Methods
The association between clinicopathological parameters and the expression levels of the candidated miRNAs were analyzed by using the Kaplan–Meier survival analysis. The cell growth, invasion and migration potential, and clone formation were observed to detect the functions of the miRNAs in HNSCC cells.
Results
In the 34 HNSCC tissues with lymph node metastasis, the expression level of miR-645 was 0.54 ± 0.12, and the expression level was 0.22 ± 0.05 in the 28 tissues with non lymph node metastasis (p = 0.017). In patients with HNSCC, higher level of miR-645 expression significantly correlates with worse overall survival (p = 0.04). Ectopic expression of miR-645 promoted cell invasion and migration.
Conclusions
miR-645 play a key role in cell invasion and metastasis and their expression correlates with overall survival in the patients with HNSCC.
Similar content being viewed by others


Background
MicroRNAs (miRNAs) are endogenous RNAs that play important gene-regulatory roles in animals via sequence-specific interactions with the 3′UTR of cognate mRNA targets, causing suppression of translation and mRNA decay [1, 2]. Nucleotides 2–7, from the 5′ end of the miRNAs, are referred to as the “seed” and are critical for hybridization to the targets [3], It has been firmly established that miRNAs regulate many key cellular processes such as cell growth, differentiation and apoptosis [4, 5]. Subsets of miRNAs have been identified as potential diagnostic and prognostic markers in malignant tumors [6–8]. Many evidences suggest that the regulatory capacity of miRNAs is dysregulated and exploited in malignant tumors [9]. Several miRNAs are up-regulated in specific tumors appears a general trait of human cancers which playing a causal role in the transformed phenotype [10–12], and the mechanisms are remain to be known further. Although the number of verified human miRNAs is still expanding, the functions of only a few of them have been described.
Head and neck squamous cell carcinoma (HNSCC) ranks sixth among cancers worldwide [13], includes tumors of the oral cavity, oropharynx, and larynx. Survival rates for HNSCC have remained unchanged throughout the last three decades, and half of all cases die within 5 years of diagnosis [11]. The presence of lymph node metastasis affects more than 50 % of HNSCC patients and it is one of the most important prognostic indicators associated with poor long survival rates [14, 15]. There are some biomarkers for the classification diagnosis, individual treatment and prognosis of HNSCC investigated by miRNAs array and studies have shown altered miRNAs profiles in HNSCC compared to their normal tissue [16–21]. miR-645 has been reported to be disrupt-expression in pathological states [22–25], however, the role of miR-645 in metastasis of cancer has not been reported.
In this study, we examined the expression of miR-645 in HNSCC samples by gene chips and further confirmed in HNSCC samples and HNSCC cell lines using real-time PCR. We found that miR-645 levels were up-regulated in HNSCC tissues and highly invasive cell lines. Furthermore, we have investigated the mechanism of miR-645 in HNSCC cancer cell lines. These results show that exogenous overexpression of miR-645 promotes the invasion and migration of HNSCC cells in vitro.
Methods
Cell culture
The human HNSCC cell lines, HN4 and HN12 were kindly provided by Shanghai Key Laboratory of Stomatology [26–30]. These cell lines were cultured in DMEM supplemented with 10 % heat-inactivated FBS (GIBCO BRL, NY, USA), penicillin (100 units/ml) and streptomycin (100 μg/ml) at 37 °C in a humidified 5 % CO2 atmosphere.
Tissue samples and reagents
Tissue samples from patients undergoing curative treatment for definitely diagnosed HNSCC were obtained by surgery, with half of each sample quickly frozen in liquid nitrogen and stored at −80 °C until use and the other half embedded in paraffin for pathological examination. All patients selected in the study were informed consent in advance. In parallel, a separate cohort of 47 patients also was assembled from a large pool of patients in the database based on histologic diagnosis of HNSCC who had undergone radical surgery. We retrospectively reviewed the medical records of patients with HNSCC. In this study, we retrospectively reviewed the medical records of patients. Total RNAs were extracted from paraffin blocks using the high pure miRNA isolation kit according to the manufacturer’s protocol (Roche, Switzerland) before further analysis.
Both the miR-645 inhibitor and its mimics were purchased from GenePharma (Shanghai, China). The high pure miRNA isolation kit was purchased from Roche (Basel, Switzerland). The miRcute miRNA qPCR detection kit and miRcute miRNA qPCR detection kit were purchased from TIANGEN BIOTECH (Beijing, China).
Invasion assays and wound-healing experiment
In vitro invasion assays were performed to analyze the invasive potential. A total of 8 × 104 various cells in 200 μl serum-free DMEM medium were plated onto BD BioCoat™ Matrigel™ Invasion Chamber (8 μm pore size; BD Biosciences) and the lower chamber was immediately filled with 500 μl of DMEM medium with 10 % FBS as a chemoattractant. After 24 h of incubation in a humidified atmosphere containing 5 % CO2 at 37 °C, the non-invading cells are removed from the upper surface of the membrane by a cotton swab and the membranes were then fixed with methanol and stained by 0.2 % crystal violet. For wound-healing experiments, cells were plated in 6-well plates, transfected as indicated, and cultured to confluency. Cells were serum-starved and scraped with a P200 tip (time 0), and the number of migrating cells is counted from pictures (5 fields) taken at the indicated time points.
Clony formation
Twenty-four hours after transfection, HNSCC cells (1 × 105 cells per plate) were plated in 100-mm culture dishes and incubated with 600 μg/ml G418 in final concentration for 14 days to allow colonies formation. The colonies were then washed twice with PBS, fixed with 70 % ethanol and stained with Coomassie Blue. Colonies of more than 50 cells were counted under a dissecting microscope. The data from colony formation were showed as mean ± SD from at least three independent experiments, each being performed in triplicate.
Statistical analysis
Statistical analyses for real-time PCR and the in vitro analysis were performed with software from SPSS 13.0 (standard version 13.0; SPSS Inc., Chicago, IL, USA). The results of the cell proliferation assay, colony formation assay, and in vitro invasion assay were evaluated by Student’s t tests. Patients were divided in two groups based on the median of the miR-645 expression values. Tumors were then classified as high miR-645 group if the expression value was equal to or above the median and as low miR-645 group if the expression value was below the median. The correlation between miR-645 expression and the disease-free survival probability were estimated by using Kaplan–Meier survival analysis. A P < 0.05 was taken as the level of significance.
Results
The association of miR-645 expression with metastatic rates in patients with HNSCC
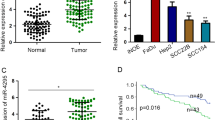
We first measured mature miR-645 levels in a group of tissue specimens from the HNSCC patients. In the 76 HNSCC tissues with lymph node metastasis, the expression level of miR-645 was 2.71 ± 0.24, and the expression level was 1.58 ± 0.23 in the 51 tissues with non lymph node metastasis (p = 0.001, Fig. 1a). The results showed that the miR-645 expression level in the primary HNSCC samples with lymph node metastasis was significantly higher than that in the tissues without lymph node metastasis. The correlations between the miR-645 expression level and clinical pathological characteristics of are summarized in Table 1. Statistically significant associations between the miR-645 expression levels and metastatic rates were observed. In the HNSCC tissues with severe histological signs (vascular emboli, perineural invasion, diffuse infiltration), the expression level of miR-645 was also significantly higher than the expression level in the tissues with non-severe histological signs (Fig. 1c–e). However, there was no significant correlation between the expression level of miR-645 and age, sex, tumor size, site, smoking history, alcohol history (Table 1).

Clinical association of miR-645 with HNSCC patients. a Mature miR-645 levels were measured in HNSCC samples by real-time PCR. b Kaplan–Meier graph representing the probability of disease-free survival in HNSCC patients from the “Milan-INT” dataset stratified. The log-rank test P value reflects the significance of the association between low miR-645 level and disease-free survival. c–e Mature miR-645 levels were compared between HNSCC samples with severe histological signs (vascular emboli, perineural invasion, diffuse infiltration) and the tissues with non-severe histological signs by real-time PCR
We next measured mature miR-645 levels in a collection of HNSCC patients with clinical history. Patients were divided in two groups, with, respectively high or low levels of miR-645. Remarkably, when tested using the Kaplan–Meier survival analysis, the miR-645 “low” group displayed a significant longer disease-free survival when compared to the “high” group (Fig. 1b). These data suggested a possible link between miR-645 expression and tumor progression.
miR-645 promotes cell proliferation
HN4 and HN12 cell lines were established from primary HNSCC tissue and the lymph node metastatic tissue from the same patient, respectively. We used these cell lines to investigate how gain or loss of function of miR-645 impacted cell biological behavior. The HN12 cells displayed high migration capacities and contained a relatively high level of miR-645 (Fig. 2a). First, we assessed the growth of miR-645-transfected and miR-NC-transfected HN4 cells after transient transfection. As shown in Fig. 2b, miR-645 was able to increase the proliferation of miR-645-transfected cells compared with miR-NC-transfected cells significantly at day 3 and 5 (P < 0.05, Student’s t test) (Fig. 2b). We further tested if endogenous expression of miR-645 was required for HNSCC invasion in the cancer cell line HN12. For this purpose, we silenced miR-645 and this treatment led to an approximately 1.5-fold decline in growth properties (Fig. 2c).

miR-645 promotes HNSCC. a Expression levels of miR-645 in cellular models of metastatic progression were tested by real-time PCR. Values related to the nonmetastatic, less aggressive cell line (HN4) are normalised to U6 and shown as the mean and SD. b, c Cell growth curves: proliferation of phenotypically stable indicated cell lines was monitored by the CCK-8 assay. d Linear regression analyse the correlation between miR-645 and IFIT2
Ectopic expression of miR-645 promotes cancer metastasis
In light of the preceding data, we aimed to determine more directly if miR-645 plays a causal role in the aggressive traits of HNSCC cancer cells. We used this cell line to investigate how gain or loss of function of miR-645 impacted cell migration and invasion, which are hallmarks of metastatic capacity. In the transwell assays shown in Fig. 3b, down regulation of miR-645 in HN12 cells decreased invasive abilities 1.6-fold compared to the same cells expressing miR-NC. We further tested if endogenous expression of miR-645 was required for cell invasion in HNSCC cancer cell line HN4. For this purpose, we upregulated miR-645 and this treatment led to an approximately 1.8-fold augmentation in invasive properties (Fig. 3a). Furthermore, the pro-migration effects of miR-645 were observed in wound-healing assays in HN4 and HN12 cells (Fig. 3c, d).

miR-645 promotes cell invasion and migration. a, b Representative pictures of cells migrated through the filter, stained with crystal violet, and taken at the same magnification and absolute quantifications as cells that had invaded through the transwell. c, d Wound-healing assay showing that gain of miR-645 promotes cell migration and loss of miR-645 suppresses cell migration
Ectopic expression of miR-645 promotes single cell clone proliferation
Single cell clone proliferation ability is considered to be hallmarks of metastatic capacity. We aimed to determine more directly if miR-645 improve the single cell clone proliferation ability. We used this cell line to investigate how gain or loss of function of miR-645 impacted the single cell clone proliferation ability. In the clony formation shown in Fig. 4a, up-regulation of miR-645 in HN4 cells increased single cell clone proliferation ability 1.5-fold compared to the same cells expressing miR-NC. We further tested if endogenous expression of miR-645 was required for cell invasion in HNSCC cancer cell line HN12. For this purpose, we down-regulated miR-645 and this treatment led to an approximately 1.6-fold reduction in single cell clone proliferation properties (Fig. 4b).

miR-645 promotes single cell clone proliferation. a, b Representative pictures of single cell clone proliferation, stained with crystal violet, colony formation assay in the group of 1 × 105 cells for miR-NC transfected and miR-645-transfected cells
Discussion
Although a global reduction of miRNA abundance appears to be a general trait of human cancers, playing a causal role in the metastatic phenotype [11, 12, 31], several miRNAs are up-regulated in tumors [10], recently, miRNAs have been shown to be related to tumor metastasis [30], providing a new perspective on the metastatic process. Nonetheless, The role of miRNAs in HNSCC has been widely investigated. Here, we will focus on miRNA-645 promotes HNSCC cancer metastasis.
In this study, we described for the first time the miR-645 is markly upregulated in metastatic HNSCC in large samples. We also found that miR-645 is closely related to the cancer metastasis and miR-645 “high expression” group displayed a significantly poorer disease-free survival rates. The miR-645 promote the HNSCC cells proliferation, invasion, migration and the single cell clone proliferation ability. The properties is the malignant characters of oncogenes. We have proven that the down-regulation of miR-645 is crucial in HNSCC metastasis and demonstrated that miR-645 acts as a putative oncogene.
Recently, miRNAs have been proved to be related with tumor metastasis [32–34], providing a new perspective on the metastatic process. Nonetheless, the role of miRNAs in HNSCC metastasis is little known. This study first proves that miR-645 is up-regulated in metastatic HNSCC. Aberrant patterns of miRNA expression are implicated in human diseases including HNSCC. miR-645 plays a variety of important functions in physiology and pathology [22, 24]. Recent literature reports that the miR-645 play important effort in the malignant progressing of ovarian cancer and adenocarcinoma of gastric esophagea [23, 25]. As miRNAs function mainly through the inhibition of multiple target genes and study report that miR-645 inhibits apoptosis by targeting tumor suppressor IFIT2 [25]. In our study, the inverse correlation between miR-645 and IFIT2 was confirmed by real-time PCR in the panel of 127 HNSCC tissues. We support that the IFIT2 may be the target gene of miR-645 (Fig. 2d). Meantime we support that the miRNAs play their efforts though the complicate gene net. This result may preliminary explain the function of miR-645, but the mechanism is still to be research.
Conclusion
In conclusion, our results have proven that miR-645 plays a causal role in the metastases of HNSCC. These findings have implications for understanding the mechanism of HNSCC, and miR-645 may be a valuable maker and target for prevention or adjuvant therapy in HNSCC.
References
Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–33.
Ambros V. The functions of animal microRNAs. Nature. 2004;431(7006):350–5.
Guo H, Ingolia NT, Weissman JS, Bartel DP. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature. 2010;466(7308):835–40.
Schickel R, Boyerinas B, Park SM, Peter ME. MicroRNAs: key players in the immune system, differentiation, tumorigenesis and cell death. Oncogene. 2008;27(45):5959–74.
Zhang J, Guo H, Qian G, Ge S, Ji H, Hu X, Chen W. MiR-145, a new regulator of the DNA fragmentation factor-45 (DFF45)-mediated apoptotic network. Mol Cancer. 2010;9:211.
Kent OA, Mendell JT. A small piece in the cancer puzzle: microRNAs as tumor suppressors and oncogenes. Oncogene. 2006;25(46):6188–96.
Calin GA, Croce CM. MicroRNA signatures in human cancers. Nat Rev Cancer. 2006;6(11):857–66.
Hou J, Lin L, Zhou W, Wang Z, Ding G, Dong Q, Qin L, Wu X, Zheng Y, Yang Y, et al. Identification of miRNomes in human liver and hepatocellular carcinoma reveals miR-199a/b-3p as therapeutic target for hepatocellular carcinoma. Cancer Cell. 2011;19(2):232–43.
Ventura A, Jacks T. MicroRNAs and cancer: short RNAs go a long way. Cell. 2009;136(4):586–91.
Volinia S, Calin GA, Liu CG, Ambs S, Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc Natl Acad Sci USA. 2006;103(7):2257–61.
Kumar MS, Lu J, Mercer KL, Golub TR, Jacks T. Impaired microRNA processing enhances cellular transformation and tumorigenesis. Nat Genet. 2007;39(5):673–7.
Ozen M, Creighton CJ, Ozdemir M, Ittmann M. Widespread deregulation of microRNA expression in human prostate cancer. Oncogene. 2008;27(12):1788–93.
Tran N, Rose BR, O’Brien CJ. Role of human papillomavirus in the etiology of head and neck cancer. Head Neck. 2007;29(1):64–70.
Ferlito A, Rinaldo A, Devaney KO, MacLennan K, Myers JN, Petruzzelli GJ, Shaha AR, Genden EM, Johnson JT, de Carvalho MB, et al. Prognostic significance of microscopic and macroscopic extracapsular spread from metastatic tumor in the cervical lymph nodes. Oral Oncol. 2002;38(8):747–51.
Kowalski LP, Sanabria A. Elective neck dissection in oral carcinoma: a critical review of the evidence. Acta Otorhinolaryngol Italica Organo Ufficiale Della Societa Italiana di Otorinolaringologia e Chirurgia Cervico-Facciale. 2007;27(3):113–7.
Ramdas L, Giri U, Ashorn CL, Coombes KR, El-Naggar A, Ang KK, Story MD. miRNA expression profiles in head and neck squamous cell carcinoma and adjacent normal tissue. Head Neck. 2009;31(5):642–54.
Wong TS, Liu XB, Chung-Wai Ho A, Po-Wing Yuen A, Wai-Man Ng R, Ignace Wei W. Identification of pyruvate kinase type M2 as potential oncoprotein in squamous cell carcinoma of tongue through microRNA profiling. Int J Cancer. 2008;123(2):251–7.
Childs G, Fazzari M, Kung G, Kawachi N, Brandwein-Gensler M, McLemore M, Chen Q, Burk RD, Smith RV, Prystowsky MB, et al. Low-level expression of microRNAs let-7d and miR-205 are prognostic markers of head and neck squamous cell carcinoma. Am J Pathol. 2009;174(3):736–45.
Hui AB, Lenarduzzi M, Krushel T, Waldron L, Pintilie M, Shi W, Perez-Ordonez B, Jurisica I, O’Sullivan B, Waldron J, et al. Comprehensive MicroRNA profiling for head and neck squamous cell carcinomas. Clin Cancer Res Off J Am Assoc Cancer Res. 2010;16(4):1129–39.
Chang SS, Jiang WW, Smith I, Poeta LM, Begum S, Glazer C, Shan S, Westra W, Sidransky D, Califano JA. MicroRNA alterations in head and neck squamous cell carcinoma. Int J Cancer. 2008;123(12):2791–7.
Avissar M, Christensen BC, Kelsey KT, Marsit CJ. MicroRNA expression ratio is predictive of head and neck squamous cell carcinoma. Clin Cancer Res. 2009;15(8):2850–5.
Zhong Y, Pei YH, Wang J, Chen J, Jiang SS, Gong JB. MicroRNA expression profile in myocardial bridging patients. Scand J Clin Lab Invest. 2014;74(7):582–7.
Shih KK, Qin LX, Tanner EJ, Zhou Q, Bisogna M, Dao F, Olvera N, Viale A, Barakat RR, Levine DA. A microRNA survival signature (MiSS) for advanced ovarian cancer. Gynecol Oncol. 2011;121(3):444–50.
Rosenbluth EM, Shelton DN, Wells LM, Sparks AE, Van Voorhis BJ. Human embryos secrete microRNAs into culture media—a potential biomarker for implantation. Fertil Steril. 2014;101(5):1493–500.
Feng X, Wang Y, Ma Z, Yang R, Liang S, Zhang M, Song S, Li S, Liu G, Fan D, et al. MicroRNA-645, up-regulated in human adencarcinoma of gastric esophageal junction, inhibits apoptosis by targeting tumor suppressor IFIT2. BMC Cancer. 2014;14:633.
Miyazaki H, Patel V, Wang H, Ensley JF, Gutkind JS, Yeudall WA. Growth factor-sensitive molecular targets identified in primary and metastatic head and neck squamous cell carcinoma using microarray analysis. Oral Oncol. 2006;42(3):240–56.
Yeudall WA, Crawford RY, Ensley JF, Robbins KC. MTS1/CDK4I is altered in cell lines derived from primary and metastatic oral squamous cell carcinoma. Carcinogenesis. 1994;15(12):2683–6.
Benke EM, Ji Y, Patel V, Wang H, Miyazaki H, Yeudall WA. VEGF-C contributes to head and neck squamous cell carcinoma growth and motility. Oral Oncol. 2010;46(4):e19–24.
Wang H, Patel V, Miyazaki H, Gutkind JS, Yeudall WA. Role for EPS8 in squamous carcinogenesis. Carcinogenesis. 2009;30(1):165–74.
Sun Q, Zhang J, Cao W, Wang X, Xu Q, Yan M, Wu X, Chen W. Dysregulated miR-363 affects head and neck cancer invasion and metastasis by targeting podoplanin. Int J Biochem Cell Biol. 2013;45(3):513–20.
Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA, et al. MicroRNA expression profiles classify human cancers. Nature. 2005;435(7043):834–8.
Shoushtari AN, Szmulewitz RZ, Rinker-Schaeffer CW. Metastasis-suppressor genes in clinical practice: lost in translation? Nature Rev Clin Oncol. 2011;8(6):333–42.
Lujambio A, Calin GA, Villanueva A, Ropero S, Sanchez-Cespedes M, Blanco D, Montuenga LM, Rossi S, Nicoloso MS, Faller WJ, et al. A microRNA DNA methylation signature for human cancer metastasis. Proc Natl Acad Sci USA. 2008;105(36):13556–61.
Tavazoie SF, Alarcon C, Oskarsson T, Padua D, Wang Q, Bos PD, Gerald WL, Massague J. Endogenous human microRNAs that suppress breast cancer metastasis. Nature. 2008;451(7175):147–52.
Authors’ contributions
QS, SC, XZ, ZF carried out the molecular studies. QS drafted the manuscript. WC, XL, QS participated in the design of the study. XH, MS performed the statistical analysis. MY, JZ conceived of the study, and participated in its design and coordination. SC helped to draft the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The human HNSCC cell lines, HN4 and HN12 were kindly provided by Shanghai Key Laboratory of Stomatology.
Grant support
This work was supported by Grants of the National Natural Science Foundation of China (NSFC 81402231), the basic and frontier technology research projects by Science and Technology Department of Henan province (142300410315) and the funding from Youth Foundation of The First Affiliated Hospital of Zhengzhou University and the oral and maxillofacial surgery academician workstation of Zhengzhou (152PYSGZ040).
Compliance with ethical guidelines
Competing interests The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Additional information
Qiang Sun and Shuai Chen contributed equally to this work
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Sun, Q., Chen, S., Zhao, X. et al. Dysregulated miR-645 affects the proliferation and invasion of head and neck cancer cell. Cancer Cell Int 15, 87 (2015). https://doi.org/10.1186/s12935-015-0238-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-015-0238-5