
Abstract
R. toruloides is an oleaginous yeast, with diverse metabolic capacities and high tolerance for inhibitory compounds abundant in plant biomass hydrolysates. While R. toruloides grows on several pentose sugars and alcohols, further engineering of the native pathway is required for efficient conversion of biomass-derived sugars to higher value bioproducts. A previous high-throughput study inferred that R. toruloides possesses a non-canonical l-arabinose and d-xylose metabolism proceeding through d-arabitol and d-ribulose. In this study, we present a combination of genetic and metabolite data that refine and extend that model. Chiral separations definitively illustrate that d-arabitol is the enantiomer that accumulates under pentose metabolism. Deletion of putative d-arabitol-2-dehydrogenase (RTO4_9990) results in > 75% conversion of d-xylose to d-arabitol, and is growth-complemented on pentoses by heterologous xylulose kinase expression. Deletion of putative d-ribulose kinase (RTO4_14368) arrests all growth on any pentose tested. Analysis of several pentose dehydrogenase mutants elucidates a complex pathway with multiple enzymes mediating multiple different reactions in differing combinations, from which we also inferred a putative l-ribulose utilization pathway. Our results suggest that we have identified enzymes responsible for the majority of pathway flux, with additional unknown enzymes providing accessory activity at multiple steps. Further biochemical characterization of the enzymes described here will enable a more complete and quantitative understanding of R. toruloides pentose metabolism. These findings add to a growing understanding of the diversity and complexity of microbial pentose metabolism.
Similar content being viewed by others


Introduction
Economically viable biorefineries upgrading lignocellulosic biomass to value-added products using microbial platforms will need to efficiently utilize all four major sugar monomers—d-glucose, d-xylose, d-mannose, and l-arabinose [1]. While d-glucose is the preferred substrate for most organisms, many have little to no capacity to metabolize d-xylose, the second largest component of biomass [2,3,4]. A deeper understanding of pentose catabolism in diverse organisms is required in order to engineer strains with utilization efficiency on par with that of d-glucose.
Rhodosporidium toruloides (Rhodotorula toruloides) is an oleaginous yeast capable of consuming many diverse substrates (including waste office paper, cassava starch, and macroalgae hydrolysate) [5,6,7]. It is able to tolerate and degrade potential inhibitors in biomass hydrolysates and non-traditional carbon streams (including furfural, 5-HMF, vanillin, syringaldehyde, vanillic acid, levulinic acid, and acetic acid) [8, 9], making it a prime candidate for Acetyl-CoA (AcCoA)-derived bioproducts such as lipids and terpenes. However, R. toruloides suffers from appreciably slower d-xylose growth relative to d-glucose [10] and an understudied pentose metabolism.
d-xylose and l-arabinose metabolism occurs primarily via cofactor-dependent oxidoreductase and redox-neutral isomerase pathways in eukaryotes and prokaryotes, respectively [11]. The d-xylose oxidoreductase pathway includes reduction via d-xylose reductase (XR), xylitol oxidation via xylitol dehydrogenase (XDH), and phosphorylation to d-xylulose-5-phosphate (Xu5P) via d-xylulose kinase (XK). Similarly, eukaryotic l-arabinose metabolism begins with l-arabinose reduction via l-arabinose reductase (or XR), l-arabitol oxidation via l-arabitol-4-dehydrogenase (LA4DH), and l-xylulose reduction via l-xylulose reductase (LXR). Figure 1 illustrates the 4 pathways describing catabolism of the major pentose monomers of deconstructed biomass. However, deviations from the norm exist (e.g., the anaerobic fungus Piromyces sp. strain E2 uses d-xylose isomerase (XI) and XK to metabolize d-xylose [12]).

Canonical fungal and bacterial l-arabinose and d-xylose metabolism are additionally overlaid. Included are most known or putative routes for d-arabitol and xylitol interconversion. Cofactor annotations were left generally as NAD(P)H instead of species-specific. Please refer to Fig. 14 for gene-protein-reaction rules inferred from this study. XR broad-substrate-specificity d-xylose reductase, LA4DH l-arabitol-4-dehydrogenase, LA2DH l-arabitol-2-dehydrogenase, LXR l-xylulose reductase, LAI l-arabinose isomerase, LRK l-ribulose kinase, LRPE l-ribulose-5-phosphate 4-epimerase, XI d-xylose isomerase, XDH xylitol dehydrogenase, DA4DH d-arabitol-4-dehydrogenase, DA2DH d-arabitol-2-dehydrogenase, RK d-ribulose kinase, XK d-xylulose kinase, DAK d-arabitol kinase, X5PP xylitol-5-phosphate phosphatase, Ru5P d-ribulose-5-phosphate, Xu5P d-xylulose-5-phosphate, DAPDH d-arabitol-phosphate dehydrogenase, DRPE d-ribulose-5-phosphate 3-epimerase, DAPP d-arabitol phosphate phosphatase, X5PDH xylitol-5-phosphate dehydrogenase, PPP pentose phosphate pathway, d-arabitol-1(5)-P indicates either d-arabitol-1-phosphate or d-arabitol-5-phosphate, depending on the precursor it was converted from, rxns reactions
A recent study of multi-omics analysis and metabolic model curation surmised—with RNAseq, proteomics, and functional genomics—that R. toruloides possesses non-canonical pentose metabolism through d-arabitol, d-ribulose, and d-ribulose-5-phosphate (Ru5P) [13] (Fig. 1). However, R. toruloides has a putative XK (encoded by RTO4_16850), but RNA transcript levels are near the limit of detection and no XK peptides were detected on both pentose and d-glucose [13]. Simultaneously, putative genes RTO4_9990 and RTO4_14368 (encoding d-arabitol-2-dehydrogenase (DA2DH) and d-ribulose kinase (RK), respectively) were strongly expressed at the peptide level and mutants in these genes had strong growth defects on nearly all pentose substrates examined, whereas mutants in XK had no growth defects in any condition tested [13]. Supporting these data, Jagtap et al. reported that R. toruloides excretes d-arabitol with d-xylose as the sole carbon source; however, they postulated that d-arabitol is a dead-end overflow metabolite to maintain redox homeostasis under high d-xylose utilization conditions alongside traditional flux through XK [14]. Under this model, deletion of RTO4_9990 (putative DA2DH) should have a minor effect on d-xylose growth, with eventual complete or near-complete growth recovery, contradicted by Kim et al. [13]. An additional study by Jagtap et al. with metabolomics and transcriptomics on several carbon sources recapitulates and reinforces the findings of Kim et al., showing no XK transcription on d-xylose with a simultaneous upregulation of the putative alternative route via d-arabitol [13, 15]. The authors updated the model in congruence with the findings of Kim et al.; however, no further pathway characterization or verification was performed.
Production of d-arabitol (and other polyols) from d-glucose or glycerol (with Xu5P or Ru5P precursors) is evident in many fungi [16,17,18,19,20,21], or l-arabitol from l-arabinose [22, 23], hinting that other fungi might possess alterations to canonical pentose metabolism. Production of d-arabitol from d-xylose or l-arabinose is less characterized, although some studies exist.
For example, Saha et al. showed that Zygosaccharomyces rouxii is able to produce d-arabitol and/or xylitol from many carbon sources including d-xylose, d-glucose, d-xylulose, d-fructose, and d-mannose, proposing an unverified pentose model accounting for their observations [24, 25]. A previous study of purified extracts of a different Z. rouxii strain fed [5-14C] d-xylulose demonstrated NADH-linked DA4DH reduction of d-xylulose to d-arabitol [26], with no activity on d-arabitol, but some activity on xylitol (XDH activity), suggesting putative DA4DH is irreversible—either intrinsically so or due to thermodynamic constraints of the tested condition. From other work, it was concluded that DA2DH activity is responsible for production of d-arabitol from d-glucose [27]. Unfortunately, d-arabitol production from d-xylose was not assessed, preventing a definitive understanding in Z. rouxii, although production seems likely via simultaneous DA4DH and DA2DH activity. In the fungus Pichia anomala d-arabitol, xylitol, and ribitol is produced solely from d-xylose, in addition to d-arabitol and xylitol solely from d-glucose [28]. Zhang et al. characterized the putative arabitol dehydrogenase in vitro showing reversible DA4DH and irreversible XDH (d-xylulose-forming only) activities, both NADH-dependent, but with highest activity on d-arabitol. However, there was no verification of their theoretical model of P. anomala pentose metabolism, so details of d-xylose metabolism remain unclear. A study of Candida maltosa grown on d-xylose mother liquor produced mostly xylitol, with a small fraction of d-arabitol [29, 30]. The authors then tested for growth solely on d-arabitol without observing xylitol production. They surmised an irreversible reaction of d-xylulose reduction (to d-arabitol) and proposed a potential d-xylose catabolic pathway accounting for d-arabitol production that includes both canonical (XR, XDH, XK) and non-canonical routes (XR, XDH, DA4DH, DA2DH). However, no in-depth characterization of the pathway or responsible genes was carried out to confirm this model. Candida arabinofermentans and Pichia guilliermondii fed 13C-labeled l-arabinose produced both labeled d-arabitol and labeled l-arabitol, as detected via NMR. However, the authors inferred that that d-arabitol was produced from d-ribulose via the PPP as opposed to directly from d-xylulose, and hypothesized that this only serves as a means to regenerate NAD+ during oxygen-limiting conditions rather than a mainstay for pentose metabolism [31, 32]. Additional examples and a more comprehensive survey of arabitol and other polyols produced by various yeasts can be found elsewhere [20, 33,34,35,36,37,38,39].
Taken together, data from different fungi suggest a diversity of polyol metabolism linked to pentose catabolism, though in most cases the data are fragmented with uncertainty as to which polyols are intermediates in the main pathway, and which may be side products. Also, while previous work by Kim et al. [13] provides solid evidence that a non-canonical pathway exists in R. toruloides and identifies several enzymes involved, high-throughput fitness studies are limited in their precision, especially for cases of overlapping enzyme function. In order to engineer R. toruloides for optimal pentose conversion, a more complete picture of the catabolic pathway and the enzymes mediating each step is required. To this end, we systematically probed the functions of each of the major putative R. toruloides pentose genes via genomic deletions and selective complementation by heterologous XI and XK. Growth phenotyping on representative pentose substrates, metabolite profiling, and enantiomer determination of arabitol accumulated in the growth medium were employed to piece together a clear picture of this unusual pentose metabolism and further validate previous high-throughput observations. Lastly, we show that the pentose pathway is functionally redundant at nearly every step and explore an unusual substrate-specific bypass to our proposed pentose metabolic model.
Results
Growth phenotyping of putative pentose catabolic enzyme deletions
As in most eukaryotes, the first step in d-xylose metabolism in R. toruloides is likely via an XR as opposed to an XI [40]. Protzko et al. found a putative l-glyceraldehyde and general pentose reductase (encoded by RTO4_9774) critical for d-galacturonic acid metabolism. In vitro enzyme activity assays showed that substrates of the reductase include l-glyceraldehyde, l-arabinose, and d-xylose, and NADPH. This broad substrate specificity is not unusual amongst fungi [41, 42]. Although the preferred substrate of the reductase is not d-xylose, we still assessed its role in d-xylose metabolism. An RTO4_9774 deletion resulted in diminished, but not abolished, growth on d-xylose as the sole carbon source, indicating XR redundancy (Fig. 2). Alignment of characterized fungal XRs from Aspergillus niger and Trichoderma reesei to R. toruloides suggests many additional candidates for this activity.

Growth of WT vs putative d-xylose reductase deletion complemented with XI on 40 g/L d-xylose medium. Solid lines are the average of 3 biological replicates; shaded regions indicate 100% percentile intervals; OE: overexpression; A.U.: Arbitrary Units
There is no evidence that R. toruloides has a functional XI, allowing us to employ XI in our investigation of the pentose assimilation pathway. To test its functionality, codon-optimized XI (from Lachnoclostridium phytofermentans) was randomly integrated (via Agrobacterium tumefaciens-mediated transformation (see methods)) into ΔRTO4_9774 (i.e., ΔRTO4_9774 + OE XI), driven by a strong promoter (Rhodotorula graminis Tef1; Fig. 2), recovering growth of the deletion mutant. This indicates that d-xylulose (the product of XI) is likely a native metabolite of pentose metabolism, and that the rate-limiting step of pentose metabolism is probably either downstream of d-xylulose (provided sufficient L. phytofermentans XI expression and activity) or at the point of transport into the cell.
Following reduction of d-xylose, xylitol is likely oxidized to d-xylulose via XDH (Fig. 1). RTO4_8988 mutants have considerable fitness defects on multiple carbon sources (l-arabinose, l-arabitol, l-lyxose, xylitol), and modest defects on d-xylose, d-xylulose [13]. Thus, RTO4_8988 likely plays a promiscuous role in l-arabitol and d-xylose metabolism. Orthologous sequences from the filamentous fungi, T. reesei, with empirical data obtained from cell-free extracts and purified enzyme assays include an NADPH-dependent d-mannitol 2-dehydrogenase encoded by lxr1 [43], and an NADPH-dependent LXR encoded by lxr3 [44], the latter showing promiscuous polyol-forming activity on many substrates in addition to l-xylulose—notably d-xylulose [44]. Additionally, characterized LXR1 from A. niger shows weak similarity to only a single gene, RTO4_8988 [45, 46]. Unsurprisingly, ΔRTO4_8988 grown on select pentose substrates shows multiple growth defects in our proposed pentose metabolism model (Fig. 3). Namely, growth defects are observed on d-xylose, xylitol, d-xylulose, l-xylulose, or l-arabitol. The progressively worsening growth defects of RTO4_8988 mutants as the carbon source is moved upstream in the pentose utilization pathway definitively supports RTO4_8988 d-arabitol-4-dehydrogenase (DA4DH) activity and possibly XDH, LXR activities.

Growth curves of WT, ΔRTO4_8988, ΔRTO4_8988 + OE XK on 5 g/L per sugar. Solid lines are the average of 3 biological replicates; shaded regions indicate 100% percentile intervals
Under canonical fungal metabolism, d-xylulose is converted to Xu5P via XK (Fig. 1). Comparative sequence analysis to characterized fungal XKs reveals that R. toruloides RTO4_16850 likely encodes an XK [47, 48]; however, under no conditions tested were RTO4_16850 peptides detected or any fitness defects observed upon deletion via RB-TDNAseq fitness profiling [13]. Either regulatory mechanisms, improper conditions, coding mutations, or cofactor balancing are suppressing expression and activity. To explore the functionality of RTO4_16850, a plasmid driven by a strong native promoter (P14 Tef1; [49]) expressing native RTO4_16850 sequence was randomly integrated into ΔRTO4_9990 (deletion without growth on d-xylose), and screened for growth on d-xylose as the sole carbon source. No growth was observed in any of the 48 transformants.
With strong evidence of no native XK functionality, XK from A. niger was codon optimized, overexpressed on a plasmid driven by the strong Rhodotorula graminis Tef1 promoter, and randomly integrated in ΔRTO4_8988 (i.e., ΔRTO4_8988 + OE XK). Growth on four of five sugars with observed growth deficits in ΔRTO4_8988 was partially or fully recovered upon XK complementation (Fig. 3). Most notable is the vastly improved growth of ΔRTO4_8988 + OE XK relative to WT on d-xylose (Fig. 3A), implying pentose metabolism is possibly rate limited downstream XDH, and RTO4_8988 may encode minor XDH activity. Interestingly, ΔRTO4_8988 + OE XK has an exacerbated growth deficit on xylitol relative to ΔRTO4_8988, potentially due to unbalanced redox homeostasis in this condition. ΔRTO4_8988 + OE XK grown on l-arabitol and l-xylulose shows partial recovery relative to WT, supporting RTO4_8988 LXR activity (Fig. 3D–E). As expected, the d-xylulose growth deficit from loss of RTO4_8988 DA4DH activity is complemented by ΔRTO4_8988 + OE XK (Fig. 3C). Growth of ΔRTO4_8988 + OE XK on d-arabitol is not impacted by XK expression, possibly due to reaction irreversibility of DA4DH, thermodynamic constraints, or cofactor/redox imbalance. We further explored the effects of XI and XK overexpression in ΔRTO4_8988 on d-xylose via random integration with the Rhodotorula graminis Tef1 promoter (Fig. 4). ΔRTO4_8988 + OE XI has an identical growth rate to ΔRTO4_8988; however, once XK is overexpressed with ΔRTO4_8988 + OE XI (Fig. 4), growth surpasses WT (similar to Fig. 3A), indicating XR and XDH are not rate limiting. The lack of XI improving growth is similar to Fig. 2 (ΔRTO4_9774 + OE XI).

WT, ΔRTO4_8988, ΔRTO4_8988 + OE XI + XK, ΔRTO4_8988 + OE XI biomass growth curves on 5 g/L d-xylose. Solid lines are the average of 3 biological replicates; shaded regions indicate 100% percentile intervals
Cross referencing RB-TDNAseq mutant fitness, another putative XDH—RTO4_16452—has moderate fitness defects upon deletion on xylitol and l-arabitol, with elevated expression on d-xylose and l-arabinose [13]. Characterized orthologs from Aspergillus oryzae, Candida tropicalis, Arxula adeninivorans, and A. niger exhibit NAD+-dependent XDH activity [46, 50,51,52,53]. However, when ∆RTO4_16452 was grown on 6 pentose intermediates, a growth defect was only observed on l-arabitol, supporting LA4DH activity (Fig. 5). Various fungi have LA4DHs that function as promiscuous XDHs. For example, in A. oryzae, LA4DH is active on several polyols—ribitol, l-arabitol, xylitol—indicating redundant XDH activity, with la4dh or xdh deletions equally improving xylitol titers [54, 55]. Seiboth et al. showed T. reesei XDH NAD+-dependent activity on xylitol and d-xylulose, but inactivity on l-arabitol and l-arabinose, with its deletion complemented by LA4DH [56]. Notably, RTO4_16452 protein alignment to putative XDH RTO4_8988 shows no similarity, but significant similarity to RTO4_12977 (i.e., 70% coverage and 41% identity)—another putative LA4DH and potential XDH.

WT, ΔRTO4_12977, ΔRTO4_16452 biomass growth curves on 5 g/L per sugar. Solid lines are the average of 3 biological replicates; shaded regions indicate 100% percentile intervals
RTO4_12977 shows significant expression on d-xylose and l-arabinose and, when deleted, moderate fitness defects on l-arabitol, l-lyxose, and l-arabinose [13]. Growth of ΔRTO4_12977 on 6 pentose sugars supports possible minor LA4DH function with no obvious growth detriments on d-xylose, xylitol, or l-xylulose (Fig. 5). Orthologous fungal sequences indicate LA4DH with partial XDH activities (NAD+-linked) [54, 55, 57,58,59,60]. Interestingly, a sequence alignment within the R. toruloides genome shows that RTO4_12977 is strikingly similar to RTO4_12974 (99% coverage and 91% identity); however, ΔRTO4_12974 showed no growth deficits on any of the 6 pentose sugars. Furthermore, RTO4_12974 data from Kim et al. shows no fitness defects upon deletion on any pentose intermediate as well as no transcripts on either d-xylose or l-arabinose [13]. Proteomics data displays an abundance of RTO4_12974 peptides, but due to the very high sequence similarity with RTO4_12977, unique peptides are sparse. Therefore, RTO4_12974 is likely a pseudogene.
In the absence of an endogenous functional XK in R. toruloides, d-xylulose is likely converted to d-arabitol via a DA4DH—a critical step distinguishing canonical vs non-canonical pentose metabolism (Fig. 1). Only two characterized fungal DA4DHs exist, one from the rust fungus, Uromyces fabae [61], but no clear ortholog is evident in the R. toruloides genome. Heterologous expression of ARD1 from U. fabae in Saccharomyces cerevisiae extracts showed NADPH-linked conversion of both d-xylulose and d-ribulose to d-arabitol in isolation (demonstrating DA2DH and DA4DH activities), not in context with global pentose metabolism, so it is unclear how d-xylose metabolism proceeds in U. fabae, although the authors posit d-arabitol is derived from PPP intermediates. Weak potential orthologs consist of ~ 55% coverage and ~ 30% identity relative to U. fabae DA4DH, including RTO4_12977, RTO4_16452, and a pair of similar, putative alcohol/2,3-butanediol dehydrogenases RTO4_9634 and RTO4_13641. The second characterized NADH-dependent DA4DH with additional activity as an XDH is from P. anomala [28]. Alignment to the R. toruloides genome shows ~ 60% coverage and ~ 40% identity to RTO4_9990 and RTO4_8988. RTO4_8905 and RTO4_9837 were two additional putative DA4DHs assigned in Kim et al. [13]; however, single deletions had no growth deficits in any condition tested.
Following d-xylulose reduction, d-arabitol is probably converted to d-ribulose via DA2DH (Fig. 1). In RB-TDNAseq fitness profiling, ΔRTO4_9990 had no growth defect on d-ribulose, but had severe fitness defects on l-arabitol, d-xylose, l-lyxose, l-arabitol, xylitol, d-xylulose, and d-arabitol [13]. This strongly implies RTO4_9990 encodes a DA2DH. In vitro characterizations of orthologs from C. tropicalis, Pichia stipitis, P. anomala, Gluconobacter oxydans, C. albicans, Ambrosiozyma monospora predict mainly an NAD+-dependent DA2DH, followed by possible DA4DH, XDH, and LXR activities [20, 28, 62,63,64,65]. Interestingly, significant protein alignment (93% coverage and 40% identity) of RTO4_9990 only exists with RTO4_8988 and no other R. toruloides genes, highlighting potential functional overlap. We screened ΔRTO4_9990 on 8 pentose substrates and observed mild growth on l-arabitol, l-ribulose, but robust growth on d-ribulose, indicative of DA2DH activity (Figs. 6 and 7). This no-growth phenotype is mirrored in C. albicans, whereby deletion of putative DA2DH eliminates growth on d-arabitol and d-arabinose [20]. Next, we overexpressed A. niger XK in ΔRTO4_9990 and tested for growth complementation on 6 pentose substrates (Fig. 6). Identical or better growth relative to WT was observed on l-xylulose, l-arabitol, and d-xylose, signifying RTO4_9990 does not have major roles upstream d-xylulose. No growth complementation was observed on xylitol, despite full growth recovery on d-xylose, l-arabitol, and l-xylulose, possibly attributed to cofactor or redox imbalance (similar to ΔRTO4_8988 + OE XK; Fig. 3B). Near complete growth recovery is seen on d-xylulose, demonstrating R. toruloides does not have an active native XK under these conditions. No growth complementation was observed on d-arabitol, possibly due to cofactor/redox imbalance, thermodynamic or enzymatic irreversibility—similar to Z. rouxii DA4DH [26]—or RTO4_9990 also encodes DA4DH activity.

Growth curves of WT, ΔRTO4_9990, ΔRTO4_9990 + OE XK on 5 g/L per sugar. Solid lines are the average of 3 biological replicates; shaded regions indicate 100% percentile intervals

Averaged biological triplicate, 240-h end-point OD600 measurements of WT, ΔRTO4_9990, ΔRTO4_14368 grown on 10 g/L per sugar
After d-arabitol oxidation, d-ribulose is most likely converted to Ru5P (Fig. 1) via putative RK encoded by RTO4_14368, orthologous to characterized S. cerevisiae RK [66]. Like RTO4_9990, RTO4_14368 is highly upregulated on pentose sugars l-arabinose and d-xylose, but unlike ΔRTO4_9990, ΔRTO4_14368 exhibits major fitness defects on d-ribulose in addition to other pentoses [13]. We constructed ΔRTO4_14368 and observed growth across 4 pentose substrates (l-ribulose, d-ribulose, l-arabitol, d-arabitol; Fig. 7) in addition to d-xylulose, xylitol, and d-xylose (Fig. 8). ΔRTO4_14368 exhibited no growth on any of the 7 substrates tested, implying that all pentose metabolism proceeds through RK.

Growth curves of WT, ΔRTO4_14368 on 5 g/L per sugar. Solid lines are the average of 3 biological replicates; shaded regions indicate 100% percentile intervals
Metabolite excretion profiles of key pentose mutants
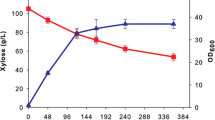
To complement growth phenotyping data, we collected pentose intermediate time-course data from select strains grown on 40 g/L d-xylose with 40 g/L glycerol (Fig. 9). ΔRTO4_8988 is the only strain that accumulates d-xylulose (Fig. 9E), supporting RTO4_8988 DA4DH activity (Fig. 3C). Glycerol consumption is uninhibited, but d-xylose utilization is heavily impacted; however, at peak titers (48 h), nearly 50% of consumed d-xylose is temporarily converted to xylitol, supporting XDH activity (Fig. 9D). In vitro characterization of P. anomala arabitol dehydrogenase (38% identify, 70% coverage to RTO4_8988) shows reversible DA4DH activity and irreversible XDH activity (xylitol to d-xylulose) [28]. After 48 h, though an additional 10 g/L d-xylose are consumed, 4 g/L of xylitol are also consumed. This may reflect increased expression of other enzymes in the pathway over time, shifting the rate-limiting step to d-xylose import or XR activity.

WT, ΔRTO4_9990, ΔRTO4_16452, ΔRTO4_8988 time-series measurements of supernatants grown on 40 g/L d-xylose + 40 g/L glycerol. WT did not produce any detectable d-arabitol or xylitol with glycerol as the sole carbon source. ΔRTO4_9990 is unable to grow on d-xylose (Fig. 6A); hence, all strains were supplemented with glycerol to aid biomass production. Solid lines are the average of 3 biological replicates; shaded regions indicate 100% percentile intervals
ΔRTO4_9990 converted the majority of d-xylose to d-arabitol with a small fraction to xylitol that was eventually consumed (Fig. 9C–D), consistent with RTO4_9990 encoding a DA2DH (Fig. 6). If RTO4_9990 additionally encoded for significant DA4DH activity, we would expect to see an accumulation of d-xylulose (similar to ΔRTO4_8988), but we do not (Fig. 9E). Curiously, ΔRTO4_9990 is able to consume both d-xylose and glycerol more rapidly than WT, hinting that RTO4_9990 itself might be one of the suspected rate-limiting steps in pentose metabolism. Lastly, ΔRTO4_16452 glycerol and d-xylose utilization were similar to WT (Fig. 9A–B); however, there was a modest, but significant temporal decrease and corresponding increase of d-arabitol and xylitol, respectively, relative to WT—supporting RTO4_16452 XDH activity (Fig. 9C–D). No excretion of d-xylulose was observed in ΔRTO4_16452, matching that of WT (Fig. 9E).
Verification of arabitol enantiomer production
In fungi, few studies have definitively verified the actual production pathway of arabitol or distinguished the enantiomer produced [31], which is important for downstream applications [33]. Neither of these two inquiries have been satisfactorily investigated in R. toruloides or related Rhodotorula species; therefore, we performed chiral separations of d-arabitol, l-arabitol standards, and the supernatant from ΔRTO4_9990 grown on glycerol plus d-xylose (Fig. 9C) via gas chromatography-mass spectrometry (GC–MS; see methods). Indeed, Fig. 10 shows that the arabitol isomer produced from d-xylose is d-arabitol.

Derivatized d-arabitol and l-arabitol standards compared to derivatized sample supernatant of end-point culture of ΔRTO4_9990 grown on 40 g/L d-xylose + 40 g/L glycerol (Fig. 9). Solid lines are the average of 3 (biological) replicates; shaded regions indicate 100% percentile intervals
Enzymatic d-arabitol-2-dehydrogenase redundancy
In Fig. 6, ΔRTO4_9990 did not grow on any substrate except for l-arabitol, with slow growth between 40 and 100 h (Fig. 6D), suggesting latent DA2DH redundancy in addition to activity encoded via RTO4_9990. In Fig. 9, a carbon balance between d-xylose, d-xylulose, d-arabitol, and xylitol was approximately closed until mid-run, then approximately 10% of carbon was unaccounted for by the end of the experiment. The onset of this carbon balance gap coincided with the onset of slow growth of ∆RTO4_9990 (Fig. 6D). To test if latent expression of RTO4_16850 was responsible for ∆RTO4_9990’s slow growth on l-arabitol, we constructed a double-deletion strain (i.e., ΔRTO4_9990 ΔRTO4_16850); however, mild growth was still observed on l-arabitol. ΔRTO4_9990 ΔRTO4_16850 was then adapted on 40 g/L l-arabitol (see methods) to generate ΔRTO4_9990 ΔRTO4_16850*. The evolved strain and controls were then cultured on 6 pentose substrates, including l-arabinose (Fig. 11). The evolved strain (ΔRTO4_9990 ΔRTO4_16850*) grew faster than the parent strain on both l-arabitol and l-arabinose, surpassing WT biomass yields with a similar growth rate. However, ΔRTO4_9990 ΔRTO4_16850* is still not able to grow on any other pentose substrate. Further, the evolved strain and controls were grown on 40 g/L l-arabinose, monitoring pentose intermediates (Fig. 12). Over 10 g/L of d-arabitol accumulated in cultures of the evolved strain. This, coupled with xylitol excretion, provides strong evidence of unknown redundant DA2DH activity alongside activity encoded via RTO4_9990. One potential homology-based candidate is RTO4_8988, the only gene that shows any similarity to RTO4_9990 (93% coverage and 40% identity); however, no growth defects were observed for RTO4_8988 mutants on d-arabitol in Fig. 3F. If the late growth on l-arabitol and l-arabinose is occurring through alternative DA2DH function, we would expect to see growth halted if RTO4_14368 is deleted in a ΔRTO4_9990 ΔRTO4_16850 background. Indeed, after 21 days of culturing on 40 g/L l-arabitol, no growth could be measured for the triple-deletion strain (ΔRTO4_9990 ΔRTO4_16850 ΔRTO4_14368), whereas significant growth was measured for ΔRTO4_9990 ΔRTO4_16850 (Fig. 13).

WT, ΔRTO4_9990, ΔRTO4_9990 ΔRTO4_16850, ΔRTO4_9990 ΔRTO4_16850* biomass growth curves on 10 g/L per sugar. Solid lines are the average of 3 biological replicates; shaded regions indicate 100% percentile intervals

WT, ΔRTO4_9990, ΔRTO4_9990 ΔRTO4_16850, ΔRTO4_9990 ΔRTO4_16850* time-series supernatant measurements from cultures grown on 40 g/L l-arabinose. Solid lines are the average of 3 biological replicates; shaded regions indicate 100% percentile intervals

Averaged biological triplicate, end-point OD600 measurements of ΔRTO4_9990 ΔRTO4_16850 and ΔRTO4_9990 ΔRTO4_16850 ΔRTO4_14368 grown on 40 g/L l-arabitol
Discussion
Recent studies of R. toruloides are concentrated on metabolic engineering, especially lipid production from biomass hydrolysates [67,68,69], with less attention to aspects of non-lipid metabolism [13, 15, 40, 49, 70, 71]. More of these studies are needed to identify rate-limiting steps, enzyme redundancy, and cofactor preference of major catabolic pathways to improve productivity, yields, titers, and efficiency of bioproduction. We present here a dataset probing metabolism of l-arabinose and d-xylose (abundant components of renewable biomass) in the non-model fungus R. toruloides which utilizes a non-traditional pathway for pentose metabolism, resulting in xylitol and d-arabitol accumulation from d-xylose.
RNAseq, proteomics, and functional genomics [13], together with d-arabitol accumulation in two R. toruloides strains [14, 72], as well as documented d-arabitol production from non-l-arabinose sources in other yeasts [33], strongly suggest R. toruloides l-arabinose and d-xylose metabolism does not occur via canonical fungal XR, XDH, and XK. Our growth complementation data (Fig. 6) and d-arabitol accumulation data (Figs. 9 and 12) strongly support an alternative pathway through d-arabitol. Moreover, it is an absolute requirement for DA2DH activity encoded via RTO4_9990 to robustly metabolize any pentose substrates. Similarly, expressing a functional RK (encoded via RTO4_14368) is an absolute requirement to metabolize any pentose substrate studied (Figs. 7 and 8)—the only studied enzyme with no known functional redundancy. Lastly, the coordinated transcriptional upregulation of RTO4_14368 and RTO4_9990 strongly suggests simultaneous flux occurs through these steps during d-xylose and l-arabinose metabolism [13]. Figure 14 is an updated pathway reflecting all data collected in this study, accompanied by Table 1, a tabular summary of supporting data for all gene-protein-reaction rules.

Gene-protein-reaction rules of putative l-arabinose and d-xylose metabolism in R. toruloides incorporating all data in this study. (*): Genes with annotation uncertainty due to insufficient evidence (see Table 1); (**): Evidence suggests RK activity is solely encoded by RTO4_14368; Some reactions have multiple enzymes (yellow stars) mediating catalysis, possibly encoded by genes outside the subset depicted here. XR broad-substrate-specificity d-xylose reductase, LA4DH l-arabitol-4-dehydrogenase, LA2DH l-arabitol-2-dehydrogenase, LXR l-xylulose reductase, XDH xylitol dehydrogenase, DA4DH d-arabitol-4-dehydrogenase, DA2DH d-arabitol-2-dehydrogenase, RK d-ribulose kinase, XK d-xylulose kinase, DRPE d-ribulose-5-phosphate 3-epimerase, Xu5P d-xylulose-5-phosphate, PPP pentose phosphate pathway, Ru5P d-ribulose-5-phosphate, rxns reactions, red ‘X’ denotes that R. toruloides does not carry flux via XK under conditions tested, despite possession of an XK-encoding gene, RTO4_16850
Under d-xylose conditions, no evidence exists for expression of a functional R. toruloides XK (RTO4_16850) [13, 15]. This is corroborated by our inability to complement growth of ΔRTO4_9990 with RTO4_16850 expression under a strong endogenous promoter Tef1 [49]. Unexpectedly, XK is highly transcriptionally upregulated in the presence of acetate (whereas the rest of pentose metabolism is downregulated), but the corresponding protein abundance, function, and role XK plays regarding acetate metabolism is unknown [15]. Further investigation into RTO4_16850 functionality (e.g., in vitro kinetics or growth complementation in XK-deficient systems) is warranted. Nonfunctional or dormant pathways are not uncommon. In Y. lipolytica, WT is incapable of utilizing both d-xylose [73], but a study complementing mutant E. coli with genes encoding putative xylose-catabolizing proteins from Y. lipolytica demonstrated functional XR, XDH, and XK [74]. WT overexpression of endogenous XK and XDH resulted in robust growth on d-xylose, without the need for adaptation. Similarly, Y. lipolytica does not consume l-arabinose despite transcriptomics and enzyme activities showing a potential pathway may be active [75]. Culturing an engineered strain (that utilizes d-xylose) on a mixture of d-xylose and l-arabinose, a dramatic improvement in l-arabinose consumption was observed, suggesting l-arabinose catabolism can exist but is dormant due to inhibitory regulation, in addition to rate-limiting LA4DH activity.
Relative to Figs. 1, 14 shows a few alternative/putative pathways to metabolize d-xylose that have limited evidence in fungi, or are not consistent with our data. One example is xylitol conversion to xylitol-5-phosphate (X5P), reported in Streptococcus mutans OMZ 176 [76], followed by X5P dehydrogenase (X5PDH) conversion to Xu5P prior to PPP entrance [77, 78]. Xu5P can be reduced via d-arabitol phosphate dehydrogenase (DAPDH) to d-arabitol-1-phosphate (DA1P) [78, 79], followed with dephosphorylation by broad-specificity polyol phosphate phosphatase (POPP) to d-arabitol, such as one from bacteria [80] or fungi [81]. The S. cerevisiae POPP identified from Xu et al. is active on d-glycerol-3-phosphate, ribitol-5-phosphate, and sorbitol-6-phosphate; however, with POPP active site structural similarity, the authors also speculate catalysis of d-arabitol-5-phosphate (DA5P), X5P, and erythritol-4-phosphate [81]. Analogously, fungi first reduce d-fructose-6-phosphate to mannitol-1-phosphate, then dephosphorylate to mannitol, as opposed to the reverse [82]. Finally, d-arabitol can arise from dephosphorylation of DA5P [20, 77,78,79].
l-arabinose and d-xylose metabolism are alternating series of reduction and oxidation steps, presenting a difficult task of cofactor balancing and redox homeostasis, a possible reason R. toruloides, A. niger [83], and other fungi have functional pentose enzyme redundancy [31]. Typically, different cofactor preference patterns exist for each step amongst molds and yeast [65], with exceptions such as XRs from 3 different yeasts (S. cerevisiae, P. stipitis, Candida parapsilosis) displaying unique cofactor preferences—solely NADPH, both, or mostly NADH, respectively [84,85,86]. Figure 14 shows the predominant cofactors participating in each step of the R. toruloides pathway as predicted by homology to characterized enzymes, but these predictions require experimental validation. Furthermore, in vitro characterization of enzyme kinetics, substrate preference, and cofactor usage might elucidate interesting selective growth of certain substrate-strain combinations tested such as ∆RTO4_8988 + OE XK, ∆RTO4_9990 + OE XK, or ΔRTO4_9990 ΔRTO4_16850* (Figs. 3B, 6B, 11C–F, respectively).
l-ribulose is readily metabolized by WT (Fig. 7) through a pathway yet to be elucidated. This may occur by isomerization to l-arabinose (via l-arabinose isomerase) or conversion to l-ribulose-5-phosphate (via l-ribulose kinase) (Fig. 1). Alternatively, l-ribulose could be converted to l-arabitol via an l-arabitol-2-dehydrogenase (LA2DH; Fig. 1), as demonstrated in purified extracts of the fungus Penicillium chrysogenum [87] and in the bacterium Pantoea ananatis expressing xytf[88, 89], with no clear orthologs in R. toruloides. Regardless of mechanism, ΔRTO4_9990 exhibits slow growth only on l-ribulose (not on d-ribulose), notably with the same final OD600 on l-arabitol (Fig. 7). In contrast, ΔRTO4_14368 neither grows on d-ribulose nor l-ribulose. Together, these data indicate that l-ribulose metabolism is upstream of RTO4_9990, likely part of l-arabinose metabolism (and possibly catalyzed via promiscuous LA2DH activity of unknown origin; Fig. 14). Secondly, this necessitates that d-ribulose is the product of DA2DH, with d-arabitol as the substrate (as there is no known reaction converting l-arabitol to d-ribulose).
Conclusion
We have strongly improved upon and verified results from Kim et al. [13], positing that the primary route of R. toruloides d-xylose and l-arabinose metabolism proceeds through a common intermediate of fungal pentose catabolism—xylitol—followed by oxidation to d-xylulose, reduction to d-arabitol, oxidation to d-ribulose, phosphorylation to Ru5P, and entrance into the PPP. This route is most consistent with evidence of d-arabitol accumulation of ΔRTO4_9990 on d-xylose and the necessity of RTO4_14368 to metabolize any pentose intermediate. This unusual metabolism can be engineered to intricately control sugar and sugar alcohol product profiles of d-ribulose, d-arabitol, xylitol, and d-xylulose, four promising chemicals that can be made from sustainable biomass feedstocks [90]. A continued, multi-faceted approach to understand fundamental metabolism of R. toruloides and related fungi will help accelerate metabolic engineering efforts toward bioproducts by identifying potential rate-limiting steps and genes responsible for encoding enzymatic redundancy of major catabolic pathways.
Methods
Strains and sequences
Rhodosporidium toruloides (a.k.a. Rhodotorula toruloides, a.k.a Rhodotorula graciis) strain IFO 0880 (a.k.a NBRC 0880) was obtained from the Biological Resource Center, NITE (NBRC), Japan. All strains named in this work are available to order through the Agile BioFoundry parts registry at https://public-registry.agilebiofoundry.org. The registry website also hosts all applicable plasmid sequences. Applicable strains and plasmid sequences are listed by figure in Additional file 1. Protein identification numbers used in this manuscript are from the R. toruloides genome version 4, available on Mycocosm, the US Department of Energy Joint Genome Institute fungal genome repository [91]. Selection markers used in R. toruloides were hygromycin, G418, zeocin resistance cassettes using the R. toruloides Tub2 promoter and terminator (see sequences on the Agile BioFoundry parts registry).
For strains constructed by homologous recombination (e.g., full-deletion mutants), the parental strain was wild type. Homologous recombination and non-homologous end-joining (i.e., for generating randomly integrated mutants) was achieved by transforming R. toruloides via TDNA insertion with 1 kbp homology arms to the targeted locus by Agrobacterium tumefaciens-mediated transformation as described in [92]. Strain construction methods are listed for each strain in Additional file 1. For all deletion mutants, successful deletion was confirmed by diagnostic PCR at the altered locus. Plasmids with heterologous gene expression were codon optimized via the high-CAI method (i.e., the most used codons in R. toruloides). For construction of overexpression strains by random insertion, ~ 48 randomly selected transformants were screened for growth in liquid culture with 100 µg/mL of the appropriate selective agent, comparing growth to WT, and selecting the best-performing strain for further analysis.
Media and growth conditions
All chemicals used in this study were from Sigma Aldrich unless otherwise stated. l-xylulose, d-xylulose, l-ribulose, and d-ribulose (XYU-009, XYU-001, RBU-005, and RBU-004, respectively) were purchased from Omicron Biochemicals. For regular strain maintenance and transformation, cells were grown in 10 g/L yeast extract, 20 g/L peptone, 20 g/L glucose (YPD). All strains were first grown on YPD agar plates (15 g/L agar), followed by picking of individual colonies to obtain biological replicates.
For Figs. 2, 3, 4, 5, 6, 7, 8, 9, 11, 12 and 13, cells were subcultured in YPD overnight, washed with water, inoculated at an optical density at 600 nm (OD600) of 0.1 (< 0.001 for Figs. 7 and 13) in 800 µL (5 mL Figs. 4, 12, 13) experimental medium and grown for the allotted time in the Figure (240 h or 21 days for Figs. 7 and 13, respectively) in a long-neck culture tube (Figs. 4, 12, 13) or a microtiter plate format: 48-well M2P Labs Flower Plate MTP-48-B (2000 µL total volume) at 1300 RPM agitation (1000 RPM for Figs. 7 and 9, 200 RPM for Figs. 4, 12, 13), 30 °C, and 85% relative humidity (50% for Figs. 4, 12, 13) in a BioLector Pro (BioLector 1 for Fig. 11) high-throughput microbioreactor (M2P Labs-Beckman Coulter) or in a 3 mm throw shaking incubator (Figs. 7 and 9). The M2P Labs Flower Plate allows for small-scale cultivation at high oxygen transfer rates [93]. For Figs. 6, 7, 11, 12 and 13, the final experimental media contained 1.7 g/L yeast nitrogen base without ammonium sulfate and amino acids (BD 233520), 5 g/L ammonium sulfate, 75 mM KH2PO4, 25 mM K2HPO4, 5 g/L each sugar (Figs. 7 and 11 10 g/L each sugar; 5 g/L d-xylose, 40 g/L l-arabinose, 40 g/L l-arabitol (Figs. 4, 12, 13, respectively)), pH 6.2. For Fig. 2, the experimental medium contained 1.7 g/L yeast nitrogen base without ammonium sulfate and amino acids (BD 233520), 40 g/L d-xylose, 203 mg/L MgCl2*6H2O, 246 mg/L MgSO4*6H2O, 2.5 mM KCl, 10 g/L yeast extract (Y1625), 3.5 mg/L ethylenediaminetetraacetic acid (EDTA), 27.8 mg/L FeSO4*7H2O, 70 mg/L C6H8O7*H2O, pH 5.6. For Figs. 3, 4, 5, 6 and 8, the experimental medium contained 1.7 g/L yeast nitrogen base without ammonium sulfate and amino acids (BD 233520), 5 g/L ammonium sulfate, 5 g/L each sugar (or 5 g/L d-xylose for Fig. 4), 100 µM, FeSO4, 400 µg/L thiamine HCl, 400 µg/L pyridoxine HCl, 180 mM KH2PO4, 20 mM K2HPO4, 176 mg/L nitrilotriacetic acid, 2.5 g/L MgSO4•7H2O, 2 g/L MgCl2•6H2O, 120 mg/L MnSO4•4H2O, 1 mM Na2SO4, 118 mg/L NaCl, 36 mg/L FeSO4•7H2O, 11.8 mg/L CoSO4•7H2O, 11.8 mg/L CaCl2•2H2O, 11.8 mg/L ZnSO4•7H2O, 1.2 mg/L CuSO4•5H2O, 1.2 mg/L AlK(SO4)2•12H2O, 11.8 mg/L H3BO3, Na2MoO4•2H2O, 2.5 mM KCl, pH 5.6. For Fig. 9, experimental medium contained 1.7 g/L yeast nitrogen base without ammonium sulfate and amino acids (BD 233520), 5 g/L ammonium sulfate, 40 g/L d-xylose, 40 g/L glycerol, 0.79 g/L complete supplement mix, 5 g/L yeast extract, 2 g/L MgSO4*7H2O, 100 mM KH2PO4, 10 M KOH-adjusted pH to 5. Periodic samples (40 uL, 250 µL, 250 µL for Figs. 4, 9 and 12, respectively) were aspirated for downstream analysis. Evaporation-corrected samples were taken and final OD600 measurements taken (Figs. 7 and 13).
Adaptive evolution
Generation of the adapted strain ΔRTO4_9990 ΔRTO4_16850* began with a colony of ΔRTO4_9990 ΔRTO4_16850 isolated from plating on YPD agar. A single colony was picked and serially passaged every 3–5 days by transferring 5 µL of cells from the previous culture to a new well of 800 µL fresh media containing 1.7 g/L yeast nitrogen base without ammonium sulfate and amino acids (BD 233520), 5 g/L ammonium sulfate, 75 mM KH2PO4, 25 mM K2HPO4, 40 g/L l-arabitol, pH 6.2. Cells were grown in a microtiter plate format: 48-well M2P Labs Flower Plate (MTP-48-B) at 1000 RPM agitation, 30 °C, and 85% relative humidity in a 3 mm throw shaking incubator. Serial transfers occurred a total of 8 times. The final generation was stopped and plated onto YPD agar. Afterwards, to isolate a pure strain and test for stability of the phenotype, serial plating was completed on YPD agar by choosing a single colony each time, for a total of 5 times. 8 colonies were chosen from the final plating and tested in biological triplicate relative to WT on 40 g/L l-arabitol for improved growth. The most consistently reproducible isolate was chosen for further analysis and named ΔRTO4_9990 ΔRTO4_16850*.
GC–MS analysis
End-point culture samples of ΔRTO4_9990 from Fig. 9 were processed and analyzed as described previously [31]. Briefly, 50 μl of each sample were evaporated to dryness under a stream of nitrogen, dissolved in dichloromethane (200 μl) and trifluoroacetic anhydride (400 μl), heated at 80 °C for 30 min, followed by nitrogen stream drying, and then redissolved in dichloromethane. GC–MS analysis was performed using a Chiraldex G-TA glass capillary column (ASTEC, Sigma 73035AST) and a single quadrupole Agilent GC–MS set at 70 eV. The helium carrier gas flow rate was set at 1 mL/min, and a four-step program was followed: 90 °C for 13 min, 0.8 °C/min up to 110 °C, 4 °C/min up to 180 °C, and 10 min at 180 °C. The injection port and ion source temperatures were maintained at 180 °C. Identification of the sample enantiomer was completed by retention time comparison to pure l-arabitol and d-arabitol standards.
Sugar and sugar alcohol quantification
Sugars were quantified on a Dionex Ultimate 3000 system UHPLC (Agilent Technologies) using an Aminex HPX-87C column (Bio-Rad 1250095) and Thermo Scientific RefractoMax 520 Refractive Index Detector (RID) held at 35 °C. Prior to analysis, samples were diluted to 1:10 and filtered through a 0.45 µM polypropylene membrane microplate filter (Agilent 200983–100) by centrifugation at 3000 RCF for 3 min. Samples were run for 26 min using an isocratic HPLC-grade water mobile phase at 0.6 mL/min and 85 °C. Quantification was completed via peak area measurements compared to standard curves of pure compounds within their linear range of detection.
Availability of data and materials
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request. All strains named in this work are available to order through the Agile BioFoundry parts registry at https://public-registry.agilebiofoundry.org. The registry website also hosts all applicable plasmid sequences.
References
Narisetty V, Cox R, Bommareddy R, Agrawal D, Ahmad E, Pant KK, et al. Valorisation of xylose to renewable fuels and chemicals, an essential step in augmenting the commercial viability of lignocellulosic biorefineries. Sustain Energy Fuels. 2021;6:29–65.
Yang S, Vera JM, Grass J, Savvakis G, Moskvin OV, Yang Y, et al. Complete genome sequence and the expression pattern of plasmids of the model ethanologen Zymomonas mobilis ZM4 and its xylose-utilizing derivatives 8b and 2032. Biotechnol Biofuels. 2018;11:125.
Cunha JT, Soares PO, Romaní A, Thevelein JM, Domingues L. Xylose fermentation efficiency of industrial Saccharomyces cerevisiae yeast with separate or combined xylose reductase/xylitol dehydrogenase and xylose isomerase pathways. Biotechnol Biofuels. 2019;12:20.
Li H, Alper HS. Enabling xylose utilization in Yarrowia lipolytica for lipid production. Biotechnol J. 2016;11:1230–40.
Nora LC, Cassiano MHA, Santana ÍP, Guazzaroni M-E, Silva-Rocha R, da Silva RR. Mining novel cis-regulatory elements from the emergent host Rhodosporidium toruloides using transcriptomic data. Front Microbiol. 2022;13:1069443.
Nair AS, Sivakumar N. Enhanced production of biodiesel by Rhodosporidium toruloides using waste office paper hydrolysate as feedstock: optimization and characterization. Fuel. 2022;327:125174.
Shaigani P, Awad D, Redai V, Fuchs M, Haack M, Mehlmer N, et al. Oleaginous yeasts- substrate preference and lipid productivity: a view on the performance of microbial lipid producers. Microb Cell Fact. 2021;20:220.
Osorio-González CS, Saini R, Hegde K, Brar SK, Lefebvre A, Avalos-Ramírez A. Inhibitor degradation by Rhodosporidium toruloides NRRL 1588 using undetoxified wood hydrolysate as a culture media. Biomass Bioenerg. 2022;160:106419.
Saini R, Hegde K, Brar SK, Vezina P. Advanced biofuel production and road to commercialization: an insight into bioconversion potential of Rhodosporidium sp. Biomass Bioenerg. 2020;132:105439.
Osorio-González CS, Saini R, Hegde K, Brar SK, Lefebvre A, Avalos RA. Crabtree effect on rhodosporidium toruloides using wood hydrolysate as a culture media. Fermentation. 2022;9:11.
Kim D, Woo HM. Deciphering bacterial xylose metabolism and metabolic engineering of industrial microorganisms for use as efficient microbial cell factories. Appl Microbiol Biotechnol. 2018;102:9471–80.
Harhangi HR, Akhmanova AS, Emmens R, van der Drift C, de Laat WTAM, van Dijken JP, et al. Xylose metabolism in the anaerobic fungus Piromyces sp. strain E2 follows the bacterial pathway. Arch Microbiol. 2003;180:134–41.
Kim J, Coradetti ST, Kim Y-M, Gao Y, Yaegashi J, Zucker JD, et al. Multi-omics driven metabolic network reconstruction and analysis of lignocellulosic carbon utilization in Rhodosporidium toruloides. Front Bioeng Biotechnol. 2020;8:612832.
Jagtap SS, Rao CV. Production of D-arabitol from D-xylose by the oleaginous yeast Rhodosporidium toruloides IFO0880. Appl Microbiol Biotechnol. 2018;102:143–51.
Jagtap SS, Deewan A, Liu J-J, Walukiewicz HE, Yun EJ, Jin Y-S, et al. Integrating transcriptomic and metabolomic analysis of the oleaginous yeast Rhodosporidium toruloides IFO0880 during growth under different carbon sources. Appl Microbiol Biotechnol. 2021;105:7411–25.
Bisping B, Baumann U, Simmering R. Effects of immobilization on polyol production by Pichia farinosa. In: Immobilized Cells - Basics and Applications, Proceedings of an International Symposium organized under auspices of The Working Party on Applied Biocatalysis of the European Federation of Biotechnology Noordwijkerhout. Elsevier; 1996. p. 395–401.
Bernard EM, Christiansen KJ, Tsang SF, Kiehn TE, Armstrong D. Rate of arabinitol production by pathogenic yeast species. J Clin Microbiol. 1981;14:189–94.
Yoshikawa J, Habe H, Morita T, Fukuoka T, Imura T, Iwabuchi H, et al. Production of D-arabitol from raw glycerol by Candida quercitrusa. Appl Microbiol Biotechnol. 2014;98:2947–53.
Diamantopoulou P, Papanikolaou S. Biotechnological production of sugar-alcohols: focus on Yarrowia lipolytica and edible/medicinal mushrooms. Process Biochem. 2023;124:113–31.
Wong B, Leeson S, Grindle S, Magee B, Brooks E, Magee PT. D-arabitol metabolism in Candida albicans: construction and analysis of mutants lacking D-arabitol dehydrogenase. J Bacteriol. 1995;177:2971–6.
Egermeier M, Russmayer H, Sauer M, Marx H. Metabolic flexibility of Yarrowia lipolytica growing on glycerol. Front Microbiol. 2017;8:49.
Saha BC, Bothast RJ. Production of L-arabitol from L-arabinose by candida entomaea and Pichia guilliermondii. Appl Microbiol Biotechnol. 1996;45:299–306.
Kordowska-Wiater M, Kuzdraliński A, Czernecki T, Targoński Z, Frąc M, Oszust K. The ability of a novel strain scheffersomyces (Syn. Candida) shehatae isolated from rotten wood to produce arabitol. Pol J Microbiol. 2017;66:335–43.
Saha BC, Sakakibara Y, Cotta MA. Production of D-arabitol by a newly isolated Zygosaccharomyces rouxii. J Ind Microbiol Biotechnol. 2007;34:519–23.
Li X, Zhang Y, Zabed HM, Yun J, Zhang G, Zhao M, et al. High-level production of d-arabitol by Zygosaccharomyces rouxii from glucose: metabolic engineering and process optimization. Bioresour Technol. 2023;367:128251.
Blakley ER, Spencer JFT. Studies on the formation OFD -ARABITOL by osmophilic yeasts. Can J Biochem Physiol. 1962;40:1737–48.
Ingram JM, Wood WA. Enzymatic basis for D-ARBITOL production by Saccharomyces rouxii. J Bacteriol. 1965;89:1186–94.
Zhang G, Lin Y, He P, Li L, Wang Q, Ma Y. Characterization of the sugar alcohol-producing yeast Pichia anomala. J Ind Microbiol Biotechnol. 2014;41:41–8.
Cheng H, Wang B, Lv J, Jiang M, Lin S, Deng Z. Xylitol production from xylose mother liquor: a novel strategy that combines the use of recombinant Bacillus subtilis and Candida maltosa. Microb Cell Fact. 2011;10:5.
Wang H, Li L, Zhang L, An J, Cheng H, Deng Z. Xylitol production from waste xylose mother liquor containing miscellaneous sugars and inhibitors: one-pot biotransformation by Candida tropicalis and recombinant Bacillus subtilis. Microb Cell Fact. 2016;15:82.
Fonseca C, Neves AR, Antunes AMM, Noronha JP, Hahn-Hägerdal B, Santos H, et al. Use of in vivo 13C nuclear magnetic resonance spectroscopy to elucidate L-arabinose metabolism in yeasts. Appl Environ Microbiol. 2008;74:1845–55.
Fonseca C, Romão R, Rodrigues de Sousa H, Hahn-Hägerdal B, Spencer-Martins I. L-Arabinose transport and catabolism in yeast. FEBS J. 2007;274:3589–600.
Kordowska-Wiater M. Production of arabitol by yeasts: current status and future prospects. J Appl Microbiol. 2015;119:303–14.
Ravikumar Y, Razack SA, Ponpandian LN, Zhang G, Yun J, Huang J, et al. Microbial hosts for production of D-arabitol: current state-of-art and future prospects. Trends Food Sci Technol. 2022;120:100–10.
Erian AM, Sauer M. Utilizing yeasts for the conversion of renewable feedstocks to sugar alcohols—a review. Bioresour Technol. 2022;346:126296.
Onishi H, Suzuki T. Microbial production of xylitol from glucose. Appl Microbiol. 1969;18:1031–5.
Lin C-C, Hsieh P-C, Mau J-L, Teng D-F. Construction of an intergeneric fusion from Schizosaccharomyces pombe and Lentinula edodes for xylan degradation and polyol production. Enzyme Microb Technol. 2005;36:107–17.
Quarterman J, Slininger PJ, Kurtzman CP, Thompson SR, Dien BS. A survey of yeast from the Yarrowia clade for lipid production in dilute acid pretreated lignocellulosic biomass hydrolysate. Appl Microbiol Biotechnol. 2017;101:3319–34.
Harcus D, Dignard D, Lépine G, Askew C, Raymond M, Whiteway M, et al. Comparative xylose metabolism among the ascomycetes C. albicans, S. stipitis and S. cerevisiae. PLoS ONE. 2013;8:e80733.
Protzko RJ, Hach CA, Coradetti ST, Hackhofer MA, Magosch S, Thieme N, et al. Genomewide and enzymatic analysis reveals efficient d-galacturonic acid metabolism in the basidiomycete yeast Rhodosporidium toruloides. mSystems. 2019. https://doi.org/10.1128/mSystems.00389-19.
Yoon BH, Jeon WY, Shim WY, Kim JH. L-arabinose pathway engineering for arabitol-free xylitol production in Candida tropicalis. Biotechnol Lett. 2011;33:747–53.
Seiboth B, Metz B. Fungal arabinan and L-arabinose metabolism. Appl Microbiol Biotechnol. 2011;89:1665–73.
Metz B, de Vries RP, Polak S, Seidl V, Seiboth B. The Hypocrea jecorina (syn. Trichoderma reesei) lxr1 gene encodes a D-mannitol dehydrogenase and is not involved in L-arabinose catabolism. FEBS Lett. 2009;583:1309–13.
Metz B, Mojzita D, Herold S, Kubicek CP, Richard P, Seiboth B. A novel L-xylulose reductase essential for L-arabinose catabolism in Trichoderma reesei. Biochemistry. 2013;52:2453–60.
Mojzita D, Vuoristo K, Koivistoinen OM, Penttilä M, Richard P. The, “true” L-xylulose reductase of filamentous fungi identified in Aspergillus niger. FEBS Lett. 2010;584:3540–4.
Witteveen CFB, Weber F, Busink R, Visser J. Isolation and characterization of two xylitol dehydrogenases from Aspergillus niger. Microbiology. 1994;140:1679–85.
vanKuyk PA, de Groot MJ, Ruijter GJ, de Vries RP, Visser J. The Aspergillus niger D-xylulose kinase gene is co-expressed with genes encoding arabinan degrading enzymes, and is essential for growth on D-xylose and L-arabinose. Eur J Biochem. 2001;268:5414–23.
Komeda H, Yamasaki-Yashiki S, Hoshino K, Asano Y. Identification and characterization of D-xylulokinase from the D-xylose-fermenting fungus. Mucor circinelloides FEMS Microbiol Lett. 2014;360:51–61.
Nora LC, Wehrs M, Kim J, Cheng J-F, Tarver A, Simmons BA, et al. A toolset of constitutive promoters for metabolic engineering of Rhodosporidium toruloides. Microb Cell Fact. 2019;18:117.
Ko BS, Kim J, Kim JH. Production of xylitol from D-xylose by a xylitol dehydrogenase gene-disrupted mutant of Candida tropicalis. Appl Environ Microbiol. 2006;72:4207–13.
Böer E, Wartmann T, Schmidt S, Bode R, Gellissen G, Kunze G. Characterization of the AXDH gene and the encoded xylitol dehydrogenase from the dimorphic yeast Arxula adeninivorans. Antonie Van Leeuwenhoek. 2005;87:233–43.
Tran LH, Kitamoto N, Kawai K, Takamizawa K, Suzuki T. Cloning and expression of a NAD+-dependent xylitol dehydrogenase gene (xdhA) of Aspergillus oryzae. J Biosci Bioeng. 2004;97:419–22.
Lima LHA, do Pinheiro Amaral CG, de Moraes LMP, de Freitas SM, Torres FAG. Xylitol dehydrogenase from Candida tropicalis: molecular cloning of the gene and structural analysis of the protein. Appl Microbiol Biotechnol. 2006;73:631–9.
Mahmud A, Hattori K, Hongwen C, Kitamoto N, Suzuki T, Nakamura K, et al. NAD+-dependent xylitol dehydrogenase (xdhA) and L-arabitol-4-dehydrogenase (ladA) deletion mutants of Aspergillus oryzae for improved xylitol production. Biotechnol Lett. 2013;35:769–77.
Suzuki T, Tran LH, Yogo M, Idota O, Kitamoto N, Kawai K, et al. Cloning and expression of NAD+-dependent L-arabinitol 4-dehydrogenase gene (ladA) of Aspergillus oryzae. J Biosci Bioeng. 2005;100:472–4.
Seiboth B, Hartl L, Pail M, Kubicek CP. D-xylose metabolism in Hypocrea jecorina: loss of the xylitol dehydrogenase step can be partially compensated for by lad1-encoded L-arabinitol-4-dehydrogenase. Eukaryot Cell. 2003;2:867–75.
Kim B, Sullivan RP, Zhao H. Cloning, characterization, and engineering of fungal L-arabinitol dehydrogenases. Appl Microbiol Biotechnol. 2010;87:1407–14.
de Vries RP, Flipphi MJ, Witteveen CF, Visser J. Characterization of an Aspergillus nidulans L-arabitol dehydrogenase mutant. FEMS Microbiol Lett. 1994;123:83–90.
Sullivan R, Zhao H. Cloning, characterization, and mutational analysis of a highly active and stable L-arabinitol 4-dehydrogenase from Neurospora crassa. Appl Microbiol Biotechnol. 2007;77:845–52.
Sukpipat W, Komeda H, Prasertsan P, Asano Y. Purification and characterization of xylitol dehydrogenase with l-arabitol dehydrogenase activity from the newly isolated pentose-fermenting yeast Meyerozyma caribbica 5XY2. J Biosci Bioeng. 2017;123:20–7.
Link T, Lohaus G, Heiser I, Mendgen K, Hahn M, Voegele RT. Characterization of a novel NADP(+)-dependent D-arabitol dehydrogenase from the plant pathogen Uromyces fabae. Biochem J. 2005;389(Pt 2):289–95.
Cheng H, Li Z, Jiang N, Deng Z. Cloning, purification and characterization of an NAD-Dependent D-Arabitol dehydrogenase from acetic acid bacterium. Acetobacter suboxydans Protein J. 2009;28:263–72.
Hallborn J, Walfridsson M, Penttilä M, Keränen S, Hahn-Hägerdal B. A short-chain dehydrogenase gene from Pichia stipitis having D-arabinitol dehydrogenase activity. Yeast. 1995;11:839–47.
Murray JS, Wong ML, Miyada CG, Switchenko AC, Goodman TC, Wong B. Isolation, characterization and expression of the gene that encodes D-arabinitol dehydrogenase in Candida tropicalis. Gene. 1995;155:123–8.
Verho R, Putkonen M, Londesborough J, Penttilä M, Richard P. A novel NADH-linked l-xylulose reductase in the l-arabinose catabolic pathway of yeast. J Biol Chem. 2004;279:14746–51.
Singh C, Glaab E, Linster CL. Molecular identification of d-Ribulokinase in budding yeast and mammals. J Biol Chem. 2017;292:1005–28.
Saini R, Osorio-Gonzalez CS, Hegde K, Kaur Brar S, Vezina P. A co-fermentation strategy with wood hydrolysate and crude glycerol to enhance the lipid accumulation in Rhodosporidium toruloides-1588. Bioresour Technol. 2022;364:127821.
Chen N, Xu C, Guo X, Shim H. Effects of sodium and magnesium supplement on lipid production and wastewater treatment by Rhodosporidium toruloides. Renew Energy. 2022;199:919–28.
Osorio-González CS, Saini R, Hegde K, Brar SK, Lefebvre A, Avalos RA. Carbon/nitrogen ratio as a tool to enhance the lipid production in Rhodosporidium toruloides-1588 using C5 and C6 wood hydrolysates. J Clean Prod. 2023;384:135687.
Coradetti ST, Pinel D, Geiselman GM, Ito M, Mondo SJ, Reilly MC, et al. Functional genomics of lipid metabolism in the oleaginous yeast Rhodosporidium toruloides. Elife. 2018. https://doi.org/10.7554/eLife.32110.
Dinh HV, Suthers PF, Chan SHJ, Shen Y, Xiao T, Deewan A, et al. A comprehensive genome-scale model for Rhodosporidium toruloides IFO0880 accounting for functional genomics and phenotypic data. Metab Eng Commun. 2019;9:e00101.
Pinheiro MJ, Bonturi N, Belouah I, Miranda EA, Lahtvee P-J. Xylose metabolism and the effect of oxidative stress on lipid and carotenoid production in rhodotorula toruloides: insights for future biorefinery. Front Bioeng Biotechnol. 2020;8:1008.
Drzymała-Kapinos K, Mirończuk AM, Dobrowolski A. Lipid production from lignocellulosic biomass using an engineered Yarrowia lipolytica strain. Microb Cell Fact. 2022;21:226.
Rodriguez GM, Hussain MS, Gambill L, Gao D, Yaguchi A, Blenner M. Engineering xylose utilization in Yarrowia lipolytica by understanding its cryptic xylose pathway. Biotechnol Biofuels. 2016;9:149.
Ryu S, Trinh CT. Understanding functional roles of native pentose-specific transporters for activating dormant pentose metabolism in Yarrowia lipolytica. Appl Environ Microbiol. 2018. https://doi.org/10.1128/AEM.02146-17.
Assev S, Rölla G. Evidence for presence of a xylitol phosphotransferase system in Streptococcus mutans OMZ 176. Acta Pathol Microbiol Immunol Scand B. 1984;92:89–92.
Hausman SZ, London J. Purification and characterization of ribitol-5-phosphate and xylitol-5-phosphate dehydrogenases from strains of Lactobacillus casei. J Bacteriol. 1987;169:1651–5.
Kentache T, Milohanic E, Cao TN, Mokhtari A, Aké FM, Ma Pham QM, et al. Transport and catabolism of pentitols by listeria monocytogenes. J Mol Microbiol Biotechnol. 2016;26:369–80.
Povelainen M, Eneyskaya EV, Kulminskaya AA, Ivanen DR, Kalkkinen N, Neustroev KN, et al. Biochemical and genetic characterization of a novel enzyme of pentitol metabolism: D-arabitol-phosphate dehydrogenase. Biochem J. 2003;371(Pt 1):191–7.
Kuznetsova E, Proudfoot M, Gonzalez CF, Brown G, Omelchenko MV, Borozan I, et al. Genome-wide analysis of substrate specificities of the Escherichia coli haloacid dehalogenase-like phosphatase family. J Biol Chem. 2006;281:36149–61.
Xu Y-F, Lu W, Chen JC, Johnson SA, Gibney PA, Thomas DG, et al. Discovery and functional characterization of a yeast sugar alcohol phosphatase. ACS Chem Biol. 2018;13:3011–20.
Hult K, Veide A, Gatenbeck S. The distribution of the NADPH regenerating mannitol cycle among fungal species. Arch Microbiol. 1980;128:253–5.
Chroumpi T, Peng M, Aguilar-Pontes MV, Müller A, Wang M, Yan J, et al. Revisiting a “simple” fungal metabolic pathway reveals redundancy, complexity and diversity. Microb Biotechnol. 2021;14:2525–37.
Lee J-K, Koo B-S, Kim S-Y. Cloning and characterization of the xyl1 gene, encoding an NADH-preferring xylose reductase from Candida parapsilosis, and its functional expression in Candida tropicalis. Appl Environ Microbiol. 2003;69:6179–88.
Verduyn C, Van Kleef R, Frank J, Schreuder H, Van Dijken JP, Scheffers WA. Properties of the NAD(P)H-dependent xylose reductase from the xylose-fermenting yeast Pichia stipitis. Biochem J. 1985;226:669–77.
Kuhn A, van Zyl C, van Tonder A, Prior BA. Purification and partial characterization of an aldo-keto reductase from Saccharomyces cerevisiae. Appl Environ Microbiol. 1995;61:1580–5.
Chiang C, Knight SG. L-Arabinose metabolism by cell-free extracts of Penicillium chrysogenum. Biochim Biophys Acta. 1961;46:271–8.
Sakakibara Y, Saha BC. Isolation of an operon involved in xylitol metabolism from a xylitol-utilizing Pantoea ananatis mutant. J Biosci Bioeng. 2008;106:337–44.
Sakakibara Y, Torigoe K. Biochemical characterization of l-arabitol 2-dehydrogenase from Pantoea ananatis. J Biosci Bioeng. 2012;113:715–8.
Werpy T, Petersen G. Top Value Added Chemicals from Biomass: Volume I -- Results of Screening for Potential Candidates from Sugars and Synthesis Gas. US Department of Energy; 2004.
Grigoriev IV, Nikitin R, Haridas S, Kuo A, Ohm R, Otillar R, et al. MycoCosm portal: gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014;42:D699-704.
Zhuang X, Kilian O, Monroe E, Ito M, Tran-Gymfi MB, Liu F, et al. Monoterpene production by the carotenogenic yeast Rhodosporidium toruloides. Microb Cell Fact. 2019;18:54.
Funke M, Diederichs S, Kensy F, Müller C, Büchs J. The baffled microtiter plate: increased oxygen transfer and improved online monitoring in small scale fermentations. Biotechnol Bioeng. 2009;103:1118–28.
Acknowledgements
Sandia National Laboratories is a multi-mission laboratory managed and operated by National Technology and Engineering Solutions of Sandia LLC., a wholly-owned subsidiary of Honeywell International Inc., for the U.S. Department of Energy's National Nuclear Security Administration under contract DE-NA0003525. The views and opinions of the authors expressed herein do not necessarily state or reflect those of the United States Government or any agency thereof. Neither the United States Government nor any agency thereof, nor any of their employees, makes any warranty, expressed or implied, or assumes any legal liability or responsibility for the accuracy, completeness, or usefulness of any information, apparatus, product, or process disclosed or represents that its use would not infringe privately owned rights. The Department of Energy will provide public access to these results of federally sponsored research in accordance with the DOE Public Access Plan (http://energy.gov/downloads/doe-public-access-plan).
Funding
This study was part of the Agile BioFoundry (https://agilebiofoundry.org) supported by the U. S. Department of Energy, Energy Efficiency and Renewable Energy, Bioenergy Technologies Office, through contract DE-AC02-05CH11231 between Lawrence Berkeley National Laboratory and the U.S. Department of Energy. A portion of this research was performed on a project award (https://doi.org/10.46936/reso.proj.2020.51637/60000235) from the Environmental Molecular Sciences Laboratory, a DOE Office of Science User Facility sponsored by the Biological and Environmental Research program under Contract No. DE-AC05-76RL01830.
Author information
Authors and Affiliations
Contributions
PAA planned experiments, constructed strains, wrote the main manuscript text and contributed to data analysis for all figures. STC planned experiments, constructed strains, performed experiments, and contributed to data analysis. JMG supervised all work at Sandia National Laboratories. All authors reviewed and edited the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Strains used in this study with strain IDs and part numbers for relevant plasmids for retrieval on the Agile BioFoundry public registry (https://public-registry.agilebiofoundry.org).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Adamczyk, P.A., Coradetti, S.T. & Gladden, J.M. Non-canonical d-xylose and l-arabinose metabolism via d-arabitol in the oleaginous yeast Rhodosporidium toruloides. Microb Cell Fact 22, 145 (2023). https://doi.org/10.1186/s12934-023-02126-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-023-02126-x