
Abstract
Background
Endolysins are peptidoglycan hydrolases with promising use as environment-friendly antibacterials mainly when used topically. However, in general, endolysin expression is hampered by its low solubility. Thus, a critical point in endolysin industrial production is optimizing their expression, including improvement of solubility and recovery from cell extracts.
Results
We report the expression of two endolysins encoded in the genome of phages infecting Staphylococcus aureus. Expression was optimized through changes in the concentration of the inducer and growth temperature during the expression. Usually, only 30–40% of the total endolysin was recovered in the soluble fraction. Co-expression of molecular chaperones (DnaK, GroEL) or N-term fusion tags endowed with increased solubility (DsbC, Trx, Sumo) failed to improve that yield substantially. Inclusion of osmolytes (NaCl, CaCl2, mannitol, glycine betaine, glycerol and trehalose) or tensioactives (Triton X-100, Tween 20, Nonidet P-40, CHAPS, N-lauroylsarcosine) in the cell disruption system (in the absence of any molecular chaperone) gave meager improvements excepted by N-lauroylsarcosine which increased recovery to 54% of the total endolysin content.
Conclusion
This is the first attempt to systematically analyze methods for increasing yields of recombinant endolysins. We herein show that neither solubility tags nor molecular chaperones co-expression are effective to that end, while induction temperature, (His)6-tag location and lysis buffer additives (e.g. N-lauroylsarcosine), are sensible strategies to obtain higher levels of soluble S. aureus endolysins.
Similar content being viewed by others

Background
The rise in antibiotic resistant bacteria is a threat to human health due to the limited therapeutic options developed over the last years. The efficacy of the few new molecules that have been added to the antibacterial portfolio were, in most cases, quickly compromised due to bacterial resistance that arose and spread quickly. This situation calls for developing novel and radically different methods to counterattack bacterial infections [1, 2]. In this scenario, being outsmarted by microorganisms, the idea of using phages and their lytic enzymes as new defensive antibacterial materials has resurfaced. After a century of being reported for the first time, phages are still at the center of a debate about their utilization. This debate slows down the badly needed use of phage cocktails to treat human and animal infections caused by drug-resistant bacteria. Many opinions favor or oppose the use of phages. However, phage cocktails are already applied for non-human use [3, 4]. A possible way to overcome the stalemate on phage utilization is by supporting the use of endolysins, potent phage-encoded enzymes with peptidoglycan hydrolase activity. Although the chemical structure of peptidoglycan is relatively conserved, subtle changes in their composition allow for the grouping of bacteria according to their chemotypes, reflecting PG slight but important diversity. This explains why endolysins are able to discriminate and destroy the cell envelope of distinct bacterial species. A number of attributes of these peptidoglycan-hydrolases make them attractive candidates for classic antibiotic replacement, among them: narrow lytic spectrum, nanomolar binding affinity, relatively easy manipulation for custom design and unlikely evolution of resistance (since they target essential structures of cell wall). Endolysins are made at the late stage of phage infection and are expressed along with holins, proteins that, by making pores in the cell membrane, allow the endolysins to reach their peptidoglycan (PG) target [5, 6]. Of note, it has been demonstrated that endolysins from phages infecting Gram (+) bacteria are highly effective when added externally [7].
Over the past two decades, these enzymes were proven to have strong antibacterial activity against Gram-positive pathogens when applied extracellularly, and some of them are currently in clinical trials (CF-301, ClinicalTrials.gov, NCT04160468) or already in the biopharmaceutical market (Staphefekt™, https://www.staphefekt.com) [8]. Nevertheless, one of the main hurdles in recombinant production of endolysins is their low recovery in the soluble form. There are a number of reviews addressing endolysin features as alternative antimicrobials [9,10,11]; however, there is little or incomplete information on the reasons for their relatively low recovery from cell extracts at the bench scale. Despite that, at least one commercial use of an endolysin of Gram (+) phage origin has been reported [8], demonstrating that it is feasible to overcome the purification hurdle.
These endolysins have a modular structure consisting of a domain that determines the specific binding to the bacterial cell wall (CBD) and one or two enzymatically active domains (EAD) that can cleave the different bonds present in peptidoglycan. This modular structure gives an experimental frame to propose the design of tailor-made endolysins accommodating EADs with different specificities against one or more Gram (+) bacteria. Simultaneously, it creates a unique opportunity to develop enzymatic antibacterial molecules with improved killing activity [12].
However, a major unsolved problem is the low solubility of most of the reported endolysins [13,14,15], a point that could dissuade the industrial production of these enzymes. So far, there has not been any systematic approach to address the expression of endolysins and their recovery from the bacterial cell extract. A recent report by Zydsiecki et al. described the successful use of matrix assisted refolding of truncated forms of the endolysin LysK [16], a promising alternative worth addressing. Notwithstanding that, the adoption of such approach would require a comprehensive testing on full length endolysins.
A different approach is reported herein, addressing the effect on endolysin recovery of different growth and expression conditions, the co-expression of several molecular chaperones as well as the use of solubility tags fused to the endolysin encoding genes. Finally, the effect of the composition of the buffer for cell disruption, including osmolytes, detergents, and salts, was also analyzed.
Results
Sequence alignment and domain organization of endolysins
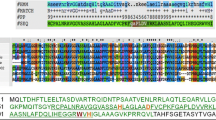
Cg and MatN were identified during the sequencing of bacteriophages active on Staphylococcus aureus isolated in our lab and chosen on the basis of their phylogenetic divergence (Suárez et al., submitted to Microbiology Spectrum, Spectrum00334-22). The amino acid sequences of Cg and MatN do not present remarkable differences when compared to the well characterized endolysins LysK, LysGH15 and LysH5 [14, 17, 18], showing high amino acid identity and similarity through their EADs (CHAP [IPR007921] and Amidase [IPR002502]) and CBD (SH3b [IPR003646]) as well as comparable domain organization (Fig. 1). Furthermore, hydrophobicity plots of these and other endolysins obtained with the Kyte-Doolittle scale showed no predominance of hydrophobic residues in any region of the amino acid sequence (Additional file 1).

MatN and Cg domain architecture and alignment with reference endolysins. Prediction of the domain architecture was analyzed using InterPro. All endolysins are composed of a CHAP (cysteine, histidine-dependent amidohydrolase/peptidase; green), Ami (Amidase; red) and SH3b (blue) domain. Multiple alignment analysis of our endolysins MatN and Cg with endolysins previously reported in the bibliography (LysK, LysGH15 and LysH5) was done using Clustal Omega. Identical residues are shaded in gray and increasing degree of residue conservation correlates with darkening of the gray scale of the bar above the sequences
Effect of inducer concentration and induction temperature on the yield of soluble endolysin
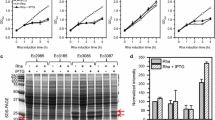
Since the vast majority of the reports on identification of novel endolysins neither provide complete information on the expression yields (Table 1) nor address the reasons behind the low solubility of endolysins, we studied the influence of different factors during the process of induction and lysis. When the effect of the growth temperature during the induction stage was analyzed, we found a clear and reproducible difference in endolysin production on induced cultures kept at low temperature (20 °C) compared to 25 °C and 30 °C (Fig. 2). Cg endolysin induction at 20 °C promoted a significantly higher proportion of soluble endolysin (p = 0.002) when compared to higher induction temperatures.

Effect of Temperature on recombinant endolysins MatN and Cg solubility and yield. A, B E. coli C43(DE3) harboring pET-32a-MatN/Cg was induced at 20, 25 °C, and 30 °C by addition of 1 mM IPTG. Cells were harvested after 20 h of incubation with constant shaking and processed to obtain soluble (S) and insoluble (I) fractions. Equivalent protein amounts (10 µg) of each sample were loaded on 12% SDS-PAGE. Upon gel electrophoresis, gels were stained with Coomassie Brilliant Blue R-250 and unstained following standard protocols. Control (CTL) lanes correspond to cellular lysates of uninduced cultures. A sample of purified (His)6-MatN/Cg endolysin (57 kDa/57.7 kDa) was loaded on P lane. C, D Total amounts of recombinant endolysin (column height) and the ratio of soluble/insoluble protein (SP, gray dotted pattern; IP, diagonal lines pattern) were calculated after background normalization with ImageJ, open-source image processing software. Endolysin expression and solubility were analyzed in biological triplicates. Error bars represent SD of mean values. **p < 0.005 (soluble endolysin ratio of 20 °C group vs. higher induction temperature groups)
Expression of the endolysin genes cloned in a pET-32a vector was induced by the addition of IPTG at various concentrations. Our results showed that the amount of endolysin produced did not increase at IPTG concentrations ranging from 0.1 to 1.0 mM. Moreover, we did not detect substantial changes in the amount of either soluble or insoluble endolysin (Fig. 3).

Effect of inducer concentration on recombinant endolysins MatN and Cg solubility and yield. A, B E. coli C43(DE3) harboring pET-32a-MatN/Cg was induced at 20 °C by the addition of three different IPTG concentrations (0.1; 0.5 and 1 mM). Cells were harvested after 20 h of incubation with constant shaking and processed to obtain soluble (S) and insoluble (I) fractions. Equivalent protein amounts (10 µg) of each sample were loaded on 12% SDS-PAGE. Upon gel electrophoresis, gels were stained with Coomassie Brilliant Blue R-250 and unstained following standard protocols. Control (CTL) lanes correspond to cellular lysates of uninduced cultures. A sample of purified (His)6-MatN/Cg endolysin (57 kDa/57.7 kDa) was loaded on P lane. C, D Total amounts of recombinant endolysin (column height) and the ratio of soluble/insoluble protein (SP, gray dotted pattern; IP, diagonal lines pattern) were calculated after background normalization with ImageJ, open-source image processing software. Endolysin expression and solubility were analyzed in biological triplicates. Error bars represent SD of mean values
The use of solubility tags does not improve endolysin solubility
To enhance the yield of soluble endolysin; we resorted to using three widely used solubility tags (DsbC, Trx and Sumo) [19,20,21] placed as N-term fusions to the endolysin genes. Surprisingly, despite DsbC being identified by high throughput screening as an efficient partner for recombinant protein solubilization, it did not increase endolysin content in the soluble fraction. On the contrary, this tag caused a reduction in total recombinant endolysin produced. Trx also caused a reduction in the synthesis of the recombinant enzyme although less drastically than DsbC. In this regard, (His)6-MatN total expression was on average 3 times higher than (His)6-DsbC/(His)6-Trx MatN, (p < 0.001), and (His)6-Cg total expression was 0.5 times higher than (His)6-Trx-Cg (p = 0.035). Thus, these two fusion constructs bearing N-term tags failed to produce a clear benefit in the level of soluble endolysin present in the cell extract (Fig. 4).

Solubility N-term tags performance on soluble endolysin yields. A, B SDS PAGE showing the position of each fusion tag to recombinant endolysins; C, D E. coli C43(DE3) harboring pET-32a modified with N-term solubility tags (DsbC, Sumo and Trx) in frame fusions to MatN/Cg genes was induced at 20 °C by addition of 1 mM IPTG. Cells were harvested after 20 h of incubation with constant shaking and processed to obtain soluble (S) and insoluble (I) fractions. Equivalent protein amounts (10 µg) of each sample were loaded on 12% SDS-PAGE. Upon gel electrophoresis, gels were stained with Coomassie Brilliant Blue R-250 and unstained following standard protocols. Control (CTL) lanes correspond to cellular lysates of uninduced E. coli C43(DE3) harboring pET32a-MatN/Cg. E, F Total amounts of recombinant endolysin (column height) and the ratio of soluble/insoluble protein (SP, gray dotted pattern; IP, diagonal lines pattern) were calculated after background normalization with ImageJ, open-source image processing software. Endolysin expression and solubility were analyzed in biological triplicates. Error bars represent SD of mean values. **p < 0.005, *p < 0.05 (total endolysin expression of N-His group vs. N-Tags groups)
(His)6 Tag localization affects MatN solubility
Since the level of expression of a recombinant protein can vary when having either an N-term or a C-term (His)6 tag [22], we set to determine the effect of the former modification on the expression of MatN (Fig. 5). C-term (His)6 tag construct prompted a statistically significant increase of soluble endolysin when compared to the N-term bearing construction (p = 0.008).

(His)6 tag orientation on MatN soluble yields. A E. coli C43(DE3) harboring pET-32a N-term- (his)6 MatN or pET-22b C-term-(his)6 MatN were induced at 20 °C by addition of 1 mM IPTG. Cells were harvested after 20 h of incubation with constant shaking and processed to obtain soluble (S) and insoluble (I) fractions. Equivalent protein amounts (10 µg) of each sample were loaded on 12% SDS-PAGE. Upon gel electrophoresis, gels were stained with Coomassie Brilliant Blue R-250 and unstained following standard protocols. A sample of purified N-term-His MatN endolysin (57 kDa) was loaded on P lane. B Total amounts of recombinant endolysin (column height) and the ratio of soluble/insoluble protein (SP, gray dotted pattern; IP, diagonal lines pattern) were calculated after background normalization with ImageJ, open-source image processing software. Endolysin expression and solubility were analyzed in biological triplicates. Error bars represent SD of mean values. *p < 0.05 (comparison between soluble endolysin ratios)
Co-expression of chaperone proteins does not improve yields of soluble endolysin
Expression of proteins acting as chaperones during translation has been a field of research for several years, both at the level of understanding fundamental processes of native protein production in different biological systems as well as a way to increase the stability of recombinant proteins in various expression systems [23]. Chaperone proteins would help stabilize nascent polypeptides facilitating their correct folding into mature proteins. The availability of cloned sets of chaperones (DnaK and GroEL) in a plasmid compatible with our expression vector led us to test their co-expression along with our chosen endolysin clones. Our results showed that even when the mentioned chaperones were co-expressed with the endolysins MatN and Cg, a meager increase in the solubility of MatN and no noticeable increase in the solubility of Cg was seen (Fig. 6).

Co-expression of chaperones with MatN and Cg endolysins for solubility improvement. A, B E. coli C43(DE3) harboring pET-32a-MatN/Cg and chaperone (chap) encoding pG-KJE8 was induced at 20 °C by l-arabinose (0.25 mg/mL) and Tetracycline (5 ng/mL) 1 h before 1 mM IPTG addition. Cells were harvested after 20 h of incubation with constant shaking and processed to obtain soluble (S) and insoluble (I) fractions. Equivalent protein amounts (10 µg) of each sample were loaded on 12% SDS-PAGE. Upon gel electrophoresis, gels were stained with Coomassie Brilliant Blue R-250 and unstained following standard protocols. Control (CTL) lanes correspond to cellular lysates of uninduced E. coli C43(DE3) harboring pET32a-MatN/Cg. Black arrow heads indicate overexpression of endolysin, dnaK and groEL chaperones. C, D Total amounts of recombinant endolysin (column height) and the ratio of soluble/insoluble protein (SP, gray dotted pattern; IP, diagonal lines pattern) were calculated after background normalization with ImageJ, open-source image processing software. Endolysin expression and solubility were analyzed in biological triplicates. Error bars represent SD of mean values
Post-expression steps in endolysin production: impact of the use of detergents and osmolytes during the extraction process
Our results described above suggested that endolysin solubility could not be significantly influenced by modification of growth conditions, solubility tags and chaperones co-expression. Thus, we investigated the effects of adding detergents and different osmolytes during the cell pellet disruption [24, 25]. To this aim, we initially tested several detergents which are commonly used for bacterial protein extraction (Tween 20, Triton-X100, CHAPS, N-lauroylsarcosine, and Nonidet P40). We have also tested low molecular weight osmolytes such as polyalcohols, salts, and sugars (i.e., NaCl, CaCl2, mannitol, trehalose, glycerol, glycine betaine) which help to stabilize proteins. Our results showed a marginal improvement in the amount of extracted endolysin except for N-lauroylsarcosine when using detergents. Glycerol, trehalose or glycine betaine added to the lysis buffer increased the yield of soluble MatN endolysin two-fold while the effect on Cg endolysin was less pronounced (see Additional file 2).
To confirm these observations, we performed new assays that included the best performing osmolytes and N-lauroylsarcosine at different concentrations (0.25; 0.5 and 1% w/v) (Fig. 7). Supplementation of the lysis buffer with N-lauroylsarcosine caused a strong increment in the soluble yield of the endolysins in a concentration-dependent manner. NLS at 1% w/v increased both total and soluble MatN recovery (p = 0.022, p = 0.047) and the soluble recovery of Cg (p = 0.005). The addition of 0.5% w/v N-lauroylsarcosine to the extraction buffer did not affect endolysin activity as judged by zymograms, thus supporting its use (Additional file 3). Osmolytes added at the protein extraction step only marginally improved the recovery of endolysins in the soluble fraction.

Impact of the use of detergents and osmolytes during the extraction process. A, B E. coli C43(DE3) harboring pET-32a MatN/Cg was induced at 20 °C by adding 1 mM IPTG. Cells were harvested after 20 h of incubation with constant shaking and processed to obtain soluble (S) and insoluble (I) fractions. Cell pellets lysis was performed by a bead-beating approach in lysis buffer (50 mM Tris–HCl pH 8.0, 150 mM NaCl, 1% glycerol, and 1 mM PMSF) supplemented with various concentrations of N-lauroylsarcosine (N-LS), 25% glycerol, 0.75 M trehalose or 1 M glycine betaine. Equivalent protein amounts (10 µg) of each sample were loaded on 12% SDS-PAGE. Upon gel electrophoresis, gels were stained with Coomasie Brilliant Blue R-250 and unstained following standard protocols. C, D Total amounts of recombinant endolysin (column height) and the ratio of soluble/insoluble protein (SP, gray dotted pattern; IP, diagonal lines pattern) were calculated after background normalization with ImageJ, open-source image processing software. Endolysin expression and solubility were analyzed in biological triplicates. Error bars represent SD of mean values. *p < 0.05 (total MatN endolysin and ratio of soluble Cg endolysin vs. none additive group)
Discussion
As part of a large sequencing project on S. aureus phage genomics, our group has isolated and sequenced more than 30 phages active on this pathogen, making available the sequence of their endolysins (Suárez et al., submitted to Microbiology Spectrum, Spectrum00334-22). Upon phylogenetic analysis, we expressed two of them, which were distant from an evolutionary standpoint. Yield of recombinant endolysins is of outmost importance when considering their industrial production. From that perspective, we adopted a systematic strategy addressing the expression (temperature and inducer concentration, cloning strategy, use of solubility tags and co-expression of molecular chaperones) or recovery (lysis buffer composition: osmolytes and detergents) steps.
Our results indicate that cloning strategy potentially impact in endolysin expression depending on the position of the insert (C-end or N-end) regarding the (His)6 tag, in agreement with previous reports [22].
As mentioned, production at the bench scale of soluble recombinant endolysins targeting S. aureus was reported by many groups [15, 17, 18, 26,27,28,29,30,31,32,33]; however, inspection of the published data did not reveal how much protein was lost in the insoluble fraction in each case and often, the extraction conditions and final yields were not stated (Table 1). A traditional and straightforward strategy to improve the solubility of recombinant proteins in bacterial expression systems consists of tuning down expression through low growth temperatures or reducing the concentration of the chemical inducer of the expression system. In this way, peptide crowding in the cytoplasm leading to insolubility could be prevented [34,35,36]. According to the literature as exemplified in Table 1, growth temperatures ranging between 10–25 °C were the most used ones, and yields—when stated—were very uneven ranging from 0.5 to 80 mg/L [18, 26, 30, 33]. In our hands, the highest yields, approximately 38–42% of soluble MatN and Cg (25 mg/L of induced culture), were obtained upon expression at 20 °C for 20 h using 1 mM IPTG.
Our attempts to increase solubility by the introduction of N-term fusion partners such as Trx (~ 12 kDa), SUMO (~11 kDa) and DsbC (~ 23 kDa) failed to produce the expected effect; in fact, we obtained the opposite result since total protein expression decreased when compared to the His-tagged constructs. These results are not only counter-intuitive but also have not been reported for any other recombinant protein [34, 37]. The strategy of adding solubility tags as means to increase the yield of endolysins is debatable, as most of those tags represent a 20–50% increase of the fusion protein molecular weight. Elimination of unwanted peptide sequences would also require the addition of an additional protease recognition site in the construct. Thus, taken as a whole, this approach may not be cost/benefit useful unless a large proportion of the synthesized endolysin is recovered in the soluble cell extract fraction.
In a different approach to improve soluble endolysin recovery, the co-expression of molecular chaperones (such as DnaK, GroEL and TF, reviewed in [23, 34]) was tested. These chaperones would prevent aggregation by acting on exposed hydrophobic patches of the recombinant endolysin, thus stabilizing misfolded or partially folded molecules. Disappointingly, this approach did not increase the amount of soluble endolysins to a convenient level. There are several reports pointing out that chaperones sometimes are by themselves responsible for undesirable side effects, from proteolysis to reduced yields or recombinant protein aggregation [38].
There are numerous environmental factors affecting the protein stability during cell lysis, such as type of buffer, pH, salt concentrations, presence of stabilizing agents (osmolytes, detergents), among others [24, 25]. These lysis media parameters or additives can either act by diminishing aggregation and protein–protein interactions or by stabilizing newly synthetized polypeptides. Interestingly, Filatova et al. [39] analyzed the effect of low molecular weight osmolytes and cations on the thermal stability of LysK; their results point out that dilution of the enzyme, addition of trehalose, sorbitol, glycerol, Mg2+ or Ca2+ increased LysK stability 100-fold. However, none of these factors have previously been evaluated as mediators of enhanced endolysin recovery. Thus, we tested variables that could be compatible with industrial scaling up. Our results showed that N-lauroylsarcosine was the most effective detergent at concentrations of 0.5–1.0 w/v, increasing the recovery of soluble MatN and Cg up to more than 50% of the total endolysin (Fig. 7). During the lysis of the cell pellet, both cytoplasmic and outer membrane components will interact with recombinant proteins, trapping them through hydrophobic and anionic surfaces. The detergent may prevent aggregation by disrupting associations with highly insoluble, aggregation-prone outer membrane components [40]. As described by Žydziecki et al. [16], there is a proven interaction between the positively charged endolysin and nucleic acids, in this aspect, the detergent may displace the nucleic acid avoiding endolysin intermolecular interactions.
In summary, our results taken as a whole suggest that the use of N-lauroylsarcosine during cell lysis, may increase the yields of soluble recombinant full-length endolysins.
Conclusions
To our knowledge, this is the first systematic approach to evaluate factors influencing endolysins expression and extraction conditions. We found that low induction temperatures (20 °C) benefit the recovery of soluble enzyme and that over the range of 0.1–1 mM IPTG, there is no appreciable changes in endolysin yields. The position of the (His)6-tag in the polypeptide as well as extra amino acid residues produced by the cloning strategy may be potentially detrimental. Neither the use of fusions to solubility enhancers (Trx, DsbC and Sumo proteins) nor the co-expression of molecular chaperones increased the yield of recombinant endolysins. Our examination of the effect of adding tensioactives or osmolytes to the lysis buffer showed that 25% glycerol, 0.75 M trehalose and 1 M glycine betaine would benefit endolysins recovery but, anionic detergent N-lauroylsarcosine was the only addition making an visible difference in soluble protein yields with no impairment of enzymatic activity.
Clearly, there is plenty of room to improve expression of recombinant endolysins other than the ones approached in this work, namely alternative expression systems, host strains, different purification protocols, etc. However, we show here in that the use of lysis buffer additives (e.g., N-lauroylsarcosine), is a sensible strategy to increase the levels of soluble S. aureus endolysins.
Methods
Reagents, bacterial strains, media, and growth conditions
E. coli DH5α and E. coli C43(DE3) strains were used as cloning host and protein expression host, respectively. Strains were grown in Luria–Bertani (LB) medium at 37 °C with shaking at 150 rpm unless indicated otherwise. Ampicillin (Ap, 100 µg/mL) and Chloramphenicol (Cm, 20 µg/mL) were added when required.
Endolysin cloning
The isolation, characterization and genomic sequencing of a set of bacteriophages active on Staphylococcus aureus will be reported elsewhere (Suárez et al., submitted to Microbiology Spectrum, Spectrum00334-22). Bioinformatics analysis using BLAST with LysK endolysin gene sequence as query allowed for the identification of 25 putative endolysin encoding genes. Cg and MatN endolysin encoding genes representing different phylogeny groups (41.53% identity, E = 5.10–4), as determined using MEGA 6.0 molecular evolutionary analysis software, were chosen for cloning and expression studies. DNA isolation of the chosen bacteriophages was carried out as previously described [41].
Cg and MatN encoding genes were cloned by Restriction Free Cloning (RFC) [42] into pET32a vector as described by Correa et al. [43]. This vector suite enables the parallel cloning of the same PCR product into different expression vectors harboring a selection of solubility fusion TAGs. The endolysin genes were fused as a C-terminal partner with (His)6-thioredoxin A (his6-Trx), (His)6-small ubiquitin-like modifier protein (his-SUMO), (His)6-disulfide bond isomerase C (his6-DsbC), or (His)6 tag alone. MatN gene was also cloned by restriction digestion and ligation (RD + L) into NdeI-BamHI digested pET-22b to generate a C-terminal (His)6 tag version of the recombinant endolysin. PET32a vector was also used when the impact of the expression of molecular chaperones on endolysin solubility was determined. All constructs were confirmed by sequencing. Primers used for endolysin cloning are listed in Table 2.
Endolysin expression
Flasks containing 10 mL of LB supplemented with the appropriate antibiotics were inoculated with an aliquot (1/100 v/v) of stationary cultures of E. coli C43(DE3) harboring pET-32a-Cg or pET32a-MatN plasmids. Cultures were incubated at 37 °C with constant shaking (150 rpm) until they reached OD600 = 0.6–0.8. Induction was started by the addition of various IPTG concentrations (ranging from 0.1 to 1 mM), and growth was continued for 20 h at 20 °C, 25 °C or 30 °C. Induced cells (10 mL) were harvested by centrifugation at 3500g for 5 min at 4 °C and the pellet was re-suspended in 1 mL of lysis buffer (50 mM Tris–HCl pH 8.0, 150 mM NaCl, 1% glycerol and 1 mM PMSF). Disruption of cells was accomplished using a bead beater homogenizer (Fast-Prep®) following 6 cycles of 15 s bursts and 2 min rest on ice protocol. Lysates were centrifuged at 16,000g for 30 min at 4 °C to separate soluble and insoluble fractions; insoluble pellets were re-suspended thoroughly in 250 µL lysis buffer and both fractions were either processed immediately or kept frozen at − 80 °C until use.
Cellular lysate analysis
Cellular lysates were analyzed by 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Protein concentration was determined by Bradford Coomassie Brilliant Blue assay [44] to ensure loading of equivalent protein mass (10 µg) on each lane. Upon gel electrophoresis, gels were stained with Coomasie Brilliant Blue and unstained following standard protocols [45]. Following image acquisition, changes in soluble endolysin content under each experimental condition were visualized and quantitated using ImageJ (open-source image processing software). The ratio of soluble/total recombinant endolysin was calculated after normalization with background values obtained from the pixel densities of bands corresponding to endogenous proteins. Unless indicated otherwise, endolysin expression and solubility was analyzed in biological triplicates.
Chaperone co-induction assay
The plasmid pG-KJE8 (TAKARA BIO INC) encoding chaperones (dnaK and groEL) and their co-factors (dnaJ-grpE-groES), was transformed into the expression strain E. coli C43 (DE3) harboring the chosen endolysin plasmid. Co-expression of chaperones was induced by L-arabinose (0.25 mg/mL) and Tetracycline (5 ng/mL) 1 h before IPTG addition. Once induced, cultures were grown with shaking for 20 h at 20 °C, harvested by centrifugation at 3500g for 5 min at 4 °C and the pellet was re-suspended in 1 mL of lysis buffer.
Solubility tags expression assay
Endolysin genes were cloned using the RFC approach in a series of modified pET-32a vectors [43] that enables cloning of fragments as fusion with a selection of solubility tags with a single megaprimer pair. Oligonucleotides for megaprimer generation were designed according to Bond and Naus [42]. In brief, the amplification conditions were as follows: 30 s at 98 °C followed by 30 cycles of 98 °C for 10 s, 62 °C for 30 s and 72 °C for 30 s with a final extension step at 72 °C for 5 min. Megaprimers were purified by extraction from agarose gel. The integration of megaprimers into the vectors were done under the following reaction parameters: 30 s at 98 °C and 25 cycles of 98 °C for 10 s, 60 °C for 30 s and 72 °C for 5 min with a final extension step at 72 °C for 7 min. Subsequently, an aliquot (10 µL) of the reaction mixture was digested with 1 µL of DpnI (Thermo Scientific) ON at 37 °C in order to remove the parental plasmid, and this was transformed into 100 µL of competent DH5α E. coli cells.
Lysis buffer supplementation for solubility enhancement
Buffer supplementation with osmolytes or detergents were tested as solubility enhancers by incorporation of each one at the following concentrations into the standard lysis buffer before cell disruption: 300 mM NaCl, 25% v/v glycerol, 0.5 M mannitol, 0.75 M trehalose, and 1 M glycine betaine; detergents were added at 0.5% v/v in the case of Tween 20, Triton-X100 or Nonidet P40; or 0.5% w/v in the case of CHAPS or N-lauroylsarcosine.
Statistical analysis
Statistical analysis was performed using JASP (Version 0.11, JASP Team, 2019). Normal distribution of data sets was evaluated through Shapiro–Wilk test, designed to test normality for small data-size, and homoscedasticity was judged by Levene's test. The two tailed Student t-test was used when comparing means from two experimental groups, whereas analysis of variance (ANOVA) followed by Tukey’s post-hoc test was chosen for multiple comparisons. When criteria of Normal distribution or equality of variances were not met, we applied non-parametric tests. Statistically significant differences are denoted by asterisks (*p < 0.05, **p < 0.005).
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- RFC:
-
Restriction free cloning
- PG:
-
Peptidoglycan
- CBD:
-
Cell binding domain
- EAD:
-
Enzymatically active domain
- PMSF:
-
Phenylmethylsulfonyl fluoride
- PBS:
-
Phosphate buffer saline
- DTT:
-
Dithiothreitol
References
Antibiotic resistance [Internet]. [cited 2022 Jan 24]. Available from: https://www.who.int/news-room/fact-sheets/detail/antibiotic-resistance.
Marston HD, Dixon DM, Knisely JM, Palmore TN, Fauci AS. Antimicrobial resistance. JAMA J Am Med Assoc. 2016;316:1193–204.
Domingo-Calap P, Delgado-Martínez J. Bacteriophages: protagonists of a post-antibiotic era. Antibiotics. 2018;7:1–16.
Kutateladze M, Adamia R. Bacteriophages as potential new therapeutics to replace or supplement antibiotics. Trends Biotechnol Elsevier Current Trends. 2010;28:591–5.
Hermoso JA, García JL, García P. Taking aim on bacterial pathogens: from phage therapy to enzybiotics. Curr Opin Microbiol. 2007;10:461–72.
Nelson DC, Schmelcher M, Rodriguez-Rubio L, Klumpp J, Pritchard DG, Dong S, et al. Endolysins as antimicrobials. Adv Virus Res Academic Press. 2012;83:299–365.
Murray E, Draper LA, Ross RP, Hill C. The advantages and challenges of using endolysins in a clinical setting. Viruses. 2021;13:680.
Totté JEE, van Doorn MB, Pasmans SGMA. Successful treatment of chronic Staphylococcus aureus—related dermatoses with the topical endolysin staphefekt SA.100: a report of 3 cases. Case Rep Dermatol Karger Publishers. 2017;9:19–25.
Gutiérrez D, Fernández L, Rodríguez A, García P. Are phage lytic proteins the secret weapon to kill Staphylococcus aureus? MBio. 2018;9:1–17.
São-José C. Engineering of phage-derived lytic enzymes: improving their potential as antimicrobials. Antibiotics. 2018;7:29.
Love MJ, Bhandari D, Dobson RCJ, Billington C. Potential for bacteriophage endolysins to supplement or replace antibiotics in food production and clinical care. Antibiotics. 2018;7:1–25.
De Maesschalck V, Gutiérrez D, Paeshuyse J, Lavigne R, Briers Y. Advanced engineering of third-generation lysins and formulation strategies for clinical applications. Crit Rev Microbiol Taylor and Francis Ltd. 2020;46:548–64.
Manoharadas S, Witte A, Bläsi U. Antimicrobial activity of a chimeric enzybiotic towards Staphylococcus aureus. J Biotechnol J Biotechnol. 2009;139:118–23.
O’Flaherty S, Coffey A, Meaney W, Fitzgerald GF, Ross RP. The recombinant phage Lysin LysK has a broad spectrum of lytic activity against clinically relevant Staphylococci, including methicillin-resistant Staphylococcus aureus. J Bacteriol Am Soc Microbiol (ASM). 2005;187:7161.
Daniel A, Euler C, Collin M, Chahales P, Gorelick KJ, Fischetti VA. Synergism between a novel chimeric lysin and oxacillin protects against infection by methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. 2010;54:1603–12.
Alaksandr Ž, Sergey G, Maksim P, Sergey K, Niyaz S, Uladzimir P, et al. Efficient matrix-assisted refolding of the recombinant anti-staphylococcal truncated endolysin LysKCA and its structural and enzymatic description. Protein Expr Purif Academic Press. 2020;174:105683.
Gu J, Xu W, Lei L, Huang J, Feng X, Sun C, et al. LysGH15, a novel bacteriophage lysin, protects a murine bacteremia model efficiently against lethal methicillin-resistant Staphylococcus aureus infection. J Clin Microbiol Am Soc Microbiol (ASM). 2011;49:111–7.
Obeso JM, Martínez B, Rodríguez A, García P. Lytic activity of the recombinant staphylococcal bacteriophage phiH5 endolysin active against Staphylococcus aureus in milk. Int J Food Microbiol Elsevier BV. 2008;128:212–8.
Nozach H, Fruchart-Gaillard C, Fenaille F, Beau F, Ramos OHP, Douzi B, et al. High throughput screening identifies disulfide isomerase DsbC as a very efficient partner for recombinant expression of small disulfide-rich proteins in E. coli. Microb Cell Fact. 2013;12:1–16.
Lavallie ER, Lu Z, Diblasio-Smith EA, Collins-Racie LA, Mccoy JM. Thioredoxin as a fusion partner for production of soluble recombinant proteins in Escherichia coli. Methods Enzymol Academic Press. 2000;326:322–40.
Marblestone JG, Edavettal SC, Lim Y, Lim P, Zuo X, Butt TR. Comparison of SUMO fusion technology with traditional gene fusion systems: Enhanced expression and solubility with SUMO. Protein Sci Wiley-Blackwell. 2006;15:182.
Tham HY, Song AAL, Yusoff K, Tan GH. Effect of different cloning strategies in pet-28a on solubility and functionality of a staphylococcal phage endolysin. Biotechniques. 2020;69:161–70.
Kolaj O, Spada S, Robin S, Wall JG. Use of folding modulators to improve heterologous protein production in Escherichia coli. Microb Cell Fact. 2009;8:9.
Leibly DJ, Nguyen TN, Kao LT, Hewitt SN, Barrett LK, van Voorhis WC. Stabilizing additives added during cell lysis aid in the solubilization of recombinant proteins. PLoS ONE. 2012;7:e52482.
Lebendiker M, Danieli T. Production of prone-to-aggregate proteins. FEBS Lett. 2014;588:236–46.
Cha Y, Son B, Ryu S. Effective removal of staphylococcal biofilms on various food contact surfaces by Staphylococcus aureus phage endolysin LysCSA13. Food Microbiol Academic Press. 2019;84:103245.
Fujiki J, Nakamura T, Furusawa T, Ohno H, Takahashi H, Kitana J, et al. Characterization of the lytic capability of a lysk-like endolysin, lys-phiSA012, derived from a polyvalent staphylococcus aureus bacteriophage. Pharmaceuticals. 2018;11:25.
Linden SB, Zhang H, Heselpoth RD, Shen Y, Schmelcher M, Eichenseher F, et al. Biochemical and biophysical characterization of PlyGRCS, a bacteriophage endolysin active against methicillin-resistant Staphylococcus aureus. Appl Microbiol Biotechnol. 2015;99:741–52.
Abaev I, Foster-Frey J, Korobova O, Shishkova N, Kiseleva N, Kopylov P, et al. Staphylococcal Phage 2638A endolysin is lytic for Staphylococcus aureus and harbors an inter-lytic-domain secondary translational start site. Appl Microbiol Biotechnol NIH Public Access. 2013;97:3449–56.
Kashani HH, Fahimi H, Goli YD, Moniri R. A novel chimeric endolysin with antibacterial activity against methicillin-resistant Staphylococcus aureus. Front Cell Infect Microbiol. 2017;7:290.
Lu Y, Wang Y, Wang J, Zhao Y, Zhong Q, Li G, et al. Phage endolysin LysP108 showed promising antibacterial potential against methicillin-resistant Staphylococcus aureus. Front Cell Infect Microbiol. 2021;11:668430.
Keary R, McAuliffe O, Ross RP, Hill C, O’Mahony J, Coffey A. Genome analysis of the staphylococcal temperate phage DW2 and functional studies on the endolysin and tail hydrolase. Bacteriophage Taylor & Francis. 2014;4:e28451.
Jun SY, Jung GM, Yoon SJ, Oh MD, Choi YJ, Lee WJ, et al. Antibacterial properties of a pre-formulated recombinant phage endolysin, SAL-1. Int J Antimicrob Agents Elsevier. 2013;41:156–61.
Costa S, Almeida A, Castro A, Domingues L. Fusion tags for protein solubility, purification, and immunogenicity in Escherichia coli: the novel Fh8 system. Front Microbiol Front Res Found. 2014;5:63.
Rosano GL, Ceccarelli EA. Recombinant protein expression in Escherichia coli: advances and challenges. Front Microbiol. 2014;5:1–17.
Rosano GL, Morales ES, Ceccarelli EA. New tools for recombinant protein production in Escherichia coli: a 5-year update. Protein Sci Wiley-Blackwell. 2019;28:1412–22.
Waugh DS. Making the most of affinity tags. Trends Biotechnol Elsevier Curr Trends. 2005;23:316–20.
Martínez-Alonso M, García-Fruitós E, Ferrer-Miralles N, Rinas U, Villaverde A. Side effects of chaperone gene co-expression in recombinant protein production. Microb Cell Fact. 2010;9:1–6.
Filatova LY, Becker SC, Donovan DM, Gladilin AK, Klyachko NL. LysK, the enzyme lysing Staphylococcus aureus cells: specific kinetic features and approaches towards stabilization. Biochimie Elsevier Masson SAS. 2010;92:507–13.
Frankel S, Sohn R, Leinwand L. The use of sarkosyl in generating soluble protein after bacterial expression. Proc Natl Acad Sci USA. 1991;88:1192–6.
Abatángelo V, Peressutti Bacci N, Boncompain CA, Amadio AA, Carrasco S, Suárez CA, et al. Broad-range lytic bacteriophages that kill Staphylococcus aureus local field strains. PLoS ONE. 2017;12:1–22.
Bond SR, Naus CC. RF-Cloning.org: an online tool for the design of restriction-free cloning projects. Nucleic Acids Res Oxford University Press. 2012;40:209.
Correa A, Ortega C, Obal G, Alzari P, Vincentelli R, Oppezzo P. Generation of a vector suite for protein solubility screening. Front Microbiol. 2014;5.
Ernst O, Zor T. Linearization of the Bradford protein assay. J Vis Exp. 2010;38:1918.
Sambrook J, Russell DW. SDS-polyacrylamide gel electrophoresis of proteins. Cold Spring Harb Protoc. Cold Spring Harbor Laboratory; 2006;2006:pdb.prot4540-pdb.prot4540.
Acknowledgements
Not applicable.
Funding
HRM acknowledges funding from ANPCyT, Argentina (PICT 2018-0867).
Author information
Authors and Affiliations
Contributions
Conception and design of the work: HRM and CLB. Acquisition and analysis of data: CS, CAB and CLB. Interpretation of data: HRM and CLB. Drafted the work: HRM and CLB. Substantively revised the manuscript: HRM, CLB and EAC. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1
: Hydrophobicity plots by the Kyle-Doolittle scale. Endolysin hydrophobic regions prediction.
Additional file 2
: Impact of different detergents and osmolytes on soluble Cg and MatN endolysin recovery. Lysis buffer additives screening
Additional file 3
: Zymograms of purified endolysins. SDS–Polyacrylamide gels containing autoclaved RN 4220 S. aureus cells were used to visualize the peptidoglycan hydrolyzing activity of recombinant His-MatN and His-Cg extracted in lysis buffer with 0.5% w/v N-Lauroylsarcosine or without the detergent.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Balaban, C.L., Suárez, C.A., Boncompain, C.A. et al. Evaluation of factors influencing expression and extraction of recombinant bacteriophage endolysins in Escherichia coli. Microb Cell Fact 21, 40 (2022). https://doi.org/10.1186/s12934-022-01766-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-022-01766-9