
Abstract
Background
Circular RNAs (circRNAs), a novel class of non-coding RNAs, play an important regulatory role in pulmonary arterial hypertension (PAH); however, the specific mechanism is rarely studied. In this study, we aimed to discover functional circRNAs and investigate their effects and mechanisms in hypoxia-induced pulmonary vascular remodelling, a core pathological change in PAH.
Methods
RNA sequencing was used to illustrate the expression profile of circRNAs in hypoxic PAH. Bioinformatics, Sanger sequencing, and quantitative real-time PCR were used to identify the ring-forming characteristics of RNA and analyse its expression. Then, we established a hypoxia-induced PAH mouse model to evaluate circRNA function in PAH by echocardiography and hemodynamic measurements. Moreover, microRNA target gene database screening, fluorescence in situ hybridisation, luciferase reporter gene detection, and western blotting were used to explore the mechanism of circRNAs.
Results
RNA sequencing identified 432 differentially expressed circRNAs in mouse hypoxic lung tissues. Our results indicated that circ-Ntrk2 is a stable cytoplasmic circRNA derived from Ntrk2 mRNA and frequently upregulated in hypoxic lung tissue. We further found that circ-Ntrk2 sponges miR-296-5p and miR-296-5p can bind to the 3′-untranslated region of transforming growth factor-β1 (TGF-β1) mRNA, thereby attenuating TGF-β1 translation. Through gene knockout or exogenous expression, we demonstrated that circ-Ntrk2 could promote PAH and vascular remodelling. Moreover, we verified that miR-296-5p overexpression alleviated pulmonary vascular remodelling and improved PAH through the TGF-β1/p38 MAPK pathway.
Conclusions
We identified a new circRNA (circ-Ntrk2) and explored its function and mechanism in PAH, thereby establishing potential targets for the diagnosis and treatment of PAH. Furthermore, our study contributes to the understanding of circRNA in relation to PAH.
Similar content being viewed by others


Background
Pulmonary arterial hypertension (PAH) is a progressive and fatal cardiopulmonary disease characterised by irreversible pulmonary vascular remodelling, wherein right heart hypertrophy and heart failure are the leading causes of death [1]. Pulmonary artery smooth muscle cell (PASMC) over-proliferation is involved in PAH development, thus playing a crucial role in pulmonary vasculature reconstruction owing to its excellent ability to regulate contraction and synthesis dynamically [2]. Regrettably, the current PAH therapies mainly aim to enhance the NO and prostacyclin pathways and inhibit the endothelin pathway. However, only symptoms of the disease can be alleviated through this approach, failing to effectively reverse the hallmark pathological changes in pulmonary vascular remodelling. Consequently, the five-year mortality rate of PAH remains high [3, 4]. Therefore, finding novel targets to inhibit PASMC over-proliferation is crucial.
In recent years, non-coding RNAs (ncRNAs) have been widely reported to play an important role in the development of PAH and other human diseases [5]. Circular RNAs (circRNAs), as a class of highly conserved regulatory ncRNAs, are derived from exons, introns, or intergenic regions through back-splicing [6, 7]. Numerous studies have reported that circRNA can sponge microRNA (miRNA) to alter physiological and pathological states in organisms [8,9,10]. Due to their unique covalent closed-loop structure, circRNAs exhibit superior stability compared with linear RNA [11,12,13]. Combined with the strong species preservation, tissue specificity, and expression abundance in mammals, circRNAs hold promise as ideal biomarkers for disease diagnosis and intervention [12,13,14]. CircRNA function and mechanism in PAH have attracted increasing attention in recent years [15].
However, studies on the function and mechanism of circRNA in PAH are still rare [16, 17]. Therefore, constructing the circRNA expression profile in this disease and identifying new functional circRNAs are of great significance for understanding their role in PAH and developing new therapeutic targets.
In this study, we compared the circRNA expression profile in the lung tissues of normoxic and hypoxic mice. We further investigated the function and mechanism of the newly identified circRNA in PAH at both the in vitro and in vivo levels, suggesting its potential application as a novel therapeutic target for PAH.
Methods
Reagents
The primary antibodies used in the present study are shown in Additional file 2: Table S1.
Cell culture and treatment
Male C57BL/6 mice (6–8 weeks old, 20–25 g) were used for PASMCs isolation. The mice were anaesthetised via isoflurane inhalation (1–2%, 0.6–1 L/min). The lungs were quickly removed and transferred to a phosphate-buffered saline (PBS) solution containing 1% penicillin and streptomycin (Gibco, USA). Under an anatomical microscope (NIKON, Japan), the lung tissue around the pulmonary artery was cleared first, and then the extravascular fascia was peeled off. Using microforceps, the endothelial cells were scraped from the blood vessel lining. The pulmonary artery was snipped and digested in 0.2% collagenase I (Gibco, USA) at 37 °C for 40 min. The primary cells were cultured in Dulbecco's modified eagle medium (Gibco, USA) supplemented with 20% fetal bovine serum (Gibco, USA) and 1% penicillin and streptomycin at 37 ℃ under 5% CO2 for 3–5 days. The third and fourth generations of PASMCs were used in this study. The PASMCs were divided into the following groups: 1. normoxia (NOR), HYP, HYP + si-circ-Ntrk2 negative control (SI-NC), HYP + si-circ-Ntrk2 (SI, 50 nM); 2. NOR, HYP, and HYP + circ-Ntrk2 plasmid treatment NC (T-NC), HYP + circ-Ntrk2 plasmid treatment (T, 50 nM); 3. NOR, HYP, HYP + miR-296-5p inhibitor-NC (inhibitor-NC), HYP + miR-296-5p inhibitor (Inhibitor, 100 nM); 4. NOR, HYP, HYP + miR-296-5p mimic negative control (Mimic-NC), HYP + Mimic (50 nM); 5. NOR, HYP, HYP + SI-NC + Inhibitor-NC, HYP + SI + Inhibitor-NC, HYP + SI + Inhibitor. The NOR group (21% O2, 74% N2, and 5% CO2) and the HYP group (5% O2, 90% N2, and 5% CO2) were incubated at 37 °C for 48 h.
Quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA was extracted by a RNeasy® Mini Kit (QIAGEN, Hilden, Germany). RNase R (Beyotime, Shanghai, China) was used to detect circRNA. Genomic DNA (gDNA) was isolated using a Genomic DNA Extraction Kit (Omega Bio-Tek, Norcross, GA, USA). Quantification of RNA and gDNA was performed using the TOROIVD® qRT Master Mix and SYBR Green qPCR Master Mix (TOROIVD, Shanghai, China). miRNA was isolated using the miRNA Extraction Kit (Sangon Biotech, Shanghai, China), and a Bulge-Loop miRNA qRT-PCR Starter Kit (RiboBio, Guangzhou, China) was used to perform real-time PCR. The circRNA and mRNA levels were normalised to those of β-actin, and miRNA was normalised to that of U6 and determined using the 2−ΔΔCt method. The primer sequences used in this study are shown in Additional file 3: Table S2.
Immunofluorescence
Lung tissues were fixed using a tissue fixator overnight, permeated with 0.1% Triton X-100, and then blocked with 5% bovine serum albumin. Then, sections were incubated with anti-alpha-smooth muscle actin antibodies (1:100) overnight at 4 °C. After rinsing the sections with PBS three times, we incubated the sections with fluorescence-conjugated secondary antibodies. The nuclei were counterstained with 4,6-diamidino-2-phenylindole (DAPI). The fluorescence microscope (Leica DMi8, Wetzlar, Germany) was used to capture fluorescent images.
Western blot assay
Lung tissues were homogenised with a Protein Extraction Reagent (BOSTER, Wuhan, China) using an automatic homogeniser (FastPrep-24 5G, MP Biomedicals, Santa Ana, CA, USA). Supernatants were collected after the homogenates were centrifuged (12,000 rpm, 4 °C) for 30 min. PASMC proteins were extracted with Protein Extraction Reagent (BOSTER). Western blotting was performed as previously described [18] using antibodies specific for PCNA (1:1000), cyclin D1 (1:1000), transforming growth factor-β1 (TGF-β1) (1:1000), p38 MAPK (1:1000), and β-tubulin (1:1000).
Proliferation assays
Cells were incubated in 96-well plates for 48 h with a density of 1 × 104 cells per well. Cell proliferation was analysed with 5-ethynyl-20-deoxyuridine (EdU, Abcam, Waltham, MA, USA) and cell counting kit-8 (CCK8, Dojindo, Japan) assay kits.
Sequencing analysis
Total RNA was isolated, as mentioned above. After the completion of product inspection, the NEBNext® Multiplex Small RNA Library Prep Set for Illumina (NEB, Ipswich, MA, USA) was used for sRNA library construction. Then, the sRNA libraries were sequenced by the HiSeq 2500 platform (Illumina, San Diego, CA, USA).
RNA target prediction
Putative circRNA and miRNA targets were identified by analysis tools, including circBase (http://www.circbase.org/), miRanda (http://www.microrna.org/microrna/home.do), TargetScan (http://www.targetscan.org/), miRWalk (http://www.ma.uni-heidelberg.de/apps/zmf/mirwalk/), miRDB (http://mirdb.org/miRDB/), and miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/index.html).
Dual luciferase reporter assay
Dual luciferase reporter analysis of potential binding sites between circ-Ntrk2 and miR-296-5p was performed using the miRanda web tool, whereas targeted binding between miR-296-5p and TGF-β1 was analysed using TargetScan, miRDB, miRTarBase, and miRWalk web tools. Circ-Ntrk2 wild-type (circ-Ntrk2-WT) and circ-Ntrk2 mutant (circ-Ntrk2-MUT) carrying the mutated binding sites for miR-296-5p, as well as TGF-β1 wild-type (TGF-β1-WT) and TGF-β1 mutant carrying the mutated binding sites for miR-296-5p (TGF-β1-MUT) were cloned into the pmiR-RB-ReportTM vector (RiboBio); the primers are shown in Additional file 3: Table S2. PASMCs were transfected with 100 ng pmirGLO vector, 50 nM miR-296-5p mimics, and their negative control (miR-NC) using Attractene Transfection Reagent (Qiagen, Germantown, MD, USA). After 48 h, the Dual Luciferase® Reporter Assay System (E1910, Promega, Madison, WI, USA) was used to measure firefly and Renilla luciferase activities.
Oligonucleotide design and transfection
For circ-Ntrk2 and miR-296-5p knockdown or overexpression, small interfering RNA (siRNA, si-circ-Ntrk2), circ-Ntrk2 plasmid, miR-296-5p inhibitor, and mimic were obtained from RiboBio. The siRNA, mimic, and inhibitor were transfected using the Ribo FECT™ CP transfection kit (RiboBio). Attractene Transfection Reagent (Qiagen, USA) was used for plasmid transfection. The sequences used are shown in Additional file 3: Table S2.
Animal models
Male C57BL/6 mice (6–8 weeks, 20–25 g) were obtained from Vital River Laboratory Animal Technology (Beijing, China). To verify the function of circ-Ntrk2, we randomly divided the mice into four groups: NOR, HYP, HYP + SI-NC, and HYP + SI (20 nmol, i.p.). To identify the function of miR-296-5p, we randomly divided the mice into four groups: NOR, HYP, HYP + miR-296-5p agomir-NC (Agomir-NC), and HYP + Agomir (20 nmol, i.p.). To investigate whether circ-Ntrk2 aggravated HYP-induced PAH by inhibiting the negative regulatory effects of miR-296-5p on TGF-β1, we randomly divided the mice were into five groups: NOR, HYP, HYP + SI-NC + miR-296-5p antagomir NC (ANT-NC), HYP + SI + ANT-NC, and HYP + SI + ANT (50 nmol, i.p.). The HYP group mice were raised by supplying 10% O2 for 21 days, while the NOR group was housed in ambient air.
Echocardiography and measurement of hemodynamic
The mice were anaesthetised via isoflurane inhalation (1–2%, 0.6–1 L/min) and were allowed to moult. High-Resolution Small Animal Ultrasound Imaging System (Vevo 3100, Fujifilm Visual Sonics, Canada) was used to measure the pulmonary artery velocity time integral (PAVTI) and pulmonary artery acceleration time (PAAT). PowerLab 8/35 Multichannel Biological Signal Recording System (AD Instruments, Bella Vista, AUS) was used to record right ventricular systolic pressure (RVSP). After the mice were sacrificed, the right ventricle (RV) was dissected, and the weights of the RV, septum (S), and left ventricle (LV) were recorded. The ratio of RV to (LV + S) weight was calculated to assess the degree of RV hypertrophy.
Measurement of pulmonary vascular remodelling and collagen deposition
Lung tissue sections were stained with haematoxylin and eosin (H&E). Representative pulmonary arteries with diameters ranging from 25 to 100 µm were selected for further analysis. The ratio of pulmonary artery wall thickness (WT) to total thickness (TT, WT/TT) and the ratio of wall area (WA) to total area (TA, WA/TA) were analysed by Image-Pro Plus 6.0 (Media Cybernetics, Bethesda, MD, USA). Sections were stained with Masson’s trichrome. ImageJ 1.8.0.112 software (NIH, Bethesda, MD, USA) was used to calculate the ratio of collagen fibre area to pulmonary artery WA.
RNA fluorescence in situ hybridisation (FISH)
Digoxin-labelled sense or antisense probes were synthesised for the circ-Ntrk2 junction sequence (Additional file 3: Table S2). Hybridisation was performed using a FISH kit (RiboBio), and the nuclei were counterstained with DAPI.
Statistical analysis
GraphPad Prism 7.0 (Graph Pad Software Inc., San Diego, CA, USA) was used for statistical analysis of the data, and the results are presented as mean ± standard deviation (SD). Student’s t-test and one-way analysis of variance were used for two-group comparisons and multiple comparisons, respectively. Statistical significance was set at P < 0.05.
Results
Circ-Ntrk2 upregulates in the lung tissues of HYP-induced PAH mice and PASMCs
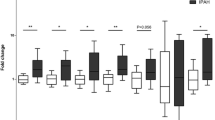
To study the expression profile and functional mechanism of circRNAs in HYP-induced PAH, we constructed a PAH mouse model (Additional file 1: Fig. S1a and b). Further, we screened 432 differentially expressed circRNAs (filtered by log2 (fold change >|1|) and P < 0.05) in HYP-exposed tissues compared with those in NOR using high-throughput transcriptome sequencing (Fig. 1a and b). Among these circRNAs, 148 were significantly upregulated and 284 were downregulated; mmu_circ: chr13:58960530–58962916 showed the most significant changes. Comparing our results with circBase (http://www.circbase.org/) revealed 30 recorded circRNAs, where mmu_circ: chr13:58960530–58962916 was also found among them. Three circRNAs (mmu_circ: chr13:58960530–58962916, mmu_circ: chr16:31,153,792–31,160,474, and mmu_circ: chr3:123304359–123337258) that were stably upregulated under hypoxic conditions were selected for further verification (Fig. 1c). The qPCR results showed that mmu_circ: chr13:58960530–58962916 was most significantly upregulated in hypoxic lung tissues and PASMCs, consistent with the sequencing results (Fig. 1d). Therefore, it was selected as the object for subsequent research. Also, since it is derived from a protein-coding locus Ntrk2 located on chromosome 13qB1 (Fig. 1e), we named this circRNA “circ-Ntrk2.”

Circ-Ntrk2 is upregulated in lung tissues of hypoxia-induced PAH mice and PASMCs. a Heat map analysis of circular RNA (circRNA) in lung tissues from hypoxia-exposed mice. b Volcano map analysis of circRNAs. c The differentially expressed circRNAs in the sequencing results were matched with the circBase data. d Quantitative polymerase chain reaction (qPCR) analysis of circRNA expression in hypoxic mouse lung tissues and PASMCs (n = 5). e Position of circ-Ntrk2 in its host gene Ntrk2. f Circ-Ntrk2 was spliced from the end of the mRNA Ntrk2 single exon; Sanger sequencing was used to verify the correctness of the cyclization site. g PCR assay with divergent and convergent primers showing the amplification of circRNA from cDNA or genomic DNA (gDNA), while GAPDH was used as a negative control. The gel or blot images have been cropped and the complete images have been submitted separately (Additional file 4). h qRT-PCR analysis of circ-Ntrk2 and Ntrk2 mRNA after treatment with RNase R. i Fluorescence in situ hybridisation (FISH) of lung tissues of mice exposed to hypoxia. All values are presented as the mean ± SD. *P < 0.05, **P < 0.01
Circ-Ntrk2 is a stable cytoplasmic circRNA derived from Ntrk2 mRNA
To further verify the ring structure of circ-Ntrk2, we first confirmed its junction site by Sanger sequencing analysis. The genomic structure showed that circ-Ntrk2 is looped by the seventh and ninth exons of the Ntrk2 gene (Fig. 1f). Circ-Ntrk2 was validated via PCR using divergent primers from cDNA rather than gDNA (Fig. 1g). Reverse-transcribed cDNA after RNase R treatment using qPCR showed no significant changes in circ-Ntrk2 compared with the untreated group, while the GAPDH gene and Ntrk2 mRNA were significantly downregulated, indicating good circ-Ntrk2 resistance to RNase R, further supporting the possibility of its cyclisation (Fig. 1h). FISH demonstrated that circ-Ntrk2 was preferentially localised within the PASMC cytoplasm (Fig. 1i). These results indicated that circ-Ntrk2 is a stable cytoplasmic circRNA derived from Ntrk2 mRNA.
Circ-Ntrk2 knockdown alleviated pulmonary vascular remodelling and improved PAH
To investigate the role of circ-Ntrk2 in PAH, we knocked down circ-Ntrk2 using siRNA (Fig. 2a and Additional file 1: Fig. S1c), and the efficiency was determined using qRT-PCR (Additional file 1: Fig. S1d). Echocardiography revealed that PAAT and PAVTI were apparently shorter under HYP, with circ-Ntrk2 silencing, which was reversed (Fig. 2b). Additionally, circ-Ntrk2 silencing attenuated the increase in RVSP and RV/(LV + S) induced by HYP (Fig. 2c). H&E staining and immunofluorescence results indicated that silencing circ-Ntrk2 reversed HYP-induced pulmonary vascular remodelling (Fig. 2d). Masson staining showed that circRNA knockdown improved perivascular collagen deposition in the disease state. In contrast, H&E staining of the right ventricular wall showed that circ-Ntrk2 knockdown significantly improved HYP-induced right ventricular cardiomyocyte hypertrophy (Fig. 2e). Western blotting verified that knockdown of circ-Ntrk2 reversed HYP-induced upregulation of proliferation-related proteins such as PCNA and cyclin D1 (Fig. 2f). These results suggested that circ-Ntrk2 knockdown can inhibit pulmonary vascular remodelling and alleviate PAH. In vitro, CCK8 and EdU results indicated that the proliferation ability of PASMCs increased under HYP, which was inhibited after silencing circ-Ntrk2 and enhanced after overexpression of circ-Ntrk2 (Fig. 2g and h). These results indicated that circ-Ntrk2 knockdown inhibits PASMCs proliferation in vitro and reverses pulmonary vascular remodelling to alleviate PAH in vivo.

Circ-Ntrk2 knockdown alleviates pulmonary vascular remodelling and improves PAH. a Construction of small interfering RNA (siRNA) based on the target sequence. All treatment groups received injections twice weekly for modelling for 21 days. b Echocardiographic measurements and images illustrating pulmonary artery acceleration time (PAAT) and pulmonary arterial velocity time integral (PAVTI) in the NOR, HYP, HYP + SI-NC, and HYP + SI groups are shown (n = 5). c Right ventricular systolic pressure (RVSP) and the right ventricle (RV)/left ventricle (LV) + septum (S) weight ratio in the NOR, HYP, HYP + SI-NC, and HYP + SI groups (n = 5). d Morphological analysis of the pulmonary artery was performed using H&E staining and immunofluorescence (n = 5). e Detection of collagen deposition around blood vessels using Masson staining (up) (n = 5) and H&E staining of myocardial cells in right ventricular wall (bottom) (n = 5). f Analysis of PCNA (proliferating cell nuclear antigen) and cyclin D1 expression in lung tissues (n = 5) and PASMCs (n = 6). The gel or blot images have been cropped and the complete images have been submitted separately (Additional file 4). g EdU in PASMCs (n = 6). h CCK-8 assay in PASMCs (n = 6). All values are presented as the mean ± SD. *P < 0.05, **P < 0.01
Circ-Ntrk2 acts as a sponge for miR-296-5p
To verify the existence of the circRNA-miRNA axis under the specific pathological conditions of PASMCs, we predicted miRNAs that were probably adsorbed by circ-Ntrk2 using the bioinformatics software miRanda (http://www.microrna.org/microrna/home.do) (Fig. 3a) and detected their expression by qPCR. Under hypoxic conditions, miR-296-5p expression was notably downregulated both in vivo and in vitro (Fig. 3b). PASMCs were transfected with circ-Ntrk2 siRNA and then exposed to HYP for 48 h. Moreover, under the same conditions, the downregulation of miRNA expression was reversed after the knockdown of circ-Ntrk2, consistent with the competitive endogenous RNA mechanism (Fig. 3c); we therefore used miR-296-5p in subsequent analyses. To verify whether circ-Ntrk2 binds to miR-296-5p, we performed FISH and showed that circ-Ntrk2 could co-localise with miR-296-5p in the cytoplasm of PASMCs (Fig. 3d). We then predicted that there were three possible target sequence regions between circ-Ntrk2 and miR-296-5p using TargetScan (http://www.targetscan.org/) (Fig. 3e). Subsequently, we constructed dual luciferase vector plasmids and found that the ratio of firefly to Renilla luciferase for wild-type and target sites (position 553–573) without mutant circ-Ntrk2 was visibly decreased compared with those of the NC, whereas those of the target site with mutant circ-Ntrk2 did not change (Fig. 3f). These results indicated that circ-Ntrk2 could act as a sponge for miR-296-5p.

Circ-Ntrk2 acts as a sponge for miR-296-5p. a miRanda bioinformatics software predicted miRNAs that could bind to circ-Ntrk2. b Quantitative polymerase chain reaction (qPCR) analysis of miR-296-5p expression in mouse lung tissues and PASMCs (n = 6). c PASMCs were transfected with si-circ-Ntrk2, and qPCR was used to detect miR-296-5p expression (n = 6). d Fluorescence in situ hybridization (FISH) was used to observe the colocalization of circ-Ntrk2 and miR-296-5p. e TargetScan predicted binding sites and secondary structures of circ-Ntrk2 and miR-296-5p. f Dual luciferase vector and mutant plasmids were designed based on binding sites between circ-Ntrk2 and miR-296-5p. Dual luciferase assays were used to validate the interactions between circ-Ntrk2 and miR-296-5p (n = 6). All values are presented as the mean ± SD. *P < 0.05, **P < 0.01
MiR-296-5p overexpression alleviated pulmonary vascular remodelling and improved PAH
To determine whether miR-296-5p plays a role in PAH, we validated its function both in vivo and in vitro. Echocardiography and right cardiac pressure showed that miR-296-5p supplementation markedly improved pulmonary arterial pressure under hypoxic condition (Fig. 4a–c, Additional file 1: Fig. S1e). H&E staining and immunofluorescence showed that HYP-induced pulmonary vascular remodelling was alleviated by miR-296-5p addition (Fig. 4d). Masson staining showed that miR-296-5p could alleviate HYP-induced collagen deposition around the pulmonary artery, and cardiac H&E staining suggested that miR-296-5p could modify pathological changes in HYP-induced cardiac hypertrophy (Fig. 4e). Western blotting verified that miR-296-5p could reverse HYP-induced upregulation of proliferation-related proteins, such as PCNA and cyclin D1 (Fig. 4f). In vitro, the results of CCK8 and EdU experiments showed that the miR-296-5p mimic decreased the EdU incorporation rate and CCK8 value under HYP (Fig. 4g and h). These results suggested that miR-296-5p overexpression inhibits HYP-induced proliferation of PASMCs. Taken together, miR-296-5p inhibits HYP-induced PASMC proliferation to alleviate pulmonary vascular remodelling and improve PAH.

MiR-296-5p overexpression alleviates pulmonary vascular remodelling and improves PAH. a Agomir constructed for miR-296-5p was used to upregulate miR-296-5p. All treatment groups received injections twice weekly for modelling for 21 days. b Echocardiographic measurements and images illustrating pulmonary artery acceleration time (PAAT) and pulmonary arterial velocity time integral (PAVTI) in the NOR, HYP, HYP + Agomir-NC, and HYP + Agomir groups are shown (n = 5). c Right ventricular systolic pressure (RVSP) and the right ventricle (RV)/left ventricle (LV) + septum (S) weight ratio in the NOR, HYP, HYP + Agomir-NC, and HYP + Agomir groups (n = 5). d Analysis of pulmonary artery remodelling was performed using H&E staining and immunofluorescence (n = 5). e Detection of collagen deposition around blood vessels using Masson staining (up) (n = 5) and H&E staining of myocardial cells in right ventricular wall (bottom) (n = 5). f Analysis of PCNA (proliferating cell nuclear antigen) and cyclin D1 expression in the lung tissues (n = 5) and PASMCs (n = 6). The gel or blot images have been cropped and the complete images have been submitted separately (Additional file 4). g EdU in PASMCs (n = 6). h CCK-8 assay in PASMCs (n = 6). All values are presented as the mean ± SD. *P < 0.05, **P < 0.01
Circ-Ntrk2 sponges miR-296-5p to promote pulmonary vascular remodelling and PAH
To further confirm whether circ-Ntrk2 affected PAH through miR-296-5p, we treated cells with circRNA and miRNA in vitro and in vivo and detected the phenotypic and pathological changes. Echocardiography and right cardiac pressure showed that the moderating effect of circ-Ntrk2 knockdown on pulmonary artery pressure under HYP was reversed by the miR-296-5p antagomir (Fig. 5a–c, Additional file 1: Fig. S1f). Similarly, H&E staining and immunofluorescence showed that the alleviating effect of circ-Ntrk2 knockdown on HYP-induced pulmonary vascular remodelling was also reversed by the miR-296-5p antagomir (Fig. 5d). Moreover, cardiac H&E staining suggested that circ-Ntrk2 knockdown could correct the pathological changes in HYP-induced myocardial hypertrophy and could be inhibited after exogenous supplementation with miR-296-5p antagomir. Masson’s trichrome staining showed a similar trend in collagen deposition (Fig. 5e). Consistently, miR-296-5p reversed the regulation of circ-Ntrk2 on the expression of PCNA and cyclin D1 under HYP (Fig. 5f). In in vitro recovery experiments, including EdU and CCK8, results showed that circ-Ntrk2 siRNA inhibited the HYP-induced proliferation of PASMCs, whereas the miR-296-5p inhibitor reversed this effect (Fig. 5g and h). Circ-Ntrk2 could promote PASMC proliferation and pulmonary vascular remodelling to regulate PAH progression by adsorbing miR-296-5p.

Circ-Ntrk2 sponges miR-296-5p to promote pulmonary vascular remodelling and PAH. a All treatment groups received si-circ-Ntrk2 or miR-296-5p antagomir injections twice weekly for modelling for 21 days. b Echocardiographic measurements and images illustrating pulmonary artery acceleration time (PAAT) and pulmonary arterial velocity time integral (PAVTI) in the NOR, HYP, HYP + SI-NC + ANT-NC, HYP + SI + ANT-NC, and HYP + SI + ANT groups are shown (n = 5). c Right ventricular systolic pressure (RVSP) and the right ventricle (RV)/left ventricle (LV) + septum (S) weight ratio in the NOR, HYP, HYP + SI-NC + ANT-NC, HYP + SI + ANT-NC, and HYP + SI + ANT groups (n = 5). d Analysis of pulmonary artery remodelling was performed using H&E and immunofluorescence. e Detection of collagen deposition around blood vessels using Masson staining (up) (n = 5) and H&E staining of myocardial cells in right ventricular wall (bottom) (n = 5). f Analysis of PCNA (proliferating cell nuclear antigen) and cyclin D1 expression in the lung tissues (n = 5) and PASMCs (n = 6). The gel or blot images have been cropped and the complete images have been submitted separately (Additional file 4). g EdU in PASMCs (n = 6). h CCK-8 assay in PASMCs (n = 6). All values are presented as the mean ± SD. *P < 0.05, **P < 0.01
Circ-Ntrk2 sponges miR-296-5p to activate the TGF-β1/p38 MAPK pathway and promotes pulmonary vascular remodelling and PAH
Bioinformatics software was used to predict the possible target proteins of miR-296-5p (Fig. 6a). Our study revealed that TGF-β1 expression was significantly increased under HYP, and its upregulation was inhibited in circ-Ntrk2 knockdown cells and that p38 MAPK showed similar changes (Fig. 6b). Therefore, it was selected for follow-up research. Dual luciferase reporter assays showed that the ratio of firefly to Renilla luciferase activity in cells expressing wild-type TGF-β1 was distinctly lower than that in the NC; however, there were no significant changes in cells transfected with a binding site mutant TGF-β1 (Fig. 6c and d). Western blotting showed that transfection with miR-296-5p mimics inhibited TGF-β1 and p38 MAPK expression, compared with that in NC. Contrasting results were obtained after transfection with the inhibitor in HYP-induced smooth muscle cells (Fig. 6e). In further experiments, we found that circ-Ntrk2 knockdown could inhibit TGF-β1 and p38 MAPK expression under HYP; however, this effect was reversed after miR-296-5p downregulation (Fig. 6f). Altogether, these results demonstrated that antagonising circ-Ntrk2 could alleviate PAH by inhibiting PASMC proliferation and reversing pulmonary vascular remodelling through the miR-296-5p/TGF-β1/p38 MAPK signal transduction axis (Fig. 7).

Circ-Ntrk2 might promote PAH through the miR-296-5p/TGF-β1 axis. a TargetScan, miRDB, miRTarBase, and miRWalk bioinformatics software predicted miR-296-5p target proteins. b Western blotting was used to verify the expression of TGF-β1 and p38 MAPK in lung tissues (n = 5) and PASMCs (n = 6) from the NOR, HYP, HYP + SI-NC, and HYP + SI groups. c TargetScan predicted binding sites and secondary structures of miR-296-5p and TGF-β1. d Dual luciferase vector and mutant plasmids were designed based on binding sites between miR-296-5p and TGF-β1. Dual luciferase assays were used to validate the interactions between miR-296-5p and TGF-β1 (n = 6). e Analysis expression of TGF-β1 and p38 MAPK in lung tissues (n = 5) and PASMCs (n = 6) after intervention of miR-296-5p. f Analysis expression of TGF-β1 and p38 MAPK in lung tissues (n = 5) and PASMCs (n = 6) after co-intervention of miR-296-5p and circ-Ntrk2. The gel or blot images have been cropped and the complete images have been submitted separately (Additional file 4). All values are presented as the mean ± SD. *P < 0.05, **P < 0.01

Potential molecular mechanisms of circ-Ntrk2 in PAH. Circ-Ntrk2 functions as an accelerator of PAH by sponging miR-296-5p to activate the TGF-β1/p38 MAPK pathway, and consequently promotes PASMC proliferation and pulmonary artery remodelling
Discussion
In this study, we identified a novel circRNA named circ-Ntrk2 and evaluated its role and mechanism in PAH pathogenesis. First, circ-Ntrk2 was upregulated in a mouse model of PAH in the lung tissue. Second, silencing circ-Ntrk2 inhibited HYP-induced PASMC proliferation and reversed HYP-induced PAH in mice. Third, circ-Ntrk2 acted as a molecular sponge for miR-296-5p to regulate the TGF-β1/p38 MAPK pathway. Overall, our results indicate that circ-Ntrk2 can sponge miR-296-5p to activate the TGF-β1/p38 MAPK signalling axis to promote the progression of PAH and pulmonary vascular remodelling.
PAH is a heterogeneous disease characterised by pulmonary vascular remodelling, which often leads to death from right heart failure [19, 20]. PASMC proliferation and endothelial cell dysfunction are involved in PAH occurrence and development, with the former playing a core pathological role [21,22,23]. Current research suggests that PAH cannot be completely cured due to the lack of drugs that can effectively reverse pulmonary vascular remodelling [24,25,26].
NcRNA is a class of transcribed RNA with no or weak protein-coding ability, mainly including circRNAs, long ncRNAs (lncRNAs), and miRNAs [5]. Numerous studies have shown that ncRNAs play a vital role in regulating human diseases, such as PAH [27, 28]. CircRNAs, as a novel class of regulatory ncRNA, showed insensitivity to RNase R and were more stable than lncRNA and miRNA [11,12,13]. Due to their abundance, species preservation, tissue specificity, stability, and regulatory functional roles, circRNAs have been identified as potential biomarkers and therapeutic targets for various diseases, including PAH [13,14,15, 29, 30]. However, most circRNAs have not been thoroughly verified experimentally, especially in PAH [31,32,33]. Previous studies have shown that circRNAs play multiple important roles in cellular physiology by acting as miRNA sponges, templates for protein translation, transcriptional regulators, and RBP-binding molecules [34], with sponge mechanisms being the most widely studied [30]. Jiang et al. [17] identified a novel circRNA named circ-calm4, demonstrating its ability to promote PASMC proliferation by sponging miR-337-3p and regulating HYP-induced PASMC pyroptosis by sponging miR-124-3p. Lu et al. [35] reported that circ-SMOC1 inhibits metabolic reprogramming in PAH by sponging miR-329-3p. Huang et al. [36] found that hsa_circ_0016070 could accelerate PAH progression by targeting miR-340-5p. These results provide strong evidence that circRNAs can regulate PAH by sponging miRNAs and suggest that their role in PAH is worth further research and investment. In this study, we identified a novel functional circRNA localised in the cytosol of PASMCs using high-throughput sequencing, Sanger sequencing, and FISH and named it “circ-Ntrk2”. It was confirmed that circ-Ntrk2 knockdown could prominently inhibit PASMCs proliferation and relieve pulmonary vascular remodelling and right ventricular hypertrophy in vivo and in vitro.
We further predicted the miRNAs targeted by circ-Ntrk2 through bioinformatics analysis and showed that circ-Ntrk2 could directly target miR-296-5p. Studies have shown that miR-296-5p is involved in various diseases. Li et al. [37, 38] reported that miR-296-5p regulates gastric cancer cell proliferation. Shi et al. [39] reported that miR-296-5p overexpression inhibits hepatocellular carcinoma progression. However, changes in the expression and role of miR-296-5p in PAH have not been confirmed. Our study is the first to demonstrate that miR-296-5p is downregulated in hypoxic PAH and that its overexpression inhibits PASMC proliferation and mediates the regulatory mechanism of circ-Ntrk2 in PAH.
The important role of the TGF pathway in PAH has been well established [40,41,42,43]. Cai et al. [44] reported that TGF-β1 activation could induce excessive PASMC proliferation and reduce apoptosis through STAT3, thereby driving pulmonary vascular remodelling. Moreover, p38 MAPK is widely recognised as a negative factor in PAH [45, 46], and the presence of TGF-β1/p38 MAPK pathway has been reported in numerous diseases, including PAH [47,48,49]. Therefore, we further investigated whether the TGF-β1/p38 MAPK pathway is involved in the process by which circ-Ntrk2 affects PAH. Interestingly, the bioinformatic analysis showed that TGF-β1 is a potential downstream target of miR-296-5p. We also demonstrated the targeted regulatory relationship between miR-296-5p and TGF-β1 and found that HYP-induced TGF-β1 and p38 MAPK upregulation was inhibited by circ-Ntrk2 knockdown, while miR-296-5p supplementation reversed this change. In conclusion, our study showed that antagonising circ-Ntrk2 could alleviate PAH by inhibiting PASMC proliferation and reversing pulmonary vascular remodelling through the miR-296-5p/TGF-β1/p38 MAPK signal transduction axis.
Our study identified a new circRNA (circ-Ntrk2) and further explored its function and mechanism, which is an important addition to circRNA research in the field of PAH. However, because of the complexity of the circRNA regulatory network, only the representative network was identified in this study. Whether another circ-Ntrk2-miRNA-mRNA regulatory axis exists under our pathological conditions will be the direction of our future studies.
Conclusions
In this study, we demonstrated that circ-Ntrk2 expression is upregulated in hypoxic lung tissues and PASMCs and that its deficiency could improve pulmonary vascular remodelling and relieve PAH via the miR-296-5p/TGF-β1/p38 MAPK signal transduction axis, thus establishing potential targets for PAH diagnosis and treatment. Furthermore, our study contributes to the understanding of circRNA in relation to PAH.
Availability of data and materials
All relevant data and materials are stored at the Key Laboratory of Heart and Lung of Wenzhou Medical University and are available from the first author and corresponding author on reasonable request.
Abbreviations
- CircRNA:
-
Circular RNA
- ncRNA:
-
Non-coding RNA
- miRNA:
-
MicroRNA
- PAH:
-
Pulmonary arterial hypertension
- PASMCs:
-
Pulmonary artery smooth muscle cells
- PBS:
-
Phosphate-buffered saline
- PCNA:
-
Proliferating cell nuclear antigen
- TGF-β1:
-
Transforming growth factor-β1
- NC:
-
Negative control
- NOR:
-
Normoxia
- HYP:
-
Hypoxia
- siRNA:
-
Small interfering RNA
- mut:
-
Mutant
- RV:
-
Right ventricle
- RVSP:
-
Right ventricular systolic pressure
- S:
-
Septum
- LV:
-
Left ventricle
- H&E:
-
Hematoxylin–eosin
- WT:
-
Wall thickness
- TT:
-
Total thickness
- WA:
-
Pulmonary artery wall area
- PAAT:
-
Pulmonary artery acceleration time
- PAVTI:
-
Pulmonary artery velocity time integral
- gDNA:
-
Genomic DNA
- FISH:
-
Fluorescence in situ hybridization
- CCK-8:
-
Cell counting kit-8
- EdU:
-
5-Ethynyl-20-deoxyuridine
- qRT-PCR:
-
Quantitative real-time polymerase chain reaction
- DAPI:
-
4,6-Diamidino-2-phenylindole
References
Thenappan T, Ormiston ML, Ryan JJ, Archer SL. Pulmonary arterial hypertension: pathogenesis and clinical management. BMJ. 2018;360: j5492.
Lechartier B, Berrebeh N, Huertas A, Humbert M, Guignabert C, Tu L. Phenotypic diversity of vascular smooth muscle cells in pulmonary arterial hypertension: implications for therapy. Chest. 2022;161:219–31.
Hassoun PM. Pulmonary arterial hypertension. N Engl J Med. 2021;385:2361–76.
Ruopp NF, Cockrill BA. Diagnosis and treatment of pulmonary arterial hypertension: a review. JAMA. 2022;327:1379–91.
Slack FJ, Chinnaiyan AM. The role of non-coding RNAs in oncology. Cell. 2019;179:1033–55.
Qu S, Yang X, Li X, Wang J, Gao Y, Shang R, Sun W, Dou K, Li H. Circular RNA: a new star of noncoding RNAs. Cancer Lett. 2015;365:141–8.
Wilusz JE, Sharp PA. Molecular biology. A circuitous route to noncoding RNA. Science. 2013;340:440–1.
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen B, Luo Y, Lyu D, Li Y, Shi G, Liang L, Gu J, He X, Huang S. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat Commun. 2016;7:11215.
Hansen TB, Jensen TI, Clausen BH, Bramsen JB, Finsen B, Damgaard CK, Kjems J. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495:384–8.
Li H, Xu J-D, Fang X-H, Zhu J-N, Yang J, Pan R, Yuan S-J, Zeng N, Yang Z-Z, Yang H, Wang X-P, Duan J-Z, Wang S, Luo J-F, Wu S-L, Shan Z-X. Circular RNA circRNA_000203 aggravates cardiac hypertrophy via suppressing miR-26b-5p and miR-140-3p binding to Gata4. Cardiovasc Res. 2020;116:1323–34.
Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer M, Loewer A, Ziebold U, Landthaler M, Kocks C, le Noble F, Rajewsky N. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495:333–8.
Werfel S, Nothjunge S, Schwarzmayr T, Strom T-M, Meitinger T, Engelhardt S. Characterization of circular RNAs in human, mouse and rat hearts. J Mol Cell Cardiol. 2016;98:103–7.
Jeck WR, Sharpless NE. Detecting and characterizing circular RNAs. Nat Biotechnol. 2014;32:453–61.
Xia S, Feng J, Lei L, Hu J, Xia L, Wang J, Xiang Y, Liu L, Zhong S, Han L, He C. Comprehensive characterization of tissue-specific circular RNAs in the human and mouse genomes. Brief Bioinform. 2017;18:984–92.
Wang J, Zhu M, Pan J, Chen C, Xia S, Song Y. Circular RNAs: a rising star in respiratory diseases. Respir Res. 2019;20:3.
Zhang J, Li Y, Qi J, Yu X, Ren H, Zhao X, Xin W, He S, Zheng X, Ma C, Zhang L, Wu B, Zhu D. Circ- calm4 serves as an sponge to regulate Myo10 (Myosin 10) and promote pulmonary artery smooth muscle proliferation. Hypertension. 2020;75:668–79.
Jiang Y, Liu H, Yu H, Zhou Y, Zhang J, Xin W, Li Y, He S, Ma C, Zheng X, Zhang L, Zhao X, Wu B, Jiang C, Zhu D. Circular RNA Calm4 regulates hypoxia-induced pulmonary arterial smooth muscle cells pyroptosis via the Circ-Calm4/miR-124-3p/PDCD6 axis. Arterioscler Thromb Vasc Biol. 2021;41:1675–93.
Huang X, Wu P, Huang F, Xu M, Chen M, Huang K, Li G-P, Xu M, Yao D, Wang L. Baicalin attenuates chronic hypoxia-induced pulmonary hypertension via adenosine A receptor-induced SDF-1/CXCR4/PI3K/AKT signaling. J Biomed Sci. 2017;24:52.
Maron BA, Abman SH, Elliott CG, Frantz RP, Hopper RK, Horn EM, Nicolls MR, Shlobin OA, Shah SJ, Kovacs G, Olschewski H, Rosenzweig EB. Pulmonary arterial hypertension: diagnosis, treatment, and novel advances. Am J Respir Crit Care Med. 2021;203:1472–87.
Rhodes CJ, Sweatt AJ, Maron BA. Harnessing big data to advance treatment and understanding of pulmonary hypertension. Circ Res. 2022;130:1423–44.
Crosswhite P, Sun Z. Molecular mechanisms of pulmonary arterial remodeling. Mol Med. 2014;20:191–201.
Leopold JA, Maron BA. Molecular mechanisms of pulmonary vascular remodeling in pulmonary arterial hypertension. Int J Mol Sci. 2016;17:761.
Veith C, Schermuly RT, Brandes RP, Weissmann N. Molecular mechanisms of hypoxia-inducible factor-induced pulmonary arterial smooth muscle cell alterations in pulmonary hypertension. J Physiol. 2016;594:1167–77.
Cheng F, Loscalzo J. Pulmonary comorbidity in lung cancer. Trends Mol Med. 2018;24:239–41.
Chan SY, Loscalzo J. Pathogenic mechanisms of pulmonary arterial hypertension. J Mol Cell Cardiol. 2008;44:14–30.
Brown LM, Chen H, Halpern S, Taichman D, McGoon MD, Farber HW, Frost AE, Liou TG, Turner M, Feldkircher K, Miller DP, Elliott CG. Delay in recognition of pulmonary arterial hypertension: factors identified from the REVEAL Registry. Chest. 2011;140:19–26.
Zhou G, Chen T, Raj JU. MicroRNAs in pulmonary arterial hypertension. Am J Respir Cell Mol Biol. 2015;52:139–51.
Deng L, Han X, Wang Z, Nie X, Bian J. The landscape of noncoding RNA in pulmonary hypertension. Biomolecules. 2022;12:796.
Chen L-L. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat Rev Mol Cell Biol. 2020;21:475–90.
Ali MK, Schimmel K, Zhao L, Chen C-K, Dua K, Nicolls MR, Spiekerkoetter E. The role of circular RNAs in pulmonary hypertension. Eur Respir J. 2022;60:2200012.
Sun X, Chen R, Yao X, Zheng Z, Wang C, Chen J, Cheng J. circGSAP: a new clinical biomarker for idiopathic pulmonary hypertension? Am J Respir Crit Care Med. 2022;205:252–3.
Wang J, Zhu M-C, Kalionis B, Wu J-Z, Wang L-L, Ge H-Y, Chen C-C, Tang X-D, Song Y-L, He H, Xia S-J. Characteristics of circular RNA expression in lung tissues from mice with hypoxia-induced pulmonary hypertension. Int J Mol Med. 2018;42:1353–66.
Yuan P, Wu W-H, Gong S-G, Jiang R, Zhao Q-H, Pudasaini B, Sun Y-Y, Li J-L, Liu J-M, Wang L. Impact of circGSAP in peripheral blood mononuclear cells on idiopathic pulmonary arterial hypertension. Am J Respir Crit Care Med. 2021;203:1579–83.
Kristensen LS, Andersen MS, Stagsted LVW, Ebbesen KK, Hansen TB, Kjems J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. 2019;20:675–91.
Lu G-F, Geng F, Deng L-P, Lin D-C, Huang Y-Z, Lai S-M, Lin Y-C, Gui L-X, Sham JSK, Lin M-J. Reduced CircSMOC1 level promotes metabolic reprogramming via PTBP1 (Polypyrimidine Tract-Binding Protein) and miR-329–3p in pulmonary arterial hypertension rats. Hypertension. 2022:101161HYPERTENSIONAHA12219183.
Huang C-X, Jiang Z-X, Du D-Y, Zhang Z-M, Liu Y, Li Y-T. Hsa_circ_0016070/micro-340-5p axis accelerates pulmonary arterial hypertension progression by upregulating TWIST1 transcription via TCF4/β-catenin complex. J Am Heart Assoc. 2022;11: e024147.
Li T, Lu YY, Zhao XD, Guo HQ, Liu CH, Li H, Zhou L, Han YN, Wu KC, Nie YZ, Shi YQ, Fan DM. MicroRNA-296-5p increases proliferation in gastric cancer through repression of Caudal-related homeobox 1. Oncogene. 2014;33:783–93.
Rong D, Lu C, Zhang B, Fu K, Zhao S, Tang W, Cao H. CircPSMC3 suppresses the proliferation and metastasis of gastric cancer by acting as a competitive endogenous RNA through sponging miR-296-5p. Mol Cancer. 2019;18:25.
Shi D-M, Li L-X, Bian X-Y, Shi X-J, Lu L-L, Zhou H-X, Pan T-J, Zhou J, Fan J, Wu W-Z. miR-296-5p suppresses EMT of hepatocellular carcinoma via attenuating NRG1/ERBB2/ERBB3 signaling. J Exp Clin Cancer Res. 2018;37:294.
Guignabert C, Humbert M. Targeting transforming growth factor-β receptors in pulmonary hypertension. Eur Respir J. 2021;57:2002341.
Long L, Crosby A, Yang X, Southwood M, Upton PD, Kim D-K, Morrell NW. Altered bone morphogenetic protein and transforming growth factor-beta signaling in rat models of pulmonary hypertension: potential for activin receptor-like kinase-5 inhibition in prevention and progression of disease. Circulation. 2009;119:566–76.
Davies RJ, Holmes AM, Deighton J, Long L, Yang X, Barker L, Walker C, Budd DC, Upton PD, Morrell NW. BMP type II receptor deficiency confers resistance to growth inhibition by TGF-β in pulmonary artery smooth muscle cells: role of proinflammatory cytokines. Am J Physiol Lung Cell Mol Physiol. 2012;302:L604–15.
Kudryashova TV, Shen Y, Pena A, Cronin E, Okorie E, Goncharov DA, Goncharova EA. Inhibitory antibodies against activin A and TGF-β reduce self-supported, but not soluble factors-induced growth of human pulmonary arterial vascular smooth muscle cells in pulmonary arterial hypertension. Int J Mol Sci. 2018;19:2957.
Cai Z, Li J, Zhuang Q, Zhang X, Yuan A, Shen L, Kang K, Qu B, Tang Y, Pu J, Gou D, Shen J. MiR-125a-5p ameliorates monocrotaline-induced pulmonary arterial hypertension by targeting the TGF-β1 and IL-6/STAT3 signaling pathways. Exp Mol Med. 2018;50:1.
Kojonazarov B, Novoyatleva T, Boehm M, Happe C, Sibinska Z, Tian X, Sajjad A, Luitel H, Kriechling P, Posern G, Evans SM, Grimminger F, Ghofrani HA, Weissmann N, Bogaard HJ, Seeger W, Schermuly RT. p38 MAPK inhibition improves heart function in pressure-loaded right ventricular hypertrophy. Am J Respir Cell Mol Biol. 2017;57:603–14.
Vanderpool RR, Tang H, Rischard F, Yuan JXJ. Is p38 MAPK a dark force in right ventricular hypertrophy and failure in pulmonary arterial hypertension? Am J Respir Cell Mol Biol. 2017;57:506–8.
Jia D, He Y, Zhu Q, Liu H, Zuo C, Chen G, Yu Y, Lu A. RAGE-mediated extracellular matrix proteins accumulation exacerbates HySu-induced pulmonary hypertension. Cardiovasc Res. 2017;113:586–97.
Sheares KKK, Jeffery TK, Long L, Yang X, Morrell NW. Differential effects of TGF-beta1 and BMP-4 on the hypoxic induction of cyclooxygenase-2 in human pulmonary artery smooth muscle cells. Am J Physiol Lung Cell Mol Physiol. 2004;287:L919–27.
Zhang L, Li Y, Chen M, Su X, Yi D, Lu P, Zhu D. 15-LO/15-HETE mediated vascular adventitia fibrosis via p38 MAPK-dependent TGF-β. J Cell Physiol. 2014;229:245–57.
Acknowledgements
The authors thank the Scientific Research Center of Wenzhou Medical University and Medical Research Center of the First Affiliated Hospital of Wenzhou Medical University for assistance.
Funding
This work was supported by the Chinese National Natural Science Foundation [82170061, 81873411], and Basic scientific research Project of Wenzhou [H20210010].
Author information
Authors and Affiliations
Contributions
The research design was completed by XYH, JW and DY. LHS, XCL and XLM performed experiments and co-wrote this manuscript. Statistical analysis of the data was performed by TTX, YYZ, and SNL. XYZ and LXW optimized the manuscript to ensure that it adhered to the journal guidelines. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All animal procedures were approved by the Animal Ethics Committee of Wenzhou Medical University (wydw2021-0257). All animal experiments complied with the ARRIVE guidelines and were conducted in accordance with the U.K. Animals (Scientific Procedures) Act, 1986 and associated guidelines, EU Directive 2010/63/EU for animal experiments, or the National Research Council's Guide for the Care and Use of Laboratory Animals. All animals were executed using anaesthesia with CO2 and sacrificed at the end of study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Measurement of arterial pressure in mice and detection of drug knockout efficiency.
Additional file 2.
The primary antibodies used in this study.
Additional file 3.
The primers used in this study.
Additional file 4.
The gel or blot images.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Su, L., Li, X., Mao, X. et al. Circ-Ntrk2 acts as a miR-296-5p sponge to activate the TGF-β1/p38 MAPK pathway and promote pulmonary hypertension and vascular remodelling. Respir Res 24, 78 (2023). https://doi.org/10.1186/s12931-023-02385-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12931-023-02385-7