
Abstract
Background
A lack of clear trial evidence often hampers clinical decision-making during support of the preterm lung at birth. Protein biomarkers have been used to define acute lung injury phenotypes and improve patient selection for specific interventions in adult respiratory distress syndrome. The objective of the study was to use proteomics to provide a deeper biological understanding of acute lung injury phenotypes resulting from different aeration strategies at birth in the preterm lung.
Methods
Changes in protein abundance against an unventilated group (n = 7) were identified via mass spectrometry in a biobank of gravity dependent and non-dependent lung tissue from preterm lambs managed with either a Sustained Inflation (SI, n = 20), Dynamic PEEP (DynPEEP, n = 19) or static PEEP (StatPEEP, n = 11). Ventilation strategy-specific pathways and functions were identified (PANTHER and WebGestalt Tool) and phenotypes defined using integrated analysis of proteome, physiological and clinical datasets (MixOmics package).
Results
2372 proteins were identified. More altered proteins were identified in the non-dependent lung, and in SI group than StatPEEP and DynPEEP. Different inflammation, immune system, apoptosis and cytokine pathway enrichment were identified for each strategy and lung region. Specific integration maps of clinical and physiological outcomes to specific proteins could be generated for each strategy.
Conclusions
Proteomics mapped the molecular events initiating acute lung injury and identified detailed strategy-specific phenotypes. This study demonstrates the potential to characterise preterm lung injury by the direct aetiology and response to lung injury; the first step towards true precision medicine in neonatology.
Similar content being viewed by others
Background
Bronchopulmonary dysplasia (BPD) remains a serious chronic complication of preterm birth, with potentially life-long consequences. BPD arises from acute injurious events occurring in the under-developed preterm lung, often after ventilation begins at birth [1, 2]. This includes inappropriate ventilatory choices aimed at supporting the respiratory distress syndrome (RDS) characteristic of preterm birth [3]. The complex and dynamic cascade of injury is unlikely to be explained by a singular biological or mechanical cause. Yet, trials of preterm respiratory therapies are generally premised on a singular, unified treatment model, and have generally failed to show any impact on BPD rates [4, 5]. The characterisation of acute lung injury and BPD is also often reliant on imprecise clinical and diagnostic criteria, such as oxygen dependency [6]. This approach fails to appreciate the complex interaction between intrinsic and developmental factors and clinical interventions that shapes the development and progression from acute lung injury to BPD, nor does it enable a biological understanding of the individual patients response to treatment.
A possible solution has been explored in adults with acute respiratory distress syndrome (ARDS); using physiology, clinical data, biomarkers to stratify patients into distinct, homogenous subgroups [7]. This approach has identified ARDS phenotypes associated with diverging inflammatory responses [8,9,10] that indicate different benefit response to specific therapies, such as lung recruitment [11]. Considering ARDS phenotypes in this way provides insight into the variable outcomes from ARDS trials, and allows the potential for targeted interventions [8]. Like ARDS, the preterm RDS population exhibits a high degree of clinical and biological heterogeneity, suggesting that this population may similarly benefit from identification of biomarker-based phenotypes.
Effectively achieving lung aeration at birth is fundamental to establishing a functional residual capacity, and failing to so do correlates to the earliest point in which respiratory management can initiate lung injury [12, 13]. Various approaches that support aeration in preterm infants have been suggested targeting different physiological rationales, such as positive end-expiratory pressure (PEEP) strategies, or the use of an initial sustained lung inflation (SI) [12, 14,15,16]. It has been proposed that the different clinical and injury responses seen with these approaches reflect different injury mechanistic pathways within the lung, and thus differing phenotypes which are as yet undefined.
We have established a large biobank of lung tissue, clinical and lung imaging and mechanics data in preterm lambs. We have also previously identified protein-based phenotypes associated with gestational age [17] and duration of ventilation [18] in this biobank. Thus, the primary aim of this study was to use proteomics to provide a deeper biological understanding of acute lung injury phenotypes resulting from different aeration strategies at birth in the preterm lung with RDS. This aim will be achieved by comparing the phenotypic response to different PEEP strategies at birth in the preterm lamb. In secondary studies, we aimed to use partial least squares modelling to identify phenotype specific protein-function associations within non-dependent and dependent lung regions. By doing so we aim to design the first phenotypic approach to defining the initiation of preterm lung disease. This would provide the mechanistic foundation for refining lung protective decisions following preterm birth to the most appropriate patient or intervention-based group, and potentially provide the precision needed to meaningfully reduce the incidence of BPD [3].
Methods
This study used historic samples and data from our biobank. The measurements, physiological data acquisition and analysis and lung injury analysis methodology from these studies have been described previously [12, 19, 20]. Figure 1 provides a summary of the experimental workflow and setup relevant to the study aims. All data used in this study is available at Figshare (https://doi.org/10.26188/19252103.v1).

Experimental setup and workflow for comparison of RM-phenotypes in non-dependent (ND) and dependent (D) lung tissue (Main box). The two-step normalization workflow [50] was performed prior to data analysis (Inset box). Blue shading represents StatPEEP group, red–orange SI group and yellow-green DynPEEP. Software analysis packages highlighted in pink
Experimental instrumentation
Preterm lambs (124–127d; term ~ 145d) born by caesarean section to date-mated, betamethasone-treated ewes were instrumented before delivery [12, 19, 20].
Recruitment strategies
Before delivery, lambs were randomly assigned to receive one of the following lung recruitment strategies from birth.
Static PEEP (StatPEEP)
Positive-pressure ventilation (PPV; SLE5000, SLE Ltd, South Croydon, UK) in volume-targeted ventilation (VTV) mode (PEEP 8 cmH2O [21], inspiratory time 0.4 s, rate 60 inflations/min, set VT 7 mL/kg and maximum peak inspiratory pressure (PIP) 35 cmH2O).
Sustained inflation (SI)
SI was maintained at 35–40 cmH2O until 10 s after achievement of a volume plateau on the EIT display (maximum 180 s) [12, 19, 20, 22].
Dynamic PEEP (DynPEEP)
Commencing at 6 cmH2O during PPV + VTV (VT, 7 ml/kg; maximum PIP 35 cmH2O), PEEP increased by 2 cmH2O every 10–15 inflations during PPV to a maximum PEEP (14–20 cmH2O) based upon dynamic compliance (Cdyn) response, and then similarly decreased to a final PEEP of 8 cmH2O (170–180 s total duration) [12, 19].
On completion of the SI or DynPEEP, PPV + VTV was continued as per the StatPEEP group for 90 min [12, 19, 20]. All lambs commenced PPV using an inspired oxygen fraction (FiO2) of 0.3. FiO2 and delivered VT were adjusted to maintain peripheral oxygen saturation between 88–94% and a partial pressure of carbon dioxide (PaCO2) of 40–60 mmHg using a standardized strategy [12, 19, 23]. Poractant alfa 200 mg/kg (Chiesi Farmaceutici SpA) was administered at 10 min. At the end of the study period all lambs received a lethal dose of pentobarbitone [12, 19, 20, 22]. Gestation-matched unventilated control (UVC) lambs received a lethal dose of pentobarbitone at delivery for identification of ventilation-associated protein alterations. Immediately after animals were euthanized, the lungs were removed en bloc. Lung tissue samples for proteome analysis from the gravity-dependent and non-dependent zones of the right upper lobe were snap frozen and stored at − 80 °C until analysis.
Determination of protein composition in TMT-labelled lung tissues
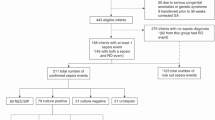
A detailed methodology describing protein extraction, TMT labelling and data extraction of the preterm lung tissue is available [24]. Inclusion in the study was based upon lung samples from our biobank that met the following criteria; ventilation strategy (StatPEEP, SI or DynPEEP), gestational age < 128d, no evidence of illness in ewe (hypoxia, sepsis, chorioamnionitis) and complete qPCR dataset. Following protein extraction only samples with a peptide quantity > 10 µg were labelled with TMT. Details of these sample reductions and final sample sizes for analysis are shown in Additional file 1: Fig. S1. This provided a final sample of 6–7 lambs in the unventilated control group and between 9 and 22 in the ventilated groups. As the first study to compare different respiratory interventions in the preterm lung using this methodology a formal power calculation was not possible. We have previously demonstrated that a sample size of 7 unventilated controls and 10 ventilated lambs identified important differences between unventilated and ventilated lambs [18, 25].
Bioinformatics and statistical analysis
Bioinformatic analysis
The Perseus software platform [26] was used to determine and visualize differential protein abundance between the unventilated control and RM-groups using Welsh’s t-test (two-sided) to account for the variability in group size with p < 0.05 considered significant. Protein class composition of significant proteins was assigned using the PANTHER tool [27]. Enriched pathways and functions associated with the proteome data sets were identified using the WebGestalt tool [28]. As per previous studies [17, 25], to address ovine database limitation in analysis, software homology of sheep to human proteins was assessed by NCBI Basic Local Alignment Search Tool (BLAST); 93% of proteins exhibited homology ≥ 75%.
Physiological and functional parameters
Physiological and functional parameters were recorded at three time points; immediately following the end of the lung recruitment strategy (approximately three minutes post-birth, matched in the StatPEEP group), and at five minutes and ninety minutes post-birth. Shapiro–Wilk normality testing confirmed the normality of data distribution, with results represented as median and interquartile range. Significant differences between gestation groups were sought using parametric and non-parametric ANOVA as appropriate (one-way ANOVA or Kruskall-Wallis ANOVA, respectively) with post hoc testing to identify intergroup differences as necessary (Holm-Šídák's or Dunn’s multiple comparison test, respectively). All statistical analysis was performed with GraphPad PRISM 9.1.2 (GraphPad Software, SanDiego, CA) and p < 0.05 considered significant. Unless otherwise stated, p values refer to ANOVA.
Assessing proteome and physiological phenotypes that differed between the RM groups
TO assess if the three treatment groups exhibited differences in the compositional proteome and physiological measures, we used sparse Partial Least Squares (sPLS) regression [29], a multivariate methodology which relates (integrates) two data matrices X (e.g. protein abundance) and Y (e.g. physiological and functional measures), and performs simultaneous variable selection in the two data sets (set at 200 proteins, 20 physiological/functional measures) in the R package MixOmics (version 3.1.1).
Results
Lambs were well-matched clinically (Additional file 1: Table S1).
Proteome profiling identified regional and ventilation strategy-specific alterations in the lung proteome
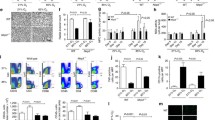
2373 proteins were analysed. The total number of differentially expressed proteins (DEPs) within the lung was highest in the SI group, followed by StatPEEP (Fig. 2A). The non-gravity dependent lung had more DEPs than the dependent lung for all ventilation groups. Co-expression of DEPs in both lung regions was approximately tenfold higher in StatPEEP and SI groups compared to DynPEEP (Fig. 2B). 56% (100 proteins) of DEPs in the non-dependent lung and 69% (14 proteins) of DEPs in the dependent lung were only expressed in a single study group (Fig. 2C). There were a higher number of decreased DEPs than increased DEPs in both lung regions of the SI and DynPEEP groups and the opposite for StatPEEP group (Fig. 2D). A total of 20 protein classes were assigned to the non-dependent lung and 18 the dependent lung (Fig. 2E). In the non-dependent lung 15 protein classes were common to all intervention groups (Additional file 1: Table S2), with defence/immunity proteins only identified in the SI group and cell adhesion molecules, structural proteins and transmembrane signal receptors absent in the DynPEEP group. Translational proteins were proportionally two-fold higher in the DynPEEP group compared to the StatPEEP and SI groups (23% versus 11 and 12%). In the dependent lung, protein class diversity was decreased by approximately 45% in the DynPEEP group compared to the other groups, except for transporter (threefold increase) and gene-specific transcriptional regulator proteins (3 to sixfold) (Additional file 1: Table S3).

Results from Orbitrap-MS analysis at 90 min of life in the non-dependent and dependent lung for the no-recruitment manoeuvre (StatPEEP), sustained inflation (SI) and dynamic stepwise positive end-expiratory pressure (DynPEEP) strategies relative to unventilated controls. Number of differentially expressed proteins (DEPs) (A), Venn diagram comparing differential protein expression between lung regions (B) and between intervention strategies within the same lung region (C). Volcano plots with DEPs highlighted in colour and the five most abundance and depleted proteins identified (D). PANTHER analysis of the protein class composition amongst DEPs identified in non-dependent and dependent lung (E)
WebGestalt over-representation analysis (ORA) revealed lung region specific pathway enrichment, with unique pathways associated with specific ventilation strategies
In cellular enrichment analysis, non-dependent lung DEPs from the DynPEEP group were cytoplasmic (Fig. 3A). Uniquely, only DEPs from the StatPEEP group were associated with adherens and cell-substrate junction, and only SI DEPs were from the extracellular region. Both the StatPEEP group and SI group were associated with the cell membrane and organelle components. A broader array of cellular components was identified in the dependent lung, largely due to association with specific organelle components in the StatPEEP (mitochondria, z disc), SI (endoplasmic reticulum, golgi and actin cytoskleleton) and DynPEEP (organelle membrane and secretory vesicle) groups.

WebGestalt identification of enriched cellular components and biological pathways in non-dependent and dependent lung exposed to either the no-recruitment manoeuvre (StatPEEP), sustained inflation (SI) or dynamic stepwise positive end-expiratory pressure (DynPEEP) strategy. Enrichment of cellular components in the non-dependent and dependent lung (A). Identified Reactome pathways with dark grey box highlighting co-enrichment of the citric acid (TCA) cycle and respiratory electron transport pathway in both StatPEEP and SI dependent lung (B). DEP mapping of StatPEEP and SI DEPs onto the citric acid (TCA) cycle and respiratory electron transport pathways (C). Inset; Venn diagram comparing DEP identification within the pathway in StatPEEP and SI groups. The threshold for identification of significant enrichments was adjusted p value < 0.05 and three or more pathways associated with the cellular component or pathway. Redundant Reactome pathways were reduced using weighted set cover
18 enriched Reactome pathways were identified in the non-dependent lung and 12 in the dependent lung (Fig. 3B). In the non-dependent lung, the inflammation response was divergent amongst RM-groups with the adaptive immune system enriched only in the StatPEEP group, as opposed to cytokine signalling pathway enrichments in SI and DynPEEP groups. Several pathways were enriched only in a single RM group and these included; Membrane trafficking and collagen formation (StatPEEP), apoptosis and translation (SI) and biological oxidations and ribosomal scanning (DynPEEP).
In the dependent lung, enriched pathways were specific to RM-groups with the exclusion of the citric acid cycle (TCA) and respiratory electron transport pathway, which was enriched in both the StatPEEP and SI groups. Amongst the DEPs associated with The TCA pathway, seven DEPs were altered in both the StatPEEP and SI groups, four DEPs were altered in only the StatPEEP group and eight DEPs were altered in only the SI group. 58% of DEPs mapped to the oxidative phosphorylation component of the TCA and respiratory electron transport pathway (Fig. 3C), with StatPEEP DEPs associated with Complexes I and II, whilst SI DEPs were associated with Complexes I–IV.
Integrative analysis identified regional and temporal protein-function associations specific to each ventilation strategy
In addition to comparing the fold change in protein abundance between ventilated and unventilated lung, we also used sPLS to test for associations between protein abundance (normalised protein reporter intensities reported in the ventilated lung) and functional measures of aeration, ventilation and clinical status. Two protein-function networks were observed in all lung regions and ventilation strategies, with an additional single protein-function network observed in the non-dependent lung of the DynPEEP group (Table 1). The highest number of protein-function associations were observed in the StatPEEP group (Table 1).
Distinct temporal associations were observed between lung regions and ventilation strategies (Table 1, Fig. 4). In the StatPEEP group the highest number of protein-function associations in either the non-dependent or dependent lung were observed after 90 min of respiratory support, and included measures of lung morphology, lung injury and oxygenation and acid–base. In contrast, in the SI group, ventilation parameters and Cdyn at 5 min accounted for 71% of protein-function relations in the non-dependent lung. In the dependent lung a single protein network was associated with the completion of the SI and 5 min ventilation parameters and Cdyn and a second network consisting of three proteins associated with acid–base at 90 min. The DynPEEP group also exhibited associations between non-dependent lung proteins and ventilation parameters at the end of the DynPEEP strategy and 5 min, as well as 90 min. 80% of associations in the DynPEEP dependent lung involved functional measures at 90 min (oxygenation and haemoglobin).

Sparse partial least squared regression and classification (sPLS) between protein abundance and functional measures in the non-dependent and dependent lung of ventilated preterm lambs receiving Static PEEP (StatPEEP), sustained inflation (SI) or a dynamic stepwise positive end-expiratory pressure (DynPEEP) strategy. sPLS in regression model was used to model a causal relationship between the proteins and functional measures. The network is displayed graphically as nodes (proteins and functional measures) and edges (relationship between nodes), with the colour edge intensity indicating the level of the association: red, positive and green, negative. Only the strongest pairwise associations were displayed, with a cut-off threshold of 0.7 (positive and negative). Coloured boxes indicate timing of functional measures (light blue = during recruitment manoeuvre (first 3 min), grey = 5 min post-birth, black = 90 min post-birth). White boxes contain the protein name with proteins identified in the DisGeNET database as associated with bronchopulmonary dysplasia, respiratory distress syndrome or respiratory failure highlighted in bold purple text. Function measure abbreviations; CRS total compliance, BE base excess, Vmin minute volume, VT tidal volume, Cdyn dynamic compliance, PIP peak inspiratory pressure, ΔP change in pressure, HR heart rate, mean BP mean blood pressure
Discussion
To our knowledge this is the first study to map the complete protein-functional responses of the preterm lung at initiation of aeration and ventilation at birth, the earliest time point in which clinical respiratory decisions can impact injury. Effective establishment of ventilation at birth is vital [30]. The optimal approach to support ventilation in the preterm lung whilst minimising injury remains unclear due to crude, late and in-direct measures of lung injury, such as death and oxygen requirement. These fail to delineate the molecular impact of such strategies on the early, acute lung injury response [31]. We have recently demonstrated that lung mechanics, regional aeration and ventilation states (such as relative overdistension and atelectasis), as well as gravity dependent expression of early injury markers, differed between SI, StatPEEP and DynPEEP strategies, suggesting each generates a different, and biologically important, injury response in the preterm lung [12, 19, 23]. Interestingly oxygenation, the primary clinical tool for defining clinical status and efficacy in trials, was not a delineator of the injury differences. Thus, our biobank was ideal to demonstrate the role of proteomic analysis to understand the response of the preterm lung to respiratory support before developing clinical trials, and use translational models to better understand the phenotype responses to different clinical approaches to the initial management of RDS.
Overall, the biggest impact on the proteome was seen in the SI group which exhibited the highest number of DEPs, whilst the DynPEEP group had fewer DEPs and a simpler protein class composition. Further highlighting the influence of ventilation strategies on protein expression, only 16% of DEPs in the non-dependent lung and 4% in the dependent lung were common to all aeration strategy groups. Protein expression exhibited a gravity-dependent pattern in all ventilation groups, with more DEPs identified in the gravity non-dependent lung than dependent, and at the individual protein level more than 88% of DEPs were identified in a single lung region alone. We have previously shown that an SI generated the greatest gravity-dependent heterogeneity in aeration and ventilation at birth that caused lung injury events specific to the regional volume state [12]. Thus, providing a mechanistic link to explain the greater DEPs especially in the non-dependent lung following a SI. In low compliance states (such as RDS) the non-dependent lung is preferentially ventilated, creating high volutrauma risk during the large mechanical forces applied during the rapid inflation of a SI. A recent large trial of SI at birth was ceased early due to a higher early mortality rate in the SI group of unknown causation [15]. Our findings, combined with the previous adverse impacts on regional aeration and surfactant efficacy in preclinical studies [12, 23, 32], provide the first indication that a SI may generate unique and acute biologically harmful changes in the preterm lung related to heterogenous rapid pressure and volume exposure throughout the lung.
Distinct gravity-dependent pathway over-representation was identified for each of the aeration strategies, suggesting differing biological events. In the non-dependent lung there were no common pathways identified between the StatPEEP and other groups, and only three common pathways were identified for SI and DynPEEP groups. In the dependent lung, only a single common pathway (R-HSA-1428517; citric acid cycle and respiratory electron transport) was identified in the StatPEEP and SI group. In the non-dependent region of the StatPEEP lung alone we observed unique identification of 30 proteins associated with an adaptive immune response phenotype. This diverse inflammatory phenotype response after only a brief period of respiratory support may provide insight into why broad anti-inflammatory agents such as corticosteroids, rather than targeted anti-inflammatory therapies, have been effective in reducing the respiratory burden of preterm RDS [33].
The SI non-dependent lung phenotype was strongly and uniquely associated with enrichment of translational proteins with 75% (15/20) of decreased proteins identified as ribosomal proteins and 67% (10/18) of increased proteins as translational proteins (initiation, elongation and release factors). Ribosome biogenesis and protein translation are finely coordinated with and essential for cell growth, proliferation, differentiation, and development [34]. Impairment of any of these two cellular processes can severely retard cell growth and perturb development [34]. Only 12 proteins were associated with a single pathway in either non-dependent (biological oxidations) or dependent lung (innate immune system) following a DynPEEP strategy. This concurs with our previous observation of improved ventilation and aeration homogeneity, lung mechanics, surfactant efficacy and subsequently reduced early lung injury mRNA markers following DynPEEP compared to SI and StatPEEP in our biobank [12, 32]. Further it provides potential mechanistic support to our hypothesis that gentle intermittent lung aeration via tidal inflations, using PEEP to maintain aeration-gains, will be less injurious than rapid lung inflation [12].
In the dependent lung the TCA cycle and respiratory electron transport pathway was identified as significant in both StatPEEP and SI dependent lung tissue, suggesting that these aeration strategies may generate significant oxidative stresses. Newborn infants encounter a dramatic surge of oxidative stress immediately after birth. The preterm lung lacks antioxidant protection [35], the risk is further exacerbated when higher oxygen concentrations are used [36]. Despite oxidative stress clearly playing a vital role in causing lung injury, antioxidant treatment has not shown clinical efficacy in preventing BPD, suggesting more complex mechanisms are involved [36, 37]. In our study we identified nuanced differences in protein abundance profiles between StatPEEP and SI lung tissues such that whilst complexes II and IV proteins exhibited similar alterations in both strategies, only SI caused decreased abundance of complex I and III proteins. Decreased enzymatic activity of complex I has previously been associated with severity of lung-stretching in a ventilated neonatal mouse model [38]. Conversely, gentler tidal volumes abrogate mitochondrial dysfunction [38]. In the current context this suggests that therapies targeting complex I may be more suited to strategies prone to over-distension such as SI.
In secondary studies we used sparse partial least squares modelling to identify the most important function-protein associations between 61 functional variables from four time-points and protein abundance in 2,373 gravity non-dependent and dependent lung protein. There are several advantages to using this approach; dimension reduction and variable selection is performed simultaneously and allows for identification of most relevant associations, sPLS regression exhibits good performance even when the sample size is much smaller than the total number of variables (as occurs with large ‘omic datasets) and both univariate and multivariate responses can be included in the analysis [39]. Highly complex and dense protein-function networks were observed in both lung regions of the StatPEEP group with most implicated functional measures involving clinical outcome measures and gene expression of lung injury. In contrast both the SI and DynPEEP groups were simpler, with protein-function associations and increased focus on associations between protein abundance and early ventilation delivery. This at first is a surprising result given that the proteome of SI lung tissue was associated with the highest numbers of DEPs. However whilst proteomic analysis facilitated the identification of proteins not previously associated with preterm lung injury, previous preterm, lung injury studies have made a priori assumptions about functional variables, such as the utility of the commonly used 6-member lung injury gene panel, first validated in a StatPEEP model 12 years ago [40]. Several recent papers have used plasma biomarker panels to define broad sub-phenotypes within ARDS [9, 41, 42]. Plasma protein biomarkers in general contributed more prominently to the phenotype definitions than most of the commonly used clinical variables in adults with ARDS [9]. Few delivery room studies have included protein biomarkers in their assessment, and when they have, single targets were used [43, 44], thereby risking missing unrecognised differences between interventions, and limiting understanding of the pathophysiology events. The diversity in protein phenotype between the three aeration strategies investigated suggests that using non-targeted approaches such as proteomics in early preterm life studies would be highly advantageous at capturing the biological impact of diverse respiratory support strategies.
Limitations
There are several limitations of this study. We, and others, have previously detailed the clinical translation limitations of our ventilation strategies and instrumented lambs generally [12, 19, 20, 23], specifically suppression of breathing and intubation [19, 22, 23, 45,46,47,48]. Lamb numbers were unbalanced between the strategies as fewer StatPEEP study animals, which acted as a ventilation ‘control’ group across studies, were available within the biobank [12, 19, 20, 23]. To avoid selection bias and knowing some samples would be lost during the extraction workflow, we included all StatPEEP, SI and DynPEEP samples rather than matching group sizes. We did not apply FDR correction on the protein p values, however we attempted to counter this within the bioinformatic analysis by limiting reporting of ORA results to cellular compartments and pathways containing at least three DEPs. The sPLS approach being based on protein abundance rather than differential expression does not require consideration of multiple testing principles. To facilitate interpretation of protein-function networks we used the sPLS approach and limited the number of variables to be included and applied an r cutoff of 0.7. Whilst this provided a clearer picture of the variables most contributing to each phenotype, important but weaker biological associations may have been missed.
Conclusions
Ideally, strategies for BPD prevention should start immediately after birth [49], focusing on supporting lung aeration and then managing RDS whilst minimising ventilator induced lung injury [4]. This study is the first to demonstrate that proteomics can identify distinct acute injury responses to different respiratory strategies at birth. Importantly, it maybe possible to use proteomics in the first hours of life to phenotype preterm infants based on the direct aetiology of their lung injury risk, an important step towards true precision respiratory support.
Availability of data and materials
The datasets generated ans analysed during the current study are available at Figshare (https://doi.org/10.26188/19252103.v1).
Abbreviations
- ARDS:
-
Acute respiratory distress syndrome
- BLAST:
-
Basic Local Alignment Search Tool
- BPD:
-
Bronchopulmonary dysplasia
- DynPEEP:
-
Dynamic PEEP
- DEPs:
-
Differentially expressed proteins
- StatPEEP:
-
Static PEEP
- ORA:
-
Over-representation analysis
- PEEP:
-
Positive end-expiratory pressure
- PIP:
-
Peak inspiratory pressure
- PPV:
-
Positive-pressure ventilation
- RDS:
-
Respiratory distress syndrome
- SI:
-
Sustained inflation
- sPLS:
-
Sparse partial least squares
- UVC:
-
Unventilated control
- VTV:
-
Volume-targeted ventilation
References
Onland W, Hutten J, Miedema M, Bos LD, Brinkman P, Maitland-van der Zee AH, et al. Precision medicine in neonates: future perspectives for the lung. Front Pediatr. 2020;8: 586061.
Stoll BJ, Hansen NI, Bell EF, Walsh MC, Carlo WA, Shankaran S, et al. Trends in care practices, morbidity, and mortality of extremely preterm neonates, 1993–2012. JAMA. 2015;314(10):1039–51.
Thebaud B, Goss KN, Laughon M, Whitsett JA, Abman SH, Steinhorn RH, et al. Bronchopulmonary dysplasia. Nat Rev Dis Primers. 2019;5(1):78.
Blazek EV, East CE, Jauncey-Cooke J, Bogossian F, Grant CA, Hough J. Lung recruitment manoeuvres for reducing mortality and respiratory morbidity in mechanically ventilated neonates. Cochrane Database Syst Rev. 2021;3: CD009969.
Bruschettini M, O’Donnell CP, Davis PG, Morley CJ, Moja L, Calevo MG. Sustained versus standard inflations during neonatal resuscitation to prevent mortality and improve respiratory outcomes. Cochrane Database Syst Rev. 2020;3: CD004953.
Jensen EA, Dysart K, Gantz MG, McDonald S, Bamat NA, Keszler M, et al. The diagnosis of bronchopulmonary dysplasia in very preterm infants. An evidence-based approach. Am J Respir Crit Care Med. 2019;200(6):751–9.
Sinha P, Calfee CS. Phenotypes in acute respiratory distress syndrome: moving towards precision medicine. Curr Opin Crit Care. 2019;25(1):12–20.
Calfee CS, Delucchi KL, Sinha P, Matthay MA, Hackett J, Shankar-Hari M, et al. Acute respiratory distress syndrome subphenotypes and differential response to simvastatin: secondary analysis of a randomised controlled trial. Lancet Respir Med. 2018;6(9):691–8.
Calfee CS, Delucchi K, Parsons PE, Thompson BT, Ware LB, Matthay MA, et al. Subphenotypes in acute respiratory distress syndrome: latent class analysis of data from two randomised controlled trials. Lancet Respir Med. 2014;2(8):611–20.
Famous KR, Delucchi K, Ware LB, Kangelaris KN, Liu KD, Thompson BT, et al. Acute respiratory distress syndrome subphenotypes respond differently to randomized fluid management strategy. Am J Respir Crit Care Med. 2017;195(3):331–8.
Wendel Garcia PD, Caccioppola A, Coppola S, Pozzi T, Ciabattoni A, Cenci S, et al. Latent class analysis to predict intensive care outcomes in acute respiratory distress syndrome: a proposal of two pulmonary phenotypes. Crit Care. 2021;25(1):154.
Tingay DG, Pereira-Fantini PM, Oakley R, McCall KE, Perkins EJ, Miedema M, et al. Gradual aeration at birth is more lung protective than a sustained inflation in preterm lambs. Am J Respir Crit Care Med. 2019;200(5):608–16.
Hooper SB, Te Pas AB, Kitchen MJ. Respiratory transition in the newborn: a three-phase process. Arch Dis Child Fetal Neonatal Ed. 2016;101(3):F266–71.
Tingay DG, Bhatia R, Schmolzer GM, Wallace MJ, Zahra VA, Davis PG. Effect of sustained inflation vs stepwise PEEP strategy at birth on gas exchange and lung mechanics in preterm lambs. Pediatr Res. 2014;75(2):288–94.
Kirpalani H, Ratcliffe SJ, Keszler M, Davis PG, Foglia EE, Te Pas A, et al. Effect of sustained inflations vs intermittent positive pressure ventilation on bronchopulmonary dysplasia or death among extremely preterm infants: the SAIL randomized clinical trial. JAMA. 2019;321(12):1165–75.
Kirpalani H, Keszler M, Foglia EE, Davis P, Ratcliffe S. Considering the validity of the SAIL trial—a navel gazers guide to the SAIL trial. Front Pediatr. 2019;7:495.
Pereira-Fantini PM, Byars SG, McCall KE, Perkins EJ, Oakley RB, Dellaca RL, et al. Plasma proteomics reveals gestational age-specific responses to mechanical ventilation and identifies the mechanistic pathways that initiate preterm lung injury. Sci Rep. 2018;8(1):12616.
Schmid C, Ignjatovic V, Pang B, Nie S, Williamson NA, Tingay DG, et al. Proteomics reveals region-specific hemostatic alterations in response to mechanical ventilation in a preterm lamb model of lung injury. Thromb Res. 2020;196:466–75.
Tingay DG, Rajapaksa A, Zannin E, Pereira-Fantini PM, Dellaca RL, Perkins EJ, et al. Effectiveness of individualized lung recruitment strategies at birth: an experimental study in preterm lambs. Am J Physiol Lung Cell Mol Physiol. 2017;312(1):L32–41.
McCall KE, Waldmann AD, Pereira-Fantini P, Oakley R, Miedema M, Perkins EJ, et al. Time to lung aeration during a sustained inflation at birth is influenced by gestation in lambs. Pediatr Res. 2017;82(4):712–20.
Probyn ME, Hooper SB, Dargaville PA, McCallion N, Crossley K, Harding R, et al. Positive end expiratory pressure during resuscitation of premature lambs rapidly improves blood gases without adversely affecting arterial pressure. Pediatr Res. 2004;56(2):198–204.
Tingay DG, Polglase GR, Bhatia R, Berry CA, Kopotic RJ, Kopotic CP, et al. Pressure-limited sustained inflation vs gradual tidal inflations for resuscitation in preterm lambs. J Appl Physiol (1985). 2015;118(7):890–7.
Tingay DG, Rajapaksa A, Zonneveld CE, Black D, Perkins EJ, Adler A, et al. Spatiotemporal aeration and lung injury patterns are influenced by the first inflation strategy at birth. Am J Respir Cell Mol Biol. 2016;54(2):263–72.
Tingay D, Pereira-Fantini PM. First description of ventilation strategy-specific phenotypes involved in initiating preterm lung injury. University of Melbourne; 2022.
Pereira-Fantini PM, Pang B, Byars SG, Oakley RB, Perkins EJ, Dargaville PA, et al. Preterm lung exhibits distinct spatiotemporal proteome expression at initiation of lung injury. Am J Respir Cell Mol Biol. 2019;61(5):631–42.
Tyanova S, Temu T, Sinitcyn P, Carlson A, Hein MY, Geiger T, et al. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat Methods. 2016;13(9):731–40.
Thomas PD, Campbell MJ, Kejariwal A, Mi H, Karlak B, Daverman R, et al. PANTHER: a library of protein families and subfamilies indexed by function. Genome Res. 2003;13(9):2129–41.
Liao Y, Wang J, Jaehnig EJ, Shi Z, Zhang B. WebGestalt 2019: gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019;47(W1):W199–205.
Le Cao KA, Rossouw D, Robert-Granie C, Besse P. A sparse PLS for variable selection when integrating omics data. Stat Appl Genet Mol Biol. 2008;7(1):Article 35.
Schmolzer GM, Kumar M, Aziz K, Pichler G, O’Reilly M, Lista G, et al. Sustained inflation versus positive pressure ventilation at birth: a systematic review and meta-analysis. Arch Dis Child Fetal Neonatal Ed. 2015;100(4):F361–8.
Manley BJ, Owen LS, Hooper SB, Jacobs SE, Cheong JLY, Doyle LW, et al. Towards evidence-based resuscitation of the newborn infant. Lancet. 2017;389(10079):1639–48.
Tingay DG, Togo A, Pereira-Fantini PM, Miedema M, McCall KE, Perkins EJ, et al. Aeration strategy at birth influences the physiological response to surfactant in preterm lambs. Arch Dis Child Fetal Neonatal Ed. 2019;104(6):F587–93.
Ramaswamy VV, Bandyopadhyay T, Nanda D, Bandiya P, Ahmed J, Garg A, et al. Assessment of postnatal corticosteroids for the prevention of bronchopulmonary dysplasia in preterm neonates: a systematic review and network meta-analysis. JAMA Pediatr. 2021;175(6): e206826.
Zhou X, Liao WJ, Liao JM, Liao P, Lu H. Ribosomal proteins: functions beyond the ribosome. J Mol Cell Biol. 2015;7(2):92–104.
Buhimschi IA, Buhimschi CS, Pupkin M, Weiner CP. Beneficial impact of term labor: nonenzymatic antioxidant reserve in the human fetus. Am J Obstet Gynecol. 2003;189(1):181–8.
Teng R-J. Oxidative stress in neonatal lung diseases. In: Chakraborti S, Chakraborti T, Das SK, Chattopadhyay D, editors. Oxidative stress in lung diseases, vol. 1. Singapore: Springer Singapore; 2019. p. 51–84.
Welty SE. Is there a role for antioxidant therapy in bronchopulmonary dysplasia? J Nutr. 2001;131(3):947S-S950.
Ratner V, Sosunov SA, Niatsetskaya ZV, Utkina-Sosunova IV, Ten VS. Mechanical ventilation causes pulmonary mitochondrial dysfunction and delayed alveolarization in neonatal mice. Am J Respir Cell Mol Biol. 2013;49(6):943–50.
Chung D, Chun H, Keles S. An Introduction to the ‘spls’ Package, Version 1.0. Madison, WI: University of Wisconsin. 2012.
Wallace MJ, Probyn ME, Zahra VA, Crossley K, Cole TJ, Davis PG, et al. Early biomarkers and potential mediators of ventilation-induced lung injury in very preterm lambs. Respir Res. 2009;10:19.
Bos LD, Schouten LR, van Vught LA, Wiewel MA, Ong DSY, Cremer O, et al. Identification and validation of distinct biological phenotypes in patients with acute respiratory distress syndrome by cluster analysis. Thorax. 2017;72(10):876–83.
Calfee CS, Janz DR, Bernard GR, May AK, Kangelaris KN, Matthay MA, et al. Distinct molecular phenotypes of direct vs indirect ARDS in single-center and multicenter studies. Chest. 2015;147(6):1539–48.
La Verde A, Franchini S, Lapergola G, Lista G, Barbagallo I, Livolti G, et al. Effects of sustained inflation or positive pressure ventilation on the release of adrenomedullin in preterm infants with respiratory failure at birth. Am J Perinatol. 2019;36(S02):S110–4.
El-Chimi MS, Awad HA, El-Gammasy TM, El-Farghali OG, Sallam MT, Shinkar DM. Sustained versus intermittent lung inflation for resuscitation of preterm infants: a randomized controlled trial. J Matern Fetal Neonatal Med. 2017;30(11):1273–8.
Hillman NH, Kemp MW, Noble PB, Kallapur SG, Jobe AH. Sustained inflation at birth did not protect preterm fetal sheep from lung injury. Am J Physiol Lung Cell Mol Physiol. 2013;305(6):L446–53.
Sobotka KS, Hooper SB, Allison BJ, Te Pas AB, Davis PG, Morley CJ, et al. An initial sustained inflation improves the respiratory and cardiovascular transition at birth in preterm lambs. Pediatr Res. 2011;70(1):56–60.
te Pas AB, Siew M, Wallace MJ, Kitchen MJ, Fouras A, Lewis RA, et al. Effect of sustained inflation length on establishing functional residual capacity at birth in ventilated premature rabbits. Pediatr Res. 2009;66(3):295–300.
te Pas AB, Siew M, Wallace MJ, Kitchen MJ, Fouras A, Lewis RA, et al. Establishing functional residual capacity at birth: the effect of sustained inflation and positive end-expiratory pressure in a preterm rabbit model. Pediatr Res. 2009;65(5):537–41.
Foglia EE, Jensen EA, Kirpalani H. Delivery room interventions to prevent bronchopulmonary dysplasia in extremely preterm infants. J Perinatol. 2017;37(11):1171–9.
Plubell DL, Wilmarth PA, Zhao Y, Fenton AM, Minnier J, Reddy AP, et al. Extended multiplexing of tandem mass tags (TMT) labeling reveals age and high fat diet specific proteome changes in mouse epididymal adipose tissue. Mol Cell Proteomics. 2017;16(5):873–90.
Acknowledgements
The authors thank Mr. Magdy Sourial and Ms. Rebecca Sutton for their assistance with management of the preterm lamb model.
Funding
This study was supported by a National Health and Medical Research Council Project Grant (1009287) and the Victorian Government Operational Infrastructure Support Program (Melbourne, Victoria, Australia), a National Health and Medical Research Council Clinical Career Development Fellowship (1053889; D.G.T.), and a National Health and Medical Research Council Program Grant (606789). Chiesi Farmaceutici S.p.A. provided the Curosurf used in this study as part of an unrestricted grant (D.G.T.) at the Murdoch Children’s Research Institute.
Author information
Authors and Affiliations
Contributions
P.P-F. Study conception and design, acquisition of data, analysis and interpretation of data, drafting manuscript, statistical analysis. KF. Critical revision of the manuscript for important intellectual content. K.E.McC., R.O., and EP. Acquisition, analysis and interpretation of clinical data, critical revision of the manuscript for important intellectual content. S.B. statistical analysis. D.G.T. Study conception and design, acquisition, analysis and interpretation of clinical data, critical revision of the manuscript for important intellectual content, obtained funding, study supervision. P.P-F. wrote the first draft with assistance from D.G.T. and K.F. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All studies were approved by the Murdoch Children’s Research Institute Animal Ethics Committee in accordance with National Health and Medical Research Council Guidelines.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Fig. S1.
Biobank assessment and proteomic sample filtering workflow. Table S1. Clinical characteristics of study population at birth. Table S2. Results from PANTHER protein class composition analysis performed on gravity non-dependent lung samples. Table S3. Results from PANTHER protein class composition analysis performed on gravity dependent lung samples.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Pereira-Fantini, P.M., Ferguson, K., McCall, K. et al. Respiratory strategy at birth initiates distinct lung injury phenotypes in the preterm lamb lung. Respir Res 23, 346 (2022). https://doi.org/10.1186/s12931-022-02244-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12931-022-02244-x