
Abstract
Background
The insulin-like growth factor (IGF-I) and growth hormone (GH) genes have been identified as major regulators of milk yield and composition, and reproductive performance in cattle. Genetic variations/polymorphism in these genes have been found to influence milk production, yield and quality. This investigation aimed to explore the association between IGF-I and GH polymorphisms and milk yield and composition, and reproductive performance in a herd consisting of 1000 Holstein–Friesian (HF) dairy cattle from El-Alamia farm. The experimental animals were 76 ± 7.25 months in age, with an average live weight of 750 ± 50.49 kg, and raised under the same conditions of feeding and weather. The studied animals were divided into three categories; high producers (n = 280), medium producers (n = 318) and low producers (n = 402).
Results
The digestion of 249 bp for IGF-I-SnaBI using the Restriction-fragment-length-polymorphism (RFLP) technique yielded two alleles; T (0.59) and C (0.41) and three genotypes; TT (0.52), TC (0.39) and CC (0.09) and this agrees with the results of DNA/gene sequencing technique. The sequencing analysis of the IGF-I gene revealed polymorphism in position 472 (C > T). Nucleotide sequencing of the amplified fragment of the IGF-I gene of different genotypes was done and submitted to the NCBI GenBank with Accession no. MH156812.1 and MH156811.1. While the digestion of 432 bp for GH-AluI using the RFLP technique yielded two alleles; A (0.81) and G (0.19) and two genotypes; AA (0.77) and AG (0.23) and this agrees with the results of DNA/gene sequencing technique. The sequencing analysis of the GH gene revealed polymorphism in the position 1758 C > G and in turn led to changes in amino acid sequence as Alanine for (A) compared to Glycine for (G). Nucleotide sequencing of the amplified fragment of the GH gene was done and submitted to the NCBI GenBank with Accession no. MH156810.1. The results of this study demonstrate the effects of variants of the GH-IGF-I somatotrophic axis on milk production and composition traits in commercial HF cattle. The greatest values of milk yield and reproductive performance were observed on IGF-I-SnaBI-TC and GH-AluI-AG genotypes. While the greatest % fat and % protein values were observed on IGF-I-SnaBI-CC and GH-AluI-AA genotyped individuals.
Conclusion
The genetic variation of the studied genes can be utilized in selecting animals with superior milk yield, composition and reproductive performance in Holstein–Friesian Dairy Cattle under subtropical conditions.
Similar content being viewed by others


Background
With the advancement of technologies and molecular genetics, livestock breeding has become more efficient and cost-effective [1, 2]. In the last few decades, exploring the role of major/ candidate genes which control livestock production has become a focus of research [3]. For instance, the genetic regulation of insulin-like growth factor-I (IGF-I) [4] and growth hormone (GH) [5] genes have been well-studied in many livestock species, including cattle [5], sheep [6], camels [7] and pigs [8]. The molecular genetic studies of IGF-I and GH genes mainly targets their association with growth and reproductive performances, carcass traits, and milk yield and composition [5, 9].
The IGF growth factor system consists of two ligands (IGF-I and IGF-II) and two cell-surface receptor types I and II. IGF-I and II are structurally related proteins, playing a key role in cell differentiation, growth, embryogenesis, regulation of metabolism and regulation of cell proliferation [10, 11]. IGF-I gene encodes a hormone similar in structure to insulin, which controls cell growth and differentiation. It, also called somatomedin C, is a protein that in humans is encoded by the IGF-I gene [12]. The IGF-I gene is located on chromosome 12 at position 12q23 in humans, on 5 in cattle, on 6 in pigs and 10 in mice [13]. IGF-I is a polypeptide of a molecular weight 7.5 kDa built of 70 amino acids. It is produced primarily by the liver as an endocrine hormone as well as in target tissues in a paracrine/autocrine fashion. In dairy cattle, IGF-I acts primarily on the mammary gland to stimulate milk synthesis [12,13,14,15]. This hormone also affects milk composition, including fat and protein content [16]. In the liver, IGF-I is dynamically regulated by lactation and energy balance [17]. Moreover, IGF-I plays an essential role in pre-and postnatal growth, muscle development, and bone formation [11]. The promoter region of the IGF-I gene contains several single nucleotide polymorphisms (SNPs), which regulate transcriptional activity and association with growth performance [7]. Studies have shown that several IGF-I-SNPs were associated with growth performance, carcass traits, and milk yield in dairy cattle [18,19,20]. Cattle with specific IGF-I-SNPs have superior growth performance and higher milk yield [9, 21].
On the other hand, the GH gene is located on chromosome 20 in cattle and on chromosome 6 in pigs. It is a single-chain polypeptide of approximately 22 kDa, composed of 190 or 191 amino acids. The GH gene encodes the GH pituitary-derived hormone [22,23,24] that plays a vital role in animal physiology. It stimulates growth, milk production, and animal reproduction in livestock [25, 26]. Studies have shown that certain GH gene polymorphisms were related to milk production traits [27, 28]. For instance, a variant of the GH gene called the A allele is associated with higher milk production in Holstein cows [28]. Several investigations reported that specific GH-SNPs have increased feed efficiency and growth rates [20, 29]. In cattle, a SNP in the GH gene was associated with a carcass weight increase of 4 kg and a 2.3% increase in feed efficiency. Similarly, pigs with the GG genotype of GH had a significant increase in feed efficiency and reduced backfat thickness. [20, 29].
IGF-I is a primary mediator of the effects of GH. The action of GH is mediated by the transmembrane GH-R [14]. The binding of GH to GH-R activates an intracellular signalling pathway that induces the transcription of many genes including the IGF-I gene. GH is made in the anterior pituitary gland, released into the bloodstream, and then stimulates the liver to produce IGF-I which in turn stimulates systemic body growth through inducing growth-promoting effects on almost every cell in the body, especially skeletal muscle, cartilage, bone, liver, kidney, nerves, skin, hematopoietic cells and lungs [18, 30]. In addition to the insulin-like effects, IGF-I can also regulate cell growth and development, especially in nerve cells, as well as cellular DNA synthesis. The GH-IGF-I system controls processes, such as; fertility, lactation milk and nutrient partitioning necessary for lactogenesis [30].
Both GH and IGF-I play a crucial role in animal growth and development [19]. There is an intricate relationship between GH and IGF-I as the former stimulates hepatic IGF-I synthesis [19, 31]. IGF-I, in turn, positively feedbacks on GH secretion and modulates its receptor expression. Moreover, IGF-I affects animal reproduction, lactation, and feed intake by interacting with the somatotropic axis [32,33,34]. Both genes have been shown to play critical roles in milk production and composition in dairy cattle, with variations in these genes affecting milk output and quality [23, 25, 26]. By understanding how these genes function and interact, researchers can work to optimize milk production and improve the quality of their dairy products [18, 24]. Therefore, understanding the genetic regulation and interaction between the GH and IGF-I genes is essential in animal breeding programs.
In this aspect, according to the studies which aimed at enhancing mastitis resistance through selection, improving mastitis-related genetic traits in cattle is imperative. Somatic cell count (SCC) is an essential trait in such selection studies and is known to have positive genetic correlations with mastitis ranging from 0.36 to 0.67 [35]. Reduction of milk SCC through proposed selection studies not only aids in improving mastitis resistance but also helps reduce mastitis incidence [36]. Conversely, mastitis significantly alters the ion composition of milk, leading to increased electrical conductivity (EC), sodium, and chloride contents, as well as elevated milk pH resulting from the mixing of blood and extracellular fluid components in inflamed quarters with secreted milk [37]. In identifying and managing these traits, milk SCC, EC, and pH-related genes or polymorphisms need to be identified and improved accordingly [38]. In dairy cows, SCC exhibited positive correlation with clinical mastitis and with body weight [39]. As such, IGF-I and GH polymorphisms may also be explored for their strength of correlations with SCC, EC or pH [40, 41].
Although IGF-I genotypes have been extensively studied, little is known about their impact on blood composition in dairy cows during the periparturient period, which spans before and after calving [29]. However, IGF-I concentrations are altered during the postpartum period exhibiting linkage to various reproductive aspects [24, 42]. To address this knowledge gap, a part of the current study was conducted to investigate the relationship between IGF-I gene polymorphisms and its concentration in the periparturient period of Holstein–Friesian (HF) dairy cows.
Few investigations are available on GH-IGF-I polymorphisms and their association with milk yield and composition, and reproductive performance in HF cattle under subtropical conditions. To the best of our knowledge, no comprehensive investigations have examined differences in both GH and IGF-I genes in association with milk traits. Thus, the present study aimed to; 1) investigate the association between the polymorphism in IGF-I gene and milk yield and composition, and reproductive performance in HF cattle. 2) Explore the relationship between different GH genotypes and milk yield and composition, and reproductive performance in HF cattle. 3) examine the association between IGF-I gene polymorphisms and its concentration in the periparturient period of HF dairy cows.
Results
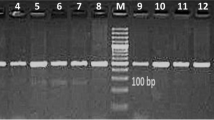
This study concerns mainly the polymorphism among tested cows for IGF-I and GH genes. It also spotlights the relationship between the differentiation of IGF-I and GH genes and milk yield and composition, and reproductive performance. PCR amplification for the tested animals produced an amplified 249 bp fragment for the IGF-I gene (Fig. 1A and Fig. S1A), and 432 bp for the GH gene (Fig. 1B, C and Fig. S1B).

A PCR amplification of the 5’-noncoding region of the bovine IGF-I gene from Holstein Frisian cattle. M, 50 bp DNA ladder. B PCR amplification of GH gene fragment from Holstein Frisian cattle. M, 100 bp DNA ladder. C Purified PCR product of GH gene (432 bp) from Holstein Frisian cattle. M, 50 bp DNA ladder
Insulin-like Growth Factor -1 (IGF-I)
PCR amplification and genotyping of IGF-I gene by RFLP
The amplified 249 bp fragment of HF cows of the IGF-I gene contains the 5’-noncoding region of the bovine IGF-I gene (Fig. 1A). A C → T substitution in the gene creates a new SnaBI restriction site, allowing for analysis using RFLP techniques. The PCR products obtained from HF cows were digested with SnaBI, resulting in three patterns: 1) Homozygous (CC) genotype with a non-digested PCR product (249 bp). 2) Homozygous (TT) genotype with 2 restricted fragments at 223 and 26 bp. 3) Heterozygote (CT) genotype with 3 restricted fragments at 249, 223, and 26 bp. The 26 bp restriction fragment was not observed on the gel (Fig. 2A).

A Agarose gel electrophoresis showing RFLP-SnaBI restriction pattern of Holstein Frisian cattle in 5’-noncoding region of the bovine IGF-I gene Lane M: 50-bp ladder marker. Lanes 1 and 2: Homozygous CC; genotype non-digested PCR product (249 bp). Lanes 3 and 4: Homozygous TT genotype with 2 restricted fragments at 223 and 26 bp. Lanes 5 and 6: heterozygote CT genotype with 3 restricted fragments at 249, 223 and 26 bp. The restriction fragment with size 26 bp has not been seen on the gel. B Result of endonuclease restriction with SnaBI using FastPCR C/T: Single nucleotide substitution, genotype TT, and C genotype CC
Sequence analysis and frequencies
The three genotypes in tested animals due to a restriction site at position 472 (TAC^GTA). Worth mentioning that this particular position (P472) has been previously identified in various cattle breeds [43, 44]. In the present study, the frequencies of the IGF-I-SnaBI alleles T and C were 0.59 and 0.41, respectively (Table 1).
Nucleotide sequencing of the amplified IGF-I gene fragment of HF cows was submitted to the NCBI GenBank (Accession no. MH156812.1 and MH156811.1) (Fig. 3A, B). The transition mutation (C > T) in the IGF-I gene was found with the detected three genotypes (Fig. 3C).

A A 249 bp sequence of IGF-I gene for Holstein Frisian genotype (AA = TT) (NCBI accession no MH156812). B a 249 bp sequence of IGF-I gene for Holstein Frisian genotype (CC) (NCBI accession no. MH156811). C The chromatogram of the sequenced 5’-noncoding region IGF-I gene showing homozygote (CC and TT) and heterozygote (CT) genotypes
Serum periparturient concentration and their association with IGF-I genotypes
An evidence for a significant (p > 0.01) association between C and T mutations in position 472 of the IGF-I gene and its serum concentration in HF cows in Egypt was found. The highest serum concentration of IGF-I was found in CC genotype cows in which CC genotype had significantly higher (p > 0.01) concentration of IGF-I at 20 d prepartum (92.44 µg/L) compared to those with TT genotype (74.58 µg/L). Also, the values of IGF-I concentration for 20 d before calving and 50 d postpartum were higher (p > 0.01) in cows with CC genotype compared to those with TT genotype. The C/T transition for the trend of IGF-I concentration between the CC and TT genotype was significantly different (p > 0.01) during the 20 d prepartum and 50 d postpartum (Table 2).
Growth hormone (GH) gene
PCR amplification of bovine GH gene
The fragment of 432 bp of the GH gene was successfully amplified from the genomic DNA of the tested cows (Fig. 1B &C). In the present study, the amplification of the GH gene produced a specific band of 432 bp and a nonspecific fragment of 120 bp (Fig. 1B). So, PCR purification from the gel was performed to isolate the specific fragment (Fig. 1C).
Genotyping of GH gene by RFLP and nucleotide and Protein sequence
The site recognized by the AluI enzyme restriction was AG^CT bases. Three AluI restriction sites produced fragment lengths of 20, 147 and 265 bp, known as [G] allele (Glycine) and three AluI restriction sites that produced fragment lengths of 20, 51, 96 and 265 bp, known as [A] allele (Alanine). A heterozygous AG genotype was then identified by fragment lengths of 20, 51, 96, 147, and 265 bp. GG genotype was not present for all samples (Fig. 4 A and B). Genotyping results produced only two genotypes AA and AG, but not GG. The genotypes frequencies were 77 and 23% for AA and AG, respectively. The frequency of A allele was very high (81%) compared to that of G allele (19%) (Table 1).

A Agarose gel electrophoresis showing RFLP-AluI restriction pattern of Holstein Frisian cattle in GH gene Lane M: 50-bp ladder marker. Lanes AA: Homozygous (AA) genotype with 4 restricted fragments at 265, 96, 51 and 20 bp. Lanes AG: heterozygote (AG) genotype with 5 restricted fragments at 265, 147,96, 51 and 20 bp. B A 432 bp sequence of GH gene for Holstein Frisian genotype (AA) (NCBI accession no. MH156810.1) with RFLP-AluI restriction pattern (4 restricted fragments at 265, 96, 51 and 20 bp) and restriction sites (AG^CT). C Amino acids comparison of amplified GH gene of tested Holstein Friesian using (MEGA-11) Molecular Evolutionary Genetics Analysis, and its site https://www.megasoftware.net
A 432 fragment from Intron 4, part of exon 4 and part of exon 5 was sequenced. Data was generated and manged by BioEdit V.7.7. (https://bioedit.software.informer.com/7.2) and GeneScan (http://hollywood.mit.edu/cgi-bin/genscanw_py.cgi) with a minimum ORF size of 20 and the start codon AGT, and the comparison of amino acids was done by using MEGA 11 (https://www.megasoftware.net). The amino acids sequence of genotype (AA) was different from that of genotype (AG) in one amino acid. The changed amino acid was Alanine for (AA) compared to Glycine for (AG) (Fig. 4C). Nucleotide sequencing of the amplified fragment of the bovine GH gene for (AA) genotype was done and submitted to the NCBI GenBank (Accession no. MH156810.1), (Fig. 5).

A A 432 bp sequence of GH gene for Holstein Frisian genotype (AA) (NCBI accession no. MH156810.1), with 4 restricted fragments at 265, 96, 51 and 20 bp. B A 432 bp sequence of GH gene for Holstein Frisian genotype (GG) with 3 restricted fragments at 20, 265 and 147
The relationship between productive and reproductive performance and the prevalence of IGF-I and GH genotypes
Insulin-like Growth Factor -1 (IGF-I)
Table 3 shows the relationship between the IGF-I-SnaBI polymorphism genotypes and productive and reproduction traits. Concerning the IGF-I gene, there were three genotypes; TT (n = 517), TC (n = 392) and CC (n = 91). The TC genotype achieved the shortest (p > 0.01) LP and DPRY followed by CC and then TT genotypes, respectively. In this aspect, the TC genotype recorded the highest TMY and 305d-MY (p > 0.01) followed by TT and then CC. Regarding the percentage of fat and protein, CC genotype was the highest (p > 0.05) followed by TT and then TC. Also, CC genotype recorded the lowest value for EC followed by CT and then TT genotypes. There were no significant differences in SCC and pH among different genotypes of IGF-I gene (Table 3).
As for reproductive performance, the TT genotype recorded the shortest (p > 0.05) FPE compared to TC and CC which had non-significantly different FPE. On the other side, the shortest DOPN and CI were observed for TC genotype (p > 0.01) compared to TT and CC genotypes. However, no significant differences in AFC, NI and GL were observed among different genotypes of IGF-I gene.
In the current study, the novel detected SNPs and their amino acids sequence for IGF-I gene could be considered as candidate SNPs for milk yield and composition, and for reproductive performance in HF cows under the subtropical conditions of Egypt (Table 1). For instance, one SNP was detected for 472 C > T of IGF-I gene in the high-producing cows (with TC genotype), this sequence resulted in a significant differentiation in LP, DPRY, TMY and 305d-MY (p > 0.01) comparing to other sequences (Table 3).
Growth hormone (GH) gene
Table 4 shows the relationship between GH/AluI polymorphism genotypes and productive and reproduction traits. There were two genotypes; AA (n = 776) and AG (n = 224) for the GH gene of the studied cows. The cows with AG genotype recorded shorter (p > 0.05) LP and DPRY compared to AA genotype. In this aspect, AG genotype achieved the highest (p > 0.01) TMY and 305d-MY compared to AA. On the other side, AA recorded the highest (p > 0.01) percentage of fat and protein. Also, AA recorded the lowest value for EC compared to AG genotype. There were no significant differences in SCC and pH among different genotypes in the GH gene.
Regarding reproductive performance, the AA recorded the shortest (p > 0.05) FPE compared to AG. On the other hand, the shortest DOPN and CI were observed on AG genotype (p > 0.01) compared to AA. There were no significant differences in AFC, NI and GL among different genotypes of GH gene.
Another novel detected SNP for 1758 C > G (Alanine / Glycine) substitution (Fig. 5) in GH gene of high-producing cows (with AG genotype) was a mutation which led to changes in the amino acid sequence; Glycine for (G) > Alanine for (A) (Fig. 4C). This sequence resulted in a significant differentiation in LP, DPRY, TMY and 305d-MY (p > 0.01) comparing to another different sequence (Table 4).
Table 5 presents the P-values for various factors influencing productive and reproductive traits in HF cows. P-values measure the probability of obtaining the observed data, or more extreme results if the null hypothesis (no association between the factor and the trait) is true. A p-value less than the predetermined significance level (0.05 or 0.01) suggests statistical significance, indicating a strong association between the respective factor and the traits under consideration. The p-values in Table 5 help assess the strength of the evidence supporting the relationship between each factor and the productive and reproductive traits of HF cows. Factors with low p-values indicate a higher degree of significance and emphasize their importance in influencing the cow's performance. On the other hand, factors with higher p-values suggest a weaker association or lack of statistically significant impact on the traits. Interpreting the p-values within the context of the specific factors examined is crucial to drawing meaningful conclusions. For example, if a particular factor related to IGF-I and GH genotypes exhibits a low p-value, it implies that it significantly affects both productive (e.g., milk production or composition) and reproductive (e.g., calving interval) traits. This finding could have significant implications for dairy farmers, as it highlights the importance of considering this factor in their breeding or management strategies. Conversely, factors with high P-values may not have a substantial impact on productive and reproductive traits.
Discussion
The implementation of genomics offers a great opportunity for dairy cattle production improvement through increased genetic progress and inclusion of new traits of economic importance in the selection programs. The identification of potential SNPs in selected groups will make the exploitation of novel genomic selection methodology in farm animals possible. Better genetic characterization of breeds under different conditions/environments can help increase selection intensity and decrease generation interval [22, 24].
Studying the polymorphism in IGF-I and GH genes are important for the dairy cattle industry as they are involved in productive and productive traits [24]. There is evidence suggesting that specific variants in these two genes may possess either positive or negative associations with milk production, reproductive performance, and growth rate in dairy cattle in addition to some other species [22, 24]. Identifying polymorphisms should aid farmers and animal breeders in selecting stocks with advantageous gene variants for rapid improvement in productivity and, hence, profitability [15, 45, 46]. Additionally, verification of the genetic basis underlying the inheritance of these traits should help in developing targeted management strategies that maximize the dairy herd performance [20]. Therefore, exploring the polymorphism in IGF-I and GH genes is crucial for the development of the modern dairy industry.
IGF-I gene encodes a hormone similar in structure to insulin (Fig. S3), while, GH gene encodes GH hormone (Fig. S4). GH hormone, produced in the anterior pituitary gland, stimulates the release of IGF-I hormone from the liver and is of critical importance in the control of partitioning and nutrient utilization for growth, fertility, lactogenesis, and some necessary processes like development in mammalian [14, 33] (Fig. S5).
Insulin-like Growth Factor -1 (IGF-I)
In the present study, polymorphisms among the studied HF cows for IGF-I and GH genes were detected. Also, an association of SNPs in the 5’-noncoding region of the IGF-I gene and the intron 4, part of exon 4 and part of exon 5 of the GH gene with the selected traits of milk production and composition, and reproductive performance in HF cows under subtropical conditions of Egypt were revealed.
Moreover, three genotypes for IGF-I gene (TT, TC and CC) were uncovered in the tested HF cows. Cows with the TC genotype were more productive for most of the milk yield traits, unlike the milk composition traits in which the CC genotype recorded the highest percentages for fat and protein in the milk (Table 3). This agrees with the results of Siadkowska et al. [47] obtained on 662 Polish-HF cows when examining the association between IGF-I gene polymorphism and the traits of feed intake, meat production, growth rate, and milk production. The heterozygous TC genotype yielded more fat-corrected milk, value-corrected milk, milk fat, and milk protein. Also, Silveira et al. [18] working on HF cows, Czerniawska-Piątkowska et al. [48] working on Holstein and Jersey cows [20] working on Madura cattle and Hartanto et al. [29] working on Jawa-Brebes cows, and obtained differentiated milk yield and composition and reproductive performance in response to different IGF-I and GH genotypes.
The effects of IGF-I gene polymorphism on cattle production, growth performance and developmental processes including metabolism and nutrient partitioning have been well-established and documented [9, 49]. The polymorphisms of IGF-I gene were first reported in Angus cattle in 1997 using the SSCP technique, and then identified as a C/T transition at position (P472) relative to the start of the transcription site at position 512 bp upstream from the ATG codon; according to the GenBank sequence (AF210383) [43].
The SNP of IGF-I gene described in the current study was similar to that found in the subtropical two strains of Nyalawi and Metairie cows [44], also, was in agreement with that advocated in Bos taurus, according to the GenBank (AF404761 and KF202095). In the same aspect, Mullen et al. [14] reported that nine SNPs were identified across a panel of twenty- two dairy and beef cattle by sequence analysis of the 5′ promoter, 3′ regulatory regions, intronic and encompassing 5 kb of IGF-I gene. Also, several SNPs were identified in the 3′ region of IGF-I and were associated (p < 0.05) with chest width and functional survival. On the other side, four out of nine SNPs were identified for their association with protein and fat yield, milk fat concentration, SCC, carcass conformation, and carcass fat (p < 0.05). These findings side by side with the results of the present investigation show strong effects of IGF-I polymorphism on milk production, fat yield and functional survival in cattle.
In the current study, the observed frequencies of different genotypes of IGF-I gene in HF cattle were (TT = 0.52), (TC = 0.39) and (CC = 0.09). This was in agreement with the results of Yazdanpanah et al. [50] who reported three frequencies of genotypes; (TT = 0.83), (TC = 0.14) and (CC = 0.03) on Najdi cattle (n = 84). Also, Nicolini et al. [31] confirmed that the frequencies of IGF-I genotypes for HF cows (n = 70) were (TT = 0.31), (TC = 0.54) and (CC = 0.14).
On the contrary, Nicolini et al. [31] reported that there was no effect of the different IGF-I genotypes on body condition change. In this regard, Omer et al. [44] reported no differences between two strains of Baggara zebu cattle at position 472 C > T of the IGF-I gene promoter. The mutant homozygote (TT) was detected in the Mesairi cattle only with a frequency of 0.016. While the heterozygote (CT) genotype existed with low allele frequencies (0.079 and 0.068) in Mesairi and Nyalawi breeds respectively. Also, Szewczuk et al. [51] reported that there was no association between IGF-I/SnaBI and dairy production traits in Polish Holstein cattle.
Concerning reproductive traits, this study revealed that IGF-I/SnaBI of TT genotype differed significantly (p < 0.05) from that of TC for FPE, whereas IGF-I/SnaBI of TC genotype had significantly shorter DOPN and CI (p < 0.01) compared to the other genotypes. Additionally, there were no significant differences observed among different genotypes of IGF-I gene for AFC, NI, and GL.
Concerning IGF-I concentration in the blood, an association was found between different IGF-I genotypes and the blood IGF-I concentration in HF cows. The findings of this study provide compelling evidence for a significant association between mutations in position 472 of the IGF-I gene and its serum concentration in HF cows in Egypt. Specifically, the CC genotype was found to have the highest serum concentration of IGF-I, with significantly higher levels compared to the TT genotype cows at 20 d prepartum. The data also showed that the CC genotype cows had higher IGF-I concentrations both 20 d before calving and 50 d postpartum compared to the TT genotype cows. Furthermore, the C/T transition for the trend of IGF-I concentration between the CC and TT genotypes was significantly different during the 20 d prepartum and 50 d postpartum. These results are summarized in Table 2, which illustrates the comparison between the serum concentrations of different IGF-I genotypes in HF dairy cows during periparturient periods. Overall, these findings indicate a strong link between the IGF-I gene mutations and serum concentration, highlighting the potential role of genetic factors in regulating IGF-I levels in HF cows.
Briefly, the highest serum concentration of IGF-I was found in CC followed by CT as compared to TT genotypes (Table 2). This is in agreement with the results of Mirzaei et al. [52] who confirmed that the highest serum concentration of IGF-I in Polish HF cows was found in CC followed by CT and then TT genotypes, which also, was in agreement with the reports of Mehmannavaz et al. [34], Bonakdar et al. [53] and Mirzaei et al. [52] on the Iranian Holstein cattle. In this regard, Gobikrushanth et al. [54] conducted a study to investigate the factors associated with the serum concentration of IGF-I in dairy cows and its relationship with reproductive outcomes. The study involved 647 lactating Holstein cows and identified various factors, such as herd, age, parity, pre-calving body condition score, and season of blood sampling, that influenced serum IGF-I concentrations. The researchers found that serum IGF-I concentration during the first week postpartum was higher in cyclic multiparous cows compared to acyclic ones, but did not show a significant association with ovarian cyclicity status in primiparous cows. The study also established optimal serum IGF-I thresholds predictive of pregnancy to first artificial insemination (P/AI) for primiparous and multiparous cows. Primiparous cows with high IGF-I had greater odds of P/AI and a tendency for higher pregnancy risk up to 150 d postpartum compared to those with low IGF-I. Similarly, multiparous cows with high IGF-I had increased odds of P/AI. Additionally, the researchers identified multiple SNPs associated with variation in serum IGF-I concentration, some of which were in linkage disequilibrium with candidate genes related to fertility.
On the other side, Wathes et al. [55] investigated the connection between negative energy balance (EB) and immune defense in peripartum dairy cows. The study found that cows with lower IGF-I levels, indicating poor EB, experienced more health problems, altered leukocyte functionality, and reduced milk production. In contrast, cows with higher IGF-I levels exhibited better immune function and milk production.
Growth hormone (GH) gene
Internationally, three different genotypes for GH-AluI were detected in Iranian-Holstein cattle [27, 56]. In the present study, only two genotypes for the GH gene (AA and AG) were discovered in the studied cows. This is in agreement with the results of Kiyici et al. [27] on Holstein dairy cattle and with Pereira et al. [26], Curi et al. [57] and Misrianti et al. [28] who obtained only two genotypes in Brazilian Canchim, Brazilian Zebu and HF dairy cattle, respectively.
In the present investigation, the frequencies of genotypes were determined to be 77% and 23% for AA and AG, respectively. Notably, the A allele displayed a pronounced prevalence of 81% in contrast to the G allele, which exhibited a relatively modest frequency of 19% (Table 1). In this aspect, several studies have investigated the frequencies of genotypes and alleles of GH gene in different populations Lucy et al. [58] found variable frequencies of the two alleles across breeds. For HF cows, the frequencies were 0.93 and 0.07, for Brown Swiss were 1 and 0, for Jersey were 0.56 and 0.44, for Guernsey were 0.92 and 0.08, and for Ayrshire they were 0.79 and 0.21, respectively. In Holstein sires used for Artificial Insemination (AI), the frequencies of A and G alleles were 0.96 and 0.04. In another study, Kovacs et al. [59] observed genotypic frequencies of 87.05%, 12.40%, and 0.55% for AA, AG and GG genotypes in Hungarian-Holstein–Friesian (HHF). Dario et al. [60] reported 61%, 22%, and 17% frequencies for AG, AA and GG genotypes, respectively. Balogh et al. [61] found that the frequencies of AA, AG and GG genotypes were 83%, 17% and 0% in a specific population of cows. Hadi et al. [62] reported that the frequencies of AG, AA and GG genotypes were 61%, 39%, and 0%, respectively. They also observed that the A allele had a higher frequency (69%) compared to the G allele (31%).
In the current study, intron 4, part of exon 4 and part of exon 5 have been investigated for the GH gene. Previous researchers have identified polymorphisms in the 3rd and 4th introns, and 5th exon and the promoter of the GH gene in cattle. Also, recent reports show a significant relationship between polymorphisms in the GH gene and lactation performance in cattle [14, 24, 27].
This investigation revealed a genetic variation found between A and G alleles, due to mutation at the 1758 base resulting in the changing base from C to G (Fig. 5). Where the homozygous (AA) genotype was with 4 restricted fragments at 265, 96, 51 and 20 bp. While the heterozygote (AG) genotype was with 5 restricted fragments at 265, 147, 96, 51 and 20 bp (Fig. 4A). These findings are similar to the results of Misrianti et al. [28].
In the present investigation, cows with the GH-AluI-AG genotype were more productive for most of the milk yield traits unlike milk composition traits, where the GH-AluI-AA genotype recorded the highest percentages for fat and protein in milk (Table 4). This agrees with the results of Kovacs et al. [59] who confirmed that AG genotype showed to be advantageous for 305d-MY, while AA genotype recorded the highest percentages for fat and protein. Also, the current findings are in agreement with the results of Nugroho et al. [20] on Madura cattle who reported that the GH-AluI-AA genotype was discovered to have higher performance in milk fat and protein content, body and carcass weights compared the GH-AluI-AG genotype, while the GH-AluI-AG genotype was found to have higher performance in milk yield compared to GH-AluI-AA genotype. Moreover, Yardibi et al. [63] reported that the variant genotypes; AA, AG and GG of the GH-AluI gene had a positive correlation with percentages of fat and protein contents of milk. On the contrary, Shaidullin [64] confirmed that the highest level of milk productivity was found in animals with GH-AluI of AA genotype with a significant advantage over full-aged cows, GH-AluI of AG and GH-AluI of GG in terms of milk yield of 280 kg (p < 0.001) and 509 kg (p < 0.001), by the amount of milk fat of 9.1 kg (p < 0.001) and 18.5 kg (p < 0.01), by the amount of milk protein of 7.9 kg (p < 0.01) and 14.8 kg (P < 0.01).
In the present study, the heterozygous AG genotype exhibited changes in the amino acid sequence in position (P114) as Glycine replaced Alanine in AA genotype. In this regard, Lucy et al. [58] and Lucy et al. [30] reported that cytosine (C) substituted guanine (G) at position 2141 causing an amino acid change from Alanine to Glycine at residue 127 of the GH polypeptide. Moreover, the associations between milk production traits and Glycine (G) allele have been confirmed [27, 49, 59]. On the other hand, a substitution favouring Alanine (A) allele was achieved [64] in several cattle breeds.
Concerning reproductive performance, the GH-AluI-AA genotype showed differences in FPE (p < 0.05) compared to the GH-AluI-AG genotype. However, the latter had significantly shorter DOPN and CI (p < 0.01) compared to the GH-AluI-AA genotype. Additionally, no significant differences were observed among different genotypes of the GH gene for AFC, NI and GL. This agrees with the results of Amiri et al. [65] who reported that the individuals with the GH-AluI-AG genotype had significantly shorter DOPN and CI (p < 0.01) compared to the GH-AluI-AA genotype.
However, the results are contradictory with the studies by Lechniak et al. [66] and Lechniak et al. [25] who did not detect any significant relationship between GH-AluI gene polymorphism and reproductive performance especially for bulls’ sperm characteristics or parameters of in-vitro fertilization and embryo development.
Briefly, the greatest TMY and 305d-MY values and the best reproductive performance were observed on IGF-I-SnaBI-TC and GH-AluI-AG genotyped cows. While the greatest % fat and % protein values were observed on IGF-I-SnaBI-CC and GH-AluI-AA genotyped cows (Tables; 3 and 4).
On the other side, Lucy et al. [58] revealed that dairy cows having small mature size like Jersey breed had high frequency of GH-G allele, and those having large mature size like Holstein breed had high frequency of GH-A allele. Otherwise, Balogh et al. [61] reported that animals carrying genotype AG were prone to higher basal insulin levels (p = 0.064), a longer time to reach half of the maximal and basal insulin concentrations (p = 0.035 and p = 0.054, respectively) and larger insulin area under the curve (p = 0.032). Expanding on this relationship, Mullen et al. [67] confirmed that there was an association between GH genotypes and carcass traits as well as SCC and body condition score (BCS). Also, Mullen et al. [14] found an association between genetic variation in GH gene and fertility, pregnancy rate and overall pregnancy rate. Moreover, Hadi et al. [62] reported that GH-AluI-AA genotype reduced dystocia, compared to GH-AluI-AG. Overall, these studies shed light on the intricate associations between GH genotypes and various phenotypic traits, including size, insulin levels, carcass traits, fertility, and dystocia.
GH-IGF-I system controls
IGF-I and GH genotypes are of significant importance when it comes to milk production and quality as well as fertility performance in HF dairy cattle. The somatotrophic axis, which includes GH and IGF-I, plays a crucial role in regulating growth and development in cattle, affecting traits such as milk yield, growth rate, body composition, and fertility (Fig. S5).
In this regard, reliable studies confirmed that the somatotrophic axis, which essentially consists of growth hormone-releasing hormone (GHRH), GH, IGF-I and II and their associated binding proteins (GHBP, IGFBP1-6) and receptors (GHRHR, GHR, IGF-IR and IGF-IIR), plays a key role in the metabolism and physiology of mammalian growth [21, 68]. The somatotrophic axis (GH-IGF) is a key regulator of animal growth and development and affects performance traits that include milk yield, growth rate, body composition, and fertility [14, 19, 23], (Fig. S6). GH and IGF-I are major regulators of postnatal metabolism, growth and consequently play critical roles in the control of mammary gland development, lactation, growth processes, and fertility in cattle [30, 69]. The actions of GH vary significantly in several physiological states [70], but the net effect of this hormone throughout early lactation supports a helpful role for the indirect actions of GH on lipolysis and gluconeogenesis [11] and attenuated growth-promoting actions and support by IGF-I in peripheral tissues [71]. Within the dairy cow, the per-parturient reduction in IGF-I synthesis is related to a concomitant reduction in the liver-specific GH receptor type 1A (GHR1A) [72].
In light of the above facts and findings in the present investigations, understanding the impact of IGF-I and GH genotypes on milk production, quality, and fertility in HF dairy cattle, especially under subtropical conditions in Egypt is essential.
Conclusions
In conclusion, studying IGF-I and GH genes has shed light on their roles in livestock growth and development. The genetic regulation of these genes has increased the efficiency of selecting superior-value animals in meat, production, and reproduction programs. The molecular genetic studies of IGF-I and GH genes have demonstrated their potential for livestock improvement. Greatest milk yield and composition values, and reproductive performance were observed on IGF-I-SnaBI-TC and GH-AluI-AG genotyped individuals. While the greatest % fat and % protein values were observed on IGF-I-SnaBI-CC and GH-AluI-AA genotyped individuals. The genetic variation of these genes can be utilized in selecting animals with superior milk yield, growth performance, feed efficiency and meat quality. Continued research in the area of genetic regulation of these genes is necessary to further explore their roles in livestock breeding and production.
Methods
Animals and Sampling
The present investigation is part of a project aiming to assess HF cattle (n = 1000) under subtropical conditions (Egypt) in order to aid the characterization of cattle genetic resources and genome analysis in this area for milk yield and composition, and reproductive performance.
A total of 1000 HF dairy cattle from El-Alamia commercial dairy farm (belonging to Universal Company for Agricultural Development and Soil Reclamation, herd located at Nubaria region in the K 90 Alex-Cairo desert road, Egypt) (Fig. S7) were investigated for milk production and composition, and reproductive performance.
Management of animals
The calving period for the tested animals was constrained between January 2016 and January 2018. The experimental cows were 76 ± 7.25 months in age, with an average live weight of 750 ± 50.49 kg, randomly selected from the respective groups of contemporaries born within 7–11 months. Cows were housed free in open semi-shaded yards, nourished under the prevailing feeding conditions. The cows were fed according to the INRA feeding system for ruminants on a complete Total Mixed Ration (TMR) diet consisting mainly of wilted grass silage, corn silage, beet pulp, cotton seed, soybean, barley, and concentrate mix and vitamin mixture, supplemented with minerals. Water was available excessively. The cows were raised under consistent conditions of nutrition and weather, and milked twice daily at 07:00 and 16:00 h and were classified according to milk yield, specifically the daily milk yield (DMY) into; high producer cows (n = 280) with a DMY above 35 kg, medium producer cows (n = 318) with a DMY between 25 ~ 35 kg, and low producer cows (n = 402) with a DMY below 25 kg.
Traits of concern
The traits of concern were; 1) Lactation characteristics: lactation length (LP), dry period length (DPRY), total milk yield (TMY) and adjusted milk yield (305d-MY). 2) Milk composition: fat percentage (% fat) and protein percentage (% protein). 3) Milk Quality; somatic cell count (SCC), electrical conductivity (EC) and acidity (pH). 4) Reproductive performance: days open (DOPN), calving interval (CI), gestation length (GL), the first postpartum estrus (FPE) and the number of inseminations (NI). These traits were recorded professionally for the studied cows for several lactations from the 1st to 4th. 5) Serum concentration of IGF-I (µg/L) for different milk yield/groups of HF dairy cows.
Genes of concern
In the current study, the polymorphism in IGF-I and GH genes of HF cows were investigated for their association with milk yield and composition, and reproductive performance of cattle.
Milk samples
Two 50 ml milk samples were taken monthly on a specific test day from the morning (07:00 am) and evening milkings (4:00 pm) of each cow and were kept at 4 °C until used to determine milk composition until the fourth month of lactation. A total of 2770 milk samples were obtained and utilized during the 4th lactation of cows.
Milk SCC measurements were taken with the help of The NucleoCounter®-SCC-100™ equipment (Chemometec, Bohemia, New York, USA), while, EC and pH measurements were taken with the help of Milkana Multi-Test milk analyser (http://mtm-solutions.com/en/products/detail/Milkana-MULTI---TEST).
Blood samples
Blood samples for genetic analysis
A blood sample of 5 ml from venous blood was collected separately from the jugular vein of each tested cow using venojects. Blood samples were treated with 0.5 ml of 2.7% EDTA (Pspark, U.K), as an anticoagulant, kept in an icebox and transferred immediately to the lab and stored at -80 °C up to the genetic analysis. All procedures carried out with the use of animals were approved by the Ethics Commission, permission No. AU082211211117), Faculty of Agriculture (Al-Shatby), Alexandria University, Egypt.
Blood samples for serum IGF-I concentration
Also, a total of 280 healthy cows from the three different IGF-I genotypes TT (n = 100), TC (n = 100) and CC (n = 80) were chosen randomly to measure periparturient serum IGF-I concentration. The blood samples were collected from the jugular vein of each cow into a separate tube (Guangzhou Improve Medical Tech. Co. Ltd., China) without anticoagulants for biochemical indices calculation. The blood samples have been taken 20 days before the expected calving date (prepartum), and 25 and 50 d postpartum of the studied cows in the spring season (March–May). Within 15–20 min. after collection, serum was separated by centrifugation (1600 × g/13 min) and stored at -20 °C until further analysis. IGF-I in the serum was measured using the IGF-I-ELISA Assay kit (Eagle Biosciences, Boston-Massachusetts, USA). The inter and intra-assay coefficients of variation were 6.8% and 7.4%, respectively, and the sensitivity was 3.3 ng/mL.
DNA isolation, amplification, manipulation and sequencing
DNA isolation
Total genomic DNA was isolated from the blood samples (n = 1000) of the selected experimental cows using a DNA isolation kit (Tiangen Biotech, Beijing, China). The DNA samples were separated by electrophoresis on 1.0–1.2% agarose in 0.5 × TBE buffer according to Sambrook and Fritsch [73] after adding 0.5 μg/ml ethidium-bromide for quality assessment purpose. The electrophoresis run was performed using apparatus with a power supply and visualized by an ultraviolet transilluminator and Gel-documentation system (Chemi.Doc™ XRS + with Image Lab™ Software, BIO-RAD, USA). The purity and integrity of DNA were appropriate, and the OD260/280 was 1.82.
Amplification and manipulation
The specificity of the PCR primers targeting the IGF-I gene (249 bp fragment, 5’-noncoding region of the bovine IGF-1 gene) and GH gene (432 bp fragment, fragment from Intron 4, part of exon 4 and part of exon 5) were previously tested by Ge et al. [43] and Balogh et al. [61], respectively. The primer sequences, amplified region and product size of IGF-I and GH genes are shown in Table 6. The primers were synthesized by (Shanghai-Sangon Biolo. Engin. Tech. & Ser. Co., Ltd). The amplification was performed using (Green-Super.mix, TaKaRa, Japan). The PCR conditions are shown in Table 6. The amplification was carried out using a Thermo-cycler Gene Amp 6700 (Applied Bio-system, USA) and the products were separated by electrophoresis on 0.8% agarose and visualized by UV trans-illuminator and gel documentation system (Chemi.Doc™ XRS + with Image Lab™ Software, BIO-RAD, USA).
Nucleotide sequence analysis
Automated DNA sequence analysis was carried out on both strands by the DNA sequencing service lab of the Korean Research Institute of Bioscience and Biotechnology with an ABI Prism 3100 apparatus for both IGF-I and GH genes. Database similarity searches were performed with the FASTA network service at the National Centre for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov). Also, the resulting sequences were analysed using MEGA 11, and Blast 2.0 software to detect SNPs between sequences. The sequences were deposited in GenBank. Moreover, the results of endonuclease restriction were carried out using FastPCR (http://primerdigital.com/fastpcr.html). Analysis of translated protein of IGF-I and GH sequences of the tested cows was generated and manged by BioEdit V.7.7. (https://bioedit.software.informer.com/7.2) and GeneScan (http://hollywood.mit.edu/cgi-bin/genscanw_py.cgi) with a minimum ORF size of 20 and the start codon AGT.
Restriction-fragment-length-polymorphism (RFLP) and electrophoresis
The RFLP was used to detect genotyping differences between and within tested cows using the PCR of target genes. The PCR amplicons of the IGF-I (249 bp) gene were digested with SnaBI (Jena Bioscience, Germany) and of GH (432 bp) with AluI (Bio-search Technologies, USA) separately. Defining restriction sites, before digestion with restriction enzymes was achieved by the NEB cutter program (http://www.labtools.us/nebcutter-v2-0). The RFLP-PCR reaction volume was 25 μl, consisting of 12 μl H2O, 2 μl 10X HaeIII buffer (Jena Bioscience, Germany), 1 μl (5 unit/ul) restriction enzyme in addition to 10 μl amplified DNA. All reactions were incubated at 37 °C for 16 h. Twenty μl of each reaction were separated by electrophoresis on 2.5% agarose gel and visualized by UV trans-illuminator and gel documentation system (Chemi.Doc™ XRS + with Image Lab™ Software, BIO-RAD, USA).
Statistical analysis
Analysis of variance (ANOVA) and duncan test
The analysis primarily consisted of two steps: analysis of variance (ANOVA) and post-hoc tests. First, ANOVA was performed to evaluate whether significant differences existed in the serum concentration of IGF-I among the three genotypes across the periparturient periods (20 days prepartum, 25 days postpartum, and 50 days postpartum). Also, Significant differences among means were tested using the Duncan test.
Normality assessment and genotype effects analysis
All milk yield, milk composition, reproductive traits, and the serum concentration of IGF-I variables were tested for normality by Shapiro–Wilk test from the UNIVARIATE procedure of SAS (SAS, 2009), and the results indicated that all data were distributed normally (W ≥ 0.90). The genotype effects were analyzed using the GLM procedure of SAS by adapting the following model:
where Yijklmnop is the mean value of the variable; Ri is the random effect of sire (i = 1,…..100 +), Fj is the fixed effect of the IGF-I genotype (j = 1,…., 3), Gk is the fixed effect of the GH genotype (k = 1 and 2), Al is the fixed effect of the calving year (l = 2016, 2017 and 2018), Sm is the fixed effect of the calving season (m = 1,…. 4), Ln is the fixed effect of the parity (n = 1,2,3,4), Co is the fixed effect of the milk production levels category (m = low, medium, high) and eijklmnop is the residual error. Significant differences among means were tested using the Duncan test.
Genetic indices and equilibrium analysis
The genetic indices of the studied animals: Heterozygosity (Ho) and Heterozygosity expected (HE) were calculated according to Nei’s methods [74, 75]. Also, the Hardy–Weinberg equilibrium (HWE) was determined using Michael H. Court’s (2005–2008) calculator [76].
Availability of data and materials
The datasets generated and analyzed in this study are accessible in the Genbank repository with the accession numbers;
* NCBI Accession no.:MH156810.1 (https://www.ncbi.nlm.nih.gov/nuccore/MH156810.1),
* NCBI Accession no.:MH156811.1 (https://www.ncbi.nlm.nih.gov/nuccore/MH156811.1),
* NCBI Accession no.:MH156812.1 (https://www.ncbi.nlm.nih.gov/nuccore/MH156812.1).
Additional relevant information and results can be found in this manuscript and its supplementary files, including Figures, Tables, and Supplementary Figures.
Abbreviations
- IGF-I :
-
Insulin-like Growth Factor-I gene
- GH :
-
Growth Hormone gene
- SNPs:
-
Single Nucleotide polymorphisms
- HF:
-
Holstein–Friesian
- SCC:
-
Somatic Cell Count
- LP:
-
Lactation length
- DPRY:
-
Dry period length
- TMY:
-
Total milk yield
- 305d-MY:
-
Adjusted milk yield
- % fat:
-
Fat percentage
- % protein:
-
Protein percentage
- EC:
-
Electrical conductivity
- pH:
-
Acidity
- DOPN:
-
Days open
- CI:
-
Calving interval
- GL:
-
Gestation length
- FPE:
-
First postpartum estrus
- NI:
-
Number of inseminations
References
Forero DA, Chand V. Methods in molecular biology and genetics: looking to the future. BMC Res Notes. 2023;16(1):26.
Džermeikaitė K, Bačėninaitė D, Antanaitis R. Innovations in Cattle farming: application of innovative technologies and sensors in the diagnosis of diseases. Animals. 2023;13(5): 780.
Saleh AA, Rashad AM, Hassanine NN, Sharaby MA. Candidate genes and signature of selection associated with different biological aspects and general characteristics of goat. Emerging Animal Species. 2022;5:100013.
Barrera SS, Naranjo-Gomez JS, Rondón-Barragán IS. Thermoprotective molecules: effect of insulin-like growth factor type I (IGF-1) in cattle oocytes exposed to high temperatures. Heliyon. 2023;11;9(3):e14375. https://doi.org/10.1016/j.heliyon.2023.e14375.
Yousief MY, Owaid JM, Jaafer AA. Relationship between polymorphisms of growth hormone gene with milk yield and its components in Iraqi cattle bred of Janoubi, Cross and Holstein. Iran Jo Ichthyology. 2023;10:144–9.
Kader Esen V, Esen S. Association of the IGFI 5′ UTR Polymorphism in meat-type sheep breeds considering growth, body size, slaughter, and meat quality traits in Turkey. Veterinary Sciences. 2023;10(4):270.
Al-Sharif MM, Radwan HA, Hendam BM, Ateya AI. Exploring single nucleotide polymorphisms in GH, IGF-I, MC4R and DGAT1 genes as predictors for growth performance in dromedary camel using multiple linear regression analysis. Small Rumin Res. 2022;207: 106619.
Mege RA, Montolalu FM, Manampiring N, Rayer DJJ. GHR mRNA expression in the liver and muscles of local pigs from The Islands in North Sulawesi, Indonesia. International Joint Conference on Science and Technology. 2020;2020:190–5.
Dar MR, Singh M, Thakur S, Verma A. Exploring the relationship between polymorphisms of leptin and IGF-1 genes with milk yield in indicine and taurine crossbred cows. Trop Anim Health Prod. 2021;53:1–8.
van Doorn J. Insulin-like growth factor-II and bioactive proteins containing a part of the E-domain of pro-insulin-like growth factor-II. BioFactors. 2020;46(4):563–78.
Soliman A, Ahmad S, Hamed N, Alyafei F, Alaaraj N, Soliman N. Maternal, placental, and fetal Insulin-Like Growth Factor-I (IGF-I) and IGF Binding proteins (IGFBPs) in Diabetic pregnancies: Effects on fetal growth and birth size. World Journal of Advanced Research and Reviews. 2023;17(2):287–95.
Fang J, Zhang X, Chen X, Wang Z, Zheng S, Cheng Y, Liu S, Hao L. The role of insulin-like growth factor-1 in bone remodeling: a review. International Journal of Biological Macromolecules. 2023;238:124125.
Zhang W, Lin S, Jiao Z, An L, Xie T, Wu J, Zhang L. The mouse CircGHR regulates proliferation, differentiation and apoptosis of hepatocytes and myoblasts. Genes. 2023;14(6): 1207.
Mullen MP, Berry DP, Howard DJ, Diskin MG, Lynch CO, Giblin L, Kenny DA, Magee DA, Meade KG, Waters SM. Single nucleotide polymorphisms in the insulin-like growth factor 1 (IGF-1) gene are associated with performance in Holstein-Friesian dairy cattle. Front Genet. 2011;2:3.
Kumar S, Dahiya S, Magotra A, Ratwan P, Bangar Y. Influence of single nucleotide polymorphism in the IGF-I gene on performance and conformation traits in Munjal sheep. Zygote. 2023;31(1):70–7.
Worku HA. The Role of Exogenous Bovine Somatotropin on Lactation and Nutrient Mobilization in Dairy Animals: A Review. Glob J Anim Sci Res. 2023;11(1). http://www.gjasr.com/index.php/GJASR/article/view/158.
Knop R, Cernescu H. Effects of negative energy balance on reproduction in dairy cows. Lucrări Stiinłifice Medicină Veterinară. 2009;42(2):198–205.
Silveira PAS, Butler W, da Silva TC, Barros CC, Corrêa MN, Schneider A. Association of polymorphisms in the IGF-I, GHR and STAT5A genes with serum IGF-I concentration and reproductive performance of Holstein dairy cows. Anim Reprod Sci. 2019;211: 106206.
Brown-Borg HM. Growth hormone, not IGF-I is the key longevity regulator in mammals. The Journals of Gerontology. 2022;77(9):1719–23.
Nugroho T, Maharani D, Widi T. Candidacy of genes associated with economic traits for Madura cattle in Sonok area. In: IOP Conference Series: earth and environmental science: 2022. IOP Publishing. 2022;1001:012018. https://doi.org/10.1088/1755-1315/1001/1/012018.
Behzadi S, MIRAEI AS, Sadeghi M, Zamani P, Abdoli R. Association of IGF-I Gene Polymorphisms with Carcass Traits in Iranian Mehraban Sheep Using Sscp Analysis. Iran J Appl Anim Sci. 2015;5(1):121–6. https://journals.iau.ir/article_513448.html.
Saleh AA, Hammoud MH, Dabour NA, Hafez EEE, Sharaby MA. Genetic variability in GH and IGFBP-3 genes and their association with growth performance in Egyptian sheep. Applied Veterinary Research. 2022;1(3):e2022012–e2022012.
Qian Y, Berryman DE, Basu R, List EO, Okada S, Young JA, Jensen EA, Bell SR, Kulkarni P, Duran-Ortiz S, et al. Mice with gene alterations in the GH and IGF family. Pituitary. 2022;25(1):1–51. https://doi.org/10.1007/s11102-021-01191-y.
Valencia CPL, Franco LÁÁ, Herrera DH. Association of single nucleotide polymorphisms in the CAPN, CAST, LEP, GH, and IGF-1 genes with growth parameters and ultrasound characteristics of the longissimus dorsi muscle in Colombian hair sheep. Trop Anim Health Prod. 2022;54(1):82.
Lechniak D, Adamowicz T, Stanisławski D, Kaczmarek D. In vitro maturation and fertilisation of bovine oocytes in relation to GH gene polymorphism (Leu/Val). Reprod Nutr Dev. 2002;42(3):275–80.
Pereira AP, Alencar MMD, Oliveira HND, Regitano LCDA. Association of GH and IGF-I polymorphisms with growth traits in a synthetic beef cattle breed. Genet Mol Biol. 2005;28:230–6.
Kiyici JM, Akyüz B, Kaliber M, Arslan K, Aksel EG, Cinar MU. Association of GH, STAT5A, MYF5 gene polymorphisms with milk somatic cell count, EC and pH levels of Holstein dairy cattle. Anim Biotechnol. 2022;33(3):401–7.
Misrianti R, Anggraeni A, Andreas E, Sumantri C. Growth hormone gene polymorphism and its association with partial cumulative milk yields of Holstein Friesian dairy cattle. Media Peternakan. 2012;35(3):145–145.
Hartanto S, Budiyanto A, Widayanti R, Setyawan EMN, Prasetya ID. Characterization of polymorphisms in the follicle-stimulating hormone receptor and insulin-like growth factor-1 genes and their association with fertility traits in Jawa-Brebes cows. Veterinary World. 2023;16(4):711.
Lucy M, Verkerk G, Whyte B, Macdonald K, Burton L, Cursons R, Roche J, Holmes C. Somatotropic axis components and nutrient partitioning in genetically diverse dairy cows managed under different feed allowances in a pasture system. J Dairy Sci. 2009;92(2):526–39.
Nicolini P, Chilibroste P, Laborde D, Meikle A. Effect of IGF-I genotype on milk production and the metabolic endocrinology in the transition period in dairy cows in pastoral conditions. Veterinaria (Montevideo). 2013;49(190):16–27.
Lin-Su K, Wajnrajch MP. Growth hormone releasing hormone (GHRH) and the GHRH receptor. Rev Endocr Metab Disord. 2002;3:313–23.
Martin-Estal I, De La Garza R, Castilla-Cortazar I. Intrauterine growth retardation (IUGR) as a novel condition of insulin-like growth factor-I (IGF-I) deficiency. Rev Physiol Biochem Pharmacol. 2016;170:1–35.
Mehmannavaz Y, Amirinia C, Bonyadi M, Torshizi RV. Association of IGF-I gene polymorphism with milk production traits and paternal genetic trends in Iranian Holstein bulls. African Journal of Microbiology Research. 2010;4(1):110–4.
Coffey E, Vinson W, Pearson R. Potential of somatic cell concentration in milk as a sire selection criterion to reduce mastitis in dairy cattle. J Dairy Sci. 1986;69(8):2163–72.
Green M, Green L, Schukken Y, Bradley A, Peeler E, Barkema H, De Haas Y, Collis V, Medley G. Somatic cell count distributions during lactation predict clinical mastitis. J Dairy Sci. 2004;87(5):1256–64.
Zhao X, Lacasse P. Mammary tissue damage during bovine mastitis: causes and control. J Anim Sci. 2008;86(suppl_13):57–65.
Santos MV, Ma Y, Barbano DM. Effect of somatic cell count on proteolysis and lipolysis in pasteurized fluid milk during shelf-life storage. J Dairy Sci. 2003;86(8):2491–503.
Miles AM, McArt JA, Yepes FAL, Stambuk CR, Virkler PD, Huson HJ. Udder and teat conformational risk factors for elevated somatic cell count and clinical mastitis in New York Holsteins. Prev Vet Med. 2019;163:7–13.
Berry D, Lee J, Macdonald K, Stafford K, Matthews L, Roche J. Associations among body condition score, body weight, somatic cell count, and clinical mastitis in seasonally calving dairy cattle. J Dairy Sci. 2007;90(2):637–48.
Mehra R, Garhwal R, Sangwan K, Guiné RP, Lemos ET, Buttar HS, Visen PKS, Kumar N, Bhardwaj A, Kumar H. Insights into the research trends on bovine colostrum: Beneficial health perspectives with special reference to manufacturing of functional foods and feed supplements. Nutrients. 2022;14(3):659.
Saeed AM, Abushady AM. Molecular characterization of some candidate genes in pure Egyptian Buffaloes and crossbred of Italian Buffaloes. Egyptian Journal of Animal Production. 2022;59(1):9–18.
Ge W, Davis M, Hines H, Irvin K, Simmen R. Association of a genetic marker with blood serum insulin-like growth factor-I concentration and growth traits in Angus cattle. J Anim Sci. 2001;79(7):1757–62.
Omer RMA, Masri MARM, Jawasreh KI, Nour IA, Biraima ADA, Musa LM-A, Ahmed M-KA. Molecular Detection of Selected Genetic Polymorphisms in Growth Hormone and Insulin Like Growth Factor I Genes in Indigenous Sudanese Baggara Cattle. Kafkas Üniversitesi Veteriner Fakültesi Dergisi. 2018;24(2).
Putra I, Sari D, Rachmawati S, Oktaviani R, Noor R, Jakaria J. Identification of single nucleotide polymorphism c. 957A> C of PLAG1 gene and its association with growth traits in Bali cattle. J Indones Trop Anim Agric. 2021;46(3):199–208.
Saleh AA, Xue L, Zhao Y. Screening Indels from the whole genome to identify the candidates and their association with economic traits in several goat breeds. Funct Integr Genomics. 2023;23(1):58.
Siadkowska E, Zwierzchowski L, Oprzadek J, Strzalkowska N, Bagnicka E, Krzyzewski J. Effect of polymorphism in IGF-1 gene on production traits in Polish Holstein-Friesian cattle. Anim Sci Pap Rep. 2006;24(3):225–37.
Czerniawska-Piątkowska E, Szatkowska I, Zaborski D, Grzesiak W, Tabor-Osińska S, Wasielewska M, Proskura WS, Kruszyński W, Pawlina E. Single nucleotide polymorphism in the promoter region of the IGF-I gene is associated with milk production in Holstein and Jersey Cattle–is the aspect of present research still relevant in the era of genomic selection? Pak J Zool. 2021;53(6):2295.
Ramesha K, Rao A, Basavaraju M, Geetha G, Kataktalware M, Jeyakumar S. Genetic variability of bovine GHR, IGF-I and IGFBP-3 genes in Indian cattle and buffalo. South African Journal of Animal Science. 2015;45(5):485–93.
Yazdanpanah A, Roshanfekr H, Mirzadeh K, Mamouei M, Khederzadeh S. Polymorphism of insulin-like growth factor 1 gene in Najdi cattle populations. Am J Biochem Biotechnol. 2013;9(3):300–6.
Szewczuk M, Zych S, Czerniawska-Piątkowska E, Wojcik J. Association between IGFIR/i16/TaqI and IGFI/SnaBI polymorphisms and milk production traits in Polish Holstein-Friesian cows. Anim Sci Pap Rep. 2012;43(4):30(1):13–24.
Mirzaei A, Sharifiyazdi H, Ahmadi M, Ararooti T, Ghasrodashti AR, Kadivar A. The effect of polymorphism in gene of insulin-like growth factor-I on the serum periparturient concentration in Holstein dairy cows. Asian Pac J Trop Biomed. 2012;2(10):765–9.
Bonakdar E, Rahmani H, Edriss M, Tabatabaei BS. IGF-I gene polymorphism, but not its blood concentration, is associated with milk fat and protein in Holstein dairy cows. Genet Mol Res. 2010;9(3):1726–34.
Gobikrushanth M, Purfield DC, Colazo M, Wang Z, Butler S, Ambrose D. The relationship between serum insulin-like growth factor-I (IGF-I) concentration and reproductive performance, and genome-wide associations for serum IGF-I in Holstein cows. J Dairy Sci. 2018;101(10):9154–67.
Wathes DC, Becker F, Buggiotti L, Crowe MA, Ferris C, Foldager L, Grelet C, Hostens M, Ingvartsen KL, Marchitelli C. Associations between circulating IGF-I concentrations, disease status and the leukocyte transcriptome in early lactation dairy cows. Ruminants. 2021;1(2):147–77.
Sadeghi M, Mokhber M, Shahrbabak M. Genetic variation in hypothalamic-pituitary axis candidate genes and their effects on milk production traits in Iranian Holstein Cattle. Russ J Genet. 2022;58(11):1393–400.
Curi RA, Palmieri DA, Suguisawa L. Oliveira HNd, Silveira AC, Lopes CR: Growth and carcass traits associated with GH1/Alu I and POU1F1/Hinf I gene polymorphisms in Zebu and crossbred beef cattle. Genet Mol Biol. 2006;29:56–61.
Lucy M, Hauser S, Eppard P, Krivi G, Clark J, Bauman D, Collier R. Variants of somatotropin in cattle: gene frequencies in major dairy breeds and associated milk production. Domest Anim Endocrinol. 1993;10(4):325–33.
Kovacs K, Völgyi-Csík J, Zsolnai A, Györkös I, Fésüs L. Associations between the AluI polymorphism of growth hormone gene and production and reproduction traits in a Hungarian Holstein-Friesian bull dam population. Arch Anim Breed. 2006;49(3):236–49.
Dario C, Carnicella D, Ciotola F, Peretti V, Bufano G. Polymorphism of growth hormone GH1-AluI in Jersey cows and its effect on milk yield and composition. Asian Australas J Anim Sci. 2008;21(1):1–5.
Balogh O, Kovács K, Kulcsár M, Gáspárdy A, Fébel H, Zsolnai A, Fésüs L, Delavaud C, Chilliard Y, Gilbert R. Interrelationship of growth hormone AluI polymorphism and hyperketonemia with plasma hormones and metabolites in the beginning of lactation in dairy cows. Livest Sci. 2009;123(2–3):180–6.
Hadi Z, Atashi H, Dadpasand M, Derakhshandeh A, Seno MG. The relationship between growth hormone polymorphism and growth hormone receptor genes with milk yield and reproductive performance in Holstein dairy cows. Iranian journal of veterinary research. 2015;16(3):244.
Yardibi H, Hosturk GT, Paya I, Kaygisiz F, Ciftioglu G, Mengi A, Oztabak K. Associations of growth hormone gene polymorphisms with milk production traits in South Anatolian and East Anatolian Red cattle. J Anim Vet Adv. 2009;8(5):1040–4.
Shaidullin RR, Zagidullin, L.R., Akhmetov, T. M. and Khalilova, G. K. : Estimation of milk productivity of Kholmogory cows with allelic variants of prolactin and somatotropin genes. Veterinary and animal science, No 3 https://doi.org/10.28983/asjy2022i3pp75-782022.
Amiri S, Jemmali B, Ferchichi MA, Jeljeli H, Boulbaba R, Ben Gara A. Assessment of growth hormone gene polymorphism effects on reproductive traits in Holstein dairy cattle in Tunisia. Archives Animal Breeding. 2018;61(4):481–9.
Lechniak D, Machnik G, Szydlowski M, Switonski M. Growth hormone gene polymorphism and reproductive performance of AI bulls. Theriogenology. 1999;52(7):1145–52.
Mullen M, Berry D, Howard D, Diskin M, Lynch C, Berkowicz E, Magee D, MacHugh D, Waters S. Associations between novel single nucleotide polymorphisms in the Bos taurus growth hormone gene and performance traits in Holstein-Friesian dairy cattle. J Dairy Sci. 2010;93(12):5959–69.
Falchi L, Cesarani A, Mastrangelo S, Senczuk G, Portolano B, Pilla F, Macciotta NP. Analysis of runs of homozygosity of cattle living in different climate zones. Journal of Animal Science. 2023;101:skad061.
Miller BS, Rogol AD, Rosenfeld RG. The history of the insulin-like growth factor system. Hormone Research in Paediatrics. 2022;95(6):619–30.
Yang L, Li C, Song T, Zhan X: Growth hormone proteoformics atlas created to promote predictive, preventive, and personalized approach in overall management of pituitary neuroendocrine tumors. EPMA Journal 2023:1–14.
Zulu VC, Nakao T, Sawamukai Y. Insulin-like growth factor-I as a possible hormonal mediator of nutritional regulation of reproduction in cattle. J Vet Med Sci. 2002;64(8):657–65.
Kim JW. Modulation of the somatotropic axis in periparturient dairy cows. Asian Australas J Anim Sci. 2014;27(1):147.
Sambrook J, Fritsch E. Maniatis. 1989. Molecular cloning: a laboratory manual. Cold Spring Harbor: Cold Spring Harbor Laboratory Press; 1997.
Nei M, Roychoudhury AK. Sampling variances of heterozygosity and genetic distance. Genetics. 1974;76(2):379–90.
Nei M, Li W-H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc Natl Acad Sci. 1979;76(10):5269–73.
Court M, Michael H. Court's (2005–2008) online calculator. 2012.
Acknowledgements
‘‘Not applicable.’’
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). ‘‘Not applicable.’’.
Author information
Authors and Affiliations
Contributions
The work presented here was carried out in collaboration between all authors. A.S, A.R, D.E, A.EL and M.SH. defined the research theme. A.S and E.H. designed methods and experiments. A.S., A.R., N.H., and T.G carried out the field experiments. A.S. carried out the genetic investigation. A.S, A.R and E.H. organized the data, and A.S. and A.R. interpreted the results and wrote the manuscript. All authors read, reviewed and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All animals and sampling procedures in this experiment were supervised and approved by the Institutional Animal Care and Use Committee of the Faculty of Agriculture, Alexandria University, Egypt (AU082211211117). All procedures and experimental protocols were under the Guide for the Care and Use of Agricultural Animals in Research and Teaching, Federation of Animal Science Societies (FASS, 2010) https://www.aaalac.org/about/Ag_Guide_3rd_ed.pdf. Also, the study was carried out in compliance with the ARRIVE guidelines (https://arriveguidelines.org).
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Saleh, A.A., Hassan, T.G.M., EL-Hedainy, D.K.A. et al. IGF-I and GH Genes polymorphism and their association with milk yields, composition and reproductive performance in Holstein–Friesian dairy cattle. BMC Vet Res 20, 341 (2024). https://doi.org/10.1186/s12917-024-04188-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12917-024-04188-4