
Abstract
Edwardsiellosis is a bacterial fish disease that mostly occurs in freshwater farms and is characterized by a high mortality rate. Edwardsiella tarda strain was recovered from 17 fish out of 50 Nile tilapia, which were harboring clinical signs of systemic septicemia. The level of un-ionized ammonia (NH3) in the fish farm’s water was 0.11–0.15 mg/L, which was stressful for the Nile tilapia.
Sequencing of the gyrB1 gene confirmed that the isolate was E. tarda JALO4, and it was submitted to NCBI under the accession number PP449014. The isolated E. tarda harbored the virulence gene edw1 AHL-synthase (quorum sensing). In addition, the isolate was sensitive to trimethoprim and sulfamethoxazole mean while it was intermediate to florfenicol. The median lethal dose (LD50) of E. tarda JALO4 was determined to be 1.7 × 105 CFU/mL in Nile tilapia.
In the indoor experiment, Nile tilapia (45.05 ± 0.4 g), which received dietary Spirulina platensis (5 and 10 g/kg fish feed), showed optimum growth and feed utilization. Meanwhile, after receiving dietary S. platensis, the fish’s feed conversion ratio (FCR) was significantly enhanced compared to the control, which was 1.94, 1.99, and 2.88, respectively. The expression of immune-related genes interleukin (IL)-1β and tumor necrosis factor (TNF)-α were upsurged in E. tarda-challenged fish with higher intensity in S. platensis groups. Dietary S. platensis at a dose of 10 g/kg fish feed could provide a relative protection level (RPL) of 22.2% Nile tilapia challenged against E. tarda. Nile tilapia experimentally infected E. tarda, drastically altering their behavior: higher operculum movement, low food apprehension, and abnormal swimming dietary S. platensis (10 g/kg fish feed) could rapidly restore normal status.
It was concluded that Edwardsiellosis could alter Nile tilapia behavior with a high loss in fish population. Fish received dietary-S. platensis could rapidly restore normal behavior after E. tarda infection. It is recommended the incorporation of S. platensis at doses of 10 g/kg into the Nile tilapia diet to boost their immunity and counteract E. tarda infection.
Similar content being viewed by others
Introduction
Aquatic animals are poikilothermic, and their immune responses are affected by the surrounding environment, such as water parameters salinity, temperature, pH, dissolved oxygen, and ammonia compounds [1, 2]. Nile tilapia (Oreochromis niloticus) belongs to the Cichlidae family that originated from Africa, and it can tolerate a wide range of water temperature16–38 °C; beyond this level, health hazards occurred due to deterioration of several metabolic functions resulting in a decrease of feed intake and immune suppression becoming more vulnerable to opportunistic pathogens, even become lethal below 10 °C [3, 4].
Microbial diseases are a major constraint in aquaculture and cause annual fish losses, job casualties, economic crises, and food insecurity; fish pathogens are mainly opportunistic, affecting immune-depressed fish [5,6,7,8]. The genus Edwardsiella belongs to the Enterobacteriaceae family had five species that are pathogenic to fish: E. tarda, E. ictaluri, E. hoshinae, E. piscicida according to Abayneh et al. [9], and E. anguillarum according to Shao et al. [10], E. tarda is a Gram-negative bacilli that is known as highly pathogenic bacteria in both humans and aquatic animals [11, 12]. Several studies have shown phenotypic variations and interspecific diversity between isolates retrieved from different geographic regions and host species [13]. Therefore, rapid and precise detection of the causative agent is a principal issue for the control and epidemiological studies [14]. It affects many aquatic species, causing mass mortality and considerable economic losses worldwide [15]. Several reports on the occurrence of Edwardsiellosis in non-catfish species included the Rosy Barb (Puntius conchonius) [16], Nile tilapia in the Western Hemisphere [17], and zebrafish (Danio rerio) [18].
Circumstances leading to immune suppression are inevitable in aquaculture (open system), so natural immune promoters could manage Nile tilapia to withstand opportunistic pathogens.
Spirulina platensis is a microscopic, filamentous, multicellular, and photosynthetic blue-green alga commonly used as a safe and functional supplement in animal feed. S. platensis possesses a smooth body with a delicate cell wall, making it highly digestible [19, 20]. S. platensis is a nutritionally dense cyanobacteria containing appreciable levels of amino acids, fatty acids, vitamins, minerals, pigments, and digestive enzymes that are considered beneficial to health and improve fish resistance against pathogens and environmental stressors [21,22,23]. S. platensis has two distinctive antioxidant pigments (carotenoids and phycocyanin) and vitamins (E and C), in addition to chlorophyll, fucoxanthin, polyphenols, and polysaccharides [23, 24]. S. platensis enhanced nonspecific immunity of channel catfish (Ictalurus punctatus) and increased host defense mechanisms [25], Nile tilapia [26]. Meanwhile, Watanuki et al. [27] found that dietary S. platensis could stimulate the immune system of common carp (Cyprirus carpio).
In this work, we investigate the bacterial pathogen that was accompanied by high mortality in freshwater fish farms, the alteration in fish behavior and immune responses associated with bacterial infection, and the possible amelioration role of S. platensis.
Materials and methods
Case history and sampling
Nile tilapia (Oreochromis niloticus) weighing 80–120 gm were cultured in earthen ponds at a density of 30,000 per acre; paddlewheel aerators were not used on the farm. Fish experienced approximately 50 deaths/acre/day during the early summer of 2023 on a private farm in Kafrelsheikh governorate, Egypt. Moribund fish had low feed intake, anorexia, abnormal swimming behavior, lethargy, skin ulcerations, and hemorrhage.
Moribund and dead Nile tilapia from the affected fish farm were transferred to the bacteriology laboratory of fish diseases at the Animal Health Research Institute (AHRI) within 1–2 h using an ice box. The fish were examined for clinical signs and post-mortem lesions of fish diseases [28].
Examination of fish farm water
On fish farm, physical water parameters were determined including water temperature and salinity (model YSI environmental, EC300), dissolved oxygen (DO) (Aqualytic, OX24), and pH (Thermo Orion, model 420 A).
Samples of farm water were collected in septic polyethylene bottles to determine the concentration of ammonia compounds including: total ammonia nitrogen (TAN), unionized ammonia (NH3), nitrite (NO2), and nitrate (NO3), samples were transferred on ice to the laboratory of Animal Health Research Institute (AHRI), and were analyzed using a UV/Visible spectrophotometer, Thermo-Spectronic 300 [29].
Bacteriological examination
Isolation, phenotypic and biochemical identification of Edwardsiella spp
The preliminary identification of bacterial isolates: Swabs were taken from the Nile tilapia organs, kidney, heart, liver, and spleen, then cultivated into tryptic soy broth (TSB) at 30 °C/24 h. Loopfuls from the bacterial broth culture were cultivated onto agar tryptic soy with 5% sheep blood and incubated at 30 °C/24 h. Prevailed colonies were randomly selected and purified by sub-culturing onto TSA. The isolated colonies were sub-cultivated onto selective agar Salmonella-Shigella agar. Bacterial isolates were stored at -80 °C in tryptic soy broth with glycerol for further analysis. The isolate was identified using the Gram-stain and API 20E (bio-Merieux) [30, 31]. All bacterial media are manufactured by Difco, Detroit, USA.
Molecular identification of Edwardsiella spp
DNA extraction and polymerase chain reaction PCR
To identify the bacterial isolate, the bacterial DNA was extracted and examined for the presence of the E. tarda gyrB1 gene. It was also tested for virulence-determinant genes, including Cds1 chondroitinase, edwI AHL-synthase, qseC sensor protein, and pvsA vibrioferrin synthesis. The extraction process of bacterial DNA was performed using the PathoGene-spin™ DNA Extraction Kit (iNtRON Biotechnology, Seongnam, Korea). Briefly, the PCR reaction consisted of a 25 µl mix containing 12.5 µl of 2X MyTaq HS mix (Bioline, Meridian Life Science, UK); 1 µL of each primer, 100 ng of template DNA; and RNase-free water. The PCR products were separated using electrophoresis on agarose gel 1% (Applichem, Germany, GmbH) and were photographed by a gel documentation system (Alpha Innotech, Biometra). The primers and PCR conditions for gene amplification are in Table 1.
Sequencing for strain identification
The extracted DNA was adjusted to 50 ng/µl and then stored at − 20 ˚C for sequencing. The isolate of Edwardsiella strain was identified and discriminated based on DNA sequencing analysis of E. tarda gyrB1. The amplicons were excised from the gel and purified using the QIAquick extraction kit (Qiagen, Germany). Nucleotide sequencing of the amplified genes was performed using the Big Dye terminator v3.1 kit (Life Technologies, Applied Biosystems, Foster City, CA, USA) by ABI 3730xl DNA sequencer (Applied Biosystems™, USA) using a primer of the PCR reaction in both directions. The raw sequence was manually checked and edited for ambiguities using a sequence alignment editor (BioEdit v. 7.2.5) [32]. Finally, the edited sequence was submitted to the GenBank database to get the accession number.
Phylogenetic tree
The Neighbor-Joining phylogenetic tree was conducted to match the gyrB1 of Edwardsiella spp. with the accession numbers of typing strains retrieved from the GenBank database using MEGA X [33]. The phylogenetic tree of sodB gene was rooted on Vibrio anguillarum strain ATCC 43,314 (KU755359.1), which was used as an outgroup. The lineage supports were assessed using nonparametric bootstrapping with 1000 replicates. The following factors were also employed: substitutions: transversions and transitions; pattern among lineages: homogeneous; and 95% cutoff partial deletion principal.
Gene expression of immune-related genes
The impact of dietary S. platensis on the expression of pro-inflammatory genes in the head kidney of the experimental Nile tilapia. The interleukin (IL)-1β and tumor necrosis factor (TNF)-α genes (Table 1) was assessed using quantitative real-time polymerase chain reaction (RT-PCR). The RNA was extracted with Trizol reagent (iNtRON Biotechnology Inc., Korea), and samples were collected from three Nile tilapia, each group using Nanodrop D-1000 spectrophotometer (NanoDrop Technologies Inc., USA). The obtained RNA was assessed for quality and quantity and kept at − 80 °C. The β-actin was used as a housekeeping gene. The results were assessed using Eq. 2−ΔΔCT [34].
Lethal median dose and pathogenicity testing
The LD50 was determined briefly according to the procedure described by Reed and Muench [37], briefly. After acclimatization at the wet laboratory of the Animal Health Research Institute, Nile tilapia (n = 10) with a mean weight of (70.8 ± 0.3 g) were anesthetized with 150 µg/ L of tricaine methanesulfonate (MS222; Sigma, St. Louis, MO, the USA), then fish were injected. Ten fish (duplicate groups) were intraperitoneally injected with 100 µl suspension of serial tenfold dilutions of E. tarda 24 h-old culture (1 × 102, 1 × 103,1 × 104, 1 × 105, 1 × 106, 1 × 107, 1 × 108, 1 × 109, or 1 × 1010 CFU/mL). Mortality rates were recorded for 14 days, and E. tarda was re-isolated from the dead moribund fish and confirmed via PCR.
Antimicrobial sensitivity analyses
The susceptibility of isolated E. tarda strain to antimicrobial agents was determined using the disc diffusion method [38]. The bacteria was cultured onto Mueller–Hinton agar (Oxoid™) for 24 h at 30 °C, then discs were seeded on the agar, discs were containing tetracycline 30 µg, trimethoprim/sulfamethoxazole (SXT) 1.25/23.75 µg), ciprofloxacin (CIP) 5 µg, florfenicol 30 µg, erythromycin (E, 15 µg), gentamicin 10 µg, amoxicillin 10 µg, ampicillin (AMP) 10 µg, kanamycin 30 µg, cefotaxime 30 µg, and streptomycin 30 µg). The results were calculated by measuring the inhibition zones [38]. Also, the multi-drug resistance (MDR) index using the following equation [39],
Where X is the number of antibiotics that the isolate was resistant to and Y is the number of antibiotics used, if the MDR index is higher than 0.2, the bacterial isolate is multiple antibiotic-resistant.
Feeding trial
In an indoor trial to improve the performance of Nile tilapia to combat the recovered pathogens, the green algae Spirulina platensis was added to the fish diet. Nile tilapia was collected from freshwater fish farm with no history of high mortality or health troubles. Fish weighing 45.05 ± 0.4 g was tranquilized in the fish farm and then swiftly transferred in labeled plastic bags filled with 1/3 clean water and 2/3 pure oxygen. At the farm site, fish were tranquilized using anesthetic agent MS-222 (tricaine methanesulfonate) at a dose of 40 mg/L of transporting water; MS-222 is produced by Syndel, Canada. Fish were subjected to an iodine bath at 20 ppm/10 minutes (Betadine®, the active ingredient, is 5% of povidone-iodine and produced by the Nile Company for Pharmaceuticals) at the arrival in the wet laboratory at Animal Health Research Institute, Agriculture Research Center, Kafrelsheikh Egypt [40, 41].
All aquaria were well aerated via an air bubble stone diffuser connected with a plastic hose and an air blower. One-third of water was replaced with clean, fresh water day after day to maintain constant water parameters of the aquaria; temperature (27 ± 1.2 °C), salinity (0.2 ± 0.05 g/L), pH (7.2 ± 0.2), and DO (5.2 ± 0.3 mg/L), TAN (0.09 ± 0.02 mg/L), NH3 (0.02–0.01 mg/L), NO2 (0.00 mg/L), and NO3 (0.5 ± 0.01 mg/L).
With meticulous precision, the fish food (in pellet form) was soaked and fully homogenated until a paste was formed. The gelatin (Canal AquaCure, Egypt) was then added to the feed paste/S. platensis mixture, enhancing the feed’s consistency. This mixture was left to dry at room temperature and subsequently cut into small, uniform sizes.
The feeding trial lasted for four weeks prior to the experimental infection. Fish feed was offered at 0.9.00 a.m. and 03.30 p.m. at a rate of 5% b.w. fish per day, with a basal diet composition of crude protein of 30.95% and digestible energy of 3400.5 kcal/kg. The diet was purchased from the local market and manufactured by Aller Aqua® Egypt (Reg. no. 9782). Lot. no. 365,622 https://www.aller-aqua.com/.
The variation in the feeding rate of the fish’s body weight was adjusted weekly. The growth performance and feed utilization were determined using the following equations,
Where final body weight (FW), initial body weight (IW), daily weight gain (DWG) feed conversion ratio (FCR), and t = time (days).
Experimental infection
To examine the pathogenicity of isolated E. tarda, 15 fish experimental fed S. platensis and control (control positive) were injected (I/P) with 0.1 mL of a bacterial solution containing LD50 1.7 × 105 CFU/mL, the bacteria dose was adjusted to LD50 using the McFarland scale in phosphate buffer saline. At the same time, a negative control group was schooled by injecting 15 fish with pure saline 0.65 mg/l in concentration (control negative) [42]. Each dead fish was counted if E. tarda was re-isolated for 14 days. the mortality rate (MR) and relative protection level RPL [43] were estimated using the following equations:
Bacteria were re-isolated following the scheme in the above section. The phenotypic profiles of E. tarda were done according to Bergey’s recommendations [44], and biochemical characters were illustrated using API 20E strips [45].
Fish reflexes
Fish breath was assessed by operculum movement (OM), which was recorded by visually counting the number of opercular beats for 1 min at one-h intervals [46]. Scores classify feeding, capture indicators, and respected characteristics for on-farm tilapia welfare assessment [47]. Food apprehension was scored into four levels: 1: in 3: in ≤ 120 s, 2: in 120–180 s, 180–300 s, and 4: No or ≥ 360 s. Escape reflex was scored into four levels 1–4, 1: Normal swimming, 2: Excited swimming behavior, > 20 dorsal fins or low body parts on surface, 3: Swimming in different directions or low activity, and 4: Many fish floating on side and exhausted.
Histopathological examination
At the end of the feeding trial and bacterial challenge, the gills, liver, and spleen specimens were taken and preserved in buffered formalin (10%) for histopathological analyses. Specimens were dehydrated with alcohol and cleared with xylol. They were embedded in paraffin wax and cut into 4 μm sections. Specimens were stained with hematoxylin-eosin and examined using a light microscope [48].
Statistical analyses
Data concerning the ameliorating role of S. platensis to Edwardsiellosis in the experimental Nile tilapia were analyzed to determine the variance (ANOVA). Significance between the experimental groups using one-way ANOVA test and data were expressed as mean ± standard error (SE) at the significant difference of P ≤ 0.05 (the SPSS software for Windows, SPSS Inc., Chicago, IL, USA).
Results
Water parameters and natural infection
On the affected fish farm, water parameters were suitable for tilapia culture, including temperature (25.8–27.5 °C), salinity (1.23–1.41 g/L), pH (8.4–8.9), and DO (3.5–5.3 mg/L). Slight increase in ammonia compounds TAN (0.98–1.29 mg/L), NH3 (0.11–0.15 mg/L), NO2 (0.04–0.06 mg/L), and NO3 (0.88–0.93 mg/L).
It was recorded that out of 50 Nile tilapia harboring clinical signs, 17 fish were infected with E. tarda. Clinical signs were low feed intake, external hemorrhages on the body surface and fin bases, and dentated tail fin (tail rot), whereas postmortem signs were enlarged brownish-red liver, distended gall bladder, splenomegaly, and inflamed intestine with ascetic fluids (Fig. 1).
Bacteria identification and median lethal dose LD50
By using API 20E, 17 out of 50 Nile tilapia were positive for E. tarda, which was identified using API20 E with the identification number (4,144,000), a unique identifier that aids in the classification and tracking of this bacterium.
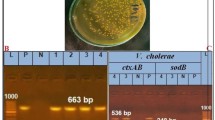
In Fig. 2, PCR analyses, the isolated E. tarda was examined for the presence gyrB1, virulence genes Cds1, edw1, qseC, and pvsA, E. tarda was positive for gyrB1 and edw1 AHL-synthase. The amplified gyrB1 gene was successfully performed from the Edwardsiella isolate using the gene-specific primers pair. The purified PCR products were then sequenced to confirm the identities of bacterial spp. at the species level (Fig. 3), and the isolated strain was identified as E. tarda JALO4 under the accession numbers PP449014 were issued by the GenBank staff. The BLAST analysis of gyrB1 gene sequences proved that bacterial strains belonged to the genus Edwardsiella, and was identified as E. tarda. The comparative sequence analysis of the current gyrB1 sequences (PP449014) revealed a high similarity index between 100% and 99.59% with relevant sequences of E. tarda (MW911830.1 and GQ387361.1). The alignment of these accession numbers (ON843829.1, MN967022.1, KF894804.1, JN700743.1, JN700742.1, JN700740.1, JN700739.1, JN700738.1, OQ361774.1, OQ557504.1, OQ557503.1, MG026726.1, MG225496.1, and MG225495.1) showed low similarity than 98% gyrB1 of E. tarda. In this study, the typing strain of Vibrio anguillarum ATCC 43,314 (KU755359.1) served as an outgroup.
To determine LD50, Nile tilapia weighting 50 g were subjected to descending ten-fold dilutions of the isolated E. tarda JALO4. The LD50 was found to be 1.7 × 105 CFU/mL at water temperature 27.5 ± 2.5 °C, salinity 0.23 g/L, pH 7.9, and DO 5.8 ± 1.5 mg/L.
In Table 2, the isolated E. tarda was sensitive to trimethoprim 1.25 µg & sulfamethoxazole 23.75 µg and intermediate to florfenicol 30 µg, meanwhile it was resistant to tetracycline, ciprofloxacin, erythromycin, gentamycin, amoxicillin, ampicillin, kanamycin, cefotaxime, and streptomycin. These findings indicate a significant challenge in treating E. tarda infections due to its resistance to a wide range of commonly used antibiotics. The obtained MDR was 0.8, indicating that E. tarda JALO4 is a multiple antibiotic-resistant bacteria.

Nile tilapia naturally infected with E. tarda. *(star): A; Tail rot, B; hemorrhage at base of operculum fin, C; hemorrhage at base of anal fin, D; hemorrhage at dorsum, E; hemorrhage at base of dorsal fin, F; hemorrhage at the abdomen. ** (2 stars): A; brownish red liver, B; enlarged gall bladder, C; slightly splenomegaly, D; slight inflamed intestine, E; remnant of abdominal fluid

Edwardsiella tarda gel electrophoresis gyrB1 and virulence genes of E. tarda JALO4. S; sample, N; negative, L; DNA ladder

Phylogenetic tree of gyrB1 gene E. tarda JALO4 (PP449014)
Indoor experimental feeding trial
Growth performance
In Table 3, Nile tilapia (45.05 ± 0.4 g) were supplemented with dietary S. platensis at a dose of 5 and 10 g/kg diet. The growth parameters FW (56.87 and 57.46 g), WG (11.67 and 12.23 g), DWG (0.42 and 0.44 g), and FI (12.2 and 23.66 g) were significantly increased in fish that received dietary S. platensis compared to the control 51.2 g, 6.53 g, 0.23 g, and 18.73 g, respectively. A significant enhancement of feed utilization was achieved in S. platensis groups as FCR was 1.94 and 1.99 compared to the control 2.88.
The expression of immune-related genes
Gene expression of pro-inflammatory IL-1β (Fig. 4) and TNF-α (Fig. 5) were affected by both dietary S. platensis and E. tarda infection. Post-infection by 3 h, IL-1β expression was significantly upsurged with E. tarda infection 2.78 and 4.42 fold-change in the head kidney of Nile tilapia received S. platensis at a dose of 5 g/kg fish feed and 10 g/kg fish feed, respectively, compared to infected control positive (+) and uninfected control negative (-) 1.12 and 0.3 fold-change, respectively. Following the trend of IL-1β expression, TNF-α expression was upsurged in response to E. tarda infection in fish that received S. platensis at doses of 5 g and 10 g/kg fish feed compared to infected control.
After 48 h post-infection, the gene expression of IL-1β and TNF-α expression significantly decreased compared with 3-h post-infection.

Gene expression of IL-1β in the head kidney of Nile tilapia experimental infected with E. tarda. Different letters indicate significant difference at P ≤ 0.05. (-); negative control fish were injected with saline, (+); positive control fish were injected with E. tarda

Gene expression of TNF-α in the head kidney of Nile tilapia experimental infected with E. tarda. Different letters indicate significant difference at P ≤ 0.05. (-); negative control fish were injected with saline, (+); positive control fish were injected with E. tarda
Experimental infection and relative protection level of S. platensis
In Fig. 6, clinical signs of the experimentally infected Nile tilapia were similar to those obtained with naturally infected ones: external body discoloration (paleness), scale loss, and slight exophthalmia. Also, post-mortem were brownish hemorrhagic liver, distended gall bladder, splenomegaly, and a partially empty intestine with greenish content.
In Table 4, MR% was decreased in the fish group that received S. platensis at a dose of 10 g/kg fish feed compared with the control and those that received 5 g /kg fish feed: 46.67%, 60%, and 60%, respectively. Supplementation of Nile tilapia with S. platensis at a dose of 10 g/kg fish feed could provide a RPL of 22.2%.

Nile tilapia experimentally infected with E. tarda. *(star): multiples hemorrhages on external body surface. ** (2 stars): A; body discoloration paleness, B; hemrrahges at base of operculum fin, C; pop-eye. *** (3 stars): A; brownish liver with hemorrhage on surface, B; enlarged gall bladder, C; splenomegaly, D; intestine with greenish content
Fish behavioral reflexes at 3 and 14 days post-infection
In Table 5, Nile tilapia experimentally infected with E. tarda showed drastic behavior changes; OM (opercular beats for 1 min period at 1 h interval) was increased regardless of S. platensis supplementations. Meanwhile, OM was higher in fish that received dietary S. platensis 55–61 opercular beats in the 3 days post-infection, restoring average values in 14 days compared to the control. Food apprehension was decreased in response to E. tarda infection, and S. platensis supplementation could enhance and restore average values after 14 days post-infection. Nile tilapia experimentally infected with E. tarda lost their normal swimming behavior, and dietary S. platensis could reserve normal behavior in the 3 days post-infection; 14 days post infections survived, and fish could normally swim.
Histopathological analyses pre- and post-infection
In Fig. 7, histological features of the gills of control fish (a), fish received 5 g (b) and 10 g (c) /kg fish feed S. platensis were normal. In contrast, fish of the control and infected with E. tarda (d) showed truncated lamellae and tips sloughing. Fish (e) received S. platensis and infected with E. tarda had lamellar aneurysm. In Fig. 8, fish in the control and fish feed S. platensis groups (a, b, and c) showed normal hepatic-parenchyma and development of pancreatic acini with the occurrence of some melano-macrophage cells. The hepatic tissue of control fish infected with E. tarda (d) showed liquifactive necrosis in the form of a wide area of lysed hepatic-parenchyma, fish received S. platensis and infected with E. tarda (e) hepatic had a small lysed area in the hepatic-parenchyma. In Fig. 9, control fish infected with E. tarda (d) showed marked depletion of lymphocytic elements without melano-macrophage centers, whereas some alterations were recorded in control (a), which were moderately sized melano-macrophage centers. In contrast, fish fed S. platensis (b) and (c) showed wide sized foci of melano-macrophage, dispersed and melano-macrophage cells. A moderate sized melano-macrophage centers in fish fed S. platensis and infected with E. tarda (e). In Fig. 10, Muscle bundles; normal histological criteria of muscle bundles in control fish (a) and fish received S. platensis (b). Muscle bundles of control fish infected with E. tarda showed foci of bacteria aggregation, vacuolation of (white arrows, c), Evidence of muscle regeneration in the form of increased cellularity, split bundles of fish received 5 g/kg fish feed S. platensis and infected with E. tarda (d) and corrugation in fish received 10 g/kg fish feed S. platensis and infected with E. tarda (e).

Normal histological criteria of gills in (a, b& c), with truncated lamellae, some showing tips sloughing (d), lamellar atelectasis (lamellar aneurysm) in (e). Note: (a); control fish, (b); fish received 5 g/kg fish feed S. platensis, (c); fish received 10 g/kg fish feed S. platensis, (d); control infect with E. tarda, (e); fish received S. platensis and infect with E. tarda. H&E X 400

Liver with evidence of liquefactive necrosis in the form of a wide area of lysed hepatic- parenchyma (d), normal hepatic parenchyma, development of pancreatic acini (a, b &c) with occurrence of some melano-macrophage cells (a&b). Small lysed area still detected in hepatic parenchyma (e) Note (a); control fish, (b); fish received 5 g/kg fish feed S. platensis, (c); fish received 10 g/kg fish feed S. platensis, (d); control infect with E. tarda, (e); fish received S. platensis and infect with E. tarda. H&E X 400

Spleen parenchyma revealing moderate melano-macrophage centers (a & e), wide-sized foci of melano-macrophage (b), dispersed melano-macrophage cells (c), and marked depletion of lymphocytic elements with absence of melano-macrophage centers (d). Note (a); control fish, (b); fish received 5 g/kg fish feed S. platensis, (c); fish received 10 g/kg fish feed S. platensis, (d); control infect with E. tarda, (e); fish received S. platensis and infect with E. tarda. H&E X 400

Muscle bundles; normal histological criteria of muscle bundles in (a & b). showing foci of bacteria aggregation in form of blue foci, vacoulation of muscle bundles (white arrows, c), Evidence of muscle regeneration in form of; increase cellularity, splitted bundles (d) and corrugation (e). Note (a); control fish, (b); fish received S. platensis, (c); fish control and infect with E. tarda, (d); fish received 5 g/kg fish feed S. platensis and infect with E. tarda (e); fish received 10 g/kg fish feed S. platensis and infect with E. tarda. H&E X 400
Discussion
In this work, the concentration of ammonia compounds of the affected fish farm was high enough to be stressful and immune depressed for Nile tilapia, including TAN (0.98–1.29 mg/L), NH3 (0.11–0.15 mg/L), NO2 (0.04–0.06 mg/L), and NO3 (0.88–0.93 mg/L) despite partial replace of the pond water. In accordance, Nile tilapia that were exposed to 0.53 and 0.265 mg/L of NH3 and infected with Clostridium perfringens type A suffered from high MR 80% and 70%, respectively [49], these results could be due to impairment of the immune responses that resulted in an upsurge of the vulnerability of fish to bacterial infection [50]. Likely, the safe maximum concentration of NH3 is 0.1 mg/L for short-term in freshwater fish [51] and 0.012 mg/L seawater for long-term in Atlantic salmon posts molts (Salmo salar L.) [52].
Nile tilapia naturally and experimentally infected with E. tarda showed similar clinical signs and post-mortem lesions that were signs of septicemic bacterial diseases, including external hemorrhages, tail rot, ascites, splenomegaly, and inflamed intestines. Similarly, both freshwater fish O. niloticus and Clarias gariepinus naturally infected with E. tarda had skin ulcers and hemorrhages, ascites, muscular hemorrhage, congested liver with splenomegaly; these signs were similar to those observed with the experimentally infected fish [53], in addition, Rao et al. [54] and Leung et al. [55] claimed that E. tarda produced bacterial toxins (various virulence factors) such as hemolysin, catalase and extracellular products (ECPs) that could attack the host cells causing tissue inflammation and damage resulting in immunodepression status in fish.
In the last decades, bacterial identification has relied on genetic-based investigations to avoid the shortcomings of conventional methods [56, 57]. In this work, naturally diseased Nile tilapia were positive for E. tarda, PCR, and sequence of gyrB1. Accordingly, Lan et al. [56] found that the gyrB1 gene could be used as a marker to screen the presence, identification, and classification of Edwardseilla species, while the edw1 gene was used to confirm the virulence and pathogenicity of E. tarda [58]. Similarly, in Egypt, isolated E. tarda harbored edwI and qseC genes [53], and pvsA gene and edw1 gene [59]. These genes (edwI, and qseC genes) are responsible for the virulence of E. tarda by regulating flagellar motility, biofilm formation, and quorum sensing [60].
In the indoor bacterial challenge test, we determined the LD50 of E. tarda to be 1.7 × 105 CFU/mL for Nile tilapia (50 g) under specific water conditions. It is important to note that LD50 can vary significantly among different fish species. For instance, different LD50 values were detected for freshwater fish [61], African catfish and Nile Tilapia [62]. These variations can be attributed to the E. tarda strain, the fish’s weight and size, and accommodation conditions. This information is crucial for understanding the impact of E. tarda on different fish species.
We found that E. tarda was sensitive to Trimethoprim & Sulfamethoxazole, showed intermediate sensitivity to florfenicol, and was resistant to several other antibiotics tetracycline, ciprofloxacin, erythromycin, gentamycin, amoxicillin, ampicillin kanamycin, cefotaxime, and streptomycin. However, Kumar et al. [63] reported that E. tarda strains were multiply resistant to antibiotics, with a high percentage of isolates retrieved from aquatic animals in West Bengal and Bihar, India. These findings underscore the urgent need for further research into alternative treatment strategies, highlighting the importance of our collective efforts in this field.
In this experiment, the growth performance was enhanced in Nile tilapia (45.05 ± 0.4 g), fed S. platensis at doses of 5 and 10 g/kg fish feed. Similar findings with different fish species were recorded by Adel et al. [64], who stated that growth performance and feed utilization of great sturgeon (Huso huso) were improved with S. platensis supplementation at a dose of 100 g/kg fish feed. Also, Mabrouk et al. [65] had similar findings with Nile tilapia fed on dietary S. platensis at 10 /kg fish feed. Some authors fed fish on S. platensis meal and had similar outcomes. Also, Rahman et al. [66] found that Stinging catfish (Heteropneustes fossilis) fed a diet containing S. platensis meal (50 g/kg fish feed) obtained significantly improved WG and DWG than the control ones. Furthermore, Nile tilapia fed dietary S. platensis could utilize feed ingredients more efficiently, so it could be considered an essential element of Nile tilapia natural diet [67].
The importance of natural barriers, including inflammatory cytokine, in the fight against bacterial infections is well known. The pro-inflammatory cytokines (IL-1β and TNF-α) are the supporting columns of humoral cell-mediated immune responses, and they activate macrophages, production of (nitric oxide reactive oxygen species, and reactive nitrogen intermediates), and autophagy [68]. In this work, gene expression of IL-1β and TNF-α were upsurged in response to E. tarda infection by 3 h, with more intensity in fish that received dietary S. platensis. Furthermore, the expression of IL-1β and TNF-α was subsided with lower intensity after 48 h post-infection. This finding was consistent with Mayer-Barber and Yan [69], who claimed that IL-1β is a rapid cytokine that responds to acute bacterial infections; it also stimulates the secretion of chemokines that are required for the optimal control neutrophil-dependent. Likely, the immunity of Nile tilapia was significantly enhanced by feeding S. platensis-diets at 1% more than 2% with a rapid down-regulation of anti-inflammatory cytokine (IL-10) [70]. Also, Awad et al. [71] found that the serum of Nile tilapia received dietary S. platensis had a low content of TNF-α and IL-10, whereas after A. hydrophila infection, a significant rise in TNF-α and a significant downregulation in IL-10 in control. This finding was inconsistent with our study as the infected fish (control) was immunosuppressed, and the inflammatory status persisted. Controversially, Song et al. [72] claimed that grass carp (Ctenopharyngodon idella) infected with A. hydrophila, intestinal pro-inflammatory cytokines (IL-1, IL-8, and TNF-α) were upregulated, and dietary S. platensis significantly reduced TNF-α and elevates IL-10. This difference was due to sampling time, as our samples were collected directly after 3 and 48 h post-infection.
In this experiment, MR% of Nile tilapia challenged with E. tarda was decreased, especially those who received 10 g S. platensis /kg fish feed compared with those of the control and ones who received 5 g S. platensis /kg fish feed, 46.67%, 60%, 60%, respectively. A 10 g S. platensis /kg fish feed dose could provide a 22.2% RPL. Consistent with these findings, dietary S. platensis had a potential immune role in different fish species, enhancing survivability and boosting the resistance against fish pathogens [64]. Similarly, African catfish and Nile tilapia injected with LD50 (0.2 ml of 104 CFU/ml) at 25 °C resulted in MR of 70% and 60% [62]. Consistently, dietary S. platensis increased the resistance against A. hydrophila infection in African Catfish [73], Cyprinus Carpio [26], and Gibel carp (Carassius gibelio) [74]. These findings could be due to the high immune response in Nile tilapia received S. platensis compared to the control. Similarly, common carp (Cyprinus carpio) fed dietary S. platensis showed high cytokines generation with rapid onset [26]. Moreover, a small dose of S. platensis (10 mg/fish /day) could improve the general health of Nile tilapia. Also, it could stimulate the immune system, making fish more resistant to infectious diseases [75]. Also, Tawfeek et al. [76] claimed that dietary Chlorella vulgaris algae could mitigate chlorpyrifos toxicity in Nile tilapia, protecting against Streptococcus agalactiae infection.
Nile tilapia experimentally infected E. tarda in this work, drastically altering their behavior: high OM, low food apprehension, and abnormal swimming. Also, it was noticed that dietary S. platensis could ameliorate these withdrawals with a fast restoration of normal status. Inconsistency acute and chronic stress in aquatic animals is usually accompanied by changes in feeding behavior, respiratory activity, escaping behavior, swimming style, and abnormal behavior [77]. Similarly, Nile tilapia experimentally infected E. tarda with lethargy and showed abnormal swimming behavior and loss of escape reflexes [61]. These changes in fish behavior could be due to insufficient oxygen supply to tissues, increasing compensatory respiration in response to stress and anemia [78, 79].
Histopathological findings revealed gills truncated lamellae and tips sloughing lamellar aneurysm in control fish infected with E. tarda. Hepatic tissue of control and S. platensis groups showed normal. In contrast, hepatic parenchyma necrosis in the form of a wide area of lysed hepatic- parenchyma, fish received S. platensis and infected with E. tarda. The spleen of fish infected with E. tarda showed marked depletion of lymphocytic elements with the absence of melano-macrophage centers. Muscles showed foci of bacteria aggregation and vacuolation. Similarly, E. ictaluri infected farmed hybrid red tilapia (Oreochromis sp.) in Southeast Asia, muscles had various clumps of rod-shaped bacteria stained with hematoxylin were also visualized; the presence of cell pyknosis and karyorrhexis was commonly found in the infected spleen with increasing numbers of melanoma-macrophage centers (MMCs). Also, they added that severe blood congestion, hemorrhage, and necrotic hepatocytes were observed in the hepatic tissue [80]. In accordance, Pirarat et al. [81] stated that lymphoid depletion and lymphoid necrosis were noticeable in control Nile tilapia that was infected with E. tarda, while focal secondary lamellar fusions and severe anemia were observed only in the gills; also they added that L. rhamnosus (probiiotic) could mitigate impacts of experimental E. tarda infection. These pathological lesions resulted from toxins produced by E. tarda while antioxidant contents of S. platensis could ameliorate such damages. Accordingly, Pseudomonas fluorescens caused oxidative stress in Nile tilapia, causing severe tissue damage, and fish fed on S. platensis (1%) showed hyperplasia of melanoma-macrophage centers and hemopoietic elements S. platensis (2%) also showed hyperplasia of melanoma-macrophage center and more activation of hemopoietic tissue [70].
Conclusion
Seventeen Nile tilapia out of 50 fish were infected with E. tarda, which harbored clinical and post-mortem signs resembling experimental fish. The level of NH3 was 0.11–0.15 mg/Lin fish farm, which was stressful for Nile tilapia. The bacterial strain was identified using API20 E (4,144,000) and molecular methods E. tarda strain and deposited in GenBank (PP449014). E. tarda (JALO4) was positive for gyrB1 and edw1 AHL-synthase, while LD50 was determined to be 1.7 × 105 CFU/mL. E. tarda was sensitive to trimethoprim & sulfamethoxazole, with an MDR of 0.8, indicating a multiple antibiotic-resistant bacteria. Higher operculum movement, low food apprehension, and abnormal swimming were observed in experimentally infected fish. The expression IL-1β and TNF-α were intensively upregulated in E. tarda-challenged fish in S. platensis groups. Dietary S. platensis at a dose of 10 g/kg fish feed could provide a higher RPL of 22.2% Nile tilapia challenged against E. tarda.
Data availability
Data is available on request from the corresponding author.
References
Velmurugan BK, Chan CR, Weng CF. Innate-immune responses of tilapia (Oreochromis mossambicus) exposure to acute cold stress. J Cell Physiol. 2019;234(9):16125–35. https://doi.org/10.1002/jcp.28270.
Sood N, Verma DK, Paria A, Yadav SC, Yadav MK, Bedekar MK, Pradhan PK. Transcriptome analysis of liver elucidates key immune-related pathways in Nile tilapia Oreochromis Niloticus following infection with tilapia lake virus. Fish Shellfish Immunol. 2021;111:208–19. https://doi.org/10.1016/j.fsi.2021.02.005.
Kumar N, Sharma SK, Sharma BK, Ojha ML, Jain M. Determination of water quality parameters with reference to different stocking densities and spinach (Spinacia oleracea) plant, grown in deep water aquaponics system. J Pharm Innov. 2023;12(4):512–8.
Sherif AH, Farag EA, Mahmoud AE. (2023)a Temperature fluctuation alters immuno-antioxidant response and enhances the susceptibility of Oreochromis niloticus to Aeromonas hydrophila challenge. Aquac Int 1–14. https://doi.org/10.1007/s10499-023-01263-9.
Assefa A, Abunna F. Maintenance of fish health in aquaculture: review of epidemiological approaches for prevention and control of infectious disease of fish. Vet Med Int. 2018;1–10. https://doi.org/10.1155/2018/5432497.
Sherif AH, Abdellatif JI, Elsiefy MM, Gouda MY, Mahmoud AE. A occurrence of infectious Streptococcus agalactiae in the farmed Nile tilapia. Egypt J Aquat Biol Fish. 2022;26(3):403–32. https://doi.org/10.21608/ejabf.2022.243162.
Sherif AH, AbuLeila RH. Prevalence of some pathogenic bacteria in caged- Nile Tilapia (Oreochromis Niloticus) and their possible treatment. Jordan J Biol Sci. 2022;15(2):239–47. https://doi.org/10.54319/jjbs/150211.
Tawfeek WS, Kassab AS, Okasha LA, Abdelsalam M, Sherif AH. The phenotypic and genetic characteristics of Pseudomonas anguilliseptica strains associated with mortalities in farmed sea bream and sea bass. Aquac Int. 2023;1–20. https://doi.org/10.1007/s10499-023-01360-9.
Abayneh T, Colquhoun DJ, Sørum H. Edwardsiella piscicida sp. nov., a novel species pathogenic to fish. J Appl Microbiol. 2013;114(3):644–54.
Shao S, Lai Q, Liu Q, Wu H, Xiao J, Shao Z, … Zhang Y (2015) Phylogenomics characterization of a highly virulent Edwardsiella strain ET080813T encoding two distinct T3SS and three T6SS gene clusters: propose a novel species asEdwardsiella anguillarum sp. nov. Syst Appl Microbiol 38(1): 36–47.
Xie HX, Lu JF, Zhou Y, Yi J, Yu XJ, Leung KY, Nie P. Identification and functional characterization of the novel Edwardsiella tarda effector EseJ. Infect Immun. 2015;83(4):1650–60.
Yang W, Wang L, Zhang L, Qu J, Wang Q, Zhang Y. An invasive and low virulent Edwardsiella tarda esrB mutant promising as live attenuated vaccine in aquaculture. Appl Microbiol Biotechnol. 2015;99:1765–77.
Austin B, Austin DA. (2012) Miscellaneous Pathogens. Bacterial Fish Pathogens: Disease of Farmed and Wild Fish, Springer Netherlands, pp. 413–441. https://doi.org/10.1007/978-94-007-4884-2_ 12.
Tinsley JW, Lyndon AR, Austin B. Antigenic and cross-protection studies of biotype 1 and biotype 2 isolates of Yersinia ruckeri in rainbow trout, Oncorhynchus mykiss (Walbaum). J Appl Microbiol. 2011;111(1):8–16. https://doi.org/10.1111/j.1365-2672.2011.05020.x.
Enany M, Al-Gammal A, Hanora A, Shagar G, El Shaffy N. Sidr Honey inhibitory effect on virulence genes of MRSA strains from animal and human origin. Suez Canal Veterinary Med J SCVMJ. 2015;20(2):23–30. https://doi.org/10.21608/scvmj.2015.64562.
Humphrey JD, Lancaster C, Gudkovs N, McDonald W. Exotic bacterial pathogens Edwardsiella tarda and Edwardsiella ictaluri from imported ornamental fish Betta splendens and Puntius conchonius, respectively: isolation and quarantine significance. Aust Vet J. 1986;63(11):369–71.
Soto E, Griffin M, Arauz M, Riofrio A, Martinez A, Cabrejos ME. Edwardsiella ictaluri as the causative agent of mortality in cultured Nile tilapia. J Aquat Anim Health. 2012;24(2):81–90.
Hawke JP, Kent M, Rogge M, Baumgartner W, Wiles J, Shelley J, … Peterson TS (2013) Edwardsiellosis caused by Edwardsiella ictaluri in laboratory populations of zebrafish Danio rerio. J Aquat Anim Health 25(3): 171–183.
Mathur M. (2018) Bioactive Molecules of Spirulina: A Food Supplement. In: M´erillon, J.- M., Ramawat, K.G, editors, Bioactive Molecules in Food. Springer International Publishing, pp. 1–22. https://doi.org/10.1007/978-3-319-54528-8_97-1.
Akter T, Hossain A, Rabiul Islam M, Hossain MA, Das M, Rahman MM, … Abdel-Tawwab M(2023) Effects of spirulina (Arthrospira platensis) as a fishmeal replacer in practical diets on growth performance, proximate composition, and amino acids profile of pabda catfish (Ompok pabda). Journal of Applied Aquaculture 35(1): 69–82. https://doi.org/10.1080/10454438.2021.1936740.
Wan D, Wu Q, Kuˇca K. (2021) Chap. 57 - Spirulina. In: Gupta, R.C., Lall, R., Srivastava, A, editors, Nutraceuticals (Second Edition). Academic Press, pp. 959–974. https://doi.org/10.1016/B978-0-12-821038-3.00057-4.
Bortolini DG, Maciel GM, Fernandes IDAA, Pedro AC, Rubio FTV, Branco IG, Haminiuk CWI. Functional properties of bioactive compounds from Spirulina spp.: current status and future trends. Food Chemistry: Mol Sci. 2022;5:100134. https://doi.org/10.1016/j.fochms.2022.100134.
Gogna S, Kaur J, Sharma K, Prasad R, Singh J, Bhadariya V, Jarial S. Spirulina-an edible cyanobacterium with potential therapeutic health benefits and toxicological consequences. J Am Nutr Assoc. 2023;42(6):559–72. https://doi.org/10.1080/27697061.2022.2103852.
Kiadaliri M, Firouzbakhsh F, Deldar H. Effects of feeding with red algae (Laurencia Caspica) hydroalcoholic extract on antioxidant defense, immune responses, and immune gene expression of kidney in rainbow trout (Oncorhynchus mykiss) infected with Aeromonas hydrophila. Aquaculture. 2020;526:735361.
Sattanathan G, Palanisamy T, Padmapriya S, Arumugam VA, Park S, Kim IH, Balasubramanian B. Influences of dietary inclusion of algae Chaetomorpha aerea enhanced growth performance, immunity, haematological response and disease resistance of Labeo rohita challenged with Aeromonas hydrophila. Aquac Rep. 2020;17:100353. https://doi.org/10.1016/j.aqrep.2020.100353.
Sherif AH, Al-Sokary ET, Rizk WF, Mahfouz ME. Immune status of Oreochromis niloticus subjected to long-term lead nitrate exposure and a Arthrospira platensis treatment trial. Environ Toxicol Pharmacol. 2020;76:103352. https://doi.org/10.1016/j.etap.2020.103352.
Watanuki H, Ota K, Tassakka ACMA, Kato T, Sakai M. Immunostimulant effects of dietary Spirulina platensis on carp. Cyprinus carpio Aquaculture. 2006;258(1–4):157–63.
Schaperclaus. Fish diseases, 1. CRC; 1992.
Rice EW, Bridgewater L. (2012) A.P.H. Association, standard methods for the examination of water and wastewater, American public health association, Washington, DC.
Buján N, Mohammed H, Balboa S, Romalde JL, Toranzo AE, Arias CR, Magariños B. Genetic studies to re-affiliate Edwardsiella tarda fish isolates to Edwardsiella piscicida and Edwardsiella anguillarum species. Syst Appl Microbiol. 2018;41(1):30–7. https://doi.org/10.1016/j.syapm.2017.09.004.
Buller NB. Bacteria from Fish and other aquatic animals: a practical identification Manual. Cambridge: CABI Publishing; 2004.
Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/N.T. Nuclic acids. Symp Ser. 1999;41:95–8.
Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 2018;35(6):1547–9.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2 –∆∆CT method. Methods. 2001;25(4):402–8.
Park SB, Kwon K, Cha IS, Jang HB, Nho SW, Fagutao FF, … Jung TS (2014) Development of a multiplex PCR assay to detect Edwardsiella tarda, Streptococcus parauberis, and Streptococcus iniae in olive flounder (Paralichthys olivaceus) J Vet Sci 15(1): 163–166.
Castro N, Osorio CR, Buján N, Fuentes JC, Rodríguez J, Romero M, Jiménez C, Toranzo AE, Magariños B. Insights into the virulence-related genes of Edwardsiella tarda isolated from turbot in Europe: genetic homogeneity and evidence for vibrioferrin production. J Fish Dis. 2016;39(5):565–76.
Reed LJ, Muench H. Simple method of estimating 50% endpoint. Am J Hyg. 1938;27:493–7.
NCCLS (National Committee for Clinical Laboratory Standards). (1999) Performance standard for antimicrobial disk and dilution susceptibility tests for bacteria isolated from animals. Approved Standard M 31A19 (11). NCCLS, Wayne, Pennsylvania.
Krumperman PH. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl Environ Microbiol. 1983;46(1):165–70. https://doi.org/10.1128/aem.46.1.165-170.
Sherif AH, Eldessouki EA, Sabry NM, Ali NG. The protective role of iodine and MS-222 against stress response and bacterial infections during Nile tilapia (Oreochromis niloticus) transportation. Aquac Int. 2022https://doi.org/10.1007/s 10499-022-00984-7.
Eldessouki EA, Salama SSA, Mohamed R, Sherif AH. Using nutraceutical to alleviate transportation stress in the Nile tilapia. Egypt J Aquat Biol Fish. 2023;27(1):413–29. https://doi.org/10.21608/ejabf.2023.287741.
Boijink CDL, Brandão DA, Vargas ACD, Costa MMD, Renosto AV. Inoculação De suspensão Bacteriana De Plesiomonas shigelloides em jundiá, Rhamdia quelen (teleostei: pimelodidae). Ciênc Rural. 2001;31:497–501.
Ruangpan L, Kitao T, Yoshida T. Protective efficacy of Aeromonas hydrophila vaccines in Nile tilapia. Vet Immunol Immunopathol. 1986;12(1–4):345–50.
Bergey DH. Aeromonas. In: Holt JG, Krieg NR, Sneath PHA, Staley JT, Williams ST, editors. Bergey’s manual of determinative bacteriology. Volume 150, 9th ed. Williams and Wilkins; 1994.
bioMerieux. (1984) Laboratory reagents and products. In: Bacterial. Barcy-L. Etiole 69260 charbonmieres-Les-Bams, France.
Rani S. Effect of water temperature on respiratory rate of fish, Cirrhinus mrigala. Indian J Fundam Appl Life Sci. 2016;6(2):13–5.
Noble C, Gismervik K, Iversen MH, Kolarevic J, Nilsson J, Stien LH, Turnbull JF, editors. (2018) Welfare Indicators for farmed Atlantic salmon: tools for assessing fish welfare. Tromsø: Nofima 351pp.
Bancroft DJ, Cook CH, Stirling RW, Turer DR. Manual of histological techniques and their diagnostic applications. Edinburgh, U.K: Churchill Livingstone; 1999.
EL-Bouhy ZM, El-Nobi G, Hassanin ME, Abd EL-Rahman AN. Effect of unionized Ammonia (UIA) on virulence of Clostridium perfringens in Oreochromis niloticus. Zagazig Vet J. 2015;43(3):64–71.
Evans JJ, Pasnik DJ, Brill GC, Klesius PH. Un-ionized ammonia exposure in Nile tilapia: toxicity, stress response, and susceptibility to Streptococcus agalactiae. N Am J Aquac. 2006;68(1):23–33.
Wedemeyer G. Physiology of fish in intensive culture systems. Springer Science & Business Media; 1996.
Fivelstad S, Schwarz J, Strømsnes H, Olsen AB. Sublethal effects and safe levels of ammonia in seawater for Atlantic salmon postsmolts (Salmo salar L). Aquacult Eng. 1995;14(3):271–80.
Algammal AM, Mabrok M, Ezzat M, Alfifi KJ, Esawy AM, Elmasry N, El-Tarabili RM. Prevalence, antimicrobial resistance (AMR) pattern, virulence determinant and AMR genes of emerging multi-drug resistant Edwardsiella tarda in Nile tilapia and African catfish. Aquaculture. 2022;548:737643.
Rao PS, Yamada Y, Leung KY. A major catalase (KatB) that is required for resistance to H2O2 and phagocyte-mediated killing in Edwardsiella tarda. Microbiol. 2003;149(9):2635–44. https://doi.org/10.1099/mic.0.26478-0.
Leung KY, Siame BA, Tenkink BJ, Noort RJ, Mok YK. Edwardsiella tarda–virulence mechanisms of an emerging gastroenteritis pathogen. Microbes Infect. 2012;14(1):26–34. https://doi.org/10.1016/j.micinf.2011.08.005.
Lan J, Zhang XH, Wang Y, Chen J, Han Y. Isolation of an unusual strain of Edwardsiella tarda from turbot and establish a PCR detection technique with the gyrB gene. J Appl Microbiol. 2008;105(3):644–51. https://doi.org/10.1111/j.1365-2672.2008.03779.x.
Das BK, Sahu I, Kumari S, Sadique M, Nayak KK. Phenotyping and whole cell protein profiling of Edwardsiella tarda strains isolated from infected freshwater fishes. Int J Curr Microbiol App Sci. 2014;3(1):235–47.
Selim A, Abdeltwab A, Elwakil R. Biofilm formation Edwardsiella tarda isolated from fresh water fishes. Benha Vet Med J. 2021;40(1):1–5. https://doi.org/10.21608/bvmj.2021.59428.1333.
Abd El-tawab AA, El-Hofy FI, El-Gohary MS, Sedek AA. Edwardesiellosis in freshwater fish with special reference for detection of some virulence genes by PCR. Int J Fish Aquat. 2020;8(5):229–34.
Weigel W. Functional characterization of Aggregatibacter actinomycetemcomitans QSEBC: a bacterial adrenergic receptor and Global Regulator of Virulence. Univ Louisville. 2015. https://doi.org/10.18297/etd/2296.
Moustafa EM, Omar AAED, Abdo WS. Insight into the virulence-related genes of Edwardsiella tarda isolated from cultured freshwater fish in Egypt. World Vet J. 2016;6(3):101–9.
Ibrahem MD, Shaheed I, Yazeed HAE, Korani H. Assessment of the susceptibility of polyculture reared African catfish and Nile tilapia to Edwardsiella tarda. J Am Sci. 2011;7(3):779–86.
Kumar P, Adikesavalu H, Abraham TJ. Prevalence of Edwardsiella tarda in commercially important finfish and shellfish of Bihar and West Bengal, India. J Coast Life Med. 2016;4:30–5. https://doi.org/10.12980/jclm.4.2016apjtd-014-0184.
Adel M, Yeganeh S, Dadar M, Sakai M, Dawood MA. Effects of dietary Spirulina platensis on growth performance, humoral and mucosal immune responses and disease resistance in juvenile great sturgeon (Huso huso Linnaeus, 1754). Fish Shellfish Immunol. 2016;56:436–44. https://doi.org/10.1016/j.fsi.2016.08.003.
Mabrouk MM, Ashour M, Labena A, Zaki MA, Abdelhamid AF, Gewaily MS, … Ayoub HF (2022) Nanoparticles of Arthrospira platensis improves growth, antioxidative and immunological responses of Nile tilapia (Oreochromis niloticus) and its resistance to Aeromonas hydrophila Aquac Res 53(1): 125–135. https://doi.org/10.1111/are.15558.
Rahman M, Al Mamun MA, Rathore SS, Nandi SK, Kari ZA, Wei LS, … Kabir MA (2023) Effects of dietary supplementation of natural Spirulina on growth performance, hemato-biochemical indices, gut health, and disease resistance to Aeromonas hydrophila of Stinging catfish (Heteropneustes fossilis) fingerling. Aquac Rep 32: 101727.
Velasquez SF, Chan MA, Abisado RG, Traifalgar RFM, Tayamen MM, Maliwat GCF, Ragaza JA. Dietary Spirulina (Arthrospira platensis) replacement enhances performance of juvenile Nile tilapia (Oreochromis niloticus). J Appl Phycol. 2016;28:1023–30. https://doi.org/10.1007/s10811-015-0661-y.
Thakur A, Mikkelsen H, Jungersen G. Intracellular pathogens: host immunity and microbial persistence strategies. J Immunol Res. 2019;1356540. https://doi.org/10.1155/2019/1356540.
Mayer-Barber KD, Yan B. Clash of the Cytokine Titans: counter-regulation of interleukin-1 and type I interferon-mediated inflammatory responses. Cell Mol Immunol. 2017;14(1):22–35.
Mahmoud MM, El-Lamie MM, Kilany OE, Dessouki AA. Spirulina (Arthrospira platensis) supplementation improves growth performance, feed utilization, immune response, and relieves oxidative stress in Nile tilapia (Oreochromis niloticus) challenged with Pseudomonas fluorescens. Fish Shellfish Immunol. 2018;72:291–300.
Awad LZ, El-Mahallawy HS, Abdelnaeim NS, Mahmoud MM, Dessouki AA, ElBanna NI. Role of dietary Spirulina platensis and betaine supplementation on growth, hematological, serum biochemical parameters, antioxidant status, immune responses, and disease resistance in Nile tilapia. Fish Shellfish Immunol. 2022;126:122–30.
Song X, Zhao J, Bo Y, Liu Z, Wu K, Gong C. Aeromonas hydrophila induces intestinal inflammation in grass carp (Ctenopharyngodon idella): an experimental model. Aquaculture. 2014;434:171–8.
Nasir NFM, Amal MNA, Omar H, Ismail A, Nasruddin NS. (2018) Growth, body composition and resistance to Aeromonas hydrophila Challenge in Juvenile African Catfish (Clarias gariepinus) Fed diets supplemented with Spirulina (Arthrospira platensis). Annu Res Rev Biol 1–15.
Cao S, Zhang P, Zou T, Fei S, Han D, Jin J, … Xie S (2018) Replacement of fishmeal by spirulina Arthrospira platensis affects growth, immune related-gene expression in gibel carp (Carassius auratus gibelio var. CAS III), and its challenge against Aeromonas hydrophila infection. Fish Shellfish Immunol 79: 265–273.
Ragap HM, Khalil RH, Mutawie HH. Immunostimulant effects of dietary Spirulina platensis on tilapia Oreochromis Niloticus. J Appl Pharm Sci. 2012;2(2):26–31.
Tawfeek WS, Kassab AS, Al-Sokary ET, Abass ME, Sherif AH. Chlorella vulgaris algae ameliorates chlorpyrifos toxicity in Nile tilapia with special reference to antioxidant enzymes and Streptococcus agalactiae infection. Mol Biol Rep. 2024;51(616):1–12. https://doi.org/10.1007/s11033-024-09535-0.
Martins CIM, Galhardo L, Noble C, Damsgard B, Spedicato MT, Zupa W, Beauchaud M, Kulczykowska E, Massabuau JC, Carter T, Planellas SR, Kristiansen T. Behavioural indicators of welfare in farmed fish. Fish Physiol Biochem. 2012;38:17–41.
Mohanty BR, Sahoo PK. Edwardsiellosis in fish: a brief review. J Biosci. 2007;32:1331–44.
Vasquez-Pineros MA, Rondon-Barragan IS, Restrepo-Batancur LF, Eslava-Moncha PR. Clinical and hematological study of experimental infection of Aeromonas hydrophila and Edwardsiella tarda on Tilapia Oreochromis spp. Orinoco Meta. 2010;14(1):0121–3709.
Dong HT, Senapin S, Jeamkunakorn C, Nguyen VV, Nguyen NT, Rodkhum C, … Rattanarojpong T (2019) Natural occurrence of Edwardsiellosis caused by Edwardsiella ictaluri in farmed hybrid red tilapia (Oreochromis sp.) in Southeast Asia. Aquaculture 499:17–23.
Pirarat N, Kobayashi T, Katagiri T, Maita M, Endo M. Protective effects and mechanisms of a probiotic bacterium Lactobacillus rhamnosus against experimental Edwardsiella tarda infection in tilapia (Oreochromis niloticus). Vet Immunol Immunopathol. 2006;113(3–4):339–47.
Acknowledgements
The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Funding
Funding Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Contributions
Lamiaa A. Okasha: Writing original draft, Formal analysis, Data curation. Jehan I. Abdellatief: Writing review & editing, Methodology. Ola H. Abd-Elmegeed: Writing – review & editing. Ahmed H. Sherif: Writing original draft, Methodology.
Corresponding author
Ethics declarations
Ethical approval
The above-described methodology was approved by the Ethics Committee at the Animal Health Research Institute (ARC-AHRI-64-24) and European Union directive 2010/63UE, and all methods were carried out in accordance with relevant guidelines and regulations. This study is reported in accordance with ARRIVE guidelines (https://arriveguidelines.org). This paper does not contain any studies with human participants by any of the authors. No specific permissions were required for access to the artificial pond in wet laboratory Animal Health Research Institute, Kafrelsheikh, Egypt. The field studies did not involve endangered or protected species.
Consent to participate
Not applicable.
Consent to publish
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

{kind=link}
Cite this article
Okasha, L.A., Abdellatif, J.I., Abd-Elmegeed, O.H. et al. Overview on the role of dietary Spirulina platensis on immune responses against Edwardsiellosis among Oreochromis niloticus fish farms. BMC Vet Res 20, 290 (2024). https://doi.org/10.1186/s12917-024-04131-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12917-024-04131-7