
Abstract
Background
The wild boar (Sus scrofa) and the Apennine wolf (Canis lupus italicus) are two wild species that have both increased their presence in the Italian territory, albeit in varying numbers. They can be occasionally found in peri-urban areas as well. Both of these species can serve as intermediate hosts for Toxoplasma gondii, as they can become infected either through the consumption of oocysts found in water, soil, or on vegetables, or through the ingestion of meat containing bradyzoites. Consequently, these animals can be regarded as key indicators of Toxoplasma presence in the wild or peri-urban environment. In our study, we examined a total of 174 wild boar meat juice and 128 wolf sera from Italy for the detection of T. gondii IgG using the indirect fluorescent antibody test (IFAT).
Results
The results showed that 40 (22.6%) of the wild boar meat juice and 34 (26.6%) of the wolf serum samples tested positive. Interestingly, there were no significant differences in seropositivity with respect to gender, age group, or the region of origin in both species.
Conclusions
Overall the results indicate a moderate exposure in both the species under investigation, highlighting the spread of T. gondii in sylvatic and periurban environments. The prevalence of T. gondii in wild boar is consistent with findings from other studies conducted in Europe. Our study, with a considerably larger sample size compared to the available research in European context, provides valuable data on the seroprevalence of T. gondii in wolves.
Similar content being viewed by others

Background
Toxoplasma gondii is a globally distributed apicomplexan protozoan. Its widespread epidemiological success can be attributed to its ability to infect both definitive and intermediate hosts through various modes [1].
Definitive hosts, primarily members of the Felidae family, facilitate the parasite’s sexual reproduction in their intestinal tract, potentially leading to the excretion of millions of oocysts into the environment. In our regions wild and domestic cats play a crucial role in perpetuating this parasite [2]. In Italy, the native European wildcat (Felis silvestris silvestris) maintains a relatively small population size in the wild, despite beingclassified as Least Concern in the IUCN Red List of Threatened Species [3]. However, free-roaming domestic cats are prevalent in rural and peri-urban regions [4].
All warm-blooded vertebrates, including humans, can serve as intermediate hosts in which cysts housing long-lasting bradyzoites develop. These hosts become infected by ingesting sporulated oocysts, although the parasite may persist through predation among them, even in the absence of a definitive host [1].
Omnivorous wild boars (Sus scrofa) are susceptible to infection through two plausible routes: ingestion of highly resistant oocysts present in water and vegetation, and consumption of remains of infected intermediate hosts [5]. Additionally, wild boars represent a potential risk to human health through the consumption of raw or undercooked game meat [6]. The wolf (Canis lupus) can also act as an intermediate host of T gondii. Despite the wolf’s primarily carnivorous diet, which includes predation on live animals, including wild boar, it has been established that they also frequently consume fruits (Rosaceae), other plant matter, and insects [7]. Consequently, both modess of infection are viable in these animals, positioning them at the apex of receptive intermediate hosts range.
The IUCN Red List of Threatened Species has classified the European assessment of Canis lupus as “Least Concern” [8]. In Italy, a subspecies of the grey wolf known as the Apennine wolf (Canis lupus italicus) has seen a population expansion throughout the Italian peninsula in recent years [9], with the exception of the islands. Over the past few decades, both the number and distribution of wolf populations in Italy have increased. Wolves have been progressively reclaiming their historic habitats, moving from the Apennines to the western areas of the Italian Alps [10, 11]. In the past decade, they have also expanded into the eastern Alps [12]. While wolves tend to prefer locations at a considerable distance from human settlements, they have been observed in close proximity to urban areas in densely populated regions [13].”
Despite being among the most heavily hunted ungulate species, wild boars have undergone a population expansion throughout Europe. In Italy, the density of wild boars has been estimated to range from 0.01 to 0.05 animals per square kilometer, increasing to as high as 2.32 to 10.5 animals per square kilometre across the entire Italian peninsula [14]. The simultaneous expansion of human-inhabited areas and the wild boar populations has facilitated the intrusion of this species into various European urban areas, including Rome [15].
In the present study, we conducted a serological survey on wolves and wild boars from different regions of Italy. The objective was to gather data on their exposure to T. gondii infection, serving as indicators of Toxoplasma presence within the wild or peri-urban environment.
Results
The wild boars displayed nearly equal representation across sex and age groups, with a notable portion originating from the Tuscany region (as reported in Table 1). Among the 177 meat juice samples, 40 (22.6%) tested positive for Toxoplasma IgG at IFAT. No statistically significant differences of seropositivity were observed in relation to sex, age groups and region of origin and between wolves and wild boar.
The region of origin of wolves and their cause of death are summarized in Table 2). Thirty-four (26.6%) out of 128 serum analysed, were positive at IFAT, with antibody titres ranging from 1:20 to 1:160. It is noteworthy that no statistically significant differences were observed to seropositivity in relation to sex, age group, geographic origin or cause of death.

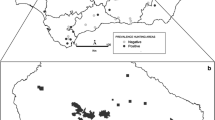
Distribution of wild boar and wolves examined; number positive/number examined
In Fig. 1 the distribution of wolves and wild boar examined are illustrated, with the number of seropositive/number of examined samples in the different Italian provinces.
In the geographical areas where there was an overlap in the sampling of wild boars and wolves (Emilia-Romagna and Tuscany regions), the seroprevalences were 18.9% and 29.6%, respectively, even though the differences were not significant.
Discussion
In this study, we evaluated the seroprevalence of T. gondii in two species, wild boar and wolves. Despite their role as intermediate hosts, these species could play a significant role in mantaining the effective continuity of the parasite’s life cycle in the wild. Both these animals can become infected through the ingestion of robust, environmentally enduring oocysts, as well as via the consumption of prey or carrion. Consequently, they serve as valuable indicators to detect the presence of T. gondii contamination within specific ecological contexts [16, 17].
During this study, we utilized two different matrices: serum samples from wolves and meat juice from wild boar. The choice of these two matrices was driven by practical considerations. In the case of wolves, which were found deceased, we were able to conduct a comprehensive necropsy, including the collection of clotted blood from the heart cavity and subsequent extraction of serum. On the other hand, for wild boars, a different approach was necessary. These animals were hunted and eviscerated before slaughtering, making it impossible to collect blood directly.Therefore, we chose meat juice as a more appropriate and easily accessible matrix in this situation.
This matrix has been used in previous studies for the detection of antibodies against T. gondii [18] as well as other zoonotic pathogens such as Trichinella sp. [19, 20], Salmonella sp., and Hepatitis E virus [21]. The use of meat juice as a matrix is particularly advantageous, as it can be easily obtained from wildlife carcasses, often found deceased, thereby providing valuable serological data that would otherwise be challenging to collect. However, it’s important to note that meat juice has been perceived as a matrix with lower sensitivity in comparison to serum, primarily due to the lower antibody concentration it contains [22]. While serological data derived from either sera or meat juice samples offer insights into an animal’s exposure to the parasite, they do not provide information concerning the presence of tissue cysts within organs, which directly relates to the risk for consumers [23, 24].
In the present study, an overall seroprevalence rate of 22.6% was observed in wild boars (ranging from 0 to 33.3% accross the different regions), and no statistically significant differences were observed among the variables considered, including age, in line with the findings of some authors [25,26,27]. Recent meta-analyses have shown that the global pooled seroprevalence of T. gondii in wild boars from 1995 to 2017 was 23%, which aligns closely with our findings [17]. However, various seroprevalence rates have been documented on wild boars in different geographical settings. For instance, in Europe, seroprevalence values ranging from 8 to 38% have been reported [17, 28,29,30]. Specifically, surveys conducted in central and southern regions of Italy, reported values ranging from 12.2% [31] to 14% [21, 32], while recent surveys in Northern Italy have identified seroprevalences spanning from 15.5% [33] to as high as 53.1% [27]. These seroprevalence differences could be related to specific local epidemiological conditions, such as variations in environmental factors, wildlife populations, or human activities, highlighting the importance of considering local risk factors in understanding the epidemiology of Toxoplasmosis.
In wolves, a seroprevalence rate of 26.6% was observed in this study. When comparing seroprevalences between wolves and wild boars, despite wolves occupying higher trophic levels and exhibiting a higher prevalence of T. gondii, no statistically significant differences were observed between these two populations. This finding aligns with the results of Dakraub et al. [4]. Reliable T. gondii seroprevalence data for wolves in European countries, including Italy, are notably scarce. Recent reports from Italy have indeed documented seropositivity in wolves, albeit with relatively small sample sizes: Dini et al. [33] identified one positive wolf out of 5 samples, while Dakraub et al. [4] reported 4 positives out of 14. In other European countries, such as Spain, a seroprevalence rate of 46.9% was observed (n = 32 wolves sampled) [34]. Due to the considerable higher sample size, the present study offers a comprehensive assessment of T. gondii seroprevalence in wolves, thereby contributing valuable data on a European scale.
In addition to its epidemiological significance, seropositivity in wolves has been associated with ecological implications, particularly in the United States. Recent research [35] demonstrated that the overlap of wolf territories with regions characterized by a high cougar population density serves as a significant predictor of T. gondii infection in wolves. Furthermore, wolves that tested positive through serological analysis were found to be more inclined to make high-risk decisions, such as dispersing and assuming leadership roles within packs [35]. These decisions have a pivotal impact on individual fitness and the broader dynamics of wolf. In the current study, we did not observe a positive correlation between seropositivity and the cause of death being a car crash. Instead, even when considering seropositivity as a factor contributing to increased wolf dispersion, it does not appear to be linked to car collision as cause of death in our sample set.
Conclusion
This study provides an update on the spread of T. gondii in sylvatic and peri urban settings, highlining a moderate exposure in both the species under investigation. Additional research endeavours should be undertaken to explore the correlation between T. gondii seropositivity in wolves and factors like dispersal rates, causes of death, and spatial overlap with other species, including humans. These studies will be able to contribute to a more comprehensive understanding of the significance of T. gondii seroprevalence, including its ecological implications.
Methods
Approximately 25 g of diaphragm tissue from wild boars were systematically collected at a specialized game meat processing facility located in the Bologna province (Emilia-Romagna region). This facility routinely receives eviscerated carcasses of hunted wild boars from various regions of Italy, encompassing Emilia-Romagna, Tuscany, and Abruzzo. Sex, and age class were determined, the latter assessed by the evaluation of the dental table. The diagnostic matrix employed in this study was the meat juice, as carcasses have already been bled and eviscerated. To extract the meat juice from the diaphragm tissue, the samples were placed in hermetically sealed plastic container, and frozen at -20 °C. Following this step, the meat samples were thawed over-night, at a controlled temperature of 4 °C. The resulting meat juice was then transferred into sterile tubes, preserved at -20 °C until use [36].
The examined wolves came mainly from Toscana and Emilia-Romagna region (Central Italy), in less extent they were collected from Calabria (south), Umbria (centre), and Veneto (north) regions. The wolves were found dead and delivered to authorized centers in order to proceed with the necropsy. Necropsy examinations on wolf carcasses were carried out at the Experimental Zooprophylactic Institute of Lombardy and Emilia-Romagna, the Wildlife and Exotic Service of the University of Bologna and at the Experimental Zooprophylactic Institute of Southern Italy. At the arrival of each carcass, a first form containing the following information was filled: subject’s identification data with the attribution of a unique ID code, the discovery location (reported as GPS coordinates), sex, weight (in kg) and nutritional status. The age of the animal was determined by assessing dental development and wear [37, 38], as well as considering body size and weight. Here, all individuals were aged using 3 categories as follows: class 1: ≤12 months; class 2: 1–2 years; class 3: > 2 years. The age determination of class 1 (based on months of life) was defined in relation to the reproductive cycle of the wolf [39]. Besides the biometrics information, phenotypic characteristics and anatomopathological activities were carried out to investigate the cause of death [40]. During necropsy the entire heart was collected, and the heart blood clot was extracted and centrifuged at 980 g for 20 min. The haemolytic serum was then collected in a 2 ml tube and stored at -20 °C until use.
A total of 177 meat juices of wild boars and 128 wolf sera were analysed for T. gondii IgG by indirect fluorescent antibody test (IFAT) following the manufacturer’s instructions (MegaFLUO TOXO-PLASMA g, MegaCor Diagnostik, Hoerbranz, Austria). As conjugated, anti-dog IgG antibody diluted in PBS at concentration of 1:64 (Anti-Dog IgG-FITC antibody, Sigma-Aldrich, Saint Louis, MO) and anti-pig. IgG antibody diluted in PBS at concentration of 1:32 (Anti-pig IgG-FITC antibody, Sigma-Aldrich, Saint Louis, MO) were used. Meat juice from wild boars with an antibody titre ≥ 1:4 were considered positive (due to the scarce concentration of antibody in this matrix) [22], while wolf serum samples with antibody titre ≥ 1:20 were considered positive (due to the haemolytic characteristics of the sera) [41].
Pearson’s χ2 test was used to correlate sex, age group, region of origin (and cause of death in wolf) with seroprevalence. Statistical significance was set at P ≤ 0.05. The Sample Size Calculator (https://www.surveysystem.com/sscalc.htm) was used to calculate 95% confidence intervals (CIs) for the observed prevalence values.
Data availability
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Tenter AM, Heckeroth AR, Weiss LM. Toxoplasma gondii: From animals to humans. Int J Parasitol. 2000;30(12–13):1217–58. https://doi.org/10.1016/s0020-7519(00)00124-7.
Montazeri M, Mikaeili Galeh T, Moosazadeh M, Sarvi S, Dodangeh S, Javidnia J, Sharif M, Daryani A. The global serological prevalence of Toxoplasma gondii in felids during the last five decades (1967–2017): A systematic review and meta-analysis. Parasit Vectors. 2020;13(1):82. https://doi.org/10.1186/s13071-020-3954-1.
Gerngross P, Ambarli H, Angelici FM, Anile S, Campbell R, Ferreras de Andres P, Gil-Sanchez JM, Götz M, Jerosch S, Mengüllüoglu D, Monterosso P, Zlatanova D. Felis silvestris. IUCN Red List Threatened Species: e. 2022. https://doi.org/10.2305/IUCN.UK.2022-1.RLTS.T181049859A181050999.en. Accessed 17 Oct 2023. T181049859A181050999.
Dakroub H, Sgroi G, D’Alessio N, Russo D, Serra F, Veneziano V, Rea S, Pucciarelli A, Lucibelli MG, De Carlo E, Fusco G, Amoroso MG. Molecular survey of Toxoplasma gondii in wild mammals of Southern Italy. Pathogens. 2023;12(3):471. https://doi.org/10.3390/pathogens12030471.
Guardone L, Armani A, Mancianti F, Ferroglio E. A review on Alaria alata, Toxoplasma gondii and Sarcocystis spp. in Mammalian game meat consumed in Europe: Epidemiology, risk management and future directions. Animals (Basel). 2022;12(3):263. https://doi.org/10.3390/ani12030263. PMID: 35158587.
Kornacka-Stackonis A. Toxoplasma Gondii infection in wild omnivorous and carnivorous animals in Central Europe - a brief overview. Vet Parasitol. 2022;304:109701. https://doi.org/10.1016/j.vetpar.2022.109701.
Meriggi A, Rosa P, Brangi A, Matteucci C. Habitat use and diet of the wolf in Northern Italy. Acta Theriol. 1991;36:141–51.
IUCN, Lista Rossa dei vertebrati italiani. 2022. https://www.iucn.it/pdf/Lista-Rossa-vertebratiitaliani-2022.pdf.
La Morgia V, Marucco F, Aragno P, Salvatori V, Gervasi V, De Angelis D, Fabbri E, Caniglia R, Velli E, Avanzinelli E, Boiani MV, Genovesi P. Stima della distribuzione e consistenza del lupo a scala nazionale 2020/2021. Relazione tecnica realizzata nell’ambito della convenzione ISPRA-Ministero della Transizione Ecologica Attività di monitoraggio nazionale nell’ambito del Piano di Azione del lupo. 2022. https://www.isprambiente.gov.it/it/attivita/biodiversita/monitoraggio-nazionale-del-lupo/file-monitoraggio/report-nazionale-lupo-20_21.pdf.
Scandura M, Apollonio M, Mattioli L. Recent recovery of the Italian wolf population: A genetic investigation using microsatellites. Mamm Biol. 2001;66:321–31.
Fabbri E, Miquel C, Lucchini V, Santini A, Caniglia R, Duchamp C, Weber JM, Lequette B, Marucco F, Boitani L, Fumagalli L, Taberlet P, Randi E. From the Apennines to the alps: Colonization genetics of the naturally expanding Italian wolf (Canis lupus) population. Mol Ecol. 2007;16(8):1661–71. https://doi.org/10.1111/j.1365-294X.2007.03262.x.
Marucco F, Avanzinelli E, Boiani MV, Menzano A, Perrone S, Dupont P, Bischof R, Milleret C, von Hardenberg A, Pilgrim K, Friard O, Bisi F, Bombieri G, Calderola S, Carolfi S, Chioso C, Fattori U, Ferrari P, Pedrotti L, Righetti D, Tomasella M, Truc F, Aragno P, La Morgia V, Genovesi P. La Popolazione Di Lupo Nelle Regioni alpine italiane 2020–2021. Relazione Tecnica Dell’Attività Di Monitoraggio Nazionale Nell’ambito del piano di Azione Del Lupo Ai Sensi della Convenzione ISPRA-MITE e nell’ambito del Progetto LIFE 18 NAT/IT/000972 WOLFALPS EU. 2022.
Zanni M, Brogi R, Merli E, Apollonio M. The wolf and the city: Insights on wolves conservation in the anthropocene. Anim Conserv. 2023;1–15. https://doi.org/10.1111/acv.12858.
Pittiglio C, Khomenko S, Beltran-Alcrudo D. Wild boar mapping using population-density statistics: From polygons to high resolution raster maps. PLoS ONE. 2018;13(5):e0193295. https://doi.org/10.1371/journal.pone.0193295.
Amendoli S, Lombardini M, Pierucci P, Meriggi A. Seasonal spatial ecology of the wild boar in a peri-urban area. Mammal Res. 2019;64:387–96. https://doi.org/10.1007/s13364-019-00422-9.
Beral M, Rossi S, Aubert D, Gasqui P, Terrier ME, Klein F, Villena I, Abrial D, Gilot-Fromont E, Richomme C, Hars J, Jourdain E. Environmental factors associated with the seroprevalence of Toxoplasma gondii in Wild Boars (Sus scrofa), France. EcoHealth. 2012;9(3):303–9. https://doi.org/10.1007/s10393-012-0786-2.
Rostami A, Riahi SM, Fakhri Y, Saber V, Hanifehpour H, Valizadeh S, Gholizadeh M, Pouya RH, Gamble HR. The global seroprevalence of Toxoplasma gondii among wild boars: A systematic review and meta-analysis. Vet Parasitol. 2017;244:12–20. https://doi.org/10.1016/j.vetpar.2017.07.013.
Opsteegh M, Swart A, Fonville M, Dekkers L, van der Giessen J. Age-related Toxoplasma gondii seroprevalence in Dutch wild boar inconsistent with lifelong persistence of antibodies. PLoS ONE. 2011;6(1):e16240. https://doi.org/10.1371/journal.pone.0016240.
Calero-Bernal R, Pérez-Martín JE, Reina D, Serrano FJ, Frontera E, Fuentes I, Dubey JP. Detection of Zoonotic Protozoa Toxoplasma gondii and Sarcocystis suihominis in wild boars from Spain. Zoonoses Public Health. 2016;63(5):346–50. https://doi.org/10.1111/zph.12243.
Villa L, Allievi C, Gazzonis AL, Ventura G, Gradassi M, Zanzani SA, Manfredi MT. Serological prevalence of Toxoplasma gondii, Neospora caninum, and Sarcoptes scabiei var. suis in wild boars (Sus scrofa) hunted in a highly anthropized area in Italy. Anim (Basel). 2023;13(11):1730. https://doi.org/10.3390/ani13111730.
Kijlstra A, Jongert E. Control of the risk of human toxoplasmosis transmitted by meat. Int J Parasitol. 2008;38(12):1359–70. https://doi.org/10.1016/j.ijpara.2008.06.002.
Puchalska M, Pyziel AM, Wiśniewski J, Steiner-Bogdaszewska Ż, Klich D, Anusz K. Prevalence of Toxoplasma gondii antibodies in wild boar (Sus scrofa) from Strzałowo Forest Division, Warmia and Mazury Region, Poland. Ann Agric Environ Med. 2021;28(2):237–42. https://doi.org/10.26444/aaem/118883.
Olsen A, Berg R, Tagel M, Must K, Deksne G, Enemark HL, Alban L, Johansen MV, Nielsen HV, Sandberg M, Lundén A, Stensvold CR, Pires SM, Jokelainen P. Seroprevalence of Toxoplasma gondii in domestic pigs, sheep, cattle, wild boars, and moose in the Nordic-Baltic region: A systematic review and meta-analysis. Parasite Epidemiol Control. 2019;5:e00100. https://doi.org/10.1016/j.parepi.2019.e00100.
Papini RA, Vannucci S, Rocchigiani G, Nardoni S, Mancianti F. Prevalence of Toxoplasma gondii and potentially zoonotic helminths in wild boars (Sus scrofa) hunted in central Italy. Mac Vet Rev. 2018;41:83–93.
Ranucci D, Veronesi F, Moretti A, Branciari R, Miraglia D, Manfredi MT, Piergili Fioretti D. Seroprevalence of Toxoplasma gondii in wild boars (Sus scrofa) from Central Italy. Parasite. 2013;20:48. https://doi.org/10.1051/parasite/2013048.
Sarno E, Costanzo N, Quaranta V, Santoro AML, Stephan R. Prevalence of IgG against hepatitis E virus, Salmonella spp., and Toxoplasma gondii in meat juice samples from wild boars hunted in Southern Italy. J Food Saf Food Qual. 2014;65:141–4.
Dini FM, Morselli S, Marangoni A, Taddei R, Maioli G, Roncarati G, Balboni A, Dondi F, Lunetta F, Galuppi R. Spread of Toxoplasma gondii among animals and humans in Northern Italy: A retrospective analysis in a one-health framework. Food Waterborne Parasitol. 2023;32:e00197. https://doi.org/10.1016/j.fawpar.2023.e00197.
Halos L, Thébault A, Aubert D, Thomas M, Perret C, Geers R, Alliot A, Escotte-Binet S, Ajzenberg D, Dardé ML, Durand B, Boireau P, Villena I. An innovative survey underlining the significant level of contamination by Toxoplasma gondii of ovine meat consumed in France. Int J Parasitol. 2010;40(2):193–200. https://doi.org/10.1016/j.ijpara.2009.06.009.
Nöckler K, Serrano FJ, Boireau P, Kapel CM, Pozio E. Experimental studies in pigs on Trichinella detection in different diagnostic matrices. Vet Parasitol. 2005;132(1–2):85–90. https://doi.org/10.1016/j.vetpar.2005.05.033.
Frey CF, Schuppers ME, Nöckler K, Marinculić A, Pozio E, Kihm U, Gottstein B. Validation of a western blot for the detection of anti-Trichinella spp. antibodies in domestic pigs. Parasitol Res. 2009;104(6):1269–77. https://doi.org/10.1007/s00436-008-1321-9.
Carella E, Caruso C, Moreno A, Di Blasio A, Oberto F, Vitale N, Masoero L. Meat juice and oral fluid as Alternatives to serum for Aujeszky Disease Monitoring in pigs. Microorganisms. 2023;11(10):2418. https://doi.org/10.3390/microorganisms11102418.
Pinto P, Bozzo G, Novello L, Terio V. Detection of Toxoplasma gondii cysts from wild boar muscles: Does it represent a risk for ready to eat food? IJFS. 2010 Oct. 27,1(8):31–4. https://www.pagepressjournals.org/index.php/ijfs/article/view/ijfs.2010.8.31.
Gazzonis AL, Gjerde B, Villa L, Minazzi S, Zanzani SA, Riccaboni P, Sironi G, Manfredi MT. Prevalence and molecular characterisation of Sarcocystis miescheriana and Sarcocystis suihominis in wild boars (Sus scrofa) in Italy. Parasitol Res. 2019;118(4):1271–87. https://doi.org/10.1007/s00436-019-06249-2.
Sobrino R, Cabezón O, Millán J, Pabón M, Arnal MC, Luco DF, Gortázar C, Dubey JP, Almeria S. Seroprevalence of Toxoplasma gondii antibodies in wild carnivores from Spain. Vet Parasitol. 2007;148(3–4):187–92. https://doi.org/10.1016/j.vetpar.2007.06.038.
Meyer CJ, Cassidy KA, Stahler EE, Brandell EE, Anton CB, Stahler DR, Smith DW. Parasitic infection increases risk-taking in a social, intermediate host Carnivore. Commun Biol. 2022;5(1):1180. https://doi.org/10.1038/s42003-022-04122-0.
Gipson PS, Ballard WB, Nowak RM, Mech LD. Accuracy and precision of estimating age of gray wolves by tooth wear. J Wildl Manag. 2000;64(3):752–8.
Brasington TJ, Hadley JM, Stahler DR, Stahler EE, Cassidy KA. A visual guide to wolf dentition and age determination. Wildl Biology, https://www.researchgate.net/publication/374921532_A_Visual_Guide_to_Wolf_Dentition_and_Age_Determination_For_Researchers_and_Wildlife_Professionals (2023).
Boitani L. Lupo Canis lupus. In: Pavan M, editors, Distribuzione e biologia di 22 specie di Mammiferi in Italia Rome (IT) IBS; 1981. p 61–67. https://www.ibs.it/distribuzionebiologia-di-22-specie-libri-vintage-vari/e/2560038844628.
Musto C, Cerri J, Galaverni M, Caniglia R, Fabbri E, Apollonio M, Mucci N, Bonilauri P, Maioli G, Fontana MC, Gelmini L, Prosperi A, Rossi A, Garbarino C, Fiorentini L, Ciuti F, Berzi D, Merialdi G, Delogu M. Men and wolves: Anthropogenic causes are an important driver of wolf mortality in human-dominated landscapes in Italy. Glob Ecol Conserv. 2021;e01892. https://doi.org/10.1016/j.gecco.2021.e01892.
Meemken D, Tangemann AH, Meermeier D, Gundlach S, Mischok D, Greiner M, Klein G, Blaha T. Establishment of serological herd profiles for zoonoses and production diseases in pigs by meat juice multi-serology. Prev Vet Med. 2014;113(4):589–98. https://doi.org/10.1016/j.prevetmed.2013.12.006.
Nardoni S, Rocchigiani G, Varvaro I, Altomonte I, Ceccherelli R, Mancianti F. Serological and molecular investigation on Toxoplasma gondii infection in wild birds. Pathogens. 2019;8(2):58. https://doi.org/10.3390/pathogens8020058.
Acknowledgements
The authors are grateful to Dr. Bruno Marasco from AUSL of Bologna, for his availability in organizing the sampling of wild boar, and Prof. Monica Caffara for the English revision.
Funding
The co-author Carmela Musto was partially supported by a research grant funded by the Vienna Science and Technology Fund (WWTF) [https://doi.org/10.47379/ESR20009].
Author information
Authors and Affiliations
Contributions
F.M.D. and R.G. designed the study and analysed data, FMD., C.M., V.M.D.N., E.B., M.S., G.M., L.B., M.D. contributed to collecting the samples. FMD carried out the analysisF.M.D. and R.G. assembled data, RG. was the supervisor, F.M.D, R.G., E.B., C.M. contributed to writing, reviewing and editing the final manuscript. All authors read and approved thefinal manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All samples were gathered in compliance with local regulations. Specifically, wild boar samples were collected by an official veterinarian as part of the official Trichinella sampling for game meat. As for wolves, the local authority collected all animals, and necropsies were conducted at specialized centers (such as IZS or University institutions) dedicated to wildlife disease control. All the methods were performed in accordance with relevant guidelines and regulations, no specific permission was required to perform the sampling.
Animal ethics declaration
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Dini, F.M., Musto, C., De Nigris, V.M. et al. Sero-epidemiological investigation on Toxoplasma gondii infection in Apennine wolf (Canis lupus italicus) and wild boar (Sus scrofa) in Italy. BMC Vet Res 20, 62 (2024). https://doi.org/10.1186/s12917-024-03922-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12917-024-03922-2