
Abstract
Background
The application of plant extracts has received great interest for the treatment of bovine mastitis. Isoliquiritigenin (ISL) is a rich dietary flavonoid that has significant antioxidative, anti-inflammatory and anticancer activities. This study was conducted to explore the protective efficacy and related mechanism of ISL against lipopolysaccharide (LPS)-stimulated oxidation and inflammation in bovine mammary epithelial cells (MAC-T) by in vitro experiments.
Results
Real-time PCR and ELISA assays indicated that ISL treatment at 2.5, 5 and 10 μg/mL significantly reduced the mRNA and protein expression of the oxidative indicators cyclooxygenase-2 and inducible nitric oxide synthase (P < 0.01), and of the inflammatory cytokines interleukin-6 (P < 0.05), interleukin-1β (P < 0.01) and tumor necrosis factor-α (P < 0.01) in LPS-stimulated MAC-T cells. Moreover, Western blotting and immunofluorescence tests indicated that the phosphorylation levels of nuclear factor kappa (NF-κB) p65 and the inhibitor of NF-κB were significantly decreased by ISL treatment, thus blocking the nuclear transfer of NF-κB p65. In addition, ISL attenuated the phosphorylation levels of p38, extracellular signal-regulated kinase and c-jun NH2 terminal kinase.
Conclusions
Our data demonstrated that ISL downregulated the LPS-induced inflammatory response in MAC-T cells. The anti-inflammatory and antioxidative activity of ISL involves the NF-κB and MAPK cascades.
Similar content being viewed by others

Background
Bovine mastitis is considered the most costly disease in dairy cattle due to inflammation of the mammary gland. Bovine mastitis is caused by infection with microbial pathogens and physical, environmental and genetic factors as well [1]. Escherichia coli is one of the most common agents of clinical mastitis. Lipopolysaccharide (LPS) is the major component of the outer membrane in Escherichia coli [2], and it activates the nuclear factor-κB (NF-κB) pathway via Toll-like receptor 4 (TLR4) dimerization [3]. Mitogen-activated protein kinases (MAPKs) are also fundamental in the control of the inflammatory response through the crosstalk with the NF-κB pathway [3]. Both MAPK and NF-κB activation promote the production of proinflammatory factors IL-1β, IL-6 and TNF-α as well as inflammatory mediators cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) [4, 5].
Antibiotics are widely applied in mastitis treatment; however, the abuse of antibiotics results in drug-resistant bacteria and antibiotic residues in the food chain that are harmful to consumers’ health [6]. As plant active ingredients are nontoxic and have medicinal and health benefits, their application in inflammation treatment has received widespread attention.
Isoliquiritigenin (ISL) is a flavonoid with a chalcone structure extracted from licorice. ISL has been reported to have pharmacological activities, including antioxidation, anti-inflammation, antiplatelet aggregation and antineoplastic properties [7, 8]. In vitro experiments have demonstrated that ISL attenuates the inflammatory response of macrophages by suppressing the homodimerization of TLR4 [9]. In vivo studies revealed that ISL inhibited NF-κB activation in septic mice, thus reducing the expression of IL-6, TNF-α and COX-2 [10, 11]. Additionally, upstream signaling pathways, including the phosphorylation of p38 in the MAPK pathway and DNA binding of NF-κB p65, were prohibited by ISL [12].
Despite these encouraging studies, it is still unclear whether ISL has a beneficial function in LPS-induced bovine mastitis. Here, we investigated the potential preventive effects of ISL in LPS-induced bovine mammary epithelial cells (MAC-T). In particular, we determined the molecular mechanism underlying the antioxidative and anti-inflammatory effects of ISL.
Results
Cytotoxicity of ISL in MAC-T cells
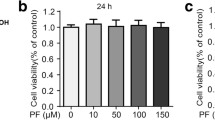
The cytotoxicity of ISL in MAC-T cells was examined by MTT assay. ISL had no effect on the viability of MAC-T cells at concentrations of 2.5, 5 and 10 μg/mL; however, cell viability was significantly decreased upon 24 h treatment with ISL at 20 and 40 μg/mL (P < 0.05, Fig. 1A). Thus, ISL concentrations of 2.5, 5 and 10 μg/mL, which showed no cytotoxicity towards MAC-T cells, were applied in further studies. To demonstrate that there was no endogenous endotoxin present and only the effects of ISL on MAC-T cells, endogenous endotoxin was tested in the cells administered ISL at 0, 2.5, 5, 10 and 20 μg/mL (Fig.1B). The results showed much lower levels of endotoxin in the blank group than in the ISL groups (P < 0.01), and the endotoxin level in each cell group was below the threshold of cytotoxic response (< 200 ng/L), indicating that the plant compound is free of endotoxin.

Viability and endotoxin test of MAC-T cells by MTT assay after 24 h of treatment with isoliquiritigenin (ISL). A Cell viability was measured using the MTT assay, where the viability of nontreatment control cells was set as 100%. B Cell endotoxin (ET) was tested using ELISA. Values represent the means ± SEM of three independent experiments. In the above bars, the same letters indicate P > 0.05; different lowercase letters indicate P < 0.05 while different uppercase letters indicate P < 0.01
Anti-inflammatory and antioxidative effects of ISL in MAC-T cells
The inflammatory effect stimulated by LPS in MAC-T cells was measured using the mRNA levels of IL-6. IL-6 mRNA levels were significantly higher in LPS-treated cells than in controls (P < 0.01), but there was no difference in IL-6 mRNA levels upon treatment with 1, 5, 10 and 20 μg/mL LPS (P > 0.05, Supplementary Fig. 1). Therefore, 1 μg/mL LPS was applied for further studies [3, 13].
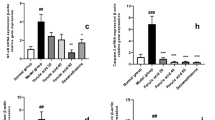
Furthermore, real-time PCR and ELISA were used to detect the expression of inflammatory markers and the inhibitory effect of ISL on inflammation. The expression of TNF-α, IL-1β and IL-6 at the mRNA and protein levels was significantly upregulated in the LPS-treated cells compared with the sham-treated cells (P < 0.01, Fig. 2). When ISL was administered, the mRNA levels of TNF-α (P < 0.01), IL-1β (P < 0.01) and IL-6 (P < 0.05) was significantly decreased, and ISL had better effects at 10 μg/mL than DEX (Fig. 2A-C). Importantly, ISL significantly inhibited the protein expression of these inflammatory biomarkers in LPS-stimulated MAC-T cells (P < 0.01, Fig. 2D-F), supporting the results of qRT-PCR experiments.

Expression analysis of proinflammatory cytokines in MAC-T cells. Cells were treated with lipopolysaccharide (LPS) at 1 μg/mL for 24 h in combination with dexamethasone (DEX) (20 μg/mL) or isoliquiritigenin (ISL) (2.5, 5 and 10 μg/mL). With β-actin as an endogenous control, real-time PCR and ELISA were used to detect mRNA (A-C) and protein (D-F) levels. Values represent the means ± SEM of four independent experiments. In the above bars, ** indicate significance at P < 0.01 between the control and the LPS treatment without DEX and ISL. Among LPS in combination with DEX and ISL treatments, the same letters indicate P > 0.05; different lowercase letters indicate P < 0.05 while different uppercase letters indicate P < 0.01
In addition, we investigated ISL influence on the expression of iNOS and COX-2, which have been reported as oxidative stress indicators. The results showed that both DEX and ISL significantly reduced LPS-induced iNOS and COX-2 mRNA abundance in MAC-T cells (P < 0.01, Fig. 3A-B). ELISA also showed the inhibitory effect of ISL on iNOS and COX-2 expression in LPS-stimulated MAC-T cells (Fig. 3C-D).

Expression analysis of COX-2 and iNOS in MAC-T cells. Cells were incubated with 1 μg/mL lipopolysaccharide (LPS) in combination with 20 μg/mL dexamethasone (DEX) or isoliquiritigenin (ISL) at 2.5, 5 and 10 μg/mL for 24 h. With β-actin as an endogenous control, real-time PCR and ELISA were used to detect mRNA (A and B) and protein (C and D) levels, respectively. Values represent the means ± SEM of four independent experiments. In the above bars, ** indicate significance at P < 0.01 between the control and the LPS treatment without DEX and ISL. Among LPS in combination with DEX and ISL treatments, the same letters indicate P > 0.05; different lowercase letters indicate P < 0.05 while different uppercase letters indicate P < 0.01
Effects of ISL on LPS-stimulated activation of MAPK and NF-κB pathways
We estimated how the NF-κB pathway, which is critical for the production of proinflammatory cytokines, was influenced. As shown in Fig. 4A-B and Supplementary Fig. 2A, the protein levels of p65, which is a key member in the NF-κB pathway, did not differ between LPS, DEX and ISL treatment, but the levels of phosphorylated p65 (p-p65) was significantly decreased in cells treated with ISL (2.5, 5 and 10 μg/mL) and DEX (P < 0.01). Moreover, ISL at 5 and 10 μg/mL increased the protein levels of IκB, which is another key member of the NF-κB pathway (P < 0.01, Fig. 4C). In addition, ISL markedly inhibited p-IκB levels increased by LPS (Fig. 4D) and dose-dependently decreased the p-p65/p65 and p-IκB/IκB ratios (P < 0.01, Supplementary Fig. 2B-C).

Detection of NF-κB activity in MAC-T cells by Western blotting. Cells were coincubated with 1 μg/mL lipopolysaccharide (LPS) and dexamethasone (DEX) (20 μg/mL) or isoliquiritigenin (ISL) (2.5, 5 and 10 μg/mL) for 24 h. The levels of p65 (A), p-p65 (B), IκB (C) and p-IκB (D) proteins were relative to β-actin. Values represent the means ± SEM of three independent experiments. In the above bars, ** indicate significance at P < 0.01 between the control and the LPS treatment without DEX and ISL. Among LPS in combination with DEX and ISL treatments, the same letters indicate P > 0.05; different lowercase letters indicate P < 0.05 while different uppercase letters indicate P < 0.01
We further determined the phosphorylation of MAPK members, critical cascades upstream of proinflammatory mediators and NF-κB. ISL administration significantly reduced LPS-induced activation of p-JNK, p-p38, and p-ERK (P < 0.01), as revealed by both their phosphorylation/total protein ratios (P < 0.01, Supplementary Fig. 3) and their phosphorylation/β-actin ratios (Fig. 5).

Detection of MAPK activity in MAC-T cells using Western blotting. Cells were coincubated with 1 μg/mL lipopolysaccharide (LPS) and dexamethasone (DEX) (20 μg/mL) or isoliquiritigenin (ISL) (2.5, 5 and 10 μg/mL) for 24 h. (A-F) The protein levels of ERK1/2, p38 , JNK, p-ERK, p-p38 and p-JNK were relative to those of β-actin, respectively. Values represent the means ± SEM of three independent experiments. In the above bars, ** indicate significance at P < 0.01 between the control and the LPS treatment without DEX and ISL. Among LPS in combination with DEX and ISL treatments, the same letters indicate P > 0.05; different lowercase letters indicate P < 0.05 while different uppercase letters indicate P < 0.01
Effect of ISL on LPS-stimulated p65 nuclear translocation
Finally, we examined the nuclear translocation of NF-κB p65 in MAC-T cells challenged with LPS using an inverted fluorescence microscope. In contrast to the uniform distribution of red fluorescently labeled p65 in the cytoplasm in the control cells, LPS administration led to p65 accumulation in the blue-labeled nuclei (Fig. 6A). However, LPS-enhanced translocation of p65 was prohibited by ISL treatment at 2.5, 5 and 10 μg/mL (Fig. 6B), similar to the effect of DEX (Fig.6A).

Immunofluorescence analysis (A) and quantification (B) of p65 nuclear translocation in MAC-T cells. Cells treated with LPS (1 μg/mL) were coincubated with DEX (20 μg/mL) or ISL (2.5, 5 and 10 μg/mL) for 24 h. p-p65 was labeled red with Cy3, while the nucleus was marked blue with DAPI. Values represent the means ± SEM of four independent experiments. In the above bars, ** indicate significance at P < 0.01 between the control and the LPS treatment without DEX and ISL. Among LPS in combination with DEX and ISL treatments, the same letters indicate P > 0.05; different lowercase letters indicate P < 0.05 while different uppercase letters indicate P < 0.01
The p65 fraction in the nucleus was further detected. As shown in Fig. 6C, LPS stimulation resulted in higher levels of p65 in the nucleus than that of the nontreatment control (P < 0.01). However, the LPS-induced nuclear content of p65 was significantly reduced after the administration of 2.5, 5 and 10 μg/mL ISL (P < 0.01), supporting the immunofluorescence results.
Discussion
To the best of our knowledge, this study is the first to demonstrate the anti-inflammatory effect of ISL in MAC-T cells. Bovine mastitis is often involved in bacterial infections during parturition and early lactation in bovine mammary glands [14]. The MAC-T cell line was established by transfection of simian virus-40 large T-antigen into bovine mammary epithelial cells; thus, the cell line has a biological response similar to that of primary cells [15] and is used as a model for studying bovine mammary gland inflammation. To this end, we chose MAC-T cells to clarify their response to ISL in this study.
It has been reported that LPS, the main component of cell membranes in Escherichia coli, triggers a comprehensive immune response in MAC-T cells via TLR4/NF-κB and MAPK cascades [16, 17]. Ma et al [13] indicated an inflammatory response of MAC-T cells after stimulation with 1 μg/ml LPS. Additionally, 1 μg/mL LPS markedly increased the mRNA and protein levels of IL-6, IL-1β and TNF-α (Fig. 2) in this study. Accordingly, 1 μg/mL LPS and ISL at 2.5, 5 and 10 μg/mL without cytotoxicity in MAC-T cells (Fig. 1) were chosen.
Numerous studies have shown that ISL can reduce the production of proinflammatory factors, including IL-6, IL-1β and TNF-α, by blocking TLR4 binding to LPS [17,18,19]. Our data also showed that ISL downregulated the expression of these proinflammatory factors in LPS-stimulated MAC-T cells (Fig. 2), suggesting the anti-inflammatory effect of ISL. Although there is no report involving ISL efficacy in mammary epithelial cells, the anti-inflammatory effects of ISL in oral squamous cell carcinoma and intestinal and retinal epithelial cells [20,21,22,23] are consistent with our results in MAC-T cells.
Other studies have shown that ISL can reduce the expression of COX-2 and iNOS in septic mice [11] and HT-29 cells [22]. COX-2 is an inducible enzyme activated by cytokines and growth factors, such as IL-1 and IL-6, while iNOS is a nitric oxide synthetase activated by pathologic stimuli. Induction of COX-2 and iNOS is responsible for the cell levels of cytotoxic mediators prostaglandin (PG) and NO, respectively [24,25,26]. Hence, both COX-2 and iNOS are considered markers of oxidative stress and inflammation. The finding that ISL downregulated LPS-enhanced expression of COX-2 and iNOS in MAC-T cells (Fig. 3) indicates the antioxidative effect of ISL on bovine mastitis.
It has been well documented that ISL inhibits NF-κB activity in neurological inflammation, hypertensive renal injury, traumatic brain injury and other inflammatory diseases in rats [11, 27, 28]. NF-κB was first discovered in B cell nuclear proteins bound to the kappa enhancer of the immunoglobulin kappa light chain gene, and it is involved in a series of physiological activities, such as cell growth, inflammation and immune response [29, 30]. NF-κB exists as an inactive complex with IκBα in the cytoplasm. Activation of NF-κB leads to IκB phosphorylation by IκB kinase (IKK) and subsequent degradation via ubiquitination, thereby releasing and translocating NF-κB p65 to the nucleus, where the transcription factor promotes transcription of its target genes, including proinflammatory cytokines, iNOS and COX-2 [31]. Our data revealed that ISL increased the abundance of IκB, but lowered the phosphorylation of IκB and p65 in LPS-stimulated cells (Fig. 4 and Supplementary Fig. 2), consequently blocking p65 translocation to the nucleus (Fig. 6 ). Therefore, the reduced expression of LPS-induced TNF-α, IL-6, IL-1β, iNOS and COX-2 can be due to the suppression of NF-κB activity.
MAPKs are a group of serine/threonine protein kinases that can be activated by a variety of stimuli and regulate the phosphorylation of downstream signaling pathways [32, 33]. MAPKs, consisting of p38, ERK and JNK, are involved in a cascade regulating NF-κB-mediated transcription of the proinflammatory cytokines, i.e. IL-6, IL-1β and TNF-α, and inflammatory mediators, such as NO, iNOS, COX-2 and prostaglandin E2 [24, 34, 35]. Previous reports indicated that ISL inhibited MAPK activities in human liver cancer and colitis [36, 37]. Our results demonstrated significant inhibition of ISL on LPS-induced phosphorylation of p38, ERK1/2 and JNK in MAC-T cells (Fig. 5). These results indicate the involvement of ISL in MAPK-mediated anti-inflammatory and antioxidative effects.
In the current study, we revealed that ISL attenuated the inflammatory and oxidative response in vitro, but did not investigated the in vivo efficacy of ISL. Especially, we have not carried out clinical trials to explore the clinical efficacy of ISL in the treatment of bovine mastitis at present. Nevertheless, findings from previous studies have highlighted the anti-inflammatory and antioxidative activities of ISL via the NF-κB, MAPK or other pathways in rats with carrageenan-induced pleurisy [38] and in mice with dextran sulfate sodium-induced colitis [37]. The above data together with the findings from the present studies will help to determine the potential application of ISL in the prevention and treatment of bovine mastitis.
Conclusions
ISL downregulats LPS-induced expression of inflammatory cytokines, i.e. TNF-α, IL-1β and IL-6, and oxidative stress indicators, which include iNOS and COX-2, in MAC-T cells. The anti-inflammatory effect of ISL involves the inhibition of the NF-κB and MAPK pathways.
Methods
Reagents and chemicals
ISL and dexamethasone (DEX) were provided by Macleans (Shanghai, China). LPS from Escherichia coli 055:B5 serotype, purity ≥ 99%, item number: L8880) and MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazole bromide] were provided by Solarbio (Beijing, China). The fluorescence quantification kits were purchased from Takara (Beijing, China). Primary antibodies were obtained commercially, including p-p65, p-IκBα, p65, IκBα, p-ERK1/2 and ERK1/2 (Bioss, Beijing, China) as well as JNK, p38, p-JNK and p-p38 (Wanleibio, Shenyang, China). Fetal bovine serum (FBS) and Dulbecco's modified Eagle's medium (DMEM) were supplied by Gibco (Suzhou, China). The ELISA and NF-κB Activation Nuclear Transport Test Kits were available from SinoBestBio (Shanghai, China) and Beyotime (Shanghai, China), respectively. Tris-buffered saline plus Tween 20 (TBST) was obtained from Solarbio (Shanghai, China).
Cell culture and treatments
MAC-T cells were kept in our laboratory and were seeded at 37°C and 5% CO2 in an incubator in DMEM basic, which contained 10% FBS, 100 U/mL streptomycin and 100 μg/mL penicillin. The cells were grown to 90-100% confluency at logarithmic phase, and then the cells were administered with 1 μg/mL LPS combined with 2.5, 5, or 10 μg/mL ISL in five replicates for 24 h. LPS (1 μg/mL) was used according to recent reports [3, 4], and ISL concentrations were determined from the cell viability experiments. Cells in LPS (1 μg/mL) without ISL and in LPS with 20 μg/mL of DEX, a drug for anti-inflammation, were used as negative and positive controls, respectively.
Cytotoxicity test
Cells were plated in a 96-well plate in 5 replicates and treated with ISL at 0, 2.5, 5, 10, 20 and 40 μg/mL for 24 h. The culture medium was replaced with 10 μL of MTT solution and 90 μL of DMEM, and the cells were incubated for another 4 h. After centrifugal precipitation at 1000 rpm for 5 min, the cells were incubated again in 110 μL of formazan solution for 10 minutes. Measurement of optical density (OD, 490 nm) was performed in xMarkTM (BIO-RAD, California, America).
Real-time PCR analysis
Cells were seeded in 12-well plates. After extraction of total RNA using the RNAiso Plus, cDNA was synthesized using the Reverse Transcription System (Takara, Beijing, China). Real-time PCRs were performed in triplicate using the TB Green® Premix Ex Taq™ II (Takara, Beijing, China) with the primers in Table 1. The melting curve analysis, PCR cycling parameters and conditions were documented recently [39]. The 2−ΔΔCt method was used to measure the mRNA levels of genes related to the gene of β-actin [13].
ELISA (enzyme-linked immunosorbnent assay) and endotoxin test
Cells were seeded in a 6-well plate and a 24-well plate for the detection of cytokines and endogenous endotoxin, respectively. Cell protein was extracted and quantified with Total Protein Extraction Kits and then BCA Protein Quantification Kits (Vazyme, Nanjing, China). The contents of IL-1β, TNF-α, IL-6, iNOS and COX-2 were detected by commercial ELISA kits (Youxuan, Shanghai, China) in triplicate, while endogenous endotoxin in cells was determined using Endotoxin (ET) ELISA kits (Jiangsu Jingmei Biological Technology, Yangcheng, China ). The OD values at 450 nm were measured in xMarkTM (BIO-RAD, California, America).
Western blotting analysis
Cells were cultured and treated as described above in a 6-well plate. Western blotting detection of IκBα, p-IκBα, p65, p-p65, p38, p-p38, JNK, p-JNK, ERK and p-ERK was performed as recently described [13]. Each experiment was repeated three times, and the β-actin protein was used as an endogenous control as described in a previous report in MAC-T cells [13]. Band intensity was measured using Image Lab software (version 5.2.1, Bio-Rad, California, America).
Measurement of p65 nuclear translocation
Immunofluorescence and ELISA assays for p65 nuclear translocation were analyzed as recently described [3, 13]. Briefly, MAC-T cells were cultured in a 6-well plate until 5,000 cells/well. The cells were incubated sequentially with the primary antibody against p-p65 and rabbit anti-goat IgG/Cy3 antibody. The nucleus was stained blue with 4',6-diamino-2-phenylindole (DAPI) and observed under a fluorescence inverted microscope (Leica, Wetzlar GER). For detection of p65 nuclear translocation, nuclear protein was extracted and quantified using a nuclear protein extraction kit and a BCA protein quantification kit (Vazyme, Nanjing, China), respectively. The levels of p65 in the nucleus were measured with commercial ELISA kits as described above.
Statistical Analysis
Statistical analysis was performed using SPSS 23.0. The data are expressed as the means ± standard error of the mean (SEM). The t-test was conducted between controls and LPS treatments without DEX or ISL. One-way analysis of variance, followed by Tukey’s post hoc test, was used to compare the significance among LPS combined with DEX and ISL treatments. Statistical significance was set at P < 0.05 or P < 0.01.
Availability of data and materials
The datasets generated and/or analyzed during the current study are available in the figshare repository, https://figshare.com/articles/figure/Anti-inflammation_of_isoliquiritigenin_in_LPS-stimulated_MAC-T_cells/19067642.
Abbreviations
- ISL:
-
Isoliquiritigenin
- NF-κB:
-
Nuclear factor kappa B
- TLR4:
-
Toll-like receptor 4
- MAPK:
-
Mitogen-activated protein kinase
- LPS:
-
Lipopolysaccharide
- MAC-T:
-
Bovine mammary epithelial cells
- IKK:
-
IκB kinase
- IL-1β:
-
Interleukin-1β
- TNF-α:
-
Tumor necrosis factor-α
- ERK:
-
Extracellular signal-regulated kinase
- JNK:
-
C-jun NH2 terminal kinase
- IL-6:
-
Interleukin-6
- COX-2:
-
Cyclooxygenase-2
- iNOS:
-
Inducible nitric oxide synthase
References
Klaas IC, Zadoks RN. An update on environmental mastitis: Challenging perceptions. Transbound Emerg Dis. 2018;65:166–85.
de Jong A, El Garch F, Simjee S, Moyaert H, Rose M, Youala M, et al. Monitoring of antimicrobial susceptibility of udder pathogens recovered from cases of clinical mastitis in dairy cows across Europe: VetPath results. Vet Microbiol. 2018;213:73–81.
Ma X, Li MM, Lu GC, Wang RH, Wei YM, Guo YF, et al. Anti-inflammation of epicatechin mediated by TMEM35A and TMPO in bovine mammary epithelial cell line cells and mouse mammary gland. J Dairy Sci. 2021;104(12):12925–38.
Wang JJ, Guo CM, Wei ZK, He XX, Kou JH, Zhou ES, et al. Morin suppresses inflammatory cytokine expression by downregulation of nuclear factor-kappa B and mitogen-activated protein kinase (MAPK) signaling pathways in lipopolysaccharide-stimulated primary bovine mammary epithelial cells. J Dairy Sci. 2016;99(4):3016–22.
Ryu SJ, Choi HS, Yoon KY, Lee OH, Kim KJ, Lee BY. Oleuropein suppresses LPS-induced inflammatory responses in RAW 264.7 cell and zebrafish. J Agric Food Chem. 2015;63(7):2098–105.
Oliver JP, Gooch CA, Lansing S, Schueler J, Hurst JJ, Sassoubre L, et al. Invited review: Fate of antibiotic residues, antibiotic-resistant bacteria, and antibiotic resistance genes in US dairy manure management systems. J Dairy Sci. 2020;103(2):1051–71.
Xin WX, Fang QL, Sun J, Kong SS, Chen LY, Huang P. Anticancer activity of Isoliquiritigenin: biological effects and molecular mechanisms. J Chinese Pharm Sci. 2019;28(10):673–86.
Zhang BY, Lai Y, Li YF, Shu N, Wang Z, Wang YP, et al. Antineoplastic activity of isoliquiritigenin, a chalcone compound, in androgen-independent human prostate cancer cells linked to G2/M cell cycle arrest and cell apoptosis. Eur J Pharmacol. 2018;821:57–67.
Feldman M, Santos J, Grenier D. Comparative evaluation of two structurally related flavonoids, isoliquiritigenin and liquiritigenin, for their oral infection therapeutic potential. J Nat Prod. 2011;74(9):1862–7.
Li W, Sun YN, Yan XT, Yang SY, Kim S, Lee YM, et al. Flavonoids from astragalus membranaceus and their inhibitory effects on LPS-stimulated pro-inflammatory cytokine production in bone marrow-derived dendritic cells. Arch Pharm Res. 2014;37(2):186–92.
Zou P, Ji HM, Zhao JW, Ding XM, Zhen ZG, Zhang X, et al. Protective effect of isoliquiritigenin against cerebral injury in septic mice via attenuation of NF-kappa B. Inflammopharmacology. 2019;27(4):809–16.
Wang KL, Hsia SM, Chan CJ, Chang FY, Huang CY, Bau DT, et al. Inhibitory effects of isoliquiritigenin on the migration and invasion of human breast cancer cells. Expert Opin Ther Targets. 2013;17(4):337–49.
Ma X, Wang R, Yu S, Lu G, Yu Y, Jiang C. Anti-Inflammatory activity of oligomeric proanthocyanidins via inhibition of NF-kappa B and MAPK in LPS-stimulated MAC-T cells. J. Microbiol Biotechnol. 2020;30:1458–66.
Li D, Fu Y, Zhang W, Su G, Liu B, Guo M, et al. Salidroside attenuates inflammatory responses by suppressing nuclear factor-kappa B and mitogen activated protein kinases activation in lipopolysaccharide-induced mastitis in mice. Inflamm Res. 2013;62(1):9–15.
Gunther J, Czabanska A, Bauer I, Leigh JA, Holst O, Seyfert HM. Streptococcus uberis strains isolated from the bovine mammary gland evade immune recognition by mammary epithelial cells, but not of macrophages. Vet Res. 2016;47:14.
Zhu XB, Liu JK, Chen O, Xue J, Huang SY, Zhu WW, et al. Neuroprotective and anti-inflammatory effects of isoliquiritigenin in kainic acid-induced epileptic rats via the TLR4/MYD88 signaling pathway. Inflammopharmacology. 2019;27(6):1143–53.
Ryman VE, Packiriswamy N, Sordillo LM. Role of endothelial cells in bovine mammary gland health and disease. Anim Health Res Rev. 2015;16(2):135–49.
Park SJ, Youn HS. Suppression of homodimerization of toll-like receptor 4 by isoliquiritigenin. Phytochemistry. 2010;71(14-15):1736–40.
Honda H, Nagai Y, Matsunaga T, Saitoh S, Akashi-Takamura S, Hayashi H, et al. Glycyrrhizin and isoliquiritigenin suppress the LPS sensor Toll-like receptor 4/MD-2 complex signaling in a different manner. J Leukoc Biol. 2012;91(6):967–76.
Hsia SM, Yu CC, Shih YH, Chen MY, Wang TH, Huang YT, et al. Isoliquiritigenin as a cause of DNA damage and inhibitor of ataxia-telangiectasia mutated expression leading to G2/M phase arrest and apoptosis in oral squamous cell carcinoma. Head Neck. 2016;38(Suppl 1):E360–E71.
Chi JH, Seo GS, Cheon JH, Lee SH. Isoliquiritigenin inhibits TNF-alpha-induced release of high-mobility group box 1 through activation of HDAC in human intestinal epithelial HT-29 cells. Eur J Pharmacol. 2017;796:101–9.
Jin XY, Sohn DH, Lee SH. Isoliquiritigenin suppresses tumor necrosis factor-alpha-induced inflammation via peroxisome proliferator-activated receptor-gamma in intestinal epithelial cells. Arch Pharm Res. 2016;39(10):1465–71.
Gnanaguru G, Choi AR, Amarnani D, D'Amore PA. Oxidized lipoprotein uptake through the CD36 receptor activates the NLRP3 inflammasome in human retinal pigment epithelial cells. Invest Ophthalmol Vis Sci. 2016;57(11):4704–12.
Chun J, Choi RJ, Khan S, Lee DS, Kim YC, Nam YJ, et al. Alantolactone suppresses inducible nitric oxide synthase and cyclooxygenase-2 expression by down-regulating NF-kappa B, MAPK and AP-1 via the MyD88 signaling pathway in LPS-activated RAW 264.7 cells. Int Immunopharmacol. 2012;14(4):375–83.
Rahman SU, Huang YY, Zhu L, Chu XY, Junejo SA, Zhang YF, et al. Tea polyphenols attenuate liver inflammation by modulating obesity-related genes and down-regulating COX-2 and iNOS expression in high fat-fed dogs. BMC Vet Res. 2020;16(1):234.
Lee JE, Lee EH, Park HJ, Kim YJ, Jung HY, Ahn DH, et al. Inhibition of inflammatory responses in lipopolysaccharide-induced RAW 264.7 cells by Pinus densiflora root extract. J Appl Biol Chem. 2018;61(3):275–81.
Zhang M, Wu YQ, Xie L, Teng CH, Wu FF, Xu KB, et al. Isoliquiritigenin protects against blood-brain barrier damage and inhibits the secretion of pro-inflammatory cytokines in mice after traumatic brain injury. Int Immunopharmacol. 2018;65:64–75.
Xiong D, Hu W, Ye ST, Tan YS. Isoliquiritigenin alleviated the Ang II-induced hypertensive renal injury through suppressing inflammation cytokines and oxidative stress-induced apoptosis via Nrf2 and NF-kappa B pathways. Biochem Biophys Res Commun. 2018;506(1):161–8.
Hayden MS, Ghosh S. NF-kappa B, the first quarter-century: remarkable progress and outstanding questions. Genes Dev. 2012;26(3):203–34.
Zhang HS, Sun SC. NF-kappa B in inflammation and renal diseases. Cell Biosci. 2015;5:12.
Hayden MS, Ghosh S. Shared principles in NF-kappa B signaling. Cell. 2008;132(3):344–62.
Sun SC, Liu ZG. A special issue on NF-kappa B signaling and function. Cell Res. 2011;21(1):1–2.
Zhu G, Shen Q, Jiang H, Ji O, Zhu L, Zhang L. Curcumin inhibited the growth and invasion of human monocytic leukaemia SHI-1 cells invivo by altering MAPK and MMP signalling. Pharm Biol. 2020;58(1):25–34.
Tian MY, Li K, Liu RN, Du JL ZDM, Ma YZ. Angelica polysaccharide attenuates LPS-induced inflammation response of primary dairy cow claw dermal cells via NF-κB and MAPK signaling pathways. BMC Vet Res. 2021;17:248.
Ren Q, Guo F, Tao S, Huang R, Ma L, Fu P. Flavonoid fisetin alleviates kidney inflammation and apoptosis via inhibiting Src-mediated NF-kappaB p65 and MAPK signaling pathways in septic AKI mice. Biomed Pharmacother. 2020;122:109772.
Wang JR, Luo YH, Piao XJ, Zhang Y, Feng YC, Li JQ, et al. Mechanisms underlying isoliquiritigenin-induced apoptosis and cell cycle arrest via ROS-mediated MAPK/STAT3/NF-κB pathways in human hepatocellular carcinoma cells. Drug Dev Res. 2019;80(4):461–70.
Choi YH, Bae JK, Chae HS, Choi YO, Nhoek P, Choi JS, et al. Isoliquiritigenin ameliorates dextran sulfate sodium-induced colitis through the inhibition of MAPK pathway. Int Immunopharmacol. 2016;31:223–32.
Gao Y, Lv XH, Yang HH, Peng LP, Ci XX. Isoliquiritigenin exerts antioxidative and anti-inflammatory effects via activating the KEAP-1/Nrf2 pathway and inhibiting the NF-κB and NLRP3 pathways in carrageenan-induced pleurisy. Food Funct. 2020;11(3):2522–34.
Wang RH, Su LF, Yu ST, Ma X, Jiang CD, Yu YX. Inhibition of PHLDA2 transcription by DNA methylation and YY1 in goat placenta. Gene. 2020;739:7.
Acknowledgements
Not Applicable.
Funding
This work was supported by the National Natural Science Foundation of China (32171673), People’s Livelihood Special Innovation Projects of CQ CSTC (cstc2017shms - kjfp0114) and the Innovation Team Building Program in Chongqing Universities. Neither entity had any role in the design of the study, in the collection, analysis and interpretation of the data, or in writing the manuscript.
Author information
Authors and Affiliations
Contributions
LMM, LGC, MX and WRH participated in conceptualization, methodology, investigation, data analysis, interpretation and the original draft writing. CXH assisted in methodology and investigation. JCD helped in conceptualization of the work, data analysis and manuscript proofreading. All authors have approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not Applicable.
Consent for publication
Not Applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplemental Fig. 1.
LPS-induction of IL-6 mRNA in MAC-T cells. Cells were respectively coincubated with LPS at 1, 5, 10 and 20 μg/mL for 24 h. Real-time PCR was used to measure IL-6 mRNA levels with β-actin as an internal control. Values represent the means ± SEM of four independent experiments. In the above bars, ** indicate significance at P < 0.01 between the control and the LPS treatment without DEX and ISL. Among LPS in combination with DEX and ISL treatments, the same letters indicate P > 0.05. Supplemental Fig. 2. Western blotting analysis of p-p65/p65 and p-IκB/IκB in MAC-T cells. (A) Western blotting. (B and C) p-p65/p65 and p-IκB/IκB, respectively. Cells were treated with 1 μg/mL lipopolysaccharide (LPS) in combination with dexamethasone (DEX) (20 μg/mL) or ISL (2.5, 5 and 10 μg/mL) for 24 h. The displayed gels were cropped from the original images in the additional files. In the above bars, ** indicate significance at P < 0.01 between the control and the LPS treatment without DEX and ISL. Among LPS in combination with DEX and ISL treatments, the same letters indicate P > 0.05; different lowercase letters indicate P < 0.05 while different uppercase letters indicate P < 0.01. Supplemental Fig. 3. Western blotting analysis of p-p38/p38, p-ERK/ERK and p-JNK/JNK in MAC-T cells. (A) Western blotting. (B-D) p-ERK/ERK, p-p38/p38 and p-JNK/JNK, respectively. Cells were incubated with 1 μg/mL lipopolysaccharide (LPS) in combination with dexamethasone (DEX) (20 μg/mL) or isoliquiritigenin (ISL) (2.5, 5 and 10 μg/mL) for 24 h. The displayed gels were cropped from the original images in the additional files. In the above bars, ** indicate significance at P < 0.01 between the control and the LPS treatment without DEX and ISL. Among LPS in combination with DEX and ISL treatments, the same letters indicate P > 0.05; different lowercase letters indicate P < 0.05 while different uppercase letters indicate P < 0.01.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, M., Lu, G., Ma, X. et al. Anti-inflammation of isoliquiritigenin via the inhibition of NF-κB and MAPK in LPS-stimulated MAC-T cells. BMC Vet Res 18, 320 (2022). https://doi.org/10.1186/s12917-022-03414-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12917-022-03414-1