
Abstract
Background
Colon cancer, a prominent contributor to global cancer-related deaths, prompts the need for innovative treatment strategies. Euphorbia resinifera O. Berg (E. resinifera) and Euphorbia officinarum subsp. echinus Hook. f. & Coss Vindt (E. echinus) and their bee-derived products have been integral to traditional Moroccan medicine due to their potential health benefits. These plants have historical use in addressing various health issues, including cancer. However, their effects against colon cancer remain unclear, and the specific mechanisms underlying their anti-cancer effects lack comprehensive investigation.
Methods
The study aimed to assess the potential anti-cancer effects of Euphorbia extract on colon cancer cell lines (DLD-1) through various techniques. The apoptosis, migration, and proliferation of DLD-1 cells were measured in DLD-1 cells. In addition, we conducted High-Performance Liquid Chromatography (HPLC) analysis to identify the profile of phenolic compounds present in the studied extracts.
Results
The extracts demonstrated inhibition of colon cancer cell migration. E. resinifera flower and E. echinus stem extracts show significant anti-migratory effects. Regarding anti-proliferative activity, E. resinifera flower extract hindered proliferation, whereas E. echinus flower extract exhibited dose-dependent inhibition. Apoptosis assays revealed E. resinifera flower extract inducing early-stage apoptosis and E. echinus flower extract promoting late-stage apoptosis. While apoptotic protein expression indicated, E. resinifera stem and propolis extracts had minimal impact on apoptosis.
Conclusion
The findings provide evidence supporting the beneficial effects of E resinifera and E. echinus extracts on colon cancer and exerting anti-cancer properties.
Similar content being viewed by others


Introduction
Cancer remains a significant global health challenge, accounting for many deaths [1]. Among the causes of cancer-related mortality, metastasis accounts for approximately 90%, and its underlying mechanisms are not yet fully elucidated [2]. Metastasis begins with the dissemination of the primary tumor from its originating organ or tissue to surrounding or adjacent structures, eventually reaching distant sites through systemic or lymphatic circulation [3].
On a global scale, colon cancer ranks as the third most lethal cancer affecting both men and women, making it one of the most prevalent forms of cancer [4]. Annually, over a quarter million individuals are diagnosed with colon cancer [5]. The development of various types of colon cancer can be attributed to genomic alterations that disrupt cell cycle control, apoptosis regulation, migration, and cell proliferation pathways [6]. However, one of the most significant disadvantages of current chemotherapies against colorectal cancer is the toxic side effects [7], such as nausea, vomiting, inhibition of bone marrow function, alopecia, and cell mutations, which patients dread [8]. For this reason, many scientists and healthcare providers have been interested in medicinal plants and their derivatives to prevent or treat cancer [9]. Owing to their structural stability, the chemical structure of the compounds, and secondary metabolites like flavonoids, alkaloids, terpenoids, etc., plants and their derivatives show relatively low toxicity with high potency [4, 10, 11].
The Euphorboaceae, well-known as the spurge family, is one of the most prominent flowering plants, containing about 300 genera, including almost 7,500 species [12]. They grow in the form of shrubs, trees, herbs, or lianas with irritant milky latex [13]. Several Euphorbia species have fundamental economic importance and are widely used in traditional medicine [14].
Morocco has diverse plant wealth due to its geographical location, climate, and diverse habitats, providing it with diverse natural flora [15], which plays a more prominent economic role [16]. Indeed, traditional medicine has always occupied a significant portion of conventional Moroccan medicine [17].
E. resinifera, called “Zakkoum in Arabic, is an endemic species of Morocco, found in the south of the Mid-Atlas, particularly in the Central High Atlas area, on the side of Beni Mellal and Azilal [18]. E. echinus) is an endemic species of North Africa in Morocco located in the southwestern region [19], locally known as” Daghmous” [20].
The most studied part of these plants is the latex, which has been investigated in chemical composition. Its pharmacological and biological properties have been known as antibacterial, antifungal, and antioxidant [21,22,23,24,25,26,27].
Despite the importance of these two species, the pharmacological activities are still unclear, and few studies have characterized the roles of extracts from different parts of these two plants. As part of the development and valorization of Moroccan aromatic and medicinal plants, two species of Euphorbia have been selected based on their uses in traditional medicine.
In the current study, we initially examined the anti-cancer activity of stems, flower petals, and propolis ethanolic extract of two Moroccan Euphorbia species, E. resinifera, and E. echinus, on human colon adenocarcinoma cell line, DLD-1. Our study is the first comparative study to evaluate the effect on the mechanism of anti-cancer activity of two plants belonging to the same family and their hive products.
Methods
The collection of the plant
The aerial parts of Euphorbia resinifera O.Berg (E. resinifera) and Euphorbia officinarum subsp. echinus (Hook. f. & Coss.) Vindt (E. echinus), i.e., stems and flower petals, were collected from the fields of Ouaouizeght (harvest location according to Merchich northern Morocco X: 410,804 Y: 179 306) in July and Tiznit (harvest location according to Merchich Sahara nord X: 778 847 Y: 770 997) in August, respectively, in Morocco during their flowering season in 2020 under the control of the High Commission for Water and Forests in two regions (Fig. 1). The wild plants were identified at the Scientific Institute - Department of Botany in Rabat, Morocco, the voucher reference codes for E. resinifera is RAB114129, and for E. echinus is RAB114130. The propolis was collected directly from the hives of the bee farms to avoid contamination of samples. After collection, samples were kept ambient.

Photos of wild Euphorbia plants in Morocco. (A) Euphorbia. resinifera from Ouaouizeght region. (B) Euphorbiaoflicinarum L. subsp. echinus from Tiznit region
HPLC analysis and purification
The dried flowers, stems, and propolis of the two Euphorbia species (49.6 g) were extracted using 70% ethanol with ultrasonication at room temperature. After evaporating the solvent in vacuo, the total extract (23.3 g) was subjected to RP C18-MPLC (10–100% MeOH), which yielded 11 fractions (E1−E11). Fraction E2 (387 mg) was separated by preparative HPLC using a 250 × 21.2 mm Phenomenex Luna C18 column (CH3CN−H2O 1:4), which yielded compound 2 (14.2 mg) and seven subfractions (M1−M7). Compound 6 (11 mg) was further purified from subfraction M3 (23.4 mg) by preparative HPLC with a 250 × 10 mm Phenomenex Luna C18 column (MeOH−H2O, 2:3). Subfraction M5 (75.4 mg) was isolated into five subfractions (M8−M12) by preparative HPLC using 40% MeOH in H2O. It yielded compound 1 (17.4 mg) and compound 4 (19.7 mg). Fraction E3 (319.8 mg) was subjected to RP C18-HPLC (CH3CN−H2O 3:7), which yielded eight subfractions (M13−M20). Compound 3 (4.9 mg) was obtained from subfraction M17 (9.4 mg) by preparative HPLC (MeOH: H2O = 9:11). Subfraction M19 (23.6 mg) was purified using preparative HPLC with 45% MeOH, which yielded compound 5 (20.5 mg).
Cells and culture conditions
DLD-1, human colon adenocarcinoma cells, and FHC cells were obtained from ATCC (Manassas, VA). Cells were grown in Dulbecco’s Modified Eagle’s Medium (DMEM) (Welgene, Korea) supplemented with 5% FBS and penicillin/streptomycin antibiotics. The medium was renewed every three days. Cells were maintained at 37 °C in a humidified incubator with 5% CO2.
Cell viability assay
The FHC and DLD-1 cells were placed in 96-well culture plates (5 × 103 cells/well). Once the cells reached confluence, they were treated with spurge extracts. After 16 h, seeding cells, a vehicle (DMSO), or different concentrations of extracts from flowers, propolis, or stem petals. These extracts were obtained from E. resinifera and E. echinus and were used at concentrations of 0, 1, 10, and 50 µg/ml. This treatment lasted for 24 h.
To assess the viability of the cells, the Quanti-MAX WST-8 Cell Viability Assay Kit reagent (Biomax, Korea) was added to the media. The mixture was then incubated at 37 °C for 30 min. After incubation, the absorbance of the solution was measured with a microplate reader at 450 nm.
Cell migration assay
DLD-1 cells (1.105 /well) were seeded in 24-well plates 24 h before the cell migration assay. Then, the cells were scratched with 200 µl pipette tips. The crude extracts at different concentrations (vehicle, 1, and 10 µg/ml) were treated and then incubated. The migration of cells was observed at 0, 24, and 48 h after incubation. The data were analyzed using DP2-BSW software [28]. The following formula calculated the migration rate in %.
Cell proliferation assay
Anti-proliferative effects of the different extracts of Euphorbia were measured with Carboxyfluorescein succinimidyl ester (CFSE) staining (Invitrogen, Waltham, MA).
Briefly, CFSE-stained DLD-1 cells were seeded at 1 × 105 cells per well in a 12-well plate and incubated overnight to allow adherence. Cells were incubated for 24 h with test samples at final concentrations of 1, 10, and 50 µg/ml and then trypsinized and inactivated. After washing twice with 4 °C PBS, the cell pellets were suspended in 500 µl ice-cold PBS at a density of 1 × 106 cells/ml and immediately analyzed by flow cytometry to measure the fluorescence.
Apoptosis measurement
Quantification of selective extract-induced apoptosis in DLD-1 cells was evaluated through flow cytometry using FITC Annexin V Apoptosis Detectionapoptosis kit [29] according to the manufacturer’s instructions (BD Biosciences, Franklin Lakes, NJ). DLD-1 Cells were seeded in 12-well plates at a density of 1 × 105 /well and treated with indicated extract with different concentrations at 50, 10, and 1 µg/ml for 24 h. As a positive control of apoptosis, Osimertinib was treated at 0.1 µM and 1 µM concentrations. Subsequently, cells were harvested and washed twice with 4 °C PBS (Biowest, France), and then the cell pellets were suspended in 100 mL ice-cold 1X binding buffer at a density of nearly 1 × 106 cells/ml and then incubated with 5 µl PE-Annexin V, and 5 µl 7-AAD for 10 min at room temperature in avoidance of light. Stained cells were detected and analyzed using flow cytometry. Finally, percentages of four different populations of cells are distinguished: unlabeled viable cells, early apoptotic, late apoptotic, and necrotic.
Western blotting
30 µg of cell lysate was subjected to Bis-Tris Gel (10%) and transferred to PVDF for western blot analysis. The membrane was blocked with a solution containing 5% non-fat dry milk and TBST for 1 h. Primary antibodies targeting Caspase 3, PARP, and BCL-2 were acquired from Cell Signaling Technology (Danvers, MA) and were applied at a 1:1,000 dilution in the blocking solution. Secondary antibodies were obtained from Jackson ImmunoResearch Labs (West Grove, PA) and were used at a 1:10,000 dilution. The western blot results were captured using the PXi Touch system (Syngene, Frederick, MD).
Statistical analysis
All analyses were carried out in triplicates. Data were presented as mean ± SD. A P value of less than 0.05 was considered significant. Statistical analyses were performed by two-way ANOVA (Tukey’s multiple comparisons test) to analyze the groups’ differences.
Results
Composition of Euphorbia extracts
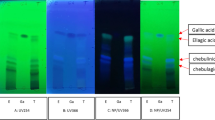
The 70% ethanol extract from the flowers, stems, and propolis of the two Euphorbia species was subjected to RP C18 Medium-pressure liquid chromatography (MPLC) and High-performance liquid chromatography (HPLC) to yield six known compounds. The compounds were elucidated using 1D and 2D NMR spectroscopy (1H, 13C, COSY, HSQC, and HMBC) and mass spectrometry. Comparison with the spectroscopic data in the literature determined the compounds as quercetin-3-O-β-D-galactospyranoside (1) [30, 31], quercetin-3-O-β-D-arabinopyranoside (2) [30, 32], quercetin-3-O-α-D-rhamnopyranoside (3) [33], helichrysin A (4), isosalipurposide (5) [34], and phenylethyl D-rutinoside (6) [35] (Fig. 2A). All compounds separated under optimal analysis conditions were analyzed by HPLC. The chromatograms are shown in Fig. 2B-G.

Structural analyses of Euphorbia plants. (A) Chemical structures of 1−6 isolated from ethanol extract of E. resinifera flowers. (B−G) HPLC chromatograms of ethanol extract from E. resinifera and E. echinus flowers. (1) quercetin-3-O-β-D-galactospyranoside; (2) quercetin-3-O-β-D-arabinopyranoside; (3) quercetin-3-O-α -D-rhamnopyranoside; (4) helichrysin A; (5) isosalipurposide; (6) phenylethyl D-rutinoside
In general, the extract of E. resinifera flowers was found to be richer in phenolic compounds than the other extract it has proven to be rich in compounds (1–6), followed by the extract of E. echinus flowers with four compounds: (1–3) and (5) The extract containing the least compounds is the extract of E. echinus stems; which showed the presence of only two compounds (3) and (5). However, the extract of the stems of E. resinifera proved to be rich in compounds (1–3). Regarding propolis extracts, the quantity was too low to identify their compounds.
Euphorbia extracts inhibit the cell migration of colon cancer cells
Cell migration is a process that plays an essential role in the progression of cancer disease [36]. In vitro-migration assay was performed to determine the potential effect of different plant extracts of the two Euphorbia species and their hive products, i.e., propolis, on DLD-1 colorectal cancer cells. The migrated distance of DLD-1 cells was measured before and 24 h after treating Euphorbia extracts at 0, 1, and 10 µg/ml (Fig. 3). After treating Euphorbia extracts, the migration of DLD-1 cells was significantly inhibited when E. resinifera flower extract (Fig. 3A, G) and E. echinus stem extract (Fig. 3E, K) were treated. This finding shows that Euphorbia extracts have an anti-migratory effect on colon cancer cells.

Euphorbia extracts inhibit the 24-hour migration of DLD-1 colon cancer cells. The migrated distance percent was measured under a microscope 24 h after the scratches on wells (scale bar = 300 μm) (A-F). Flower petals (A, D), stem (B, E), and propolis (C, F) extracts of E. resinifera (A, B, C) and E. echinus (D, E, F) were treated at 0, 1, 10 µg/ml for 24 h. The measured distance 24 h after scratches was compared to time 0 to validate the effect of Euphorbia extracts(G, E. resinifera flower petals; H, E. resinifera stems; I, E. resinifera propolis; J, E. echinus flower petals; K, E. echinus stems; L, E. echinus propolis). The measured distance was compared. Data were presented as mean + SD (n = 4). *p˂0.05, **p˂0.01, *** p˂0.001, ****p˂0.0001 and ns p˃0.05
E. resinifera flower extract showed significant inhibition of cell migration as the treated concentration was elevated to 10 µg/ml. Interestingly, the migration of DLD-1 cells was inhibited by E. resinifera stem extract at 1 µg/ml, but this inhibition was reverted when the extract was increased to 10 µg/ml. Meanwhile, propolis of E. resinifera did not show any significant change in DLD-1 migration.
In the case of E. echinus, flower and propolis extracts did not show any significant change in DLD-1 migration. However, the extract from E. echinus stem inhibited DLD-1 migration in 1 and 10 µg/ml treated groups.
Euphorbia flower extracts inhibit cell proliferation of colorectal cancer
We assessed the impact of flower extracts, which have proven to be the richest in phenolic compounds for cell proliferation of DLD-1 cells by the CFSE assay. DLD-1 cell lines were strongly affected by treatment with the flower extracts of both Euphorbia species after 24 h of treatment (Fig. 4). The flower extract of E. resinifera showed more significant inhibition of proliferation, similar for all the different concentrations tested (Fig. 4A).

Euphorbia flower extracts inhibit the proliferation of colon cancer cells. CFSE-stained DLD-1 cells were analyzed via flow cytometry after 24-hour treatment of flower petal extracts at designated concentrations of E. resinifera (A) and E. echinus (B) at 0, 1, 10, and 50 µg/ml. The signal of CFSE was measured from 1.0 × 105 events from each group (Blue, non-treated; red, treated)
When the cells were treated with E. echinus flower extracts, cell proliferation was generally demonstrated in a dose-dependent manner; a lower concentration of 1 µg/ml showed higher inhibition compared to treatment with the other concentrations (Fig. 4B).
To exclude the toxicity of Euphorbia extracts on normal colon cells, we used FHC cells and measured the proliferation by WST-8 assay. Extracts from both species showed no significant cell death in 1, 10 and 50 µg/ml (Suppl. Figure 1). However, E. resinifera propolis showed inhibition of proliferation, which might be an extreme concentration to cells (Suppl. Figure 1B). To show the effect of anti-cancer agents in DLD-1 cells, we treated 0, 0.1 and 1 µM of Osimertinib, which showed no cell death in normal FHC cells (Suppl. Figure 2 A) while DLD-1 showed significant cell death (Suppl. Figure 2B).
Euphorbia extracts induce apoptosis in colorectal cancer
Apoptosis, the programmed cell death pathway, is an essential process in various biological systems, which occurs during cell aging and development for maintaining homeostasis [37]. To determine the apoptotic effect of flower extracts, we treated the extract on DLD-1 cells for 24 h and then detached cells to propagate to Annexin V/7-AAD stain. A quantitative approach was undertaken to characterize the apoptosis induced by extracts from E. resinifera flowers, stems, and propolis, as well as E. echinus flower extract, in cell death.
During apoptosis, an early event involves the loss of plasma membrane asymmetry, leading to the translocation of phosphatidylserine (PS) to the outer leaflet of the membrane. Annexin V, a protein that binds to exposed PS, was used to assess this. By conjugating annexin V with FITC (fluorescein isothiocyanate), it becomes detectable by flow cytometry. Annexin V was combined with a vital dye called propidium iodide (PI) to distinguish apoptotic cells from dead cells. Propidium iodide is a nucleic acid intercalator that penetrates the plasma membrane of dead cells.
For the experiment, the cells were incubated with different concentrations of the extracts (0.1, 10, and 50 µg/ml) for 24 h. After the incubation, the cells were stained with annexin V-FITC/propidium iodide and then analyzed using flow cytometry (Fig. 5). After 24 h of incubation, the different concentrations of E. resinifera flower extract led to a significant decrease in cell viability. The cell viability decreased from 95.22% in the control group to 84.14% when treated with 50 µg/mL of the flower extract. Moreover, the percentage of early apoptotic cells increased from 1.83% in control to 11.12% at the same concentration of 50 µg/mL (Fig. 5). This dose-dependent increase in early-stage apoptosis suggests the potential apoptotic-inducing properties of E. resinifera flower extract.

Euphorbia extracts induce the apoptotic character of colon cancer cells. Flow cytometry analysis of Annexin V-FITC stained DLD-1 cells after 24 h exposure to 1, 10, and 50 ?g/mL of (A) E. resinifera (E.R.) flower and E. echinus (E.E.) flower and (B) E. resinifera (E.R.) stem and E. resinifera (E.R.) propolis. Non-apoptotic and non necrotic cells (Annexin V-negative; PI-negative), early apoptotic cells (Annexin V-positive; PI-negative), late apoptotic cells (Annexin V-positive; PI-positive), and necrotic cells (Annexin V-negative; PI-positive) were separately gated to analyze apoptotic events
In contrast, the flower extract of E. echinus exhibited a different pattern. Like E. resinifera, it also showed a dose-dependent increase in apoptotic cells. However, the effect was more pronounced in late-stage apoptosis. The percentage of cells in late apoptosis increased from 2.03% in control to 5.71% when treated with 50 µg/mL of E. echinus flower extract (Fig. 5B).
Additionally, we validated the effects of stem and propolis extracts of E. resinifera. When the extracts were treated to DLD-1 cells for 24 h, the apoptotic events of cells were not significantly changed with the treatment of the extracts (Fig. 5C and D).
Osimertinib was treated in DLD-1 cells as the positive control of apoptosis, and the apoptotic events were measured. At a concentration of 0.1µM Osimertinib, cell viability decreased from 73.31% (control) to 67.75%, and at 1µM, it further reduced to 45.76%. Moreover, the percentage of late apoptotic cells increased significantly from 22.49% in the control to 28.06% and 45.29% at 0.1 and 1µM Osimertinib, respectively (Suppl. Figure 3).
These findings highlight the apoptotic effects between E. resinifera and E. echinus flower extracts. While E. resinifera extract induced early apoptosis, E. echinus extract led to late apoptosis.
Mechanistic effects of Euphorbia extracts
The expression levels of numerous proteins related to the anti-cancer effect were ascertained within DLD-1 cells to elucidate potential mechanisms inherent to extracts derived from two distinct Euphorbia species.
To measure the anti-cancer effects in molecular levels, PARP, Bcl-2, Caspase-3, and Dynamin-2 were measured from the cell lysate treated with Euphorbia extracts. Propolis and stem extracts of E. echinus increase Caspase-3 expression compared to the control. Expression of Bcl-2 is also increased under the same conditions. Conversely, compared to the control, PARP expression is diminished in propolis and stem extracts. In the case of the flower extract, a notable rise in Caspase-3 expression is observed, while the levels of Bcl-2 and PARP expression do not exhibit significant changes compared to the control (Fig. 6A).

Euphorbia extracts show anti-cancer effects at molecular levels. Western blot analyses of DLD-1 cells. Cells were grown in 6-well plates overnight and then treated with E. echinus flower petals, stem, or propolis (A) and E. resinifera flower petals, stem, or propolis (B) at 0, 10 µg/ml for 24 h. The protein expression levels of PAPR, Bcl-2, Caspase-3, and Dynamin-2 were detected from the membrane. The raw data of PARP, Bcl-2 and Caspase-3 is shown in Supplementary Fig. 4
For E.resinifer, the stem extract lacks significant change in Caspase-3 expression, which indicates that this specific pathway might not have been prominently activated by the extract and stem extract could be operating in a manner independent of the classical caspase-dependent apoptotic pathway (Fig. 6B).
The reduction in PARP might reveal that cancer cells struggle to repair their DNA, possibly leading to cellular stress signals and eventual activation of apoptosis through an alternative pathway. On the other hand, the increase in Bcl-2 suggests that the extract might counter apoptosis by preserving mitochondrial membrane integrity and blocking the release of pro-apoptotic molecules.
Meanwhile, the migration marker of cancer cells, Dynamin-2, does not show significant change with the treatment of the extracts of both species. These observations match well with the subtle migration change mentioned in Fig. 3.
Discussion
Knowing the constituent molecules present in the extracts of the flowers, stems, and propolis of both species has the possible usage in therapy. This is particularly significant considering that the profile of phenolic compounds in these extracts is being examined for the first time. To achieve this, we conducted High-performance liquid chromatography (HPLC) HPLC analysis on the ethanolic extracts of all the plant parts. Selecting an appropriate extraction solvent is crucial in isolating target compounds from plant materials. In this study, 70% ethanol was chosen as the extraction solvent for the two Euphorbia species’ flowers, stems, and propolis. Selecting an appropriate extraction solvent is crucial in isolating target compounds from plant materials. In this study, 70% ethanol was chosen as the extraction solvent for the two Euphorbia species’ flowers, stems, and propolis.
In previous studies, the choice of 70% ethanol as the extraction solvent is justified by observing higher concentrations of bioactive flavonoid compounds in the ethanol extracts. This can be attributed to the increased polarity from adding water to pure ethanol. By incorporating 30% water to prepare the 70% ethanol solution, the overall polarity of the solvent was effectively improved [38, 39]. Additionally, the researchers discovered that ethanol, being less polar than water, facilitated the release of total polyphenols from cell walls due to its effectiveness in degrading the non-polar character of the cell walls. As a result, higher values of total polyphenols were observed in the ethanol extract compared to the water extract, as reported in a study by Bimakr et al. in 2010 [40]. The flower extracts of the plants exhibited a higher abundance of phenolic compounds when compared to the other extracts.
The extract obtained from E. resinifera flowers contained six compounds, including quercetin-3-O-β-D-galactospyranoside, quercetin-3-O-β-D-arabinopyranoside, quercetin-3-O-α-D-rhamnopyranoside, helichrysin A, isosalipurposide, and phenylethyl D-rutinoside. In comparison, the extract of E. echinus flowers contained four compounds, namely quercetin-3-O-β-D-galactospyranoside, quercetin-3-O-β-D-arabinopyranoside, quercetin-3-O-α-D-rhamnopyranoside, and isosalipurposide. These findings align with previous studies that have identified flavonoids, particularly quercetin glycosides, in various above-ground parts of Euphorbia species [41, 42]. The Flavonoid family has been reported to exhibit anti-proliferative activity [43] and can induce apoptosis when combined with its derivatives [44]. Several studies have investigated the effects of quercetin obtained from specialized suppliers, demonstrating its anti-proliferative and apoptosis-inducing effects on human breast cancer cells (MDA-MB-453) [45]. Additionally, research by Refolo, M.G. et al. confirmed that quercetin specifically affects the proliferation and migration of colon cancer cells (Caco2 cells) [46]. Moreover, quercetin isolated from apple peels has shown potent anti-proliferative activity against HepG2 and MCF-7 cells [47]. Furthermore, quercetin-3-O-β-D-galactopyranoside has been found to possess anti-cancer activity against skin cancer cell lines through various mechanisms, including regulating proliferation, autophagy, and apoptosis [48]. Additionally, isosalipurposide, isolated from the ethanolic extract of Helichrysum maracandicum flowers, has been reported to have anticarcinogenic activity [49]. However, no Helichrysin A and phenylethyl D-rutinoside reports exhibited any anti-cancer properties. The same compounds we detected in our extract may be responsible for the observed anti-proliferative activity in DLD-1 cells.
Euphorbia resinifera O. Berg (E. resinifera) and Euphorbia officinarum subsp. echinus (Hook. f. & Coss.) Vindt (E. echinus), two species from the Euphorbiaceae family, have a rich history of traditional use among the local Moroccan population to treat colon cancer [50].
Firstly, we determined whether the extract of Euphorbia affected both colon cancer cells and normal cells. Various techniques have been utilized when examining cell viability analyses presented in the literature. These include assays such as WST-8, MTT, Alamar Blue, cell cycle analysis, neutral red release assay, and trypan blue exclusion assay. However, it’s important to note that these diverse methods do not consistently produce similar results. [51,52,53,54]. The observations shed light on divergent cellular responses depending on the cell and extract types. Euphorbia extracts do not exhibit apparent toxicity towards DLD-1 cells and even induce enhanced cell viability under most conditions, while FHC cells are unaffected. These results concerning the DLD-1 cells can be elucidated by the fact that the WST-8 assay quantifies the conversion of the reagent into formazan, which does not directly gauge cell viability. Viable yet non-metabolically active cells may yield falsely negative outcomes [55].
In this study, we employed a range of assays to investigate and compare the mechanisms underlying the anti-cancer effects of these plants on human DLD-1 colon cancer cells in an in vitro setting. Specifically, we explored the impacts of extracts derived from various plant parts of E. resinifera and E. echinus and bee products such as propolis on cell migration, proliferation, and apoptosis. Several studies have documented the cytotoxic properties of Euphorbia latex or whole plants from specific species within the Euphorbia genus [50, 56,57,58,59,60]. However, there is a dearth of research examining the specific contributions of different parts of Euphorbia plants, specifically E. echinus and E. resinifera, in the context of cancer studies. Tumor metastases are the leading cause of cancer death. Colorectal cancer is the third most common cancer [61], and it was reported that approximately 20% of patients with colorectal cancer had been reported to present metastases at diagnosis, which can decrease the survival rate from about 56% of patients from 5 years to low survival rate [62]. The crucial roles of cell migration in cancer cell dissemination, tissue invasion, and metastasis have been extensively examined by numerous experts in the field of cancer research. Through their investigations, these researchers have provided valuable insights into the development of malignant tumors (cancers) and the underlying mechanisms involved in tumor progression [63].
In the bibliography, there is a lack of studies addressing the anti-migration activity of extracts from E. resinifera and E. echinus on cancer cells
Our results demonstrated that both species’ flower, propolis, and stem extracts tend to inhibit DLD-1 cell migration. The extracts that gave significant results after 24 h were the E. resinifera flower extract and E. echinus stem extract, which inhibited the migration of DLD-1 cells by as much as 10%. These findings suggest that these extracts possess anti-migratory properties, which can impede the progression and metastasis of colon cancer. Interestingly, the inhibitory effect of E. resinifera flower extract on cell migration became more pronounced as the concentration increased to 10 µg/ml. In contrast, the inhibition observed with E. resinifera stem extract at 1 µg/ml was diminished when the concentration was raised to 10 µg/ml. These observations indicate a potential dose-dependent effect of the extracts on cell migration. However, the stem extract of E. echinus demonstrated inhibitory effects on cell migration at both 1 µg/ml and 10 µg/ml treated concentrations. This highlights the potential anti-migratory properties associated specifically with the stem extract of E. echinus.
Later, we selected flower extracts of the two plants to analyze the anti-proliferative effect by the CFSE assay, which showed the ability to inhibit the proliferation of DLD-1 cells.
No previous data were found on the effects of E. resinifera and E. echinus extracts in human colon cancer cells, and the studies about the colon cancer effects of herbal extracts on colorectal cancers were limited. Previous reports by other researchers have documented the anti-migration of other Euphorbia species, including the hydroalcoholic extract of E. lacteal, which showed maximum inhibition (about 40%) at the concentration of 125 µg/ml against the migration of HN22 cells [64]. Another study on an ethanolic extract from seeds of E. lathyris showed a reduction in the migration capacity of 18.68% after 72 h of colon cancer cells (T84 cell line) [65]. In the same context, a study was made on propolis obtained from northern China, which showed that the ethanolic extract of this propolis showed significantly reduced cell migration at a concentration (25, 50, and 100 µg/ml) after 24 and 48 h on HepG2 hepatocellular carcinoma cells [66].
The apoptosis mechanism is one of the biomedical strategies for cancer treatment in the boundary of chemotherapy and chemoprevention [67]. In this study, Annexin V/7-AAD assay was performed to prove the mode of cell death induced by E. resinifera propolis, E. resinifera stem, E. resinifera flower, and E. echinus flower extracts, which showed significant and remarkable induction of DLD-1 apoptosis. Based on our results, Euphorbia extracts induced apoptosis in this cell line. The extract from the E. resinifera flowers shows a high ability to enhance apoptosis-mediated cell death. Therefore, the extract is considered promising as a potential anti-cancer candidate without significant toxicity to normal colon cells. So far, many plants and medicinal compounds have shown anti-cancer effects by inducing apoptosis.
After assessing the evaluation of various Euphorbia extracts for their anti-proliferative and apoptotic impacts, we unraveled the fundamental molecular mechanism responsible for initiating apoptosis. This was achieved by examining the expression of apoptosis-related proteins such as caspase 3, PARP, and Bcl-2.
The pivotal biochemical event central to the process of apoptosis was the increase in pro-apoptotic proteins and/or the decrease in molecules of anti-apoptotic proteins.
Caspase-3, a vital component in the execution of apoptosis, represents the active configuration procaspase-3. Activation of caspase-3 can arise from both the intrinsic pathway initiated by mitochondria and the extrinsic pathway activated by death receptors [68]. This, in turn, sets in motion various apoptotic processes downstream, resulting in characteristic biochemical and morphological changes within the cell. Based on the findings of this research, the increased susceptibility of DLD-1 cells to apoptosis when treated with various Euphorbia extracts notably aligns with the concurrent rise in caspase-3 activity, except for E. resinifera stem extract.
PARP, functioning as a DNA repair enzyme, becomes active in response to DNA breaks, facilitating the addition of ADP-ribose polymers to different nuclear components to accelerate repair processes. PARP holds significance for cell survival. However, the cleavage of PARP contributes to cellular breakdown and indicates cells undergoing apoptosis. In apoptosis, caspases trigger the cleavage and deactivation of PARP [69]. The decrease in PARP expression implies that DLD-1 cells treated with Euphorbia extracts could encounter disturbances in DNA repair mechanisms, thus facilitating the initiation of apoptosis. The Bcl-2 protein is an anti-apoptotic molecule, which raises its expression and causes a delay in disrupting organelles surviving cells from apoptosis [70]. These proteins are found on the outer mitochondrial membrane and play a particularly crucial role in the intrinsic apoptotic pathway [71]. The stable expression of Bcl-2 in cells treated with extracts from E. echinus flowers could suggest that this anti-apoptotic protein has not been directly affected by the ongoing anti-cancer treatment. This might imply that the cancer cells have maintained their resistance to apoptosis through a consistent Bcl-2 expression. Conversely, the increased expression of Bcl-2 in other extracts could be explained by cancer cells attempting to counteract the treatment’s effects by enhancing their resistance to apoptosis. This could also indicate an adaptive response of cancer cells to the pressure exerted by the anti-cancer treatment. Also, the observation that the expression of Dyanmin-2 remains relatively unchanged across all tested extracts compared to the control group also sparks intrigue. This suggests that the apoptotic effects induced by these extracts might occur independently of Dyanmin-2 regulation. As such, these extracts may potentially target key apoptotic pathways that exhibit limited dependence on Dyanmin-2.
In summary, this study provides an enlightening perspective on the distinct mechanisms underpinning the apoptotic effects of Euphorbia extracts. The intricate alterations in the expression of pivotal proteins hint at sophisticated interactions and the potential activation of alternative apoptotic pathways. These findings deepen our comprehension of the intricate regulatory mechanisms governing cellular fates, paving the way for promising research in anti-cancer therapy and cellular biology.
Our research focused exclusively on two species of Euphorbia, namely E. resinifera and E. echinus. As a result, generalizing the findings to other species or different types of cancer may present challenges. Conducting a broader study encompassing a wider variety of plant species and cancer types would provide a more comprehensive understanding of the anti-cancer activity of these plant extracts. Furthermore, our research primarily centered on in vitro studies. Assessing the effectiveness of plant extracts in contexts closer to reality, such as in animal models or clinical studies, would provide valuable insights and a better understanding of their therapeutic potential for patients with colon cancer.
Exploring potential interactions between Euphorbia plant extracts and commonly used anti-cancer agents is an intriguing avenue for future research. Combination studies could uncover synergistic or complementary effects, thereby enhancing the overall efficacy of treatment approaches, as many cancer therapies already employ combination strategies.
In addition, conducting pharmacokinetic studies on Euphorbia plant extracts would yield valuable information regarding their absorption, distribution, metabolism, and elimination within the body. Such data is essential for determining appropriate concentrations and dosing regimens for subsequent clinical studies. Addressing these limitations and pursuing these perspectives would contribute to a more comprehensive understanding of the anti-cancer potential of Euphorbia plant extracts and pave the way for their effective utilization in cancer treatment strategies.
Conclusions
The Euphorbia flower extract emerges as a highly promising candidate for anti-cancer therapy, as it has demonstrated, for the first time, remarkable potential in inhibiting the growth of DLD-1 colon cancer cells. Our study revealed that these flower extracts possess the capability to induce apoptosis, a vital mechanism in combating cancer. These findings strongly suggest that the flower extracts from both Euphorbia species harbor bioactive compounds that contribute to their anti-cancer activity.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
References
Stratton MR, Campbell PJ, Futreal PA. The cancer genome. Nature. 2009;458:719–24.
Chaffer CL, Weinberg RA. A perspective on cancer cell metastasis. Science. 2011;331:1559–64.
Wittekind C, Neid M. Cancer invasion and metastasis. Oncology. 2005;69(Suppl):1:14–6.
Sahoo NK, Sahu M, Pullaiah CP, Muralikrishna KS. In vitro anti-cancer activity of Eclipta alba whole plant extract on colon cancer cell HCT-116. BMC Complement Med Ther. 2020;20:1–8.
Sears CL, Garrett WS. Microbes, microbiota, and colon cancer. Cell Host Microbe. 2014;15:317–28.
Watson AJ, Collins PD. Colon cancer: a civilization disorder. Dig Dis. 2011;29:222–8.
Lichota A, Gwozdzinski K. Anti-cancer activity of natural compounds from plant and marine environment. Int J Mol Sci. 2018;19:3533.
Nguyen NH, Ta QTH, Pham QT, Luong TNH, Phung VT, Duong T-H, et al. Anti-cancer activity of novel plant extracts and compounds from Adenosma bracteosum (Bonati) in human lung and liver cancer cells. Molecules. 2020;25:2912.
Greenwell M, Rahman P. Medicinal plants: their use in anti-cancer treatment. Int J Pharm Sci Res. 2015;6:4103.
Haider F, Naqvi AA, Bagchi GD. Oil constituents of Artemisia Nilagirica var. Septentrionalis during different growth phases at subtropical conditions of north Indian plains. J Essent Oil Res. 2007;19:5–7.
Webster GL. Classification of the Euphorbiaceae. Ann Mo Bot Gard. 1994;:3–32.
Rahman A, Akter M. Taxonomy and medicinal uses of Euphorbiaceae (Spurge) family of Rajshahi, Bangladesh. Res Plant Sci. 2013;1:74–80.
Uduak AE, Kola KA. Antimicrobial activities of some Euphorbiaceae plants used in the traditional medicine of Akwa Ibom State of Nigeria. Ethnobotanical Leaflets. 2010;2010:2.
Mbayo KM, Kalonda ME, Tshisand TP, Kisimba KE, Mulamba M, Richard MK, et al. Contribution to ethnobotanical knowledge of some Euphorbiaceae used in traditional medicine in Lubumbashi and its surroundings (DRC). J Adv Bot Zool. 2016;4:1–16.
Benabid A. Flore et écosystèmes du Maroc: Evaluation et préservation de la biodiversité. 2000.
Bellakhdar J. Pharmacopée marocaine traditionnelle. Ibis; 1997.
Bouyahya A, Abrini J, Et-Touys A, Bakri Y, Dakka N. Indigenous knowledge of the use of medicinal plants in the North-West of Morocco and their biological activities. Eur J Integr Med. 2017;13:9–25.
Moujanni A, Partida L, Essamadi AK, Hernanz D, Heredia FJ, Terrab A. Physicochemical characterization of unique unifloral honey: Euphorbia resinifera. CyTA-Journal Food. 2018;16:27–35.
Moujanni A, Essamadi AK, Terrab A. L’apiculture Au Maroc: focus sur la production de miel/Beekeeping in Morocco: focus on honey production. Int J Innov Appl Stud. 2017;20:52.
Boutoub O, El-Guendouz S, Estevinho LM, Paula VB, Aazza S, El Ghadraoui L, et al. Antioxidant activity and enzyme inhibitory potential of Euphorbia resinifera and E. Officinarum honeys from Morocco and plant aqueous extracts. Environ Sci Pollut Res. 2021;28:503–17.
Wang S, Liang H, Zhao Y, Wang G, Yao H, Kasimu R, et al. New triterpenoids from the latex of Euphorbia resinifera Berg. Fitoterapia. 2016;108:33–40.
Siritapetawee J, Attarataya J, Charoenwattanasatien R. Sequence analysis and crystal structure of a glycosylated protease from Euphorbia resinifera latex for its proteolytic activity aspect. Biotechnology and Applied Biochemistry; 2021.
Li M-M, Qi Y-R, Feng Y-P, Liu W, Yuan T. Euphatexols C- G, five new triterpenoids from the latex of Euphorbia resinifera. J Asian Nat Prod Res. 2022;24:311–20.
Siritapetawee J, Khunkaewla P, Thumanu K. Roles of a protease from Euphorbia resinifera latex in human anticoagulant and antithrombotic activities. Chemico-Biol Interact. 2020;329:109223.
Smaili A, Mazoir N, Rifai LA, Koussa T, Makroum K, Benharref A, et al. Antimicrobial activity of two semisynthetic triterpene derivatives from Euphorbia officinarum latex against fungal and bacterial phytopathogens. Nat Prod Commun. 2017;12:1934578X1701200305.
Bailen M, Khamlichi MD, Benharref A, Martinez-Diaz RA, Gonzalez-Coloma A. New bioactive semisynthetic derivatives of 31-norlanostenol and obtusifoliol from Euphorbia officinarum. Nat Prod Commun. 2016;11:1934578X1601100609.
Mazoir N, Benharref A, Bailén M, Reina M, González-Coloma A, Martínez-Díaz RA. Antileishmanial and antitrypanosomal activity of triterpene derivatives from latex of two Euphorbia species. Z für Naturforschung C. 2011;66:360–6.
Liang C-C, Park AY, Guan J-L. In vitro scratch assay: a convenient and inexpensive method for analysis of cell migration in vitro. Nat Protoc. 2007;2:329–33.
Van Engeland M, Nieland LJ, Ramaekers FC, Schutte B, Reutelingsperger CP. Annexin V-affinity assay: a review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry: J Int Soc Anal Cytol. 1998;31:1–9.
Zhu Y, Liu Y, Zhan Y, Liu L, Xu Y, Xu T, et al. Preparative isolation and purification of five flavonoid glycosides and one benzophenone galloyl glycoside from Psidium guajava by high-speed counter-current chromatography (HSCCC). Molecules. 2013;18:15648–61.
Al-Yousef HM, Alqahtani AS, Ghani AS, El-Toumy SA, El-Dougdoug WI, Hassan WH, et al. Nephroprotective, cytotoxic and antioxidant activities of Euphorbia paralias. Saudi J Biol Sci. 2021;28:785–92.
Li J, Jiang H, Shi R. A new acylated quercetin glycoside from the leaves of Stevia rebaudiana Bertoni. Nat Prod Res. 2009;23:1378–83.
Fukunaga T, Nishiya K, Kajikawa I, Watanabe Y, Suzuki N, Takeya K, et al. Chemical studies on the constituents of Hyphear tanakae Hosokawa from different host trees. Chem Pharm Bull. 1988;36:1180–4.
Aydin T. Secondary metabolites of Helichrysum plicatum DC. subsp. plicatum flowers as strong carbonic anhydrase, cholinesterase and α-glycosidase inhibitors. Z für Naturforschung C. 2020;75:153–9.
Umehara K, Hattori I, Miyase T, Ueno A, Hara S. Studies on the constituents of leaves of Citrus Unshiu MARCOV. Chem Pharm Bull. 1988;36:5004–8.
Perlikos F, Harrington KJ, Syrigos KN. Key molecular mechanisms in lung cancer invasion and metastasis: a comprehensive review. Crit Rev Oncol/Hematol. 2013;87:1–11.
Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007;35:495–516.
Velavan S. Phytochemical techniques-a review. World J Sci Res. 2015;1:80–91.
Oreopoulou A, Tsimogiannis D, Oreopoulou V. Extraction of polyphenols from aromatic and medicinal plants: an overview of the methods and the effect of extraction parameters. Polyphenols Plants. 2019;:243–59.
Bimakr M, Rahman RA, Taip FS, Ganjloo A, Salleh LM, Selamat J, et al. Comparison of different extraction methods for the extraction of major bioactive flavonoid compounds from spearmint (Mentha spicata L.) leaves. Food Bioprod Process. 2011;89:67–72.
Özbilgin S, Acıkara ÖB, Akkol EK, Süntar I, Keleş H, İşcan GS. In vivo wound-healing activity of Euphorbia characias subsp. wulfenii: isolation and quantification of quercetin glycosides as bioactive compounds. J Ethnopharmacol. 2018;224:400–8.
Noori M, Chehreghani A, Kaveh M. Flavonoids of 17 species of Euphorbia (Euphorbiaceae) in Iran. Toxicol Environ Chem. 2009;91:631–41.
Kelly GS, Quercetin. Altern Med Rev. 2011;16:172–95.
Kim YJ, Jung EB, Seo SJ, Park KH, Lee MW, Lee CS. Quercetin-3-O-(2-galloyl)-α-l-rhamnopyranoside prevents TRAIL-induced apoptosis in human keratinocytes by suppressing the caspase-8-and bid-pathways and the mitochondrial pathway. Chemico-Biol Interact. 2013;204:144–52.
Choi EJ, Bae SM, Ahn WS. Anti-proliferative effects of quercetin through cell cycle arrest and apoptosis in human breast cancer MDA-MB-453 cells. Arch Pharm Res. 2008;31:1281–5.
Refolo MG, D’Alessandro R, Malerba N, Laezza C, Bifulco M, Messa C, et al. Anti proliferative and pro apoptotic effects of flavonoid quercetin are mediated by CB1 receptor in human colon cancer cell lines. J Cell Physiol. 2015;230:2973–80.
He X, Liu RH. Phytochemicals of apple peels: isolation, structure elucidation, and their anti-proliferative and antioxidant activities. J Agric Food Chem. 2008;56:9905–10.
Kong Y, Sun W, Wu P. Hyperoside exerts potentanti-cancer activity in skin cancer. Front Bioscience-Landmark. 2020;25:463–79.
Yagura T, Motomiya T, Ito M, Honda G, Iida A, Kiuchi F, et al. Anticarcinogenic compounds in the Uzbek medicinal plant, Helichrysum maracandicum. J Nat Med. 2008;62:174–8.
Mohamed A, Karima S, Nadia O. The use of medicinal plants against cancer: an ethnobotanical study in the Beni Mellal-Khenifra Region in Morocco. Eur J Integr Med. 2022;52:102137.
Stoddart MJ. WST-8 analysis of cell viability during osteogenesis of human mesenchymal stem cells. Mammalian Cell Viability: Methods Protocols. 2011;:21–5.
Jiang Z, Okimura T, Yokose T, Yamasaki Y, Yamaguchi K, Oda T. Effects of sulfated fucan, ascophyllan, from the brown Alga Ascophyllum nodosum on various cell lines: a comparative study on ascophyllan and fucoidan. J Biosci Bioeng. 2010;110:113–7.
Chen H-Y, Huang T-C, Lin L-C, Shieh T-M, Wu C-H, Wang K-L, et al. Fucoidan inhibits the proliferation of leiomyoma cells and decreases extracellular matrix-associated protein expression. Cell Physiol Biochem. 2018;49:1970–86.
Kwok AKH, Yeung CK, Lai TYY, Chan KP, Pang CP. Effects of trypan blue on cell viability and gene expression in human retinal pigment epithelial cells. Br J Ophthalmol. 2004;88:1590–4.
Aslantürk ÖS. In vitro cytotoxicity and cell viability assays: principles, advantages, and disadvantages. Genotoxicity-A predictable risk to our actual world. 2018;2:64–80.
Ahmed SR, Al-Sanea MM, Mostafa EM, Qasim S, Abelyan N, Mokhtar FA. A Network Pharmacology Analysis of cytotoxic triterpenes isolated from Euphorbia abyssinica latex supported by drug-likeness and ADMET studies. ACS Omega. 2022;7:17713–22.
El-Hawary SS, Mohammed R, Tawfike AF, Lithy NM, AbouZid SF, Amin MN, et al. Cytotoxic activity and metabolic profiling of fifteen Euphorbia Species. Metabolites. 2020;11:15.
Sumathi S, Malathy N, Dharani B, Sivaprabha J, Hamsa D, Radha P, et al. Cytotoxic studies of latex of Euphorbia antiquorum in in vitro models. J Med Plants Res. 2011;5:4715–20.
Ashraf A, Sarfraz RA, Rashid MA, Shahid M. Antioxidant, antimicrobial, antitumor, and cytotoxic activities of an important medicinal plant (Euphorbia royleana) from Pakistan. J food drug Anal. 2015;23:109–15.
Kwan YP, Saito T, Ibrahim D, Al-Hassan FMS, Ein Oon C, Chen Y, et al. Evaluation of the cytotoxicity, cell-cycle arrest, and apoptotic induction by Euphorbia hirta in MCF-7 breast cancer cells. Pharm Biol. 2016;54:1223–36.
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Cancer J Clin. 2018;68:394–424.
Tang M, Wang H, Cao Y, Zeng Z, Shan X, Wang L. Nomogram for predicting occurrence and prognosis of liver metastasis in colorectal cancer: a population-based study. Int J Colorectal Dis. 2021;36:271–82.
Vogelstein B, Kinzler KW. Cancer genes and the pathways they control. Nat Med. 2004;10:789–99.
Wongprayoon P. Cytotoxic and anti-migratory activities from hydroalcoholic extract of Euphorbia lactea haw. Against HN22 cell line. Thai Bull Pharm Sci. 2018;13:69–77.
Mesas C, Martínez R, Ortíz R, Galisteo M, López-Jurado M, Cabeza L, et al. Antitumor effect of the ethanolic extract from seeds of Euphorbia lathyris in colorectal cancer. Nutrients. 2021;13:566.
Liu H, Li J, Yuan W, Hao S, Wang M, Wang F, et al. Bioactive components and mechanisms of poplar propolis in inhibiting proliferation of human hepatocellular carcinoma HepG2 cells. Biomed Pharmacother. 2021;144:112364.
Kang K, Lee HJ, Kim CY, Lee SB, Tunsag J, Batsuren D, et al. The chemopreventive effects of Saussurea salicifolia through induction of apoptosis and phase II detoxification enzyme. Biol Pharm Bull. 2007;30:2352–9.
Elumalai P, Gunadharini DN, Senthilkumar K, Banudevi S, Arunkumar R, Benson CS, et al. Induction of apoptosis in human breast cancer cells by nimbolide through extrinsic and intrinsic pathway. Toxicol Lett. 2012;215:131–42.
Shu B, Duan W, Yao J, Huang J, Jiang Z, Zhang L. Caspase 3 is involved in the apoptosis induced by triptolide in HK-2 cells. Toxicol in Vitro. 2009;23:598–602.
Lincz LF. Deciphering the apoptotic pathway: all roads lead to death. Immunol Cell Biol. 1998;76:1–19.
Reed J, Jurgensmeier J, Matsuyama S. Bcl-2 family proteins and mitochondria. Biochim et Biophys Acta (BBA)-Bioenergetics. 1998;1366:127–37.
Acknowledgements
The authors are grateful to the Center for Research Facilities at the Chonnam National University for their assistance with the structure analysis (FT-NMR) and the Korea Basic Science Institute (KBSI) for NMR measurements and the support of the National Research Fund of South Korea (NRF-2021R1I1A3043909).
Funding
This work was supported by the grant of Research Institute of Medical Science, Daegu Catholic University (2022).
Author information
Authors and Affiliations
Contributions
RB, AZ, and SK wrote the original draft and equally contributed to this work. RB conducted experiments and collected data of Figs. 1 and 3. AZ conducted experiments and collected data of Figs. 4, 5 and 6. SK conducted experiments and collected data of Fig. 2. DK, collected and curated the data. AKE, AT and AM mentored RB. NC and JH supervised and finalized the manuscript. All authors contributed to the article and approved the submitted version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All methods comply with relevant institutional, national, and international guidelines and legislation.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Conflict of interest
The authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Benjamaa, R., Zhu, A., Kim, S. et al. Two spurge species, Euphorbia resinifera O. Berg and Euphorbia officinarum subsp. echinus (Hook.f. & Coss.) Vindt inhibit colon cancer. BMC Complement Med Ther 24, 261 (2024). https://doi.org/10.1186/s12906-024-04566-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-024-04566-3