
Abstract
Background
Antimicrobial drug resistance is a major public health threat that can render infections including wound and skin infections untreatable. The discovery of new antimicrobials is critical. Approaches to discover novel antimicrobial therapies have included investigating the antimicrobial activity of natural sources such as honey. In this study, the anti-microbial activity and chemical composition of 12 honeys from Kazakhstan and medical grade manuka honey were investigated.
Methods
Agar well diffusion and broth culture assays were used to determine anti-microbial activity against a range of skin and wound infecting micro-organisms. Folin-Ciocalteu method was used to determine the total phenol content of the honeys and non-targeted liquid chromatography analysis was performed to identify components that correlated with antimicrobial activity.
Results
In the well diffusion assay, the most susceptible micro-organisms were a clinical isolate of Methicillin resistant Staphylococcus aureus (MRSA) and Enterococcus faecalis (ATCC 19433). Buckwheat & multi-floral honey from Kazakhstan demonstrated the highest antimicrobial activity against these two micro-organisms. Kazakhstan honeys with a buckwheat floral source, and manuka honey had the highest total phenol content. Non-targeted liquid chromatography analysis identified components that correlated with anti-microbial activity as hydroxyphenyl acetic acid, p-coumaric acid, (1H)–quinolinone, and abscisic acid.
Conclusions
The Kazakhstan honeys selected in this study demonstrated antimicrobial activity against wound and skin infecting micro-organisms. Compounds identified as correlating with antimicrobial activity could be considered as potential bioactive agents for the treatment of wound and skin infections.
Similar content being viewed by others


Background
Anti-microbial drug resistance is a major worldwide health problem requiring the urgent discovery of novel therapeutic interventions [1]. The treatment of wound and skin infections with antibiotics, for example, has become less effective because of infection with antibiotic resistant strains of bacteria such as Methicillin resistant Staphylococcus aureus (MRSA) [2]. Manuka honey, produced by bees (Apis mellifera) gathering nectar from the flowering plant Leptospermum scoparium, has broad range anti-bacterial activity and is recommended for the clinical management of wound infections [3]. Common wound infecting micro-organisms include Staphylococcus aureus, Pseudomonas aeruginosa, Escherichia coli, Enterococcus faecalis and Acinetobacter baumannii [2]. In addition to wound infections, micro-organisms are associated with the etiology of a variety of skin disorders. For example, S. aureus is a cause of impetigo and furuncles and skin colonization with S. aureus is a common feature of atopic dermatitis [4]. Furthermore, Candida albicans is a cause of cutaneous candidiasis and Malassezia yeasts have been associated with seborrheic dermatitis and tinea versicolor [5]. As honey has been shown to have broad range anti-microbial activity, it is realistic to consider honey as a potential therapeutic agent for other skin disorders where micro-organisms are involved in the mechanisms of the disease.
Research has shown that components responsible for the anti-microbial activity of honey include high sugar content, low pH, hydrogen peroxide (H2O2), antimicrobial peptides, methylglyoxal (MGO) and polyphenols [6]. All honeys are unique, and the chemical composition and anti-microbial activity is often variable between different types of honey. The low moisture and high sugar content of honey exerts osmotic pressure on bacterial cells whilst low pH denatures bacterial proteins and inhibits growth. Interestingly, there is recent interest in sugar as a treatment for wounds, as well as osmotic pressure, sugar is thought to reduce amino acid breakdown in the wound leading to a reduction in wound odor [7]. Honey also contains a mixture of polyphenols derived from plant nectar, which have been described as giving each honey a polyphenol signature [8]. Polyphenols have been shown to have bactericidal effects, for example, the phenolic compound methyl syringate has been identified in manuka honey and reportedly has anti-microbial activity [9]. Polyphenols may act alone or synergistically with other polyphenols or other antimicrobial components in honey to kill micro-organisms [10]. The identification of polyphenols in honey that correlate with antimicrobial activity could help in the discovery of new antimicrobial compounds or bioactive agents of dermatological importance.
Of all the honey types, manuka has been the most extensively investigated, however, there may be others with superior antimicrobial activity that are still to be recognized. Honeys of a variety of floral sources are produced abundantly across Kazakhstan, yet the regional honeys have not been fully investigated for biological activity and potential therapeutic use. Hence, the primary aim of this study was to investigate the antimicrobial activity of selected honeys from Kazakhstan against wound and skin infecting micro-organisms and compare their activity with medical grade manuka honey. Secondly, we investigated the composition of the honeys, including pH, sugar content and total phenol content (TPC) and carried out non-targeted liquid chromatographic mass spectrometry (LC-MS) to analyze the metabolic profiles of the honeys, with the objective of identifying components responsible for the anti-microbial activity.
Methods
Microbial strains
For the well diffusion assay, the following clinical isolates obtained from the Medical Microbiology Department, Republican Diagnostic Center (RDC), Nur-Sultan, Kazakhstan were used: Methicillin resistant Staphylococcus aureus (MRSA), Acinetobacter baumannii (multi-drug resistant), Klebsiella pneumoniae (multi-drug resistant) and Candida albicans (sensitive) (Anonymous patient data showing resistance/sensitivity of the clinical isolates to a range of antibiotics or antifungals is provided in Supplementary file 1). Identification of the clinical isolates was performed using VITEK-MS: Healthcare spectrophotometer apparatus. The cefoxitin disc diffusion assay was used to confirm that the MRSA strain was methicillin resistant [11]. Other microorganisms were from the American Type Culture Collection (ATCC) and included: Pseudomonas aeruginosa (ATCC 27853), Staphylococcus aureus (ATCC 29213), Escherichia coli (ATCC 25922), Enterococcus faecalis (ATCC 19433) and Malassezia furfur (ATCC 14521). For the broth culture assay, Staphylococcus aureus (ATCC 25923), Pseudomonas aeruginosa (ATCC 27853) and Escherichia coli (ATCC 25922) were used. Results of antibiotic susceptibility testing for these three strains are shown in Tables S1a-c (Supplementary file 2).
Honey samples
The following Kazakhstan honeys (numbered 1–12) were analysed: 1. Multi-floral (East Kazakhstan*); 2. Sweet Clover (Akmola region**); 3. Sunflower (Akmola); 4. Multi-floral (East Kazakhstan); 5. Buckwheat (East Kazakhstan); 6. Buckwheat (Burabay); 7. Buckwheat & multi-floral (East Kazakhstan); 8. Buckwheat (2nd batch of honey 5); 9. Multifloral (2nd batch of honey 1); 10. Sweet clover (2nd batch of honey 2); 11. Sunflower (2nd batch of honey 3); 12. Multi-floral (2nd batch of honey 4). *Borodulikha village, towns of Zyryanovsk and Shemonaikha **Shortandy and Alexeyevka villages. All the Kazakhstan honeys were supplied by apiarists based in Nur-Sultan, Kazakhstan except for honey 6 which was purchased from a local market. The apiarists use beehive trailers to reach different regions of Kazakhstan. Activon 100% medical grade manuka honey (Advancis Medical, UK) was also tested. To determine the effects of sugar on bacterial growth, a sugar control was also produced and was based on the sugar composition of honey (consisting of 167.5 g glucose, 202.5 g fructose, 37.5 g maltose, 7.5 g sucrose dissolved in 85 mL of sterile distilled water) [12].
Determination of background microbial contamination of honey
Samples of each honey were spread onto plates containing Mueller Hinton agar (Sigma Aldrich) and incubated aerobically at 37 °C. After 24 h, plates were examined for microbial contamination.
Determination of the antimicrobial activity of honeys in vitro using a well diffusion assay
An agar well diffusion assay [13] with some modifications was adopted to determine the susceptibility of the micro-organisms to the different types of honey. Briefly, sterile glass petri dishes, 100 mm in diameter, were filled with 25 mL of nutrient agar (Himedia, India) that had been inoculated with the relevant micro-organism at a turbidity of 0.5 McFarland standard. Sabouraud dextrose agar (Himedia) was used for C. albicans and M. furfur. Using a sterile cork borer, wells of 8 mm in diameter and 3.2 mm height were cut in the agar. Samples of Kazakhstan honeys, manuka honey and sugar solutions were added to the wells in equal volumes at concentrations of 100, 75 and 50% (w/v) diluted in sterile distilled water. These concentrations were selected because of their clinical relevance (for wound healing, honey is commonly applied topically at concentrations between 90 and 100%). For the 100% honey concentrations wells were filled to the surface and for the 75 and 50% concentrations 100 μl volumes were added. Zones of inhibition were measured in mm with a clear ruler 24 h after aerobic incubation in a 37 °C incubator. All experiments were conducted in either duplicate or triplicate and repeated two, three or four times on separate occasions.
Determination of the antimicrobial activity of honeys in vitro using a broth culture assay
A broth culture assay was used to determine the antibacterial activity of the Kazakhstan honeys (7–12) and the manuka honey against S. aureus, P. aeruginosa and E. coli using the method of Schneider et al. [14]. Briefly, 75% honey broth cultures were prepared in tryptone soya broth (TSB) (Sigma Aldrich) (7.5 g of honey made up to 10 mL with TSB) then inoculated with 100 μl of an overnight culture of the micro-organism and incubated for 24 h aerobically at 37 °C at 120 rpm. Samples (1 mL) of the broths were serially diluted in 0.1 M sterile Phosphate Buffered Saline (PBS). The neat broth and serial dilutions (100 μL) were spread onto tryptone soya agar plates using a sterile spreader. Plates were placed in a 37 °C incubator for 24 h in aerobic conditions. 100% TSB control and 75% of the sugar control were also tested. Plates that had between 30 and 300 colony forming units (cfu) were counted. All experiments were conducted in triplicate and repeated at least three times on separate occasions.
Honey pH and sugar content
The pH was measured in triplicate by dipping pH test strips (GE Healthcare, UK Ltd) into each honey and reading off the pH from the manufacturer’s colour chart. The percentage total sugar content of the honeys was estimated using a hand-held refractometer (TrustTechnology Co. Ltd., Shenzhen, China) in triplicate according to the manufacturer’s instructions. Sterile water was used as a blank.
Extraction and measurement of total phenol content
Honey samples (2 × 10 g) were extracted with 20 mL of 80% (v/v) acetonitrile (ACN) containing 0.2% (v/v) formic acid (FA). Samples were vortexed for 30 s then extracted with end-over-end mixing on a blood tube rotator for 30 min at 4 °C. After centrifugation (10 min, 5o C, 2500 g), supernatants were collected. The pellets were extracted once more, and the supernatants combined. The total phenol content of the extracts was measured by the Folin method [15].
Solid Phase Extraction (SPE)
Honey extracts (~ 40 mL) were made up to 250 mL with 0.1% (v/v) FA in ultrapure water (UPW) and shaken manually to ensure dissolution. Briefly, each sample was applied to a solid phase extraction unit (Strata C18-E, GIGA SPE units, 10 g capacity; Phenomenex Ltd., UK) that had been prewashed in 0.1% (v/v) FA in ACN then equilibrated with 0.1% (v/v) FA in UPW. The unbound material, which contains organic acids and sugars, was collected. The columns were then washed with 2 volumes of UPW containing 0.1% FA. The bound extracts were eluted with 80% v/v ACN plus FA and aliquots were evaporated to dryness in a speed vacuum concentrator (Thermo Scientific, Waltham, USA). The total phenol content of bound and unbound fractions was measured as above. SPE was essential to prevent the high sugar content from obscuring the phytochemical diversity of the honeys.
Examination of phytochemical profiles by non-targeted Liquid Chromatography – Mass Spectrometry (LC-MS)
The bound SPE fractions were concentrated 10-fold using a speed vacuum concentrator then re-suspended in 250 μL of 5% (v/v) ACN containing 0.1% (v/v) FA. These were filtered using Whatman syringeless filter devices (Mini-UniPrep™) then transferred into LC-MS vials. A Thermo Dionex U3000 pump, column oven and autosampler connected to a U3000 photodiode array detector (PDAD) (Thermo Fisher Scientific UK), was used for HPLC separations. The flow rate was 300 μL/min and 20 μL of extract was injected in part-loop mode. The column and guard column (Synergi C18 Hydro-RP 80 Ä, 150 × 2.0 mm (column) and 5.0 × 2.0 mm (guard), 4 μm particle size; Phenomenex Ltd.) were maintained at 30 °C. The solvents were A, HPLC grade water, and B, HPLC grade acetonitrile, acidified with 0.1% [v/v] formic acid. The gradient program was: 0–2 min hold 2% B, 2–5 min 2–5% B, 5–25 min 5–45% B; 25–26 min 45–100% B, 26–29 min hold at 100% B, 29–30 min 100–2% B, 30–35 min hold 2% B for re-equilibration. The eluent was monitored using the PDAD in absorbance mode over 200–600 nm and three UV channel set points at 280 nm, 365 nm and 520 nm). The HPLC eluent was then transferred to a Thermo LTQ-Orbitrap XL MS operated under Xcalibur software (Thermo Fisher Scientific UK) and from 2 to 30 mins the flow was directed to the ESI-MS. Data-dependent analysis applied a primary FT full scan from 100 to 2000 m/z, followed by a secondary LTQ-IT scan to collect MS2 fragmentation data on the top 3 most intense ions [16]. The samples were analyzed in negative mode with a randomized sample order with blank extracts analyzed at the start and end of each run. Specific honey samples were also analyzed in positive mode to confirm identities.
Identification of components that correlate with antimicrobial activity
Peaks from MS chromatographs were selected using set detection criteria and processed using Xcalibur Quan software to obtain peak areas. Genstat software was used to correlate the abundance of the various components against antimicrobial activities of the honeys. Selected components that showed high correlation with antimicrobial activity were putatively identified using their MS data, exact mass and predicted formulae and MS2 properties.
Statistical analysis
For the well diffusion and broth culture assay, data was compared using a two-tailed independent Student’s t-test. P values of ≤0.05 were considered statistically significant. Correlation coefficients (R2) for total phenol content, % total sugar content and pH with antimicrobial activity were calculated using Microsoft Excel, 2010. Genstat statistical software was used to assess the correlation of the abundance of the various components against the antimicrobial activities of the honeys.
Results
Background microbial contamination of honeys
No microbial growth was detected in any of the honeys 24 h after spreading the honey on Mueller Hinton Agar.
Antimicrobial activity of Kazakhstan honeys using well diffusion assay
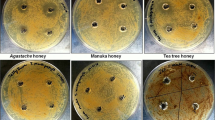
The 100% concentrations of the honeys showed the largest zones of inhibition (Fig. 1). The micro-organisms that were most susceptible to the honeys were a clinical isolate of MRSA and E. faecalis (ATCC 19443). Honey 7 (buckwheat & multi-floral), honey 8 (buckwheat) and honey 11 (sunflower) demonstrated the highest antimicrobial activity against MRSA and were significantly higher than medical grade manuka honey (p < 0.05; Fig. 2). Honey 6 (buckwheat) and honey 7 (buckwheat & multi-floral) had higher antimicrobial activity against E. faecalis than medical grade manuka honey (p < 0.05). No anti-microbial activity against C. albicans and M. furfur was detected for any honey. The sugar control demonstrated a bacteriostatic effect against MRSA, S. aureus and E. faecalis without clear zones of inhibition. For P. aeruginosa and A. baumannii, there was an inhibitory effect of the sugar control, but this was lower than the more active honeys (6.2 mm and 2.6 mm respectively). The general susceptibility of E. coli to honeys has been noted before [17] and was illustrated by the anti-microbial effect of the sugar control (mean zone of inhibition = 10.0 ± 1.4 mm). Mean zone of inhibition values were sometimes less than 8 mm in the weaker honeys and sugar controls because zones of inhibition were not always present and were thus recorded as 0.

Antimicrobial activity of (a) buckwheat & multi-floral (honey 7) and (b) medical grade manuka (Activon) honey against a clinical isolate of MRSA. Samples tested at 100, 75 and 50% (w/v)

Antimicrobial activity of 12 Kazakhstan honeys and medical grade manuka honey at 100% concentration measured using agar well diffusion assay. Results are expressed as mean zone of inhibition (mm) ± standard error of the mean. This antimicrobial data was used for the correlation analysis. * - denotes higher antimicrobial activity than medical grade manuka honey (Activon) at p < 0.05. Each honey was given a unique number (1–12). Honeys 9–12 were not tested against all bacteria
Antimicrobial activity of honeys using broth culture assay
Antibacterial activity of the 2nd batch of Kazakhstan honeys (7–12) and manuka honey against S. aureus, P. aeruginosa and E. coli was assessed using the broth culture assay (Fig. 3). Manuka honey had the highest anti-microbial activity with significant reduction in bacterial growth of all species (p < 0.05). All the Kazakhstan honeys demonstrated statistically significant anti-microbial activity against S. aureus, P. aeruginosa and E. coli when compared with the TSB control (p < 0.05). The sugar control reduced the growth of S. aureus and E. coli to 4–5 log10 cfu/ml compared to the TSB control (p < 0.05). However, the degree of inhibition caused by the sugar solutions was considerably less than the honeys (with minimum reduction to 3 log10 cfu/mL).

Anti-microbial activity of Kazakhstan honeys (7–12) and medical grade manuka honey (Activon) against S. aureus, P. aeruginosa and E. coli measured using a broth culture assay. All honeys produced significant reductions (p < 0.001)
Percentage sugar content, pH and Total Phenol Content (TPC) of the honeys
All the honeys contained a sugar content > 78%, with the highest at 84% (Table 1) and low pH (range from 2.7 to 3.8), consistent with previous reports [14]. Buckwheat honey (8) had the highest TPC followed by buckwheat honey (5), manuka honey, buckwheat & multifloral honey (7) and buckwheat honey (6), which were clearly higher than the other honeys. Interestingly, these were the same honeys that demonstrated the highest antimicrobial activity against MRSA and E. faecalis. They also had the darkest colour. However, sunflower honey (11) had good antimicrobial activity against MRSA but did not have a particularly high TPC.
Phytochemical profiling of Kazakhstan honeys by LC-MS
Each honey sample gave LC-MS profiles containing a range of components with m/z values and fragmentation (MS2) data that allowed putative identification against previous reports (Fig. 4A-D). Some abundant components were common to most, if not all, honey samples. For example, components with m/z [M-H]− values of 363, 361, 201 and 199 were present in all samples but in different amounts. These are glucosides of decanedioic and decenedioic acids and the free acids respectively, as previously identified in honeys [15]. Some components were only present in a limited number of honey samples and certain components appeared to be characteristic of certain honeys. This is particularly apparent in the manuka samples which contained components such as methyl syringate, methoxyphenyl lactic acid and the maltosyl methyl syringate derivative (m/z [M-H]− 581) unique to this monofloral honey [15, 18].

A-D. LC-MS profiles of selected Kazakhstan honey samples and medical grade manuka honey (Activon). The first panel represents UV trace at 280 nm, the second panel negative mode MS and the third panel positive mode MS profiles. BW + MF = buckwheat and multifloral honey. FSD = full scale deflection. PC = pinocembrin, C = chrysin, PB = pinobanksin, MPLA = methoxyphenyl lactic acid, PLA = phenyl lactic acid, HO-PLA = hydroxyphenyl lactic acid, pCA = p-coumaric acid
Although comparison of the LC-MS traces is informative, it was impossible to compare traces from multiple honey sources and correlate against their antimicrobial activities against the different microbes tested. Therefore, we took a non-targeted approach and quantified detectable peaks in the honey samples (Table S2) then correlated their abundance against antimicrobial activities. All honey samples were tested against MRSA, E. faecalis, A. baumannii and P. aeruginosa whereas a reduced set of honey samples were tested against K. pneumonia, E. coli and S. aureus. Interestingly, when these two sets of correlations were performed, components suggested to be antimicrobial were consistent across the two correlations (see supplementary material Table S3). Components that correlated with antimicrobial activity (Table 2) and certain components that were negatively correlated with antimicrobial activity, i.e. which appeared to promote bacterial growth, were identified (Table 3).
Discussion
In the well diffusion assay, the two Gram-positive micro-organisms, MRSA and E. faecalis were the most susceptible to the honeys tested. The Kazakhstan buckwheat & multifloral (7) honey displayed the highest antimicrobial activity against these two micro-organisms. It is clinically relevant that this honey is effective against MRSA because it is essential that new anti-microbials are found against antibiotic-resistant strains of this bacteria. Buckwheat & multifloral honey (7) and also buckwheat honey (8) from Kazakhstan could therefore be considered as a potential treatment for not only wound infections caused by S. aureus but also other disorders of the skin associated with S. aureus infection such as impetigo or impetiginized atopic dermatitis. The growth of C. albicans was not affected by any of the honeys tested in this study and this finding is consistent with reports in the scientific literature that C. albicans is more resistant to honey than many other microbial species e.g. [19]. The honeys tested in this study also did not exhibit anti-fungal activity against M. furfur, therefore it is unlikely that they would be efficacious for the treatment of skin diseases caused by C. albicans or M. furfur.
All honeys tested were significantly bactericidal against S. aureus, P. aeruginosa and E. coli as assessed by the broth culture assay. Indeed, using this assay, manuka honey was more effective than the buckwheat & multifloral honey (7), which was more effective than manuka in the well diffusion assay. This disparity may be related to the ease by which the different chemical components responsible for the anti-bacterial activities of the honeys can diffuse through the nutrient agar in the well diffusion assay. It has previously been suggested that the anti-microbial components of honey can move better in liquid broth than in agar [6, 14]. However, the use of different bacterial strains in the different assays may also be a factor.
Comparing the chemical composition of the honeys against their anti-microbial activity may indicate components responsible for the anti-microbial activity. Higher total sugar content was not associated with increased antimicrobial activity as correlation coefficients (R2) for % total sugar content and antibacterial activity against MRSA and E. faecalis, for example, were − 0.54 and − 0.35 respectively. The pH of the honeys ranged from 2.7 to 3.8 and did not correlate with antibacterial activity against MRSA and E. faecalis (R2 = 0.10 and 0.17 respectively; well diffusion data). The most effective honeys i.e. manuka honey, buckwheat honey (8) and buckwheat & multifloral honey (7) had the highest TPC and R2 values for TPC and activity against MRSA and E. faecalis were 0.42 (p = 0.14) and 0.40 (p = 0.18), respectively. This agrees with Kaskoniene et al., [20] and Fyfe et al. [15] who showed that the more effective honeys had higher polyphenol levels. However, like our findings, Fyfe et al., [15] did not find a statistically significant correlation between TPC and antimicrobial activity. Although TPC may be significant, the actual polyphenol composition of honeys may be more important, and this may explain why sunflower honey (11) was not particularly high in TPC but demonstrated good antimicrobial activity against MRSA. We did not measure the hydrogen peroxide content of the honeys, however Fyfe et al. [15] reported that hydrogen peroxide content of Scottish honeys did not differ between the effective and non-effective honeys.
Interestingly, correlation analysis also revealed that honeys that were effective against MRSA were also effective against P. aeruginosa (R2 = 0.85) and honeys that were effective against E. faecalis were also effective against K. pneumoniae (R2 = 0.86). There was a lower but still significant correlation (R2 = 0.55) between effectiveness against P. aeruginosa and A. baumannii. These correlations were not related to the bacteria being Gram-positive or Gram-negative but are likely related to the specific mechanism of action of the antibacterial components present in the honeys and the bacterial molecules they act upon. Further research is required to determine what these mechanisms are.
The correlation of composition with antimicrobial activity indicated candidate components including hydroxyphenyl acetic acid, p-coumaric acid, (1H)-quinolinone, and abscisic acid (Table 2). Hydroxyphenyl acetic acid has previously been identified in other types of honey and has been reported to have antibacterial activity against P. aeruginosa [21, 22]. Huberman et al., [22] suggested that the antibacterial activity of phenols was related to the production of free radicals which can exert an inhibitory effect on microbial proteins. Antibacterial activity of p-coumaric acid has been reported against S. aureus and E. coli [23] and was suggested to act by increasing bacterial membrane permeability, binding to DNA, and altering gene expression and bacterial replication. p-Coumaric acid has been reported to be more abundant in buckwheat honey than in manuka honey [24]. 1H-quinolinones have previously been found in honey [25] and have been shown to have antibacterial activity with their mechanism of action related to inhibition of bacterial DNA gyrase [26]. Abscisic acid (ABA) is a plant hormone which has been found in honey [15] and ABA isolated from Korean acacia honey was reported to have antibacterial activity against Helicobacter pylori [27].
On the other hand, compounds identified as potentially supporting the growth of bacteria were also identified. Unedone has previously been identified in honeys [28] and may be derived from ABA. Isorhamnetin-diglucoside is a flavonol derivative and has also been detected in other types of honey [9, 28]. There are no previous reports on the antibacterial effectiveness of these two compounds. Phloretin, a dihydrochalcone, has been shown to have antibacterial activity against Propionibacterium acnes, S. aureus, E. coli and P. aeruginosa. The antibacterial activity of phloretin was reportedly strain specific and in P. acnes may arise through binding to β-ketoacyl acyl carrier protein (ACP) synthase III (KAS III) and interference with fatty acid synthesis [29], or in S. aureus, it may interfere with fatty acid and sugar utilization [30]. Phloretin from natural sources has been shown to be bound to other compounds [31] and this may alter its antibacterial activity. The presence of Tri-p-coumaroyl spermidine has been noted in honeys [32] but its antimicrobial potential has not been reported. Fatty acids in honey such as hydroxy hexadecanedioic acid, dihydroxy palmitic acid [17] and the undefined difatty acid derivative may provide fuel for bacterial growth or provide protection from other antibacterial agents in the honeys [33].
A limitation of this study is that we did not compare the antimicrobial activity of the honeys with appropriate antibiotics serving as positive controls. This could have provided important information about the effectiveness of the honeys in comparison to antibiotics.
In the future, it would be important to investigate the anti-microbial properties of the honeys against micro-organisms associated with other skin disorders such as P. acnes, determine the minimal inhibitory concentration and hydrogen peroxide content of the honeys, and investigate the antimicrobial properties and mechanisms of the components identified as correlating with antimicrobial activity.
Conclusions
The Kazakhstan honeys tested in this study have antimicrobial activity against wound and skin infecting microorganisms. The antimicrobial compounds identified in this study could be considered as bioactive agents for potential therapeutic use in the treatment of wound or skin infections and should be investigated further.
Availability of data and materials
Some of the data generated or analysed during the study are included in this publication in supplementary information files. Other data analysed are available on reasonable request from the corresponding author.
Abbreviations
- ABA:
-
Abscisic acid
- ACN:
-
Acetonitrile
- ACP:
-
Acyl carrier protein
- ATCC:
-
American type culture collection
- BW + MF:
-
Buckwheat & multifloral
- C:
-
Chrysin
- C. albicans :
-
Candida albicans
- E. coli :
-
Escherichia coli
- ESI-MS:
-
Electrospray ionisation mass spectrometry
- FA:
-
Formic acid
- FSD:
-
Full scale deflection
- FT:
-
Fourier transform
- HO-PLA:
-
Hydroxyphenyl lactic acid
- HPLC:
-
High performance liquid chromatography
- KAS III:
-
β-ketoacyl acyl carrier protein (ACP) synthase III
- LC-MS:
-
Liquid chromatographic mass spectrometry
- M. furfur :
-
Malassezia furfur
- MGO:
-
Methylglyoxal
- MPLA:
-
Methoxyphenyl lactic acid
- MRSA:
-
Methicillin resistant Staphylococcus aureus
- P. aeruginosa :
-
Pseudomonas aeruginosa
- PB:
-
Pinobanksin
- PBS:
-
Phosphate buffered saline
- PC:
-
Pinocembrin
- PDAD:
-
Photodiode array detector
- PLA:
-
Phenyl lactic acid
- S. aureus :
-
Staphylococcus aureus
- SPE:
-
Solid phase extraction
- TPC:
-
Total phenol content
- TSB:
-
Tryptone soya broth
- UK:
-
Unknown
- UPW:
-
Ultrapure water
- UV:
-
Ultraviolet
References
World Health Organisation. Antimicrobial resistance: global report on surveillance. 2014. https://www.who.int/drugresistance/documents/surveillancereport/en/ [Accessed 8 Oct 2019].
Collier M. Recognition and management of wound infections. World Wide Wounds. United Kingdom: Surgical Materials Testing Laboratory; 2004. http://www.worldwidewounds.com/2004/january/Collier/Management-of-Wound-infections.html. Accessed 7 Oct 2019.
NHS, Lothian, Scotland. NHS Lothian Joint Formularies. Wound Section. https://www.ljf.scot.nhs.uk/LothianJointFormularies/Adult/Wound%20Section/Pages/default.aspx [Accessed, 8 Oct 2019].
Ong PY. Recurrent MRSA skin infections in atopic dermatitis. J Allergy Clin Immunol Pract. 2014;2(4):396–9. https://doi.org/10.1016/j.jaip.2014.04.007.
Gaitanis G, Magiatis P, Hantschke M, Bassukas ID, Velegraki A. The Malassezia genus in skin and systemic disease. Clin Microbiol Rev. 2012;25(1):106–41. https://doi.org/10.1128/CMR.00021-11.
McLoone P, Warnock M, Fyfe L. Honey: a realistic antimicrobial for disorders of the skin. J Microbiol Immunol Infect. 2016;49(2):161–7. https://doi.org/10.1016/j.jmii.2015.01.009.
Wu DJ, Figenshau K, Rajpara A, Aires DJ, Elston D. Sugar based dressings for wound care. J Am Acad Dermatol. 2020;82(3):e73. https://doi.org/10.1016/j.jaad.2019.09.049.
Ceksteryte V, Klazlauskas S, Racys J. Composition of flavonoids in Lithuanian honey and beebread. Biologija. 2006;2:28–33.
Oelschlaegel S, Gruner M, Wang P-N, Boettcher A, Koelling-Speer I, Speer K. Classification and characterization of Manuka honeys based on phenolic compounds and methylglyoxal. J Agric Food Chem. 2012;60(29):7229–37. https://doi.org/10.1021/jf300888q.
Alvarez M, DeBattista N, Pappano N. Synergism of flavonoids with bacteriostatic action against Staphylococcus aureus ATCC 25923 and Escherichia coli ATCC 25922. Biocell. 2006;30(1):39–42.
Centers for Disease Control and Prevention, Methicillin resistant Staphylococcus aureus (MRSA), Laboratory testing. Laboratory Testing | MRSA | CDC. Accessed on the 29 May 2021.
Muller P, Alber DG, Turnbull L, Schlothauer RC, Carter DA, Whitchurch CB, et al. Synergism between Medihoney and rifampicin against methicillin-resistant Staphylococcus aureus (MRSA). PLoS One. 2013;8(2):e57679.
Dryden M, Lockyer G, Saeed K, Cooke J. Engineered honey: in vitro antimicrobial activity of a novel topical wound care treatment. J Glob Antimicrob Resist. 2014;2(3):168–72.
Schneider M, Coyle S, Warnock M, Gow I, Fyfe L. Anti-microbial activity and composition of Manuka and Portobello honey. Phytother Res. 2013;27(8):1162–8. https://doi.org/10.1002/ptr.4844.
Fyfe L, Okoro P, Paterson E, Coyle S, McDougall GJ. Compositional analysis of Scottish honeys with antimicrobial activity against antibiotic-resistant bacteria reveals novel antimicrobial components. LWT Food Sci Technol. 2017;79:52–9.
Allwood JW, Woznicki TL, Xu Y, Foito A, Aaby K, Sungurtas J, et al. Application of HPLC-PDA-MS metabolite profiling to investigate the effect of growth temperature and day length on blackcurrant fruit. Metabolomics. 2019;15(1):12. https://doi.org/10.1007/s11306-018-1462-5.
Leyva-Jimenez FJ, Lozano-Sanchez J, Borras-Linares I, de la Luz C-GM, Mahmoodi-Khaledi E. Potential antimicrobial activity of honey phenolic compounds against gram-positive and gram-negative bacteria. LWT Food Sci Technol. 2019;101:236–45.
Fearnley L, Greenwood DR, Schmitz M, Stephens JM. Compositional analysis of manuka honeys by high resolution mass spectrometry: identification of a manuka-enriched archetypal molecule. Food Chem. 2011;132(2):948–53.
Kuncic MK, Jaklic D, Lapanje A, Gunde-Cimerman N. Anti-bacterial and anti-mycotic activities of Slovenian honeys. Br J Biomed Sci. 2012;69(4):154–8.
Kaskoniene V, Maruska A, Kornysova O, Charczun N, Ligor M, Buszewski B. Quantitative and qualitative determination of phenolic compounds in honey. Chem Tech. 2009;3:74–80.
Vasic V, Gasic U, Stankovic D, Lusic D, Vukic-Lusic D, Milojkovic-Opsenica D, et al. Towards better quality criteria of European honeydew honey: phenolic profile and antioxidant capacity. Food Chem. 2019;274:629–41. https://doi.org/10.1016/j.foodchem.2018.09.045.
Huberman L, Gallop N, Mumcuoglu K, Breuer E, Bhusare SR, Shai Y, et al. Antibacterial substances of low molecular weight isolated from the blowfly Lucilia sericata. Med Vet Entomol. 2007;21(2):127–31. https://doi.org/10.1111/j.1365-2915.2007.00668.x.
Lou Z, Wang H, Rao S, Sun J, Ma C, Li J. P-Coumaric acid kills bacteria through dual damage mechanisms. Food Control. 2012;25:550–4.
Deng J, Liu R, Lu Q, Hao P, Xu A, Zhang J, et al. Biochemical properties, antibacterial and cellular antioxidant activities of buckwheat honeys in comparison to manuka honey. Food Chem. 2018;252:243–9. https://doi.org/10.1016/j.foodchem.2018.01.115.
Makowicz E, Jasicka-Misiak I, Teper D, Kafarski P. HPTLC fingerprinting-rapid method for the differentiation of honeys of different botanical origin based on the lipophilic fractions. Molecules. 2018;23(7):1811. https://doi.org/10.3390/molecules23071811.
Hradil P, Hlavac J, Soural M, Hajduch M, Kolar M, Vecerova R. 3-Hydroxy-2-phenyl-4 (1H)-quinolinones as promising biologically active compounds. Mini-Rev Med Chem. 2009;9(6):696–702. https://doi.org/10.2174/138955709788452720.
Kim S, Hong I, Woo S, Jang H, Pak S, Han S. Isolation of abscisic acid from Korean acacia honey and anti-helicobacter activity. Pharmacogn Mag. 2017;13(Suppl 2):S170–3. https://doi.org/10.4103/0973-1296.210166.
Tuberoso CIG, Bifulco E, Caboni P, Cottiglia F, Cabras P, Floris I. Floral markers of strawberry tree (Arbutus unedo L.) honey. J Agric Food Chem. 2010;58(1):384–9. https://doi.org/10.1021/jf9024147.
Cheon D, Kim J, Jeon D, Shin HC, Kim Y. Target protein of phloretin for its anti-inflammatory and anti-bacterial activities against Propionibacterium acnes. Molecules. 2019;24(9):1319. https://doi.org/10.3390/molecules24071319.
Barreca D, Bellocco E, Lagana G, Ginestra G, Bisignano C. Biochemical and antimicrobial activity of phloretin and its glycosylated derivatives present in apple and kumquat. Food Chem. 2014;160:292–7. https://doi.org/10.1016/j.foodchem.2014.03.118.
Cao X, Wang C, Pei H, Sun B. Separation and identification of polyphenols in apple pomace by high speed counter-current chromatography and high-performance liquid chromatography coupled with mass spectrometry. J Chromatogr A. 2009;1216(19):4268–74. https://doi.org/10.1016/j.chroma.2009.01.046.
Negri G, Barreto LMRC, Sper FL, de Carvalho C, Campos MGR. Phytochemical analysis and botanical origin of Apis mellifera bee pollen from the municipality of Canavieriras Bahia State, Brazil. Braz J Food Technol. 2018;21:e2016176. https://doi.org/10.1590/1981-6723.17616.
Saito HE, Harp JR, Fozo EM. Incorporation of exogenous fatty acids protects enterococcus faecalis from membrane damaging agents. Appl Environ Microbiol. 2014;80(20):6527–38. https://doi.org/10.1128/AEM.02044-14.
Acknowledgements
We would like to thank Dr. Dinara Almabayeva of Nazarbayev University for helping us source high quality Kazakhstan honeys for the experiments described in this article.
Funding
This research was funded by the Ministry of Education and Science, Republic of Kazakhstan. IRN No. АР05135293. The funders had no role in the study design, data collection, data analyses or interpretation or in the writing of the paper.
Author information
Authors and Affiliations
Contributions
PM and GM designed the study, acquired funding, analyzed the data, wrote the original draft and critically revised and reviewed the manuscript. AZ, YK and SY performed experimental procedures and data analysis. LY was involved in funding acquisition, provision of research materials, antibiogram analysis, supervision and intellectual contributions. SV, JS CA and JWA were involved in data curation, experimental procedures, formal analysis and validation. All authors critically reviewed the manuscript and agreed to the published version of the manuscript. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 2: Table S1.
a. E. coli ATCC 25922 antibiotic susceptibility testing (antibiogram). b. P. aeruginosa ATCC 27853 antibiotic susceptibility testing (antibiogram). c. S. aureus ATCC 25923 antibiotic susceptibility testing (antibiogram). Table S2. Peak areas for all detected peaks in honey samples. Table S3. Correlation of peak abundance with antimicrobial activity against different bacteria.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
McLoone, P., Zhumbayeva, A., Yunussova, S. et al. Identification of components in Kazakhstan honeys that correlate with antimicrobial activity against wound and skin infecting microorganisms. BMC Complement Med Ther 21, 300 (2021). https://doi.org/10.1186/s12906-021-03466-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-021-03466-0