
Abstract
Background
Integrating systems biology is an approach for investigating metabolic diseases in humans. However, few studies use this approach to investigate the mechanism by which Rhizoma coptidis (RC) reduces the effect of lipids and glucose on high-fat induced obesity in rats.
Methods
Twenty-four specific pathogen-free (SPF) male Sprague–Dawley rats (80 ± 10 g) were used in this study. Serum metabolomics were detected by ultra-high-performance liquid chromatography coupled with quadrupole-time-of-flight tandem mass spectrometry. Liver tissue and cecum feces were used for RNA-Seq technology and 16S rRNA gene sequencing, respectively.
Results
We identified nine potential biomarkers, which are differential metabolites in the Control, Model and RC groups, including linoleic acid, eicosapentaenoic acid, arachidonic acid, stearic acid, and L-Alloisoleucine (p < 0.01). The liver tissue gene expression profile indicated the circadian rhythm pathway was significantly affected by RC (Q ≤ 0.05). A total of 149 and 39 operational taxonomic units (OTUs), which were highly associated with biochemical indicators and potential biomarkers in the cecum samples (FDR ≤ 0.05), respectively, were identified.
Conclusion
This work provides information to better understand the mechanism of the effect of RC intervention on hyperlipidemia and hypoglycemic effects in obese rats. The present study demonstrates that integrating systems biology may be a powerful tool to reveal the complexity of metabolic diseases in rats intervened by traditional Chinese medicine.
Similar content being viewed by others

Background
A Western diet (high fat) is increasingly popular in China [1], resulting in obesity and diabetes [2], cardiovascular diseases and atherosclerosis. However, the mechanisms underlying this diet-induced obesity remain unclear. Human symbiotic gut microbiota has recently been linked to obesity-related diseases [3,4,5]. A growing body of studies have suggested that gut microbiota and the metabolites of intestinal microbiota play an important role in the development of obesity-associated diseases [6,7,8,9,10]. Therefore, how to regulate the gut microbiota of obese individuals by diet [11, 12], drugs, or probiotics and prebiotics [13], attracts scientific attention.
With nearly 2000 years of clinical use, Rhizoma coptidis (RC) is still used today in traditional Chinese medicine (TCM). RC is widely used for treatment of obesity and its complications [14].
RC possesses common clinical therapeutic effects, including antihyperglycemia and antihyperlipidemia [14, 15]. The main pharmacological active ingredients in RC include berberine, epiberberine, coptisine and palmatine, all of which are alkaloids. Modern pharmacological studies show that the alkaloids from RC can reduce glucose, lipids, and inflammatory cytokines levels [16,17,18,19]. Jiang et al. (2004) showed that berberine is a novel, cholesterol-lowering drug that is different from statins [20]. Because RC alkaloids are mainly distributed in the liver and intestine of rats [21], the modulation of gut microbiota and its influence on lipid metabolic pathways are very important aspects of the effects of RC alkaloids on glucose and lipids. A study of B6 mice found that alkaloids could alleviate hyperlipidemia by modulating gut microbiota and bile acid pathways [22]. Additionally, the berberine of RC recently demonstrated effects on preventing high-fat-diet-induced obesity and insulin resistance by altering gut microbiota [23]. The above findings highlight the critical role of RC alkaloids in hyperlipidemia treatment in an obesity model. In addition, the clinical application of alkaloids was found to be more suitable for the obesity stage of prediabetes [24, 25]. Despite these advances, we know very little about the mechanism by which hyperlipidemia and hypoglycemic effects are affected by RC intervention in an obesity model.
Integrating systems biology reveals an efficient method for investigating the etiology of complex metabolic diseases, particularly the application of metabolomics in gut microbiota research [26]. The bacteria producing short-chain fatty acids (SCFAs) are regulated by berberine in Wistar rats [27].
In the present study, we performed an untargeted metabolomics analysis, RNA sequencing and 16S rRNA gene sequencing of gut bacteria to elucidate the mechanism by which RC mitigates the lipids and glucose effect on high-fat-induced obesity in rats.
Methods
Materials and reagents
We used HPLC-grade acetonitrile and methanol purchased from Merck (Germany). HPLC-grade formic acid was obtained from Dikma (USA). According to the determination method recorded in the China pharmacopoeia (2020 edition), we prepared the RC decoction and performed quality control (Supplementary Fig. 1 in Supplementary Information). Detailed information for RC preparation is provided in the RC decoction standards within the Supplementary Materials and Methods section in the Supplementary Information. The QIAamp Stool DNA kits (Germany) was used to extract the total DNA of the gut microbiota from cecum collected from the rats killed. The 16S rRNA gene V4 region amplification, sequencing, and bioinformatics analysis are described in detail in the Supplementary Materials and Methods section in the Supplementary Information. The total RNA in the liver was determined using the TRIzol method, details of the statistical analysis are provided in the Supplementary Materials and Methods section in the Supplementary Information.
Animal experiments
All experiments involving rats were performed with protocols approved by the Animal Ethics Committee of Jiangxi University of TCM. We purchased 24 specific pathogen-free (SPF) male Sprague–Dawley rats (80 ± 10 g) from the Hunan Slac Jynda Laboratory Animal Company (Hunan, China). After 1 week of adaptive feeding by free diet and drinking in a barrier system in the laboratory animal science and technology center of Jiangxi University of TCM, all 24 rats were randomly divided into two groups: (1) Control group (n = 8), raised with normal chow diet (containing 10% fat by energy; Supplementary Table 1 in Supplementary Information); (2) Model group (n = 16), raised with high-fat diet (containing 60% fat by energy; Supplementary Table 1 in Supplementary Information). After 8 weeks of breeding, the Model group rats were randomly separated into two groups: (1) Model group (n = 8), continually fed with high-fat diet; and (2) Model + RC group (n = 8), continuously fed with high-fat diet and additional stomach filling with the RC raw medical materials by 0.05 g/kg body weight once daily. Four weeks later (after drug intervention), all animals were euthanized by cervical dislocation after fasting but with free access to water overnight. The liver and feces were excised and collected, weighed, and frozen in liquid nitrogen immediately for further analysis. Weight and length were measured every week, and blood was collected every 2 weeks throughout the experiment. All animals were raised in the same environment and treated in the same way.
Metabolomic analysis
All blood samples were centrifuged at 4 °C (1500 rpm, 15 min) to isolate serum. In order to obtain small substances with a molecular weight less than 1000 Da, we employed conventional methanol precipitation protein methods [28]. The water ACQUITY UPLC™ I-Class Xevo G2-XS QTOF system (USA) was used to detect the metabolic profile in ESI positive-ion modes. The chromatographic conditions were carried out on Waters BEH-C18 columns (2.1 × 50 mm, 1.7 μm) to separate the serum blood samples. Optimized UPLC and MS settings for analysis in the only ESI + mode is shown in Supplementary Table 2 in the Supplementary Information.
Data processing and statistical analyses
Metabolomics data were acquired from the Masslynx software (USA) and the raw data were analyzed by Progenesis QI software (Version 2.3, UK). First, peak picking was aligned and retention time (RT) was calibrated. Then, all the data were imported into the EZinfo software (Version 3.0, Swit) for multidimensional statistical analysis after experimental design setup, including principal component analysis (PCA) and partial least-squares-discriminant analysis (PLS-DA). The quality control parameters for m/z satisfied the minimum coefficient of variation (CV) < 30%, ANOVA p value < 0.05, max fold change ≥ 2.0 and VIP > 1 and were chosen for potential biomarkers. The potential biomarkers were further identified by METLIN, KEGG, and HMDB databases as well as via Ingenuity Pathway Analysis (IPA) software [29]. Finally, the differences between the verified potential biomarkers between the Model and Model + RC groups were performed by one-way analysis of variance using the software program R (http://www.r-project.org/).
Results
RC reduces glucose in high-fat diet-induced obese rats
After 8 weeks of high-fat diet breeding, we found the weight, length, and lee’s index (LI) of the Model group (obesity) to be higher than that of the Control group raised on a normal chow diet (Supplementary Table 3 in Supplementary Information). In addition, the levels of total cholesterol (TC), glucose (GLU) and homeostasis model assessment of insulin resistance (HOMA-IR) of the Model group were also higher than those of the Control group (Supplementary Table 3 in Supplementary Information). However, after 4 weeks of oral administration of RC, only GLU and HOMA-IR were significantly reduced when compared with the Model group (Supplementary Table 4 in Supplementary Information). Meanwhile, the level of TC was still higher than that of the Control group. In addition, we analyzed the amount of food consumed after ingestion of the RC decoction. Compared with the Model group, there was no difference in the average amount of food consumed from the RC group (p > 0.05; Supplementary Table 5 in Supplementary Information).
Serum metabolic profiles and candidate biomarkers
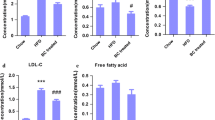
The total ion chromatograms (TIC) and basic peak intensity (BPI) of serum samples in the positive-ion mode are shown in Supplementary Fig. 2 in Supplementary Information. UHPLC-Q-TOF–MS in the positive MSE model and unadjusted continued data was used to detect small endogenous metabolites in the serum. The PLS-DA results of the Control, Model, and RC groups are shown in Fig. 1. There is a slight separation between the Control and Model groups. Furthermore, potential biomarkers were identified by Progenesis MetaScope of the Progenesis QI software based on accurate mass measurements via UHPLC-TOF–MS. The ion m/z 556.2771 of leucine encephalin was used to illustrate the process of biomarker identification. The software adopted an automatic search method with retention time correction and peak alignment. The detailed validation protocol was performed according to Zhang’s methods [28] (Supplementary Fig. 3 in Supplementary Information). The process of searching for potential biomarkers and pathways was as follows: (1) The differential metabolites between the Control and Model groups were defined as palmitic acid, linoleic acid, eicosapentaenoic acid, arachidonic acid and stearic acid, and the associated pathways were from the biosynthesis of unsaturated fatty acids, free fatty acid receptors, α-linolenic acid, linoleic acid metabolism, and the circadian clock (Supplementary Table 6 in Supplementary Information). (2) According to the comparison between the Model and RC groups, in addition to the differential metabolites identified between the Control and Model, branched chain amino acids (norvaline, L-alloisoleucine), betaine and 5-aminopentanoic acid were also found to be differential metabolites between the Model and RC groups (Supplementary Table 7 in Supplementary Information). (3) To further compare the differences among the three groups, we used the Bartlett test of homogeneity of variances in combination with a one-way ANOVA to estimate the p-values, as shown in Fig. 2. Compared to the Control group, the levels of linoleic acid and eicosapentaenoic acid were significantly decreased (p < 0.01) in the serum samples of the obese rats raised with a high-fat diet. Although the RC group did not show a reversal of this trend, the levels of eicosapentaenoic acid tended to rise.

PLS-DA score plots derived from the UHPLC-Q-TOF spectra of the serum in positive mode

Comparison of the abundance of potential biomarkers in serum from the Control, Model and RC groups. The potential biomarkers are palmitic acid, linoleic acid, eicosapentaenoic acid, arachidonic acid, stearic acid, L-alloisoleucine, norvaline, betaine, and 5-aminopentanoic acid. *p < 0.05, **p < 0.01, #p < 0.05, ##p < 0.01, * and # compared between the Control group and Model group, respectively
Differential gene expression of liver tissue
To further explore the changes of related genes in serum metabolites caused by glucose-lowering as well as lipid invariability affected by RC, we carried out RNA-Seq technology of the liver tissue (Supplementary Materials and Methods in Supplementary Information). Since most drugs are eliminated from the body by hepatic metabolism, we sequenced the total RNA in the liver tissue [30, 31]. The PCA scoring diagram helped to differentiate among the three groups. Compared with the Model group and the Control group, the RC group profile displayed an apparent returning trend (Fig. 3). Hence, the identified differentially expressed genes could contribute to the hypoglycemic effect mechanism of RC in high-fat-diet-induced obese rats.

PCA scores plots derived from the transcriptional profiling of the liver tissue
In order to understand the relationship between the differentially expressed genes, we constructed network relationships [32]. Interestingly, the circadian rhythm pathway was especially affected by the RC, in which priority was given to the Arntl gene (Fig. 4 and Supplementary Fig. 4 in Supplementary Information). Compared with the Control group, the expression levels of all seven candidate genes in the Model group were significantly decreased. However, compared to the Model group, the expression levels of these genes in the RC group were significantly increased and were close to, or equal to, the levels in the Control group.

The network of the Arntl gene of the circadian rhythm pathway
Differentiated microbiota with biochemical indicators and metabolomic biomarkers
Further studies showed that gut microbes were associated with obesity and produced large amounts of metabolites [33, 34]. To further verify the relationship between serum metabolites and biorhythm genes under the hypoglycemic effect of RC, we executed 16S rRNA gene sequencing of gut bacteria of cecum. It has been reported that cecum luminal samples are more useful for investigation of fatness-associated microbes than stool samples [35]. Hence, we used the cecum sample to study the gut microbiota. We used the association studies with a two-part model to screen out the biomarkers and biochemical indicators related operational taxonomic units (OTUs) (Supplementary Materials and Methods in Supplementary Information) [36]. Corrections were first made for the body weight and length values, and then the residuals were used for association. In the biochemical indicator results, we identified a total of 149 significant associations for 88 shared OTUs at FDR ≤ 0.05 for TC and high-density lipoprotein cholesterol (HDL-C) (Supplementary Fig. 5 in Supplementary Information). These OTUs were mainly annotated to Clostridium viride, Butyricicoccus pullicaecorum, Lachnospiracea and Ruminococcaceae.
With respect to the potential biomarkers, we identified 39 OTUs that were significantly associated with linoleic acid, eicosapentaenoic acid and arachidonic acid at FDR ≤ 0.05 (Fig. 5). These OTUs were annotated to the three dominant Phyla (Firmicutes, Bacteroidetes and Proteobacteria). Of the 39 metabolite-associated OTUs, two (Otu982 and Otu256) were shared among the three metabolites. These two OTUs were annotated to Lachnoclostridium and Ruminococcus, respectively. Lachnoclostridium showed a negative association with both linoleic acid and arachidonic acid (P = 1.00E – 04 and 3.77E – 05, respectively) and a positive association with eicosapentaenoic acid (P = 3.91E – 05). Ruminococcus was positively associated with linoleic acid, arachidonic acid and eicosapentaenoic acid (P = 1.00E – 04, 3.77E – 05 and 2.58E – 05, respectively) (Fig. 5). The OTU376 associated with TC, HDL-C, linoleic acid and arachidonic acid was classified as Clostridium scindens. Clostridium scindens showed strong negative associations with the biochemical indicators and positive associations with potential biomarkers. As for other biochemical indicators and potential biomarkers, we did not identify any significant associations at the OTU level.

The relationship between the endogenous metabolites and gut microbiota. A The results of linoleic acid. B The results of eicosapentaenoic acid. C The results of arachidonic acid. Thirty-nine OTUs were significantly associated with linoleic acid, eicosapentaenoic acid and arachidonic acid at FDR ≤ 0.05. Otu982 and Otu256 were shared among the three metabolites
Discussion
The present study aimed to elucidate the mechanism by which RC mitigates the glucose effect on high-fat-induced obesity in rats. The results of the serum biochemical indicators showed that RC could effectively lower blood glucose, which is consistent with clinical observations [37]; however, RC did not reduce the serum lipids. We noticed that the observed results were independent of the amount of food ingested. The reason for this result may be the alkaloids, which are the main components that have inhibitory activities of α-glucosidase in RC extract [38, 39].
We also evaluated the serum metabolic profiles and found that the differential metabolites were associated with the metabolites of unsaturated fatty acids and SCFAs. Linoleic acid can reduce triglyceride storage [40] and lead to a reduction in total serum cholesterol [41], diminishing glucose uptake and utilization [40]. Arachidonic acid, as a metabolite of linoleic acid, can regulate cholesterol metabolism [42]. Eicosapentaenoic acid, which belongs to a different family of polyunsaturated fatty acids, can also alleviate and/or prevent obesity [43]. Eicosapentaenoic acid inhibited hyperglycemia through a potent antioxidant mechanism [44]. These preliminary studies showed that linoleic acid and arachidonic acid both have a good effect on lipid regulation in obesity, while eicosapentaenoic acid has a good effect on hypoglycemia. This is consistent with the results observed after administration of the RC in this study. Only the level of eicosapentaenoic acid increased in the RC group. Results of these potential metabolites support the data regarding the serum biochemical indicators. After RC treatment, the level of betaine increased compared to the Model and Control groups. Additionally, a recent study suggested that supplementation of a native betaine source increased propionic acid production [45]. In addition, these differential metabolites were produced by intestinal microbiota and confirmed the effect of RC on gut microbiota [45,46,47].
We found that the difference of gene enrichment was mostly caused by sugar and lipid metabolism related genes and their pathways, except for the Dbp gene. However, one study showed that the Dbp gene binds to saturated and unsaturated fatty acids [48]. This suggested that the RC could have a great influence on the starting position of the loop in the biorhythm pathway [49]. The above results verified that blood glucose and HOMA-IR were affected by serum metabolite enrichment of the biological clock pathway. Circadian rhythm changes could have a profound effect on human health, and up to 15% of human genes have been regulated by patterns of circadian rhythm. Nearly 50% of genes involved in metabolism pathways found in the liver were under the influence of this rhythm [50].
Gut microbes play an important role in obesity. Interestingly, we found the OTUs that were associated with TC and HDL-C produced SCFAs such as Clostridium viride, Butyricicoccus pullicaecorum [51], Lachnospiracea and Ruminococcaceae [52, 53]. Regarding the differential metabolites we screened, as well as the results related to OTU, we found that these OTUs, which mainly were annotated as Lachnospiracea and Ruminococcaceae [54], were also SCFAs producing bacteria. These bacteria helped increase fecal SCFAs concentrations, promote energy intake from fiber, inhibit opportunistic pathogens, and protect the hosts against inflammation and colonic diseases [55].
At the OTU level, the biochemical indicators and differential metabolites associated OTUs were annotated as Clostridium scindens, which converted glucocorticoids into androgens by cleaving the carbon–carbon bond of 17-hydroxylated corticoids at C17-C20 [56]. According to studies of humans and rats [57], Clostridium scindens is involved in the synthesis of bile acid and may inhibit the growth of Clostridium difficile.
A recent study by De Preter (2015) described systems biology as an integrative research strategy that studies the interactions between DNA, mRNA, protein, and metabolite level in an organism [58]. Based on the integrating systems biology research strategy, Mardinoglu et al. (2018) used a multi-omics approach to characterize the resulting alterations in metabolism, transcript profiling of liver biopsies, and the gut microbiota [59]. In future studies, integrating systems biology would greatly promote glycolipids mechanism research on the complexity of metabolic diseases intervened by traditional Chinese medicine. Furthermore, the results from this study provide important insight, such as isolation of the causative microbes for RC, and mitigation of the glucose effect on high-fat-induced obesity in rats, that provide basic information for regulating the gut microbes to reduce the occurrence of obesity.
Conclusion
Collectively, by systematically combining untargeted metabolomics, RNA-Seq sequencing and 16S rRNA gene analysis techniques, which proved to be an efficient and robust way of identifying potential biomarkers from many metabolites, nine potential biomarkers were found that showed significant associations with the circadian rhythm pathway. Furthermore, a major gene (Arntl gene) was affected by RC intervention. The potential biomarkers and biochemical indicators associated with gut microbiota were mainly affected by the SCFAs producing bacteria. These results suggested that the circadian rhythm pathway may play an important role in the metabolites of serum from obese rats as well as the gut microbiota composition. We established a simple research strategy for integrating systems biology and provided information to better understand the mechanism of the effect of RC intervention on the hypoglycemic effect in obese rats.
Availability of data and materials
Serum metabolism, fecal microbiota, liver tissue expression profiles and correlation analysis data are available from the corresponding author on reasonable request.
Abbreviations
- TCM:
-
Traditional Chinese medicine
- RC:
-
Rhizoma coptidis
- SPF:
-
Specific pathogen-free
- OTUs:
-
Operational taxonomic units
- FDR:
-
False discovery rate
- RT:
-
Retention time
- PCA:
-
Principal component analysis
- PLS-DA:
-
Partial least-squares-discriminant analysis
- CV:
-
Coefficient of variation
- IPA:
-
Ingenuity Pathway Analysis
- LI:
-
Lee’s index
- TC:
-
Total cholesterol
- GLU:
-
Glucose
- HOMA-IR:
-
Homeostasis model assessment of insulin resistance
- TG:
-
Triglycerides
- HDL-C:
-
High-density lipoprotein cholesterol
- LDL-C:
-
Low-density lipoprotein cholesterol
- TIC:
-
Total ion chromatograms
- BPI:
-
Basic peak intensity
- SCFAs:
-
Short-chain fatty acids
- HPLC–UV:
-
Liquid chromatography with ultraviolet detector
- RDP:
-
Ribosomal Database Project
References
Rosa-Caldwell ME, Lee DE, Brown JL, Brown LA, Perry RA Jr, Greene ES, Carvallo Chaigneau FR, Washington TA, Greene NP. Moderate physical activity promotes basal hepatic autophagy in diet-induced obese mice. Appl Physiol Nutr Metab. 2017;42(2):148–56.
Liu Y, Su D, Zhang L, Wei S, Liu K, Peng M, Li H, Song Y. Endogenous L-carnosine level in diabetes rat cardiac muscle. Evid Based Complement Alternat Med. 2016;2016:6230825.
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027–31.
Waldram A, Holmes E, Wang Y, Rantalainen M, Wilson ID, Tuohy KM, McCartney AL, Gibson GR, Nicholson JK. Top-down systems biology modeling of host metabotype-microbiome associations in obese rodents. J Proteome Res. 2009;8(5):2361–75.
Zhao L, Zhang F, Ding X, Wu G, Lam YY, Wang X, Fu H, Xue X, Lu C, Ma J, et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science. 2018;359(6380):1151–6.
De Vadder F, Kovatcheva-Datchary P, Goncalves D, Vinera J, Zitoun C, Duchampt A, Backhed F, Mithieux G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell. 2014;156(1–2):84–96.
den Besten G, Bleeker A, Gerding A, van Eunen K, Havinga R, van Dijk TH, Oosterveer MH, Jonker JW, Groen AK, Reijngoud DJ, et al. Short-chain fatty acids protect against high-fat diet-induced obesity via a PPARγ-dependent switch from lipogenesis to fat oxidation. Diabetes. 2015;64(7):2398–408.
De Vadder F, Kovatcheva-Datchary P, Zitoun C, Duchampt A, Backhed F, Mithieux G. Microbiota-produced succinate improves glucose homeostasis via intestinal gluconeogenesis. Cell Metab. 2016;24(1):151–7.
Mollica MP, Mattace Raso G, Cavaliere G, Trinchese G, De Filippo C, Aceto S, Prisco M, Pirozzi C, Di Guida F, Lama A, et al. Butyrate regulates liver mitochondrial function, efficiency, and dynamics in insulin-resistant obese mice. Diabetes. 2017;66(5):1405–18.
Serena C, Ceperuelo-Mallafre V, Keiran N, Queipo-Ortuno MI, Bernal R, Gomez-Huelgas R, Urpi-Sarda M, Sabater M, Perez-Brocal V, Andres-Lacueva C, et al. Elevated circulating levels of succinate in human obesity are linked to specific gut microbiota. ISME J. 2018;12(7):1642–57.
Wu G, Zhang C, Wang J, Zhang F, Wang R, Shen J, Wang L, Pang X, Zhang X, Zhao L, et al. Diminution of the gut resistome after a gut microbiota-targeted dietary intervention in obese children. Sci Rep. 2016;6:24030.
Gu Y, Liu C, Zheng N, Jia W, Zhang W, Li H. Metabolic and gut microbial characterization of obesity-prone mice under a high-fat diet. J Proteome Res. 2019;18(4):1703–14.
Barengolts E. Gut microbiota, prebiotics, probiotics, and synbiotics in management of obesity and prediabetes: review of randomized controlled trials. Endocr Pract. 2016;22(10):1224–34.
Li WL, Zheng HC, Bukuru J, De Kimpe N. Natural medicines used in the traditional Chinese medical system for therapy of diabetes mellitus. J Ethnopharmacol. 2004;92(1):1–21.
Yuan L, Tu D, Ye X, Wu J. Hypoglycemic and hypocholesterolemic effects of Coptis chinensis franch inflorescence. Plant Foods Hum Nutr. 2006;61(3):139–44.
Tang J, Feng Y, Tsao S, Wang N, Curtain R, Wang Y. Berberine and Coptidis rhizoma as novel antineoplastic agents: a review of traditional use and biomedical investigations. J Ethnopharmacol. 2009;126(1):5–17.
Li JY, Wang XB, Luo JG, Kong LY. Seasonal variation of alkaloid contents and anti-inflammatory activity of Rhizoma coptidis based on fingerprints combined with chemometrics methods. J Chromatogr Sci. 2015;53(7):1131–9.
Xie W, Gu D, Li J, Cui K, Zhang Y. Effects and action mechanisms of berberine and Rhizoma coptidis on gut microbes and obesity in high-fat diet-fed C57BL/6J mice. PLoS One. 2011;6(9):e24520.
He K, Kou S, Zou Z, Hu Y, Feng M, Han B, Li X, Ye X. Hypolipidemic effects of alkaloids from Rhizoma Coptidis in diet-induced hyperlipidemic hamsters. Planta Med. 2016;82(8):690–7.
Kong W, Wei J, Abidi P, Lin M, Inaba S, Li C, Wang Y, Wang Z, Si S, Pan H, et al. Berberine is a novel cholesterol-lowering drug working through a unique mechanism distinct from statins. Nat Med. 2004;10(12):1344–51.
Wang L, Ye X, Li X, Chen Z, Chen X, Gao Y, Zhao Z, Huang W, Chen X, Yi J. Metabolism, transformation and distribution of Coptis chinensis total alkaloids in rat. Zhongguo Zhong Yao Za Zhi. 2010;35(15):2017–20.
He K, Hu Y, Ma H, Zou Z, Xiao Y, Yang Y, Feng M, Li X, Ye X. Rhizoma Coptidis alkaloids alleviate hyperlipidemia in B6 mice by modulating gut microbiota and bile acid pathways. Biochim Biophys Acta. 2016;1862(9):1696–709.
Zhang X, Zhao Y, Xu J, Xue Z, Zhang M, Pang X, Zhang X, Zhao L. Modulation of gut microbiota by berberine and metformin during the treatment of high-fat diet-induced obesity in rats. Sci Rep. 2015;5:14405.
Formisano E, Pasta A, Cremonini AL, Favari E, Ronca A, Carbone F, Semino T, Di Pierro F, Sukkar SG, Pisciotta L. Efficacy of nutraceutical combination of monacolin K, berberine, and silymarin on lipid profile and PCSK9 plasma level in a cohort of hypercholesterolemic patients. J Med Food. 2020;23(6):658–66.
Yin J, Xing H, Ye J. Efficacy of berberine in patients with type 2 diabetes mellitus. Metabolism. 2008;57(5):712–7.
Liu KY, Song YG, Liu YL, Peng M, Li HY, Li XL, Feng BW, Xu PF, Su D. An integrated strategy using UPLC-QTOF-MSE and UPLC-QTOF-MRM (enhanced target) for pharmacokinetics study of wine processed Schisandra Chinensis fructus in rats. J Pharmaceut Biomed. 2017;139:165–78.
Zhang X, Zhao Y, Zhang M, Pang X, Xu J, Kang C, Li M, Zhang C, Zhang Z, Zhang Y, et al. Structural changes of gut microbiota during berberine-mediated prevention of obesity and insulin resistance in high-fat diet-fed rats. PLoS One. 2012;7(8):e42529.
Zhang Q, Xu G, Li J, Guo X, Wang H, Li B, Tu J, Zhang H. Metabonomic study on the plasma of streptozotocin-induced diabetic rats treated with Ge Gen Qin Lian Decoction by ultra high performance liquid chromatography-mass spectrometry. J Pharm Biomed Anal. 2016;120:175–80.
Zeng ZJ, Ji YH, Huang X, Jiang YN, Ou ZM, Xiong WP, Li BT, Zhang QY, Nie P, Xu GL, et al. Integrating the Metpa and Ipa to verify the biological function of the potential biomarkers from plasma metabonomics in diabetic rats. Acta Med Mediterr. 2019;35(3):1627–31.
Shi XB, Yang SM, Zhang G, Song YG, Su D, Liu YL, Guo F, Shan LN, Cai JQ. The different metabolism of morusin in various species and its potent inhibition against UDP-glucuronosyltransferase (UGT) and cytochrome p450 (CYP450) enzymes. Xenobiotica. 2016;46(5):467–76.
Shi XB, Zhang G, Ge GB, Guo Z, Song YG, Su D, Shan LN. In vitro metabolism of auriculasin and its inhibitory effects on human cytochrome P450 and UDP-glucuronosyltransferase enzymes. Chem Res Toxicol. 2019;32(10):2125–34.
Zeng ZJ, Gao Y, Liu LY, Yan XJ, Xu GL, Liu HN, Ji YH. Quickly evaluating the synergistic effects of top anti-cancer drugs by the computer high performance computing power and complex network visualization. J Intell Fuzzy Syst. 2020;38(1):277–81.
Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444(7122):1022–3.
Munoz-Garach A, Diaz-Perdigones C, Tinahones FJ. Gut microbiota and type 2 diabetes mellitus. Endocrinol Nutr. 2016;63(10):560–8.
He MZ, Fang SM, Huang XC, Zhao YZ, Ke SL, Yang H, Li ZJ, Gao J, Chen CY, Huang LS. Evaluating the contribution of gut microbiota to the variation of porcine fatness with the cecum and fecal samples. Front Microbiol. 2016;7:2108.
Fu J, Bonder MJ, Cenit MC, Tigchelaar EF, Maatman A, Dekens JA, Brandsma E, Marczynska J, Imhann F, Weersma RK, et al. The gut microbiome contributes to a substantial proportion of the variation in blood lipids. Circ Res. 2015;117(9):817–24.
Pang B, Yu XT, Zhou Q, Zhao TY, Wang H, Gu CJ, Tong XL. Effect of Rhizoma coptidis (Huang Lian) on treating diabetes mellitus. Evid Based Complement Alternat Med. 2015;2015:921416.
Ge AH, Bai Y, Li J, Liu J, He J, Liu EW, Zhang P, Zhang BL, Gao XM, Chang YX. An activity-integrated strategy involving ultra-high-performance liquid chromatography/quadrupole-time-of-flight mass spectrometry and fraction collector for rapid screening and characterization of the alpha-glucosidase inhibitors in Coptis chinensis Franch. (Huanglian). J Pharm Biomed Anal. 2014;100:79–87.
Chang YX, Ge AH, Donnapee S, Li J, Bai Y, Liu J, He J, Yang X, Song LJ, Zhang BL, et al. The multi-targets integrated fingerprinting for screening anti-diabetic compounds from a Chinese medicine Jinqi Jiangtang Tablet. J Ethnopharmacol. 2015;164:210–22.
den Hartigh LJ, Han CY, Wang S, Omer M, Chait A. 10E,12Z-conjugated linoleic acid impairs adipocyte triglyceride storage by enhancing fatty acid oxidation, lipolysis, and mitochondrial reactive oxygen species. J Lipid Res. 2013;54(11):2964–78.
Jackson RL, Taunton OD, Morrisett JD, Gotto AM Jr. The role of dietary polyunsaturated fat in lowering blood cholesterol in man. Circ Res. 1978;42(4):447–53.
Demetz E, Schroll A, Auer K, Heim C, Patsch JR, Eller P, Theurl M, Theurl I, Theurl M, Seifert M, et al. The arachidonic acid metabolome serves as a conserved regulator of cholesterol metabolism. Cell Metab. 2014;20(5):787–98.
Li JJ, Huang CJ, Xie D. Anti-obesity effects of conjugated linoleic acid, docosahexaenoic acid, and eicosapentaenoic acid. Mol Nutr Food Res. 2008;52(6):631–45.
Mason RP, Jacob RF. Eicosapentaenoic acid inhibits glucose-induced membrane cholesterol crystalline domain formation through a potent antioxidant mechanism. Biochim Biophys Acta. 2015;1848(2):502–9.
Rink F, Bauer E, Eklund M, Mosenthin R. The effect of betaine on in vitro fermentation of carbohydrate and protein combinations under osmotic stress in pigs. J Sci Food Agric. 2012;92(12):2486–93.
McIntosh FM, Shingfield KJ, Devillard E, Russell WR, Wallace RJ. Mechanism of conjugated linoleic acid and vaccenic acid formation in human faecal suspensions and pure cultures of intestinal bacteria. Microbiology. 2009;155(Pt 1):285–94.
Piazzi G, D’Argenio G, Prossomariti A, Lembo V, Mazzone G, Candela M, Biagi E, Brigidi P, Vitaglione P, Fogliano V, et al. Eicosapentaenoic acid free fatty acid prevents and suppresses colonic neoplasia in colitis-associated colorectal cancer acting on Notch signaling and gut microbiota. Int J Cancer. 2014;135(9):2004–13.
Swamy N, Ray R. Fatty acid-binding site environments of serum vitamin D-binding protein and albumin are different. Bioorg Chem. 2008;36(3):165–8.
Dardente H, Fortier EE, Martineau V, Cermakian N. Cryptochromes impair phosphorylation of transcriptional activators in the clock: a general mechanism for circadian repression. Biochem J. 2007;402(3):525–36.
Masri S, Rigor P, Cervantes M, Ceglia N, Sebastian C, Xiao C, Roqueta-Rivera M, Deng C, Osborne TF, Mostoslavsky R, et al. Partitioning circadian transcription by SIRT6 leads to segregated control of cellular metabolism. Cell. 2014;158(3):659–72.
Eeckhaut V, Wang J, Van Parys A, Haesebrouck F, Joossens M, Falony G, Raes J, Ducatelle R, Van Immerseel F. The probiotic Butyricicoccus pullicaecorum reduces feed conversion and protects from potentially harmful intestinal microorganisms and necrotic enteritis in broilers. Front Microbiol. 2016;7:1416.
Lawley TD, Clare S, Walker AW, Stares MD, Connor TR, Raisen C, Goulding D, Rad R, Schreiber F, Brandt C, et al. Targeted restoration of the intestinal microbiota with a simple, defined bacteriotherapy resolves relapsing Clostridium difficile disease in mice. PLoS Pathog. 2012;8(10):e1002995.
Schubert AM, Rogers MA, Ring C, Mogle J, Petrosino JP, Young VB, Aronoff DM, Schloss PD. Microbiome data distinguish patients with Clostridium difficile infection and non-C. difficile-associated diarrhea from healthy controls. MBio. 2014;5(3):e01021-01014.
Zhang J, Guo Z, Xue Z, Sun Z, Zhang M, Wang L, Wang G, Wang F, Xu J, Cao H, et al. A phylo-functional core of gut microbiota in healthy young Chinese cohorts across lifestyles, geography and ethnicities. ISME J. 2015;9(9):1979–90.
De Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, Collini S, Pieraccini G, Lionetti P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci U S A. 2010;107(33):14691–6.
Ridlon JM, Ikegawa S, Alves JM, Zhou B, Kobayashi A, Iida T, Mitamura K, Tanabe G, Serrano M, De Guzman A, et al. Clostridium scindens: a human gut microbe with a high potential to convert glucocorticoids into androgens. J Lipid Res. 2013;54(9):2437–49.
Greathouse KL, Harris CC, Bultman SJ. Dysfunctional families: Clostridium scindens and secondary bile acids inhibit the growth of Clostridium difficile. Cell Metab. 2015;21(1):9–10.
De Preter V. Metabonomics and systems biology. Methods Mol Biol. 2015;1277:245–55.
Mardinoglu A, Wu H, Bjornson E, Zhang C, Hakkarainen A, Rasanen SM, Lee S, Mancina RM, Bergentall M, Pietilainen KH, et al. An integrated understanding of the rapid metabolic benefits of a carbohydrate-restricted diet on hepatic steatosis in humans. Cell Metab. 2018;27(3):559-571.e555.
Acknowledgements
We thank the Jiangxi Province Key Laboratory of TCM Etiopathogenesis, University of Jiangxi TCM for providing the analytical platform.
Funding
This research was supported by the National Natural Science Foundation of China (81760787, 81560744 and 61762051), China Postdoctoral Science Foundation (2017M622101), Scientific Foundation of the Education Department of Jiangxi Province (GJJ190643), Traditional Chinese medicine Scientific Research Subject of Jiangxi Province (2019A111, 2019B110), Health and Family Planning Commission of Jiangxi province (20195651, 20203766 and 20203773), Technology Innovation Guidance Program of Jiangxi Province (National Science and Technology Award Backup Program Cultivation Program, 20192AEI91002), Research Center of Natural Resources of Chinese Medicinal Materials and Ethnic Medicine (JXXT2018002) and the specialized research fund for the first-class discipline of Chinese herb from Jiangxi province (JXSYLXK-ZHYAO111).
Author information
Authors and Affiliations
Contributions
All authors contributed to this study. YJ and ZZ performed the experiments, analyzed the data, and wrote the manuscript; KL, JZ, PN, WX, and XL collected the samples; HL conceived and designed the experiments; and GX revised the manuscript. All authors have read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was approved by the Animal Ethics Committee of Jiangxi University of TCM (SYXK2017-0004).
Consent for publication
Not applicable.
Competing interests
The authors report no conflict of interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Supplementary Materials and Methods. Supplementary Table 1. The diet composition on normal chow diet and high-fat diet. Supplementary Table 2. UPLC elution gradient for optimized UPLC-MS methods, ESI+. A is 99.9/0.1 water/formic acid and B is 99.9/0.1 acetonitrile/formic acid. Supplementary Table 3. Pre-administration: the of results the apparent indexes and serum biochemical indexes (\(\overline{x} \pm s\)). Supplementary Table 4. After dosing: the of results the apparent indexes and serum biochemical indexes (\(\overline{x} \pm s\)). Supplementary Table 5. After dosing: the of results amount of food ingested. Supplementary Table 6. The differences metabolites and association pathways of the Control and Model groups. Supplementary Table 7. The differences metabolites and association pathways of the RC and Model groups. Supplementary Figure 1. HPLC fingerprint of the RC decoction. (A) HPLC-UV chromatogram of Batch A. (B) HPLC-UV chromatogram of Batch B. Peaks were detected at 345nm (1, coptisine; 2, palmatine; 3, berberine hydrochloride). Supplementary Figure 2. The chromatograms of the serum sample of the rats in positive mode. (A) Typical TIC chromatogram obtained from the same serum sample of the rats with positive mode. (B) Typical BPI chromatogram obtained from the same serum sample of the rats with positive mode. Supplementary Figure 3. Identification of a potential marker. MS/MS spectrum; the collision energy was 20eV~30eV. Supplementary Figure 4. The pathway enrichment of differentially expressed genes of the liver tissue. Supplementary Figure 5. The relationship between the biochemical indicators and gut microbiota. (A) The results of TC. (B) The results of HDL-C.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ji, Y., Luo, K., Zhang, J.M. et al. Obese rats intervened with Rhizoma coptidis revealed differential gene expression and microbiota by serum metabolomics. BMC Complement Med Ther 21, 208 (2021). https://doi.org/10.1186/s12906-021-03382-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-021-03382-3