
Abstract
Background
This study investigated the anticancer potential of the medicinal herb, Cleome droserifolia (CD), a local plant of the Arabian Peninsula. C. droserifolia is traditionally known for its rubefacient, anti-diabetic, anti-oxidant, and anti-inflammatory properties.
Methods
Organic fractions of the aerial parts of Cleome droserifolia harvested from the Arabian Peninsula were tested in human breast and cervical cancer cell lines for their anticancer potential. This was accomplished by using biochemical and cellular assays, including MTT, caspase Glo, western blot, and annexin V/propidium iodide-based flow cytometry analyses.
Results
Test of the dichloromethane fraction of the methanolic extract of C. droserifolia, (CDD) revealed potent cytotoxic activity (from 70 to 90%) against several human cancer cell lines, including MCF-7, MDA-MB-231, and HeLa. Further characterization of the CDD fraction in MCF-7 cells revealed that it could activate the enzymatic activity of various caspases in a statistically significant manner, and induce cleavage of both caspase 7 and poly ADB ribose polymerase (PARP) proteins, but not the ethyl acetate fraction. Test of the ability of CDD to induce early signs of apoptosis was validated by annexin V/propidium iodide assay using FACS analysis. Induction of apoptosis was completely reversed by the classic pan inhibitor of apoptosis, Z-VAD-FMK, reducing early apoptosis from 29.7 to 0.6%, confirming that CDD could induce caspase-dependent apoptosis.
Conclusions
Altogether, our results reveal that C. droserifolia is a valuable medicinal plant with bioactive molecules that can induce apoptosis in human cancer cells. Thus, this plant should be explored further for its potential as an anticancer natural therapy as well as the isolation of novel molecules with anticancer properties.
Similar content being viewed by others


Background
Cleome droserifolia (Forssk) Del. (syn. Roridula droserifolia Forssk.), locally known as “samwa”, is an aromatic flowering shrub of the genus Cleome with ~ 200 species, belonging to the family Cleomaceae [1,2,3]. Other terminologies include rem el bard, afeen, and mashta in Arabic, while forssk in English. In addition, it is also known by other names such as “spider flower” and “mountain bee plant” [2,3,4,5]. All of the Cleome species grow at similar locations with different soil types. Moist places and rocky regions are favored for some species, while others grow in black fertile soil and rainy season, regions with waste water, and some in shaded areas in red soil which develops in warm, temperate, and moist climate during rainy season [2,3,4,5]. Cleome is found in tropical and subtropical countries in the Old and New Worlds, as well as in North Africa and Indian subcontinent [2,3,4,5].
C. droserifolia is an important species of Cleome due to its historical use in traditional medicine that is becoming increasingly endangered [4, 6, 7]. Plants in the Cleome genus improve stomach aches and treat many ailments like scabies and rheumatic fever [4,5,6,7]. They have immediate effect on abdominal and rheumatic pain, control inflammation, and are also effective towards wound healing, and snake & scorpion bites [4, 6,7,8]. These effects are attributed to their rubefacient, antimicrobial, analgesic, antipyretic, antioxidant, and anti-inflammatory activities [8,9,10,11]. For example, essential oils from three different species of Cleome, including C. droserifolia, were shown to have strong antibacterial properties owing to the essential oils being enriched in sulfur- and nitrogen-containing compounds [12]. C. droserifolia especially is well-known for its hypoglycemic effects, improving carbohydrate & lipid metabolism, fighting obesity, and enhancing antioxidant activity in diabetic rats & mice [13,14,15,16,17,18,19,20]. It also has anti-urinary schistosomiasis effects [21, 22].
C. droserifolia is rich in phytochemicals and several bioactive constituents have been isolated from this species (reviewed in [23, 24]). Numerous studies of C. droserifolia have revealed the presence of flavonoids, glycosides, carbohydrates, cardenolides, saponins, sterols, tannins, catechins, triterpenes, and sesquiterpenes, such as buchariol, teucladiol, daucosterol, and a new alkaloid from the aerial parts [22,23,24,25,26,27,28,29,30]. Other than these compounds, it has the distinction of being the first plant source of diterpenoid dolabellane esters as well [31]. Other Cleome species have been shown to contain numerous flavonoids glycosides [32,33,34], kaempferol 3-glucuronide from roots [35], a new naringenin glycoside [36], three new coumarino lignoids from seeds [37], and others [23, 24, 38]. Some of these constituents are thought to be responsible for the hypoglycemic effect of Cleome in animals [17,18,19] as well as its liver-protective properties [17, 39]. Thus, the isolation of several new phytonutrients from Cleome makes it an attractive candidate for further drug discovery [23, 24, 40].
Not much is known about the anticancer potential of Cleome. Cleome gynandra, has been shown to be effective when injected in Swiss albino mice using Ehlrich’s ascites carcinoma cells [41]. Similarly, extracts from another species, Cleome arabica, has been shown to have cytotoxic effects against the mouse leukemia cell line P388 by activating apoptosis and inhibiting phosphorylation of AKT and ERK kinases induced by the epidermal growth factor signaling [42]. Some of these cytotoxic effects could be attributed to the presence of dammarane triterpenes in these extracts that have been shown to have cytotoxic effects in P388 cells in MTT assays [28].
The aim of this study was to investigate the cytotoxic potential of C. droserifolia in more detail and determine its mechanism of action as part of the collaborative work being undertaken to collect, categorize, and study the biological potential of native land and marine flora and fauna of the Sultanate of Oman. Towards this end, the native C. droserifolia species was collected and identified and the aerial parts of the plant were used to prepare various organic fractions from its methanolic extract [30]. This was followed by test of the anticancer potential of these samples on breast and cervical cancer cell lines. Our results confirm the cytotoxic potential of C. droserifolia observed in the Egyptian variety [43] and further reveal that the active anticancer agents in this species have the ability to induce potent caspase-dependent apoptotic activity in human cancer cells. This finding supports the contention that medicinal plants serve as an important and rapidly dwindling source of novel and natural biomolecules for the fight against cancer and other human ailments [7, 44].
Methods
Plant collection, extraction, and fractionation
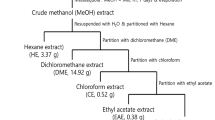
The fresh whole plant material was collected from Al-Jabel Al-Akhdar, Al-Dakhiliya region in Oman (2012) according to local and national rules and regulations. The specie was identified by a trained taxonomist at the Department of Biological Sciences and Chemistry, University of Nizwa, Oman. The voucher specimen (JCD/02/2012) was kept in the herbarium of Natural and Medical Sciences Research Center, University of Nizwa, Oman. As described earlier [30], 7.5 kg of whole aerial parts of C. droserifolia were used for extraction. These parts were dried in the dark, chopped and ground to a coarse powder, followed by organic extraction with methanol for 2–3 weeks at room temperature. After evaporation under pressure (leaving a greenish syrupy residue behind), the methanol extract (CDME) was suspended in distilled water and fractionated into n-hexane (CDH), dichloromethane (CDD), ethyl acetate (CDEA) and n-butanol (CDB) fractions (Fig. 1).

Preparation of organic fractions of Cleome droserifolia methanolic extract. Flow chart representing the steps used to prepare the various organic fractions of Cleome droserifolia (CD) starting from its methanolic extract (CDME). The residual aqueous fraction (20 g) is not shown in this scheme
To test the anticancer potential of the various Cleome droserifolia extracts/fractions, they were solubilized individually in the solvent dimethyl sulfoxide (DMSO) to prepare stock solutions. The concentration of the stock solutions varied for different extracts/fractions depending upon their form and solubility (12.5 mg/ml, 25 mg/ml, and 50 mg/ml), and then stored at -20 °C. From these stock solutions, different extract/fraction treatments were prepared for the MTT assays at twice the final concentration (100, 250, and 500 mg/mL) in media, depending upon the cell line used, since 100 μL of treatment was added to cells plated in 100 μL media on the day of treatment. Each experiment had control samples that contained the same concentration of DMSO as present in the tested fractions or their dilutions as a control for cell death induced by DMSO alone.
Cells and cell culture
The hormone responsive human breast cancer cell line, MCF-7 and the triple-negative human breast cancer cell line, MDA-MB-231, were obtained from Prof. Samir Attoub, Department of Pharmacology, College of Medicine and Health Sciences (CMHS), UAE University (UAEU), Al Ain, UAE. The normal breast epithelial cell line, MCF-10A, was obtained from ATCC (USA). The human cervical cancer cell line, HeLa, was a gift of Prof. Tahir A. Rizvi, Department of Microbiology and Immunology, CMHS, UAEU. The cell lines were cultured in the ATCC-recommended media as per instructions. All cells were maintained at 37 °C in a 5% CO2 incubator. Cells were plated a day before conducting the experiment and seeded at the cell density mentioned in the experiments, depending upon the plate and well size used for the experiment. This was followed by treatment of the cell cultures with different concentrations of the tested extracts along with appropriate dilutions of DMSO. Passage numbers of cell lines used for different experiments can be provided to the readers upon request.
MTT cell viability assay
All MTT [3-(4, 5-dimethylthiazol-2-yl)-2–5-diphenyltetrazolium bromide] assays were conducted in 96-well plates at a density of 5 × 103 cells/well/100 μL media. Appropriate normal or cancerous cell lines mentioned above were treated with 100 μL of the different concentrations of the organic extract/fraction of C. droserifolia, or DMSO (the solvent), or culture media alone, for the indicated time points. This was followed by addition of 25 μl/well of the MTT stock solution (5 mg/ml) and incubation for 3–4 h at 37 °C. DMSO was used to dissolve the formazan crystals that were formed (200 μl per well), followed by measurement of absorbance at 560 nm using the Perkin Elmer Victor TM 2030 Multi-label Reader. Cell viability was calculated as a percentage of cells treated with the test extract/fraction with cells treated with DMSO alone as a control using the same concentrations of DMSO as the test treated cells. Results were analyzed using MS Excel and GraphPad Prism v5 software.
Cell titer-GLO luminescent cell viability assay and Caspase-GLO 3/7, 8 and 9 assays
MCF-7 cells were cultured in opaque, white, 96-well plates (5000 cells/well) and treated with 50 μl of the different extract concentrations at 37 °C. The Cell Titer-Glo Luminescent Cell Viability Assay (Promega, USA) and Caspase-Glo 3/7,8 and 9 assays were used after 48 h to monitor cell viability and caspase enzyme activities, as per manufacturer’s instructions. The Infinite M200 Pro Tecan plate reader was used to measure luminescence and the resulting data was plotted as percent cell viability of experimental groups compared to DMSO-treated cells, the viability of which was taken as 100%.
Protein extraction and Western blot analysis
Total cell proteins were extracted from the treated cells and subjected to western blot analyses for investigation of cellular pathways activated upon extract/fraction treatment. Briefly, cells treated with various concentrations of the fractions were harvested by trypsinization, washed with 1 x PBS, and lysed using 100 μL of RIPA (radioimmunoprecipitation assay) lysis buffer (10 mM Tris-Cl (pH 8.0), 1 mM EDTA, 1% Triton X-100, 0.1% sodium deoxycholate, 0.1% SDS, 140 mM NaCl) per million cells supplemented with 50 μl of β-mercaptoethanol/ml RIPA and 1 mM of the serine protease inhibitor, phenylmethylsulfonyl fluoride (PMSF). Nuclei were separated from the lysed cells by spinning at 14,000 rpm for 10 min at 4 °C. The cell lysates were separated from the pellets and either used immediately or stored at -80 °C. Protein amounts in the cellular extracts were quantified using the Bio-Rad Bradford colorimetric assay.
Total protein lysates (40 μg per lane) were loaded onto 8–12% SDS-polyacrylamide mini gels (Bio-Rad) or 4–12% gradient gels (GeneScript, USA) and blotted onto Protran nitrocellulose membranes using standard protocol. After wet transfer, the membranes were blocked with 5% low fat dried milk in 1 x PBS and 0.1% Tween-20 (PBST) for 1 h and incubated at 4 °C overnight with the specified primary antibodies in 1% milk in PBST. All antibodies were purchased from Cell Signaling Technologies (USA). After washing, the blots were incubated with the appropriate secondary antibodies (anti-mouse or anti-rabbit) for 1 h and 30 min at room temperature followed by detection using the Pierce™ ECL Plus Western Blotting Substrate and visualized using Typhoon reader.
Annexin V/Propidium iodide flow cytometry analysis
Flow cytometry was used to test the ability of the effective extracts/fractions to induce apoptosis by inducing inversion of the phosphatidylserine phospholipid present at the inner plasma membrane to the outer layer of the phospholipid bilayer. This was accomplished by staining the treated and untreated cells with fluorescently labelled Annexin V (AV) and the vital dye, propidium iodide (PI) using a commercially available kit from Becton Dickenson, (Franklin Lakes, NJ USA). Ten thousand cells were counted per sample for flow cytometry using BD FACS Canto II, USA machine by counting all cells without gating to prevent sample bias. The apoptosis inhibitor, Z-VAD-FMK (Promega, USA), was used at a concentration 20 μM for 2 h prior to extract/fraction treatment. Z-VAD-FMK is a well-known pan-caspase inhibitor that is permeable to the cells. It binds to the catalytic site of caspase enzymes and inhibit their function, thereby preventing apoptosis.
Morphological studies
Control and treated cell cultures were analyzed for morphological changes by observation via an inverted light microscopic attached to a CCD camera or EVOS Cell Imaging System (Thermo-Fisher Scientific, USA). Images were saved at various magnifications at different times after treatment.
Statistical analysis
Statistical analysis was conducted using either Microsoft Excel or GraphPad Prism v5 software. In case of replicates, average values were calculated for each replicate followed by standard deviation calculations plotted as error bars on line or column graphs. Two-tailed, paired, ttest was used to calculate significant differences between groups, while extent of significance was depicted as stars, with * = p < 0.05, ** = p < 0.01 but > 0.001, and *** = p < 0.001. The MTT line graphs were generated by normalizing the values obtained for the test groups with that of their DMSO control by using the DMSO control data for each concentration tested. Same was the case for the Promega Glo assays where the effect of DMSO control was used to normalize the data obtained from the treated samples. Finally, the IC50 values for CDD were calculated by using the non-linear regression function of GraphPad Prism v7 and least square fit to estimate the final values.
Results
Screening of extracts/fractions for anticancer activity
Four organic fractions from the C. droserifolia (CD) methanolic extract (n-hexane, dichloromethane, ethyl acetate, and n-butanol) [30] were tested for their cytotoxic potential. The solvents were used with increasing polarity to ensure that bioactive molecules of a wide range of polarity were successfully extracted from the crude plant material [44]. Interestingly, the amount of n-hexane fraction obtained was ~ 45% of the total extract (53.4 g out of 117.8 g total extract; Fig. 1). This may be due to the nature of the compounds present in this species that contain high concentration of monoterpenoids, diterpenoids, cembrenes, and simple triterpenoids [23, 24, 45, 46]. We have fractionated another species of the same genus (Cleome austroarabica) and obtained more non-polar (n-hexane and chloroform) than polar compounds [47].
To determine the cytotoxic potential of these extracts/fractions, an initial screening was performed using the MTT assay in the breast cancer cell line, MCF-7 [48], and the cervical cancer cell line, HeLa [49] to determine which fraction had the potential to affect proliferation of either of these cancer cell types. The four fractions were tested in a dose- and time-dependent manner at different concentrations (50, 125, 250 μg/mL) and at two different time points (24 and 72 h) post treatment. Table 1 summarizes the results obtained. As can be seen, only the dichloromethane fraction (CDD) was able to induce significant cytotoxic effects in both MCF-7 and HeLa cultures. None of the other fractions (CDH, CDEA, or CDB) were able to induce > 20% death in either of the two cell lines.
CDD is efficient at inducing cytotoxic effects in both Normal and Cancer cells
To study cytotoxic potential of the CDD fraction further, it was tested in an expanded series of human cells, including MCF-10A, a normal breast cell line [50] and MDA-MB-231, the hard-to-treat, prototypic, triple-receptor-negative breast cancer cell line [51] along with the hormone-inducible MCF-7 and HeLa cells. First, the MTT assay was repeated in these cell lines in a dose- and time-dependent manner 24, 48, and 72 h post treatment (Fig. 2). This allowed us to compare effects of CDD in both normal and cancer cell lines at the same time and in the same manner.

The dichloromethane fraction of Cleome droserifolia methanolic extract (CDD) can induce potent cell death in normal and cancer human cell lines. Cytotoxic effect of A) 50 μg/ml, B) 125 μg/ml, and 250 μg/ml of CDD using the MTT assay on MCF-10A, MCF-7, MDA-MB-231, and HeLa cells in a dose- (a-c) and time- (d-f) dependent manner, after normalizing to the effects of DMSO solvent used to dissolve the extract. * indicates statistically significant differences between the control and treated samples (* = p < 0.05; ** p < 0.01 but > 0.001; *** = p < 0.001). The table below the figure shows the IC50 values calculated for CDD in the four cell lines tested
Figure 2 shows results of the test of CDD on the tested cell lines after normalizing the effects of DMSO on cell proliferation, an organic solvent that was used to dissolve CDD for testing purposes. As can be seen, statistically significant cell death could be observed in three of the four cell lines, even at exposure with 50 μg/mL CDD by 48 h post treatment, except for the triple-negative cell line MDA-MB-231 (Fig. 2a). All four cell lines became susceptible to CDD-induced cell death by treatment with 125 μg/mL of CDD at 24 h, leading to death of ~ 70% of MCF-10A and MCF-7 cells and ~ 60% of MDA-MB-231 and HeLa cells by 72 h post treatment (Fig. 2b). Treatment with 250 μg/mL CDD led to induction of > 90% cell death in all the three breast cell lines tested in a comparable fashion, with HeLa cells nearly catching up by 72 h post treatment (Fig. 2c).
Time-dependent analysis of CDD-induced cell death revealed that once again, the three breast cell lines were similar in their susceptibility to CDD early on with the HeLa cells nearly catching up by 72 h post treatment with the different doses of CDD (Fig. 2d-f). Interestingly, HeLa cells were consistently more susceptible to CDD-induced killing at lower doses (50 μg/mL) than the breast cells, as observed by the steeper slope of the kill curve at 48 and 72 h (Fig. 2e & f). These subtleties were reflective in the IC50 values calculated at the 250 μg/ml value 72 h post treatment which revealed that the normal MCF-10A were the most sensitive to CDD-induced killing with an IC50 value of 57.34 μg/ml, followed by HeLa and MCF-7 cells at 73.23 and 74.63 μg/mL, respectively, while MDA-MB-231 were less sensitive with an IC50 of 122.8 μg/ml. These values are in the same range as observed for the cell killing potential of two other medicinal plants that we have tested in these cell lines [52, 53]. To ensure that the cytotoxicity being observed was not due to any residual organic solvent in the tested fractions, each of the organic solvents by itself was tested on the three cancer cell lines and observed to be non-toxic (data not shown). Further NMR analysis of the organic fractions also confirmed absence of residual organic solvent, confirming our results that bioactive components within C. droserifolia are the ones that affect cell proliferation of both human breast and cervical cancer cells significantly, but at the same time, they also affect the viability of normal cells. Finally, the fact that none of the other three fractions showed any significant signs of cytotoxicity also confirmed that residual solvents in the fractions were not responsible for the cytotoxicity being observed.
CDD can induce enzymatic activity and cleavage of key apoptotic proteins
To explore the potential mechanism of action of the effective extracts, the ability of CDD to induce apoptosis was explored since apoptosis is the most widely induced type of cell death pathway observed by natural compounds [54]. MCF-7 cells were treated with CDD for 48 h and assayed for its ability to induce the activity of various caspase enzymes, a hallmark of apoptosis induction [55]. As can be seen, CDD induced potent anti-proliferative effects in MCF-7 cells (> 80%) and was also able to induce the caspase enzyme activity in a statistically significant manner (Fig. 3). This was reproducible, as observed in two independent experiments conducted in triplicates (Fig. 3a and b). The levels of caspase 8 & 9 induced were higher than that of caspase 3/7. The effect on cell viability correlated well with the results of the MTT assay (Fig. 2). All results obtained for the induction of caspases were normalized to the number of viable cells in culture to ensure that the results were not skewed due to the ensuing cell death (see Methods). Since MCF-7 cells do not express caspase 3 [56], any induction of caspase 3/7 in this system is most likely due to the induction of caspase 7.

The dichloromethane fraction of Cleome droserifolia methanolic extract (CDD) can activate the enzymatic activity of caspase 3/7, 8, and 9. MCF-7 cells were treated with 250 μg/ml of CDD for 48 h. a) and b) show results from two independent experiments. *, statistically significant differences between the control and treated samples (* = p < 0.05; ** p < 0.01 but > 0.001; *** = p < 0.001). DMSO Control, MCF-7 cells treated with media containing the same amount of DMSO as the CDD-treated samples
To confirm whether CDD tested in the Promega Glo enzyme assays for caspase activation could indeed induce apoptosis, our next aim was to confirm the activation of caspases associated with apoptosis using western blot analyses. MCF-7 cells were treated with CDD for 24 h and tested for the expression and cleavage of various caspases and poly ADP-ribose polymerase (PARP). PARP is one of the enzymes that is cleaved during the execution phase of apoptosis by activated caspases [55, 57]. Thus, activation of caspases and PARP by cleavage are two major indicators of a cell undergoing apoptosis.
Figure 4a shows the western blot analysis of CDD on MCF-7 cells 24 h post treatment. As can be seen, we could not detect a cleaved band for any of the caspase proteins and in fact, the antibody reactivity for caspase 8 and 9 was very poor for these proteins from MCF-7 cells; however, diminished amounts of procaspases 7, 8, and 9 could be observed at 24 h, suggesting that the cleavage reaction had taken place (Fig. 4a). Test of the PARP protein revealed significant reduction of full-length PARP, also suggesting its activation (Fig. 4a). To verify the results, MCF-7 cells were once again treated with the extract, but for shorter time periods in case the cleavage event had been missed due to the timing of the analysis. Figure 4b shows the results for MCF-7 cells treated for < 15 mins, 12 h, and 24 h with CDD, representing an early, intermediate, and late time point for detecting caspase cleavage. Clear cleavage of caspase 7 and PARP could be observed starting from around 15 min post treatment. The cleaved product decreased or essentially disappeared by 24 h, as observed earlier (Fig. 4b vs 4a). This observation confirms that CDD has the potential to induce apoptosis in MCF-7 cells.

The dichloromethane fraction of Cleome droserifolia methanolic extract (CDD) can induce cleavage of procaspase 7 and PARP. MCF-7 cells were treated with 250 μg/ml of CDD for a) 24 h, and b) multiple time points. Western blots were carried out on the untreated, DMSO Control, and CDD-treated cellular lysates for the specified caspase or poly ADB ribose polymerase (PARP) proteins. Beta-actin antibody was used as a loading control. DMSO-treated, DMSO control having the same amount of DMSO as the test samples
To ensure that the results being observed were truly due to the ability of CDD to cause apoptosis, we took the ethyl acetate fraction of the CD methanol extract (CDEA) that had shown an inability to cause cell death in MCF-7 cells (Table 1) and treated the MCF-7 cells with it in a similar manner (Fig. 5). Results of the western blot analysis of CDEA-treated cells revealed a complete lack of cleavage of either the procaspases 7, 8, or 9, or PARP, or decrease in their expression, confirming that the cleavage of the procaspases and PARP was truly due to the ability of CDD to induce apoptosis.

The ethyl acetate fraction of Cleome droserifolia methanolic extract (CDEA) is unable to induce apoptosis. Western blot analysis of MCF-7 cells treated with 250 μg/ml of CDEA for various time points and analyzed with the specified antibodies. Beta-actin antibody was used as a loading control. DMSO Control, control sample having the same amount of DMSO as the test samples
CDD can induce early signs of apoptosis
To confirm whether CDD could indeed induce apoptosis, we tested the potential of CDD to induce the inversion of the plasma membrane phospholipids, an early sign of apoptosis [55]. Towards this end, MCF-7 cells were treated with CDD and tested for the inversion of the phosphoserine phospholipid that is found exclusively at the inner leaflet of the plasma membrane using FITC-labelled Annexin V (AV), a carbohydrate that has high affinity for phosphoserines moieties [55]. Furthermore, to determine whether the cell membrane integrity was maintained, cells were stained with propidium iodide (PI), a vital dye that can differentiate between live and dead cells. PI can enter the cells once the cell membrane integrity has been compromised in dead cells and therefore is excluded from live cells [55].
Figure 6 shows the flow cytometric analysis of the MCF-7 cells treated with CDD 24 h post treatment. As can be seen, 99.9% of the untreated and DMSO-treated cells were observed to be in the AV(−)/PI(−) quadrant, indicative of live healthy cells (Fig. 6a and b). Treatment with an essential oil from Boswellia sacra (positive control) that induces potent apoptosis in breast cancer cells [52, 53, 58] shifted 74% of the cells to the AV(+)/PI(−) quadrant indicative of early apoptosis, and 5.5% of the cells to the AV(+)/PI(+) quadrant, indicative of late apoptosis (Fig. 6c). In comparison, 29.7% of the cells were shifted to the early apoptosis by CDD and 1.6% of the cells to the late apoptosis stage (Fig. 6d). Pre-treatment of the MCF-7 cells with Z-VAD-FMK, a classical inhibitor of apoptosis [55], was able to completely reverse the cell death being observed in the CDD-treated cultures with 98.9% of the cells appearing in the quadrant indicting healthy and live cells (Fig. 6e). These results confirm that CDD has the potential to induce apoptosis in MCF-7 cells.

The dichloromethane fraction of Cleome droserifolia methanolic extract (CDD) induces inversion of inner plasma membrane phospholipids. FACS analysis of MCF-7 cells stained with Annexin V/propidium iodide after treatment with: a media alone, b media with DMSO, cBoswellia sacra essential oil, a known apoptosis inducer, d 250 μg/ml CDD for 24 h, or E) 250 μg/ml CDD after pre-treatment with 20 μM Z-VAD-FMK for 2 h. DMSO Control, MCF-7 cells treated with the same amount of DMSO as the test samples
Microscopic analysis of cells treated with CDD for 24 h revealed that unlike the untreated and DMSO-treated cells that were found to create nice monolayers (Fig. 7a and b), the CDD-treated cells exhibited cell rounding and shrinkage, characteristic of apoptosis, as observed with Boswellia sacra (Fig. 7c and d), followed by their removal from the monolayer. Pre-treatment of cells with Z-VAD-FMK essentially maintained the healthy phenotype of the CDD-treated monolayer (Fig. 7e), confirming that indeed CDD induces apoptosis in MCF-7 cells.

The dichloromethane fraction of Cleome droserifolia methanolic extract (CDD) induces morphological changes reminiscent of apoptosis in breast cancer cells. Photomicrographs of MCF-7 cells treated with a) media alone, b) media with DMSO, c) Boswellia sacra essential oil, a known apoptosis inducer, d) 250 μg/ml of CDD for 24 h, or e) 250 μg/ml CDD after pre-treatment with 20 μM Z-VAD-FMK for 2 h. DMSO Control, MCF-7 cells treated with the same amount of DMSO as the test samples. Magnification: 400X
Effect of C. droserifolia on autophagy
Finally, we explored the ability of CDD to activate autophagy, a type II cell death pathway normally activated by nutrient deprivation, growth factor depletion, or hypoxia [59]. Induction of autophagy is actively being explored as an important cellular pathway activated by anticancer compounds [60,61,62,63]. Our earlier work with two other medicinal plants, Teucrium mascatense and Acridocarpus orientalis has shown that induction of apoptosis by natural products can activate/perturb autophagy as well [52, 53]. Therefore, we tested the ability of CDD to induce key proteins of the autophagy pathway, the autophagy receptor, SQSTM1/p62, responsible for recognizing the ubiquitinated cellular cargo targeted for degradation, and the microtubule-associated protein 1 light chain 3 expressed in the cytoplasm (LC3-I) or its cleaved form (LC3-II) that is recruited to the autophagosome via lipidation upon autophagy activation [59].
Treatment of MCF-7 breast cancer cells with 250 μg/mL of CDD led to the oligomerization of SQSTM/p62 protein (mostly dimer and some trimer formation) 24 h post treatment; the monomer, though remained the most abundant species (Fig. 8; lane 3, upper panel). By 48 h, both the dimer and trimer had increased, with the dimer being still more abundant than the trimer form, while the monomer amount decreased compared to the 24-h time point (Fig. 8; lane 4 vs lane 3; upper panel). Treatment with CDD also induced the expression of cytosolic LC3-I and the membrane-bound LC3-II by 24 h in equal proportions (Fig. 8; lane 2 vs lane 3; middle panel). This suggested recruitment of soluble LC3–1 to the developing autophagosomes, a process that continued for 48 h, as observed by the increase in LC3-II band compared to LC3-I, signifying continued activation of autophagy (Fig. 8, lanes 3 and 4; middle panel). Beta-actin served as the control with no effect of CDD either on its expression or stability Fig. 8, lanes 1–4; lower panel), confirming that the results being observed were due to CDD. These results suggest induction of autophagy in the MCF-7 cells by CDD.

The dichloromethane fraction of Cleome droserifolia methanolic extract (CDD) induces expression of key proteins of autophagy in breast cancer cells. Western blot analysis of MCF-7 cells treated with 250 μg/ml of CDD for indicated time points. Beta-actin antibody was used as a loading control. DMSO-treated, DMSO control having the same amount of DMSO as the test samples
Discussion
This study characterized the cytotoxic potential of an important medicinal plant of the Arabian Peninsula, C. droserifolia, growing in Oman. It revealed that the dichloromethane fraction of the methanol extract, CDD (Fig. 1), had effective cytotoxic activity against human cancer cells (Table 1 & Fig. 1). CDD efficiently killed not only human breast but also cervical cancer cells in a dose- and time-dependent manner (Fig. 2). Investigation of the mechanism of action of CDD revealed that it had the ability to induce caspase 7, 8, and 9 enzymatic activity (Fig. 3) and activated cleavage of caspase 7 & PARP proteins (Fig. 4). Finally, FACS analysis of cells treated with CDD for Annexin V/PI staining confirmed that apoptosis could be induced by CDD which was reversible by the classical apoptosis inhibitor, Z-VAD-FMK (Figs. 6 & 7) in the hormonally-responsive MCF-7 breast cancer cell line.
These results support the observations made by Ezzat and colleagues on the C. droserifolia species from Egypt in two human cancer cell lines: MCF-7 and the colon carcinoma cell line, HCT116 [43]. Their tests revealed that both the aqueous and ethanolic extracts of the aerial parts of C. droserifolia had cytotoxic potential in these cell types. The aqueous extract was further fractionated using four organic solvents amongst which the chloroform, ethyl acetate, and n-hexane fractions were observed to have the most active cytotoxic potential, while the n-butanol fraction had weaker cytotoxic activity. Furthermore, nine compounds (six terpenoids and two flavanol glycosides) were isolated from the chloroform and ethyl acetate fractions of which two terpenoids and both flavonols had cytotoxic potential against the two cell lines, similar to those observed with the anticancer drug, Doxorubicin [43]. We only tested fractions of the methanolic extract for its cytotoxic potential. Furthermore, we have previously reported the isolation of two known aromatic derivatives, veratrol and 2-methoxy-4-methylacetophenone, and one novel alkaloid (5-hydroxy-2-methoxy-1-methy-1H-indole-3-carbaldehyde) from the dichloromethane fraction [30]; however, their anticancer cytotoxic potential remains to be tested owing to the insufficient availability of enough pure compounds.
Other than cytotoxicity for cancer cells, CDD also was able to kill normal human breast cells (Fig. 2). The ability of CDD to induce killing of normal cells is not surprising given the fact that we tested a crude organic fraction representing a mixture of compounds, and not a pure compound. Thus, it is possible that once the active biomolecules representing the cytotoxic principals have been isolated and separated from each other, we should be able to identify molecules that are able to induce death of only cancer and not normal cells. Even if that is not possible, our identification of crude fractions with anticancer potential, especially for breast cancer cells, is significant since it has been hard to identify natural compounds that have cytotoxicity for breast cancer cells in large screenings of natural compounds [64, 65].
Interestingly, we found that dichloromethane was the best solvent at extracting the cytotoxic potential from the methanolic extract. None of the other solvents could purify molecules with any significant cytotoxic activity. This observation agrees with our studies with other plant extracts where dichloromethane was the best at isolating the anticancer potential of the plant [52, 53]. These observations suggest that the dichloromethane solvent is one of the better organic solvents at extracting anticancer bioactive molecules from medicinal plants with cytotoxic effects against human cancer cells.
Finally, we also studied how treatment with a natural plant product like CDD can affect autophagy, an important cellular catabolic pathway responsible for maintaining cell health by engulfing damaged proteins and organelles, thereby recycling nutrients needed for cell survival [59]. We observed that CDD was efficient at inducing autophagy, as assessed by the induction and oligomerization of the autophagy receptor protein SQSTM1/p62 as well as activation of the cytosolic LC3-I form followed by its recruitment to the membrane-bound LC3-II (Fig. 8). We have observed similar effects of two other natural plant products on autophagy when apoptosis was induced by their organic fractions, although the precise pattern of induction of SQSTM1/p62 and LC3 was different with each plant product [52, 53]. In fact, this pattern differed even between the organic fraction from leaves (AOD (L)) versus the stems (AOB (S)) from the same plant, Acridocarpus orientalis. For example, AOD (L) had a lower potential of inducing apoptosis and was more efficient at inducing the dimer form of SQSTM1/p62 than AOB (S), while AOB (S) was more effective at inducing apoptosis and was also more efficient at inducing both forms of LC3 than AOD (L) [53]. These observations suggest that induction of apoptosis can have direct effects on autophagy, depending upon the nature and type of stress being imposed by the constituents of the medicinal plant product.
The role of autophagy in cancer cell death and survival is quite complex, especially in the presence of apoptosis [66, 67]. Autophagy has been shown to cause cell death along with apoptosis or ensure cell survival by inhibiting apoptosis [68]. Interestingly, it may even be a requirement for activating apoptosis in cells which are faced with stresses that can activate autophagy itself, such as oxidative stress or nutrient depravation [69, 70]. Thus, although we observed activation of autophagy in the case of CDD as well as other plant organic fractions [52, 53], it is not clear whether it was a requirement or a consequence of apoptotic cell death, manifesting as a cell survival response. For example, it has been shown that oligomerization of SQATM1/p62 by oxidative stress is suggestive of induction of autophagy as a pro-survival mechanism rather than a cell death process [69, 70]. Alternatively, it could also be an independent response to compounds in CDD or the other plant extracts. Thus, tweaking this out will require further careful experimentation with inhibitors of both apoptotic and autophagic pathways, followed by cellular and biochemical assays to determine the role being played by CDD or other natural compounds in the induction of autophagy.
Conclusions
This study reveals that C. droserifolia has strong cytotoxic potential against human cancer cells. It further shows that the bioactive molecules in this plant exert their effect by inducing caspase-dependent apoptosis that could essentially be reversed by Z-VAD-FMK. Thus, C. droserifolia is a valuable medicinal plant in the fight against cancer which can also serve as a biological source of novel natural anticancer agents.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- AOB (S):
-
Acridocarpus orientalis n-butane stem fraction
- AOD (L):
-
Acridocarpus orientalis dichloromethane leaf fraction
- CDB:
-
C. droserifolia n-butanol fraction
- CDD:
-
C. droserifolia dichloromethane fraction
- CDEA:
-
C. droserifolia ethyl acetate fraction
- CDH:
-
C.droserifolia n-hexane fraction
- DMEM:
-
Dulbecco’s Modified Eagles Medium
- DMSO:
-
Dimethyl sulfoxide
- EGF:
-
Epidermal growth factor
- FBS:
-
Fetal bovine serum
- FCS:
-
Fetal calf serum
- LC3:
-
Microtubule-associated protein 1 light chain 3
- MTT:
-
[3-(4, 5-dimethylthiazol-2-yl)-2–5-diphenyltetrazolium bromide]
- PARP:
-
Poly (ADP-ribose) polymerase
- PBS:
-
phosphate-buffered saline
- PBST:
-
PBS and 0.1% Tween-20
- PMSF:
-
Phenylmethyl sulfonyl fluoride
- RIPA:
-
Radioimmunoprecipitation assay buffer
- SDS:
-
Sodium dodecyl sulfate
References
World Flora Online (2020): World Flora Online. Published on the Internet; http://www.worldfloraonline.org. Accessed 18 Jan 2020.
Batanouny K. Wild Medicinal Plants in Egypt. Academy of Scientific Research and Technology, Egypt. The World Conservation Union (IUCN), Switzerland. 1999.
Boulos L. Flora of Egypt. Cairo: Al Hadara Pub; 1999.
Aparadh V, Mahamuni R, Karadge B. Taxonomy and physiological studies in spider flower (Cleome species): a critical review. Plants Sci Feed. 2012;2:25–46.
Kamel W, El-Ghani M, El-Bous M. Cleomaceae as a distinct family in the Flora of Egypt. Afr J Plant Sci Biotech. 2010;4:11–6.
Rahman MA, Mossa JS, Al-Said MS, Al-Yahya MA. Medicinal plant diversity in the flora of Saudi Arabia 1: a report on seven plant families. Fitoterapia. 2004;75:149–61.
Moustafa ARA, Mansour SR, Alotaibi MO. Cleome droserifolia. An Egyptian natural heritage facing extinction. Asian J Plant Sci Res. 2019;9:14–21.
Sarhan WA, Azzazy HM, El-Sherbiny IM. Honey/chitosan Nanofiber wound dressing enriched with Allium sativum and Cleome droserifolia. Enhanced antimicrobial and wound healing activity. ACS Appl Mater Interfaces. 2016;8:6379–90.
Bouriche H, Selloum L, Tigrine C, Boudoukha C. Effect of Cleome arabica leaf extract on rat paw edema and human neutrophil migration. Pharm Biol. 2003;41:10–5.
Narendhirakannan RT, Subramanian S, Kandaswamy M. Anti-inflammatory and lysosomal stability actions of Cleome gynandra L. studied in adjuvant induced arthritic rats. Food Chem Toxicol. 2007;45:1001–12.
Abd El-Gawad AM, El-Amier YA, Bonanomi G. Essential oil composition, antioxidant and allelopathic activities of Cleome droserifolia (Forssk.) Delile. Chem Biodivers. 2018;15:e1800392.
Muhaidat R, Al-Qudah M, Samir O, Jacob J, Hussein E, Al-Tarawneh I, et al. Phytochemical investigation and in vitro antibacterial activity of essential oils from Cleome droserifolia (Forssk.) Delile and C. trinervia Fresen (Cleomaceae). South Afr J Bot. 2015;99:21–8.
Marles RJ, Farnsworth NR. Antidiabetic plants and their active constituents. Phytomedicine. 1995;2:137–89.
Mikhail Y. Studies on the hypoglycemic effects of Cleome drosrifolia and bran of Triticum vulgaris. Cairo: Cairo University; 2000.
Helal I, Sharaf H, Matter F. Anti-diabetic and anti-obesity effects of plant extract from Cleome droserifolia (samwa). Egyption J Hosp Med. 2002;9:85–101.
Naggar E, Bartošíková L, Žemlička M, Švajdlenka E, Rabišková M, Strnadová V, et al. Antidiabetic effect of Cleome droserifolia aerial parts: lipid peroxidation-induced oxidative stress in diabetic rats. Acta Vet Brno. 2005;74:347–52.
El-Shenawy N, Abdel-Nabi I. Comparative analysis of the protective effect of melatonin and Cleome droserifolia extract on antioxidant status of diabetic rats. Egyption J Hosp Med. 2004;14:11–25.
Al-Shenawy N, Abdel-Nabi I. Hypoglycaemic effect of Cleome droserifolia ethanolic leaves extract in experimental diabetes and their effect on non-enzymatic antioxidant glycogen content thyroid hormones and insulin levels. Diabetol Croat. 2006;35:15–22.
Motaal AA, Ezzat SM, El-Askary H. Antihyperglycemic activity and standardization of the bioactive extract of Cleome droserifolia growing in Egypt. Pharmacogn J. 2014;65:15–21.
Nicola WG, Ibrahim KM, Mikhail TH, Girgis RB, Khadr ME. Role of the hypoglycemic plant extract Cleome droserifolia in improving glucose and lipid metabolism and its relation to insulin resistance in fatty liver. Boll Chim Farm. 1996;135:507–17.
Ndamba J, Nyazema N, Makaza N, Anderson C, Kaondera KC. Traditional herbal remedies used for the treatment of urinary schistosomiasis in Zimbabwe. J Ethnopharmacol. 1994;42:125–32.
NS EL-S, MFM S, Abdel-Nabi IM. Does Cleome droserifolia have anti-schistosomiasis mansoni activity? Rev Inst Med Trop Sao Paulo. 2006;484:223–8.
Singh H, Mishra A, Mishra AK. The chemistry and pharmacology of Cleome genus: a review. Biomed Pharmacother. 2018;101:37–48.
Abdullah W, Elsayed WM, Abdelshafeek KA, Nazif NM, Singab ANB. Chemical constituents and biological activities of Cleome genus: a brief review. IJPR. 2016;8:777–87.
Fushiya S, Kishi Y, Hattori K, Batkhuu J, Takano F, Singab AN, et al. Flavonoids from Cleome droserifolia suppress NO production in activated macrophages in vitro. Planta Med. 1999;65:404–7.
Aboushoer MI, Fathy HM, Abdel-Kader MS, Goetz G, Omar AA. Terpenes and flavonoids from an Egyptian collection of Cleome droserifolia. Nat Prod Res. 2010;24:687–96.
El-Askary HI. Terpenoids from Cleome droserifolia Forssk. Del Mol. 2005;10:971–7.
Nagaya H, Tobita Y, Nagae T, Itokawa H, Takeya K, Halim AF, Abdel-Halim OB. Cytotoxic triterpenes from Cleome africana. Phytochemistry. 1997;44:1115–9.
Hussein NS, Ahmed AA, Darwish FMK. Sesquiterpenes from Cleome droserifolia. Pharmazie. 1994;49:76–7.
Hussain J, Khan H, Ali L, Khan AL, Rehman NU, Jahangir S, et al. New indole alkaloid from Cleome droserifolia. Helvetica Chimica Acta. 2015;98:719–23.
Fathy HM, Aboushoer MI, Harraz FM, Omar AA, Goetz G, Tabacchi R. Dolabellane Diterpenes from Cleome droserifolia. Natural Product Commun. 2008;3:1479–82.
Senthamilselvi MM, Kesavan D, Sulochana N. An anti-inflammatory and antimicrobial flavones glycoside from flowers of Cleome viscosa. Org Med Chem Lett. 2012;2:1–5.
Phan NM, Nguyen TP, Le TD, Mai TC, Phong MT, Mai DT. Two new flavonol glycosides from the leaves of Cleome viscosa. L Phytochem Lett. 2016;18:10–3.
Kjaer A, Thomsen H. Isothiocyanate-producing glucosides in species of Capparidaceae. Phytochemistry. 1963;2:29–32.
Chauhan J, Srivastava S, Srivastava S. Kaempferide 3-glucuronide from the roots of Cleome viscosa. Phytochemistry. 1979;18:691.
Srivastava S, Chauhan J, Srivastava S. A new naringenin glycoside from Cleome viscosa. Phytochemistry. 1979;18:2057–8.
Ray A, Chattopadhyay S, Kumar S, Konno C, Kiso Y, Hikino H. Structures of cleomiscosins, coumarinolignoids of Cleome viscosa seeds. Tetrahedron. 1985;41:209–14.
Qin GW, Hamed AI, el-Emary NA, Chen YG, Wang LQ, Cheung KK, Cheng KF. A new trinortriterpenoid from Cleome chrysantha. Planta Med. 2000;66:191–3.
Abdel-Kader MS, Alqasoumi SI, Al-Taweel AM. Hepatoprotective constituents from Cleome droserifolia. Chem Pharm Bull (Tokyo). 2009;57:620–4.
Tiwari P, Kumar B, Kaur M, Kaur G, Kaur H. Phytochemical screening and extraction: a review. Int Pharmaceutica Sci. 2011;1(1):98–106.
Bala A, Kar B, Haldar PK, Mazumder UK, Bera S. Evaluation of anticancer activity of Cleome gynandra on Ehrlich's ascites carcinoma treated mice. J Ethnopharmacol. 2010;129:131–4.
Tigrine C, Bulzomi P, Leone S, Bouriche H, Kameli A, Marino M. Cleome arabica leaf extract has anticancer properties in human cancer cells. Pharm Biol. 2013;51:1508–14.
Ezzat SM, Abdel MA. Isolation of new cytotoxic metabolites from Cleome droserifolia growing in Egypt. Z Naturforsch C. 2012;67:266–74.
Cowan MM. Plant products as antimicrobial agents. Clin Microbiol Rev. 1999;12:564–82.
Dwight O, Collins DO, Reynolds WF, Reese PB. New cembranes from Cleome spinosa. J Nat Prod. 2004;67:179–83.
Jente R, Jaklipwic J, Olatunji GA. A cembranoid diterpene from Cleome viscosa. Phytochemistry. 1990;29:666–7.
Weli AM, Al-Harrasi A, Al Baiti NH, Philip A, Hossain A, Gilani SA, et al. Biological and toxicological evaluation of aerial parts extracts of locally grown Cleome austroarabica. J King Saud University–Sci. 2020;32:753–7.
Soule HD, Vazguez J, Long A, Albert S, Brennan M. A human cell line from a pleural effusion derived from a breast carcinoma. J Natl Cancer Inst. 1973;51:1409–16.
Scherer WF, Syverton JT, Gey GO. Studies on the propagation in vitro of poliomyelitis viruses. IV. Viral multiplication in a stable strain of human malignant epithelial cells (strain HeLa) derived from an epidermoid carcinoma of the cervix. J Exp Med. 1953;97:695–710.
Soule HD, Maloney TM, Wolman SR, Peterson WD Jr, Brenz R, McGrath CM, et al. Isolation and characterization of a spontaneously immortalized human breast epithelial cell line, MCF-10. Cancer Res. 1990;50:6075–86.
Cailleau R, Young R, Olive M, Reeves WJ Jr. Breast tumor cell lines from pleural effusions. J Natl Cancer Inst. 1974;53:661–74.
Panicker NG, Balhamar SOMS, Akhlaq S, Qureshi MM, Rizvi TS, Al-Harrasi A. Identification and characterization of the caspase-mediated apoptotic activity of Teucrium mascatense and an isolated compound in human cancer cells. Molecules. 2019;24:977.
Balhamar SOMS, Panicker NG, Akhlaq S, Qureshi MM, Ahmad W, Rehman NU. Differential cytotoxic potential of Acridocarpus orientalis leaf and stem extracts with the ability to induce multiple cell death pathways. Molecules. 2019;24:3976.
Fulda S. Modulation of apoptosis by natural products for cancer therapy. Planta Med. 2010;76:1075–9.
Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007;35:495–516.
Jänicke RU. MCF-7 breast carcinoma cells do not express caspase-3. Breast Cancer Res Treat. 2009;117:219–21.
Chaitanya GV, Steven AJ, Babu PP. PARP-1 cleavage fragments: signatures of cell-death proteases in neurodegeneration. Cell Commun Signal. 2010;8:31.
Suhail MM, Wu W, Cao A, Mondalek FG, Fung KM, Shih PT, Fang YT, Woolley C, Young G, Lin HK. Boswellia sacra essential oil induces tumor cell-specific apoptosis and suppresses tumor aggressiveness in cultured human breast cancer cells. BMC Complement Altern Med. 2011;11:129.
Green DR, Llambi F. “Cell Death Signaling” in Cold Spring Harb Perspect Biol 2015;7:a006080.
Galluzzi L, Pietrocola F, Pedro JMB-S, Amaravadi RK, Baehrecke EH, Cecconi F, et al. Autophagy in malignant transformation and cancer progression. EMBO J. 2015;34:856–80.
Gali-Muhtasib H, Hmadi R, Kareh M, Tohme R, Darwiche N. Cell death mechanisms of plant-derived anticancer drugs: beyond apoptosis. Apoptosis. 2015;20:1531–62.
Chen N, Karantza V. Autophagy as a therapeutic target in cancer. Cancer Biol Ther. 2011;11:157–68.
Wang N, Feng Y. Elaborating the role of natural products-induced autophagy in cancer treatment: achievements and artifacts in the state of the art. Biomed Res Int. 2015;2015:934207. https://doi.org/10.1155/2015/934207.
Suffredini IB, Paciencia ML, Frana SA, Varella AD, Younes RN. In vitro breast cancer cell lethality of Brazilian plant extracts. Pharmazie. 2007;62:798–800.
Fouche G, Cragg GM, Pillay P, Kolesnikova N, Maharaj VJ, Senabe J. In vitro anticancer screening of south African plants. J Ethnopharmacol. 2008;119:455–61.
Yun CW, Lee SH. The roles of autophagy in Cancer. Int J Mol Sci. 2018;19:3466.
Doherty J, Baehrecke EH. Life, death and autophagy. Nat Cell Biol. 2018;20:1110–7.
Das G, Shravage BV, Baehrecke EH. Regulation and function of autophagy during cell survival and cell death. Cold Spring Harb Perspect Biol. 2012;4:a008813.
Carroll B, Otten EG, Manni D, Stefanatos R, Menzies FM, Smith GR, et al. Oxidation of SQSTM1/p62 mediates the link between redox state and protein homeostasis. Nat Commun. 2018;9:256.
Mildenberger J, Johansson I, Sergin I, Kjøbli E, Damås JK, Razani B. N-3 PUFAs induce inflammatory tolerance by formation of KEAP1-containing SQSTM1/p62-bodies and activation of NFE2L2. Autophagy. 2017;13:1664–78.
Acknowledgements
MCF-7 and MDA-MB-231 were a gift from Prof. Samir Attoub, Department of Pharmacology, College of Medicine and Health Sciences (CMHS), UAE University (UAEU). MCF-10A cell line was purchased from ATCC (USA), while HeLa cells were a gift of Prof. Tahir A. Rizvi, Department of Microbiology and Immunology, CMHS, UAEU. We thank Dr. Waqar Ahmad, Department of Biochemistry, CMHS, UAEU for his assistance with IC50 calculations and referencing.
Funding
This research was funded by grant numbers 31M122, 31R122, and 31R087 to FM from the UAE University and The Oman Research Council (TRC) (grant number: ORG/CBS/12/004) to JH and AAH. The funding bodies had no role in the design of the study, collection, analysis, or interpretation of data or in writing the manuscript.
Author information
Authors and Affiliations
Contributions
SOMSB, NGP, SA, MMQ, and NUR performed the experiments, SOMBS, NGP, and FM analyzed and interpreted the data, JH and AAH provided the extracts, SOMSB and FM wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Panicker, N.G., Balhamar, S.O.M.S., Akhlaq, S. et al. Organic extracts from Cleome droserifolia exhibit effective caspase-dependent anticancer activity. BMC Complement Med Ther 20, 74 (2020). https://doi.org/10.1186/s12906-020-2858-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-020-2858-0