
Abstract
Background
Galetin 3,6-dimethyl ether (FGAL) is a flavonoid isolated from aerial parts of Piptadenia stipulacea. Previously, FGAL was shown to inhibit both carbachol- and oxytocin-induced phasic contractions in the rat uterus, which was more potent with oxytocin. Thus, in this study, we aimed to investigate the tocolytic action mechanism of FGAL on the rat uterus.
Methods
Segments of rat uterus ileum were suspended in organ bath containing modified Locke-Ringer solution at 32 °C, bubbled with carbogen mixture under a resting tension of 1 g. Isotonic contractions were registered using kymographs and isometric contractions using force transducer.
Results
FGAL was more potent in relaxing uterus pre-contracted with oxytocin than with KCl. Additionally, FGAL shifted oxytocin-induced cumulative contractions curves to the right in a non-parallel manner, with Emax reduction, indicating a pseudo-irreversible noncompetitive antagonism of oxytocin receptors (OTR) or a downstream pathway target. Moreover, FGAL shifted CaCl2-induced cumulative contraction curves to the right in a non-parallel manner in depolarizing medium, nominally without Ca2+, with Emax reduction, suggesting the inhibition of Ca2+ influx through CaV. The relaxant potency of FGAL was reduced by CsCl, a non-selective K+ channel blocker, suggesting positive modulation of these channels. Furthermore, in presence of apamin, 4-aminopyridine, glibenclamide or 1 mM TEA+, the relaxant potency of FGAL was attenuated, indicating the participation of SKCa, KV, KATP and highlighting BKCa. Aminophylline, a non-selective phosphodiesterase (PDE) blocker, did not affect the FGAL relaxant potency, excluding the modulation of cyclic nucleotide PDEs pathway by FGAL.
Conclusion
Tocolytic effect of FGAL on rat uterus occurs by pseudo-irreversible noncompetitive antagonism of OTR and activation of K+ channels, primarily BKCa, leading to calcium influx reduction through CaV.
Similar content being viewed by others


Background
Natural products have played an important role in the process of discovery and drug development [1]. These products and their derivatives represent more than 50% of all drugs in clinical use worldwide, and plants contribute no less than 25% of this total [2]. In this context, several natural products have been isolated from species of the Fabaceae family, including benzofuranoids, essential oils, triterpenoids, alkaloids and flavonoids [3].
This family includes a plant found in the northeastern Brazilian province of Caatinga, the species Piptadenia stipulacea (Benth.) Ducke, popularly known as “Jurema-branca” [4], “Jurema-malícia-da-serra”, “Caracará” and “Calumbi” [5]. This species is widely used in folk medicine for treatment of inflammatory processes, being consumed as a decoction or tincture of its barks and leaves [6].
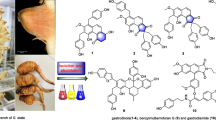
Flavonoids are low molecular weight phenolic compounds, secondary metabolites found in plants [7]. A flavonoid named galetin 3,6-dimethyl ether (FGAL) (Fig. 1) was isolated from the chloroform phase of the crude ethanolic extract obtained from the aerial parts of Piptadenia stipulacea [8]. This flavonoid presented interesting biological activities, such as antiviral [9], anticancer [10] and antioxidant [11]. In addition, the flavonoid presented spasmolytic activity on guinea pig ileum and trachea and rat aorta and uterus [12].

Chemical structure of flavonoid galetin 3,6-dimethyl ether (FGAL) [8]
In recent years and in light of the vital importance of myometrium in physiological processes, there has been remarkable progress in the understanding of uterine pathophysiology, such as embryo implantation and disorders such as dysmenorrhea and colic, which occurs by an uncontrolled uterine contractions process [13]. The exaggerated contractions culminate in pain, even leading to reduced uterine vascular flow, hypoxia and ischemia, further increasing the level of pain [14].
Some flavonoids have presented spasmolytic activity in rat uterus, such as genistein, kaempferol, quercetin [15], and isoliquiritigenin, which also showed analgesic activity in mice [16], and FGAL [12], which also presented anti-inflammatory and anti-nociceptive activities in mice [8].
Therefore, we decided to investigate the tocolytic action mechanism of FGAL on rat uterus given the importance of flavonoids as secondary metabolites produced by plants, in addition to the fact that substances able to exert tocolytic action are promising for treating uterine pain associated with smooth muscle contraction dysregulation, such as uterine colic and dysmenorrhea.
Methods
Animals
Virgin female Wistar rats (40 rats, 150–250 g) were used for all experiments. The animals were maintained under standard conditions with a 12 h light/dark cycle in a temperature-controlled environment (21 ± 1 °C). They had free access to water and food (Purina®, Brazil). All experimental procedures were performed in accordance with the guidelines approved by the Animal Research Ethics Committee (CEPA) of Laboratório de Tecnologia Farmacêutica (LTF)/Universidade Federal da Paraíba (UFPB) (protocol n° 0303/11).
Chemicals
FGAL was obtained as previously described [8]. Briefly, FGAL was isolated from the aerial parts of Piptadenia stipulacea Benth., species collected in the city of Serra Branca, Paraíba, Brazil. The sample were identified by PhD. Maria de Fátima Agra, from UFPB. A voucher, Agra et al. 3331 (JPB) was deposited in the Herbarium Lauro Pires Xavier in the Departamento de Sistemática e Ecologia from UFPB (João Pessoa, PB, Brazil).
Potassium chloride (KCl) and calcium chloride bi-hydrate (CaCl2.2H2O) were purchased from Merck & Co. Inc. (Whitehouse Station, NJ, USA). Apamin, cesium chloride (CsCl), tetraethylammonium chloride (TEA-Cl), glibenclamide, 4-aminopyridine (4-AP) and diethylstilbestrol were purchased from Sigma-Aldrich Co. (Saint Louis, MO, USA). Oxytocin was purchased from Eurofarma (Brasil). All substances were dissolved in distilled water, except glibenclamide and diethylstilbestrol, which were dissolved in ethanol PA (95%). FGAL was solubilized in Cremophor EL® (3%) plus distilled water. The final concentration of Cremophor EL® in the organ bath never exceeded 0.01% (v/v), which does not produce any observable effect on rat uterine tonus.
Tissue preparation and measurement of contractile tension
Rats were pretreated with diethylstilbestrol 1.0 mg/kg (s.c.) 24 h prior to the estrus induction experiment. The animals were sacrificed by cervical dislocation. The rat uterus was immediately removed, cleaned of connective tissue and fat and immersed in Locke-Ringer solution (in mM: NaCl, 154.0; KCl, 5.63; CaCl2.2H2O, 2.16; MgCl2.6H2O, 2.10; glucose, 5.55; and NaHCO3, 5.95) [14] and continuously bubbled with a carbogenic mixture (95% O2 and 5% CO2). A depolarizing Locke-Ringer solution (60 mM KCl), nominally without Ca2+, was also used with KCl in equimolar exchange for NaCl [15]. The segment of uterus was cut longitudinally into strips (1 to 2 cm in length and approximately 1 mm wide). Then, the strips were suspended by cotton thread in organ baths at 32 °C. The isotonic contractions were recorded on a drum of a smoky kymograph using lever, and the isometric contractions were registered by a force transducer TIM-50 (São Paulo) coupled to a data acquisition system (AECAD 04F, AQCAD 2.0.3., AVS Projects, SP). The uterus segments were stabilized with a 1.0 g resting tension (baseline) by at least 40 min. At this time, the solution was changed every 10 min.
Experimental procedures
FGAL effect on KCl- or oxytocin-induced tonic contractions
After the stabilization period, two similar concentration-response curves for 60 mM KCl or 10−2 IU/mL oxytocin (OXY) were obtained, and after the plateau phase of the second contraction, cumulative concentrations of FGAL were added to the organ bath to obtain a relaxation curve. The results were expressed as the reverse percentage of initial contraction elicited by contractile agents. The pEC50 value of FGAL was calculated and compared to both contractile agents.
FGAL effect on oxytocin-induced cumulative contractions
After the stabilization period, two similar cumulative concentration-response curves for OXY (10−5-3 × 10−1 IU/mL) were obtained (control), and then, in the presence of different concentrations of FGAL pre-incubated for 15 min, a third cumulative concentration-response curve for OXY was obtained. The maximal contraction obtained with the control concentration-response curve to OXY was taken as 100%, and all concentration-response curves in the presence of FGAL were calculated as a function of this value. pEC50 value of OXY was calculated in both absence and presence of FGAL concentrations. The antagonism exerted by FGAL was analyzed based on the values of Schild slope [17] and its potency on the pEC50 value.
FGAL effect on CaCl2-induced contractions in depolarizing medium (60 mM KCl) nominally without Ca2+
After the preparations were stabilized, Locke-Ringer solution was replaced by a depolarizing Locke-Ringer solution, nominally without Ca2+. After 30 min, 60 mM KCI was added to produce smooth muscle depolarization and remained in the bath throughout the experiment. Ten min after KCI addition, two similar cumulative concentration-response curves for CaCl2 (3 × 10−6–10−2 M) were obtained (control), and a third cumulative concentration-response curve for CaCl2 was then obtained in the presence of different concentrations of FGAL pre-incubated for 15 min. The Emax obtained with the control concentration-response curve for CaCl2 was taken as 100% and served as the reference for all concentration-response curves assessed in the presence of FGAL [15].
FGAL effect on oxytocin-induced tonic contractions in the absence and presence of K+ channel blockers
In uterine segments, some pharmacological tools were incubated for 20 min before the contraction induced by the contractile agent to investigate the participation of different types of K+ channels, as follows: 5 mM CsCl, non-selective K+ channel blocker [18]; 3 mM 4-AP, voltage-gated K+ channel (Kv) blocker [19]; 3 × 10−5 M glibenclamide, ATP-sensitive K+ channel (KATP) blocker [20]; 100 nM apamine, small-conductance Ca2+-activated K+ channel (SKCa) blocker [21]; or 1 mM TEA+, big-conductance Ca2+-activated K+ channel (BKCa) blocker [22]. After stabilization of the tonic contraction induced by 10−2 IU/mL OXY, cumulative concentrations of FGAL were added to the bath to obtain the relaxation curve. The results were expressed as the reverse percentage of initial contraction elicited by OXY. The relaxation potency of FGAL was evaluated by comparing the pEC50 values in both the absence and the presence of blockers.
FGAL effect on oxytocin-induced tonic contractions in the absence and presence of non-selective phosphodiesterase inhibitor
Aminophylline (10−4 M), a non-selective phosphodiesterase (PDE) inhibitor [23], was added to organ baths for 20 min followed by an OXY-induced tonic contraction, and FGAL was then cumulatively added to obtain the relaxation curve. The results were expressed as the reverse percentage of initial contraction elicited by OXY, and the relaxation potency of FGAL was evaluated by comparing the pEC50 values in both the absence and the presence of aminophylline.
Statistical analysis
All results were expressed as mean ± standard error of mean (SEM). Student’s t-test, for single comparisons and one-way ANOVA followed by Bonferroni’s post-test for multiple comparisons were used in the data analysis, and results were considered significant when P < 0.05. Curves and pEC50 values were calculated by non-linear regression and Schild slope and pEC’50 (negative logarithm to base 10 of molar concentration value of an antagonist that reduces the response to an agonist to 50% of its maximum effect) by linear regression [17]. All analyses were performed using GraphPad Prism® software version 5.01 (GraphPad Software Inc., San Diego, CA, USA).
Results
FGAL effect on KCl- or oxytocin-induced tonic contractions
In a concentration-dependent manner, FGAL (10−9–10−4 M, n = 5) relaxed the uterus pre-contracted with 60 mM KCl (pEC50 = 5.7 ± 0.06) or 10−2 IU/mL oxytocin (pEC50 = 7.0 ± 0.08) (Fig. 2). An analysis of the pEC50 values indicates that FGAL was more potent in inhibiting the contractions induced by oxytocin. After the control experiments, all preparations showed complete reversion of the contractile response within 2 h.

FGAL effect on tonic contractions induced by KCl or oxytocin in rat uterus. FGAL effect on tonic contractions induced by 60 mM KCl (□) or 10−2 IU/mL oxytocin (■) in rat uterus (n = 5). Symbols and vertical bars represent the mean and SEM. Student’s t test, ***P < 0.001 (KCl vs. oxytocin)
FGAL effect on oxytocin-induced cumulative contractions
FGAL 10−6 M did not present effect, however FGAL (3 × 10−6 and 10−5 M, n = 5) inhibited the cumulative concentration-response curves to oxytocin. These curves were shifted to the right in a non-parallel manner, with decreasing Emax (Table 1, Fig. 3). The FGAL pEC50’ was 5.04 ± 0.02, and Schild slope value was 0.75 ± 0.16.

Cumulative-contractions response curves to oxytocin in absence and presence of FGAL in rat uterus. Cumulative-contractions response curves to oxytocin in the absence (■) and presence of FGAL 10−6 (△), 3 × 10−6 (▲) and 10−5 M (◇) in rat uterus (n = 5). Symbols and vertical bars represent the mean and SEM
FGAL effect on CaCl2-induced contractions in depolarizing medium, nominally without Ca2+
In depolarizing Locke-Ringer solution, nominally without Ca2+, (60 mM KCl), FGAL (3 × 10−6, 10−5 and 3 × 10−5 M, n = 5) significantly reduced the CaCl2-induced maximal contractile response and promoted a concentration-dependent rightward shift of CaCl2 concentration-response curves (Table 2, Fig. 4).

Cumulative-contractions response curves to CaCl2 in absence and presence of FGAL in rat uterus. Cumulative-contractions response curves to CaCl2 in depolarizing medium, nominally without Ca2+, in the absence (■) and presence of FGAL 10−6 (□), 3 × 10−6 (●), 10−5 (○) and 3 × 10−5 M (▲) in rat uterus (n = 5). Symbols and vertical bars represent the mean and SEM
FGAL effect on oxytocin-induced tonic contractions in the absence and presence of K+ channels blockers
FGAL (10−9–10−4 M, n = 5) completely relaxed the uterus pre-contracted with 10−2 IU/mL oxytocin (pEC50 = 7.0 ± 0.08), but its relaxant potency was attenuated after pre-incubation with 5 mM CsCl (pEC50 = 5.7 ± 0.01) (Fig. 5a). The relaxant potency of FGAL was attenuated in the presence of specific K+ channel blockers (Fig. 5b): 3 mM 4-AP (pEC50 = 5.5 ± 0.01), 30 μM glibenclamide (pEC50 = 5.3 ± 0.01), 1 mM TEA+ (pEC50 = 5.0 ± 0.009) and 100 nM apamine (pEC50 = 6.4 ± 0.03).

FGAL effect on tonic contractions induced by oxytocin in absence and presence of K+-channels blockers. FGAL effect on tonic contractions induced by 10−2 IU/mL oxytocin in both absence (■) and presence of CsCl (□) (a), 4-AP (▼), apamin (◇), 1 mM TEA+ (▼) or glibenclamide (○) (b) in rat uterus (n = 5). Symbols and vertical bars represent the mean and SEM, respectively. Student’s t test, ***P < 0.001 (OXY vs. blockers + OXY)
FGAL effect on oxytocin-induced tonic contractions in the absence and presence of aminophylline
FGAL (10−9–10−4 M, n = 5) completely relaxed the uterus pre-contracted with 10−2 IU/mL oxytocin (pEC50 = 7.0 ± 0.08), and its relaxant potency was not modified in the presence of aminophylline (pEC50 = 6.7 ± 0.03) when compared with the control curve (Fig. 6).

FGAL effect on tonic contractions induced by oxytocin in absence and presence of aminophylline. FGAL effect on tonic contractions induced by 10−2 IU/mL oxytocin in absence (■) and presence (◇) of aminophylline in rat uterus. Symbols and vertical bars represent the mean and SEM, respectively
Discussion
This study investigated the mechanism of the spasmolytic effect of galetin 3,6-dimethyl ether on rat uterus, which appears to occur by non-competitive pseudo-irreversible antagonism of oxytocin receptors and downstream pathway modulation, as positive modulation of K+ channels, with greatest specificity for BKCa subtype.
In preliminary pharmacological screening, it has been shown that FGAL inhibited both oxytocin- and carbachol-induced phasic contractions in the rat uterus [12]. Thus, we decided to investigate whether the flavonoid would relax the organ pre-contracted by 60 mM KCl, which induces contraction due to electromechanical mechanism and 10−2 IU/mL OXY, a pharmaco-mechanical coupling agent. In a concentration-dependent manner, FGAL relaxed rat uterus pre-contracted by both oxytocin and KCl, but not equipotently, being more potent in relaxing uterus pre-contracted with oxytocin.
Because FGAL was more potent in inhibiting the contractions induced with oxytocin than with carbachol [12] and because it relaxed the rat uterus pre-contracted with this agonist more potently than with KCl, a cumulative concentration-response curves to this agonist were obtained to evaluate a possible antagonism of these receptors. FGAL inhibited oxytocin-cumulative curves, and these were shifted to the right, in a non-parallel manner, with a reduction and abolition of Emax, excluding a competitive antagonism. In addition, the non-competitive antagonism was confirmed, as the slope value (0.75 ± 0.16) was different from the unit [17].
Non-competitive antagonism occurs when the antagonist binds to the same agonist binding site (pseudo-irreversible antagonism) or in a separate site (allosteric antagonism) [24]. In addition, pseudo-irreversible antagonism occurs with a slow dissociation of the drug from the receptor, leading to a prolonged effect and abolishing the maximum effect of the agonist, differing from those observed in the allosteric antagonism, in which the maximum effect is not suppressed [25]. Thus, the profile of the cumulative concentration-response curves to oxytocin represents a pseudo-irreversible antagonism because the inhibition of the contractile response to oxytocin in the presence of FGAL has not reached a limiting value, a characteristic observed in allosteric antagonism. Additionally, it is not excluded the modulation of downstream pathway, as voltage-gated Ca2+ channels or K+ channels.
In rat uterus, extracellular and intracellular Ca2+ sources are important for muscular tonus development, and the Ca2+ entry in smooth muscle cells occurs primarily through voltage-gated Ca2+ channels (CaV) [26]. Because the opening of these channels is a common step in the oxytocin and KCl signaling pathways to maintain the tonic phase of contraction [27] and because FGAL relaxed the rat uterus pre-contracted by both contractile agents, it was hypothesized that FGAL could be acting by blocking the Ca2+ influx through CaV. Therefore, cumulative contractions were induced with CaCl2 in depolarizing medium, nominally without Ca2+ , in both the absence and the presence of different concentrations of FGAL. The flavonoid antagonized the CaCl2-induced contractions, as evidenced by the shift of the control curve to the right, in a non-parallel manner, with reduced Emax, indicating that the flavonoid inhibits Ca2+ influx through the CaV.
The K+ channel plays a key role in membrane potential regulation and can modulate the openness of CaV, regulating the Ca2+ influx in smooth muscle cells [28]. Substances able to open K+ channels, such as cromacalin and pinacidil, comprise a diverse group of molecules with therapeutic potential by preventing the cell excitation [29]. These compounds open K+ channels, resulting in hyperpolarization by increasing K+ efflux, and consequently reduce the cytosolic concentration of Ca2+ ([Ca2+]c), followed by smooth muscle relaxation [30, 31]. Hence, we decided to investigate the possible positive modulation of these channels using the CsCl, a non-selective K+ channel blocker as pharmacological tool [18]. The relaxant potency of FGAL was reduced approximately 22-fold in the presence of CsCl, confirming the positive modulation of K+ channels by FGAL.
In uterine smooth muscle, the most abundant and most studied subtypes of K+ channels are BKCa, SKCa, KV and KATP [32]. Accordingly, we decided to investigate the involvement of these subtypes of K+ channels in the tocolytic action of FGAL using selective blockers. The relaxation curve induced by FGAL was shifted to the right in the presence of 4-aminopyridine (36-fold), a selective KV blocker [19], glibenclamide (50-fold), a selective KATP blocker [20], apamin (4-fold), a selective SKCa blocker [21], and 1 mM TEA+ (100-fold), a BKCa blocker [22]. These results indicate that these subtypes of K+ channels are involved in the FGAL tocolytic mechanism of action on the rat uterus. Moreover, in the presence of 1 mM TEA+, the relaxant potency of FGAL was more reduced than in the presence of the other blockers, indicating that FGAL primarily positively modulates BKCa, which is important because it has been described that uterine smooth muscle presents greater expression of these channels compared with the other subtypes and that they play a fundamental role in regulating the basal tone of this organ [33].
Another important pathway involved in smooth muscle relaxation is the cyclic nucleotide phosphodiesterases (PDEs) pathway. Cyclic monophosphates of adenosine and guanosine (cAMP and cGMP, respectively) are able to activate protein kinase A (PKA) and protein kinase G (PKG), respectively, when occurs an elevation in the concentration of cAMP and cGMP in the cytosolic medium. These kinases can phosphorylate several intracellular targets, leading to smooth muscle relaxation, and one of these targets are K+ channels. PDE is responsible for cAMP and cGMP hydrolysis, resulting in their inactive products 5′-AMP and 5′-GMP, respectively, which do not activate PKA and PKG, thus stopping the cell signaling mechanism dependent on cyclic nucleotides [23]. Substances that raise the intracellular content of cAMP or cGMP show a potential relaxant effect through PDE inhibition in different tissues, including uterine smooth muscle [23].
Thus, to evaluate the involvement of this pathway, aminophylline, a non-selective PDE inhibitor [23], was used as a pharmacologic tool. The FGAL relaxant potency was not altered in both the absence and the presence of aminophylline, suggesting that the cyclic nucleotide PDEs pathway is not involved on relaxing effect induced by FGAL.
Conclusion
In conclusion, the FGAL tocolytic action mechanism in rat uterus involves a non-competitive pseudo-irreversible antagonism of oxytocin receptors and a positive modulation of K+ channels, primarily the BKCa channels subtype, which indirectly modulates the CaV, leading to a reduction in Ca2+ influx and uterine smooth muscle relaxation. Thereby, FGAL appears promising for the treatment of disorders affecting the uterine smooth muscle, such as the pain caused by colic and dysmenorrhea. However, more research is needed to better elucidate the FGAL tocolytic action mechanism in the rat uterus, such as molecular investigation of ion flow through the channels modulated by FGAL and the effect of FGAL on cytosolic calcium concentration of uterus myocytes.
Abbreviations
- BKCa :
-
Big-conductance Ca2+-activated K+ channel
- FGAL:
-
Flavonoid galetin 3,6 dimethyl ether
- KATP :
-
ATP-sensitive K+ channel
- OXY:
-
Oxytocin
- PDEs:
-
Phosphodiesterases
- SKCa :
-
Small-conductance Ca2+-activated K+ channel
References
Newman DJ, Cragg GM. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J Nat Prod. 2012;75:311–35.
Gurib-Fakim A. Medicinal plants: traditions of yesterday and drugs of tomorrow. Mol Asp Med. 2006;27:1–93.
Melo AC, Cota BB, Oliveira AB, Braga FC. HPLC quantitation of kaurane diterpene in Xylopia species. Fitoterapia. 2001;72:40–5.
Fabricante JR, Andrade LA. Análise estrutural de um remanescente de caatinga no Seridó Paraibano. Oecol Bras. 2007;11:341–9.
Florentino ATN, Araújo EL, Albuquerque UP. Contribuição de quintais agroflorestais na conservação de plantas da Caatinga, Município de Caruaru, PE, Brasil. Acta Bot Bras. 2007;21:37–47.
Albuquerque UP, Andrade LHC. Uso de recursos vegetais da Caatinga: o caso do agreste do estado de Pernambuco (Nordeste do Brasil). Interciência. 2002;27:336–46.
Middleton-Junior E, Kandaswami C, Theoharides TC. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacol Rev. 2000;52:673–751.
Queiroz AC, Lira DP, Dias TLMF, Souza ET, Matta CB, Aquino AB, Silva LHAC, Silva DJC, Mella EAC, Agra MF, Filho JMB, Araújo-Júnior JX, Santos BVO, Alexandre-Moreira MS. The antinociceptive and anti-inflammatory activities of Piptadenia stipulacea Benth. (Fabaceae). J Ethnopharmacol. 2010;128:377–83.
Elsohly HN, El-Feraly FS, Joshi AS, Walker LA. Antiviral flavonoids from Alkanna orientalis. Planta Med. 1997;63:384.
Rosenbloom RA. Compositions and methods for the treatment of leukemia, Patent Application Publication; 2006. p. 10.
Leach D, Stevenson L, Gabriel B, Beattie K. Biologically active compounds isolated from Centipeda and their biological activity as antioxidant and anti-inflammatory agents, PCT Patent Filing; 2007. p. 49.
Macêdo CL, Vasconcelos LHC, Correia ACC, Martins IRR, Lira DP, Santos BVO, Silva BA. Spasmolytic effect of galetin 3,6 dimethyl-ether, flavonoid obtained from Piptadenia stipulacea (Benth.) Ducke. J Smooth Muscle Res. 2011;47:123–34.
Aguilar H, Micthell B. Physiological pathways and molecular mechanisms regulating uterine contractily. Hum Reprod Update. 2010;16:725–44.
Bortoleto CCR. Dismenorreia primária, etiopatogenia e terapêutica. Femina. 1995;23:439–43.
Revuelta MP, Cantabrana B, Hidalgo A. Depolarization-dependent effect of flavonoids in rat uterine smooth muscle contraction elicited by CaCI2. Gen Pharmacol. 1997;29:847–57.
Shi Y, Wu D, Sun Z, Yang J, Chai H, Tang L, Guo Y. Analgesic and uterine relaxant effects of isoliquiritigenin, a flavone from Glycyrrhiza Glabra. Phytother Res. 2012;26:1410–7.
Arunlakshana O, Schild HO. Some quantitative uses of drug antagonists. Br J Pharmacol. 1959;14:48–58.
Latorre E, Oberhauser A, Labarca P, Alvarez O. Variety of calcium-activated potassium channels. Annu Rev Physiol. 1989;51:385–99.
Aaronson PI, Sarwar U, Gin S, Rockenbauch U, Connolly M, Tillet A, Watson S, Liu B, Tribe RM. A role for voltage-gated, but not Ca2+-activated, K+ channels in regulating spontaneous contractile activity in myometrium from virgin and pregnant rats. Br J Pharmacol. 2006;147:815–24.
Hughest SJ, Hollingsworth M. Relaxin as a relaxant of the isolated rat uterus: comparison with its mechanism of action in vivo. Gen Pharmacol. 1997;29:829–33.
Tsai ML, Cummings KC, Webb RC, Caruso RL. Acute inhibition of spontaneous uterine contractions by an estrogenic polychlorinated biphenyl is associated with disruption of gap junctional communication. Toxicol Appl Pharmacol. 1998;152:18–29.
Jmari K, Mironneau C, Mironneau J. Inactivation of calcium channel current in rat uterine smooth muscle: evidence for calcium- and voltage-mediated mechanisms. J Physiol. 1986;380:111–26.
Lugnier C. Cyclic nucleotide phosphodiesterase (PDE) superfamily: a new target for the development of specific therapeutic agents. Pharmacol Ther. 2006;109:366–98.
Blumenthal DK, Garrison JC. Pharmacodynamics: molecular mechanisms of drug action. In: Brunton LL, Chabner BA, Knollmann BC, editors. Goodman and Gilman's the pharmacological basis of therapeutics. San Diego: The McGraw-Hill Companies; 2011. p. 41–71.
May LT, Leach K, Sexton PM, Christopoulos A. Allosteric modulation of G protein-coupled receptors. Ann Rev Pharmacol Toxicol. 2007;47:1–51.
Catterall WA. Structure and regulation of voltage-gated Ca2+ channels. Annu Rev Cell Dev Biol. 2000;16:521–55.
Wray S. Uterine contraction and physiological mechanisms of modulation. Am J Phys. 1993;264:C1–18.
Knot HT, Brayden EJ, Nelson MT. Calcium channels and potassium channels. In: Bárány M, editor. Biochemistry of smooth muscle contraction. San Diego: Academic Press; 1996. p. 203–19.
Quest U, Cook NS. Moving together: K+-channel openers and ATP-sensitive K+-channels. Trends Pharmacol Sci. 1989;10:431–5.
Weston AH, Edwards G. Recent progress in potassium channel opener pharmacology. Biochem Pharmacol. 1992;43:47–54.
Brainard AM, Korovkina VP, England SK. Potassium channels and uterine function. Semin Cell Dev Biol. 2007;18:332–9.
Lorca RA, Prabagaran M, England SK. Functional insights into modulation of BKCa channel activity to alter myometrial contractility. Front Physiol. 2014;31:1–12.
Bender AT, Beavo JA. Cyclic nucleotide phosphodiesterases: molecular regulation to clinical use. Pharmacol Rev. 2006;58:488–520.
Acknowledgements
The authors thank CAPES and CNPq for financial support and José Crispim Duarte for providing technical assistance.
Funding
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Availability of data and materials
Availability of data: no available.
Sample of FGAL: no available.
Author information
Authors and Affiliations
Contributions
CJN is the author who mainly contributed to this research, performing literature search, pharmacological experiments, analysis of the data and writing the manuscript. SILL, PJC, VLHC and TRA were involved in acquisition, interpretation and analysis of functional pharmacological experiments. The PhD SBVO performed the phytochemical isolation of the flavonoid studied. The PhD SBA was involved in design, interpretation of the data and review of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
All experimental procedures were performed in accordance with the guidelines approved by the Animal Research Ethics Committee (CEPA) of Laboratório de Tecnologia Farmacêutica (LTF)/Universidade Federal da Paraíba (UFPB) (protocol n° 0303/11).
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Carreiro, J., Souza, I.L.L., Pereira, J.C. et al. Tocolytic action and underlying mechanism of galetin 3,6-dimethyl ether on rat uterus. BMC Complement Altern Med 17, 514 (2017). https://doi.org/10.1186/s12906-017-2007-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-017-2007-6