
Abstract
Background
In aging, muscle stiffness is considered as one of the factors associated with the reduction of force generation capability. There have been inconsistent findings on age-related alteration in the passive stiffness of quadriceps muscle in the female adults. Thus, the aim of this study was to determine the effect of aging on the shear moduli of the superficial muscle heads of the quadriceps and to explore its relationship with knee extension force.
Methods
Passive shear moduli of the rectus femoris (RF), vastus lateralis (VL), and vastus medialis (VM) were measured at rest using shear wave elastography in 20 young and 20 senior female adults. Measurements were repeated at four knee joint positions, that is, 30°, 60°, 90°, and 105° of knee flexion. Maximal isometric voluntary knee extension force was assessed at 30°, 60°, and 90° of knee flexion.
Results
As per our findings, senior adults were determined to have significantly higher passive muscle shear moduli in the RF (by 34% – 68%; all p < 0.05) and the VL muscle heads (by 13%–16%, all p < 0.05) at and beyond 60° of knee flexion. Age-related increase in the VM was evident at 105° knee flexion (by11%, p = 0.020). The RF shear modulus was negatively correlated to the maximal isometric voluntary contraction force measured at 60° (r = − 0.485, p = 0.030) in senior adults.
Conclusions
Senior female adults had greater passive stiffness at the superficial muscle heads of the quadriceps muscles when measured at long muscle length. Among the senior female adults, the passive stiffness of RF has been determined to have a negative association with the knee extensor force only at 60° knee flexion. No significant association was noted for other angles and muscles.
Similar content being viewed by others
Background
In aging, a decrease in muscle force has been assumed to be mainly induced by muscle atrophy [1, 2]. However, muscle atrophy cannot completely account for the age-induced force decrease as the force decrease (40%) has been noted to be often larger than the atrophy sign (23%) [3, 4]. The change of microstructure in a single muscle fiber, such as decreased myosin protein, slowing of myosin-actin cross-bridge in single fiber [5], reduced cross-sectional area of myosin heavy chain II fibers, and decrease in intermyofibrillar mitochondrial size with aging [6], might have contributed to the age-related muscle force decrease. In addition, muscle properties like passive muscle stiffness could also influence muscle force [7, 8] as well as muscle performance in the elderly [9]. A numerical study has demonstrated a causal association between the age-related increase in passive muscle stiffness and the reduced force generation capability with aging [10].
However, the finding that age is related to alteration of muscle stiffness is not always a consistent result. Previous animal studies have revealed higher stiffness of the aged soleus muscle [11, 12]. Similarly, the single fiber of isolated vastus lateralis (VL) muscle from an older person was noted to have greater passive stiffness compared to a younger one [13, 14]. Using qualitative ultrasound elastography in vivo, a greater muscle stiffness in biceps brachialis muscle, rectus femoris (RF), and gastrocnemius muscles was observed in elderly people [15, 16]. However, contrary results have been reported as well. For example, lower muscle stiffness was observed in the four heads of quadriceps muscle [17, 18], and comparable level of stiffness in several leg muscles has been reported in aged participants [19,20,21,22]. According to a previous study [16], age-related modulation in muscle stiffness was often influenced by muscle length, which was dependent on joint angles in vivo. It further revealed that aged muscle was stiffer than young muscle at long muscle length. However, how the joint position affects muscle stiffness was not considered in most reported studies. When the muscle length was taken into account, the muscle stiffness of triceps surae muscle in old participants was equal to that of young participants at 0° and 15° ankle dorsiflexion [23] and less than the young ones at maximal dorsiflexion [24]. Therefore, the length-dependent modulation of age on muscle stiffness has not been clarified as a whole muscle in vivo.
When the stiffness of quadriceps femoris muscle was measured at short muscle length, its relationship with extension knee force was not observed [15]. Similar result was found in medial gastrocnemius as well [25]. It should be noted that muscle stiffness was measured only at a given muscle length in these studies. As muscle stiffness was length dependent [26], it is thus necessary to explore the association between muscle stiffness and muscle force generation capacity at different muscle lengths.
In this study, our primary aim was to determine the effect of aging on the shear moduli of the superficial muscle heads of the quadriceps. Second aim was to explore the relationship between muscle shear moduli of the superficial heads of the quadriceps and knee extension force in the senior female adults. The hypotheses were senior female adults would present with greater passive muscle stiffness at long muscle length, and passive muscle passive stiffness would be negatively associated with muscle force.
Methods
In total, 20 senior healthy females (age range: 50–70 years) and 20 young healthy females (age range: 20–32 years) were included in this study. The participants’ demographic information is summarized in Table 1. All participants were free of knee pain or injury and have no history of knee surgery. They did not engage in any regular exercise training or high-intensity sport activities.
Passive muscle shear modulus measurements
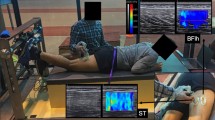
The passive muscle stiffness was first measured. The participants were positioned supine on the plinth of the isokinetic dynamometer (Cybex, Medway, MA, USA) with hip at 0° and neutral rotation, as the pelvis, trunk, and thigh were stabilized with straps. The knee joint rotation center (lateral femoris condyle) was aligned with the rotation center of the isokinetic dynamometer. Measurements were taken with the knee joint positioned at 30º, 60º, 90º, and 105º of knee flexion (0° being the knee extended) in sequence in order to avoid the stretching effect from muscle length (Fig. 1). The knee was then passively positioned at each angle and remained for 30 s before measurement started. Participants were instructed to remain relax during the whole procedure.

The participant position during measurements. A typical image of muscle shear modulus of vastus medialis (VM) muscle is presented
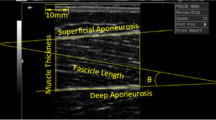
The muscle shear modulus (an index for muscle stiffness) of the dominant leg was assessed using an ultrasound scanner coupled with a 4–15 MHz linear transducer (AixplorerV4.0; Supersonic Imagine, Aix-en-Provence, France). The elastography machine was then set to the musculoskeletal preset. A well-trained examiner in musculoskeletal ultrasound examination performed all the measurements. The following three superficial heads of the quadriceps femoris muscle were then measured: RF, VL, and vastus medialis (VM). The measurement locations were marked on the skin with a waterproof pen before the test. For VM and VL, the transducer was placed 1/5 and 1/3 of the distance from the midpoint of medial and lateral patella border to the anterior superior iliac spine, respectively. For RF, the transducer was placed 1/2 of the distance from anterior superior iliac spine to the midpoint of the superior tip of the patella [4, 27]. The transducer was held perpendicularly to the skin with plentiful gel, and slight pressure was exerted on the skin. The transverse images were obtained to confirm the right muscle was measured; then, the longitudinal images were captured at one sample per second which was the temporal resolution of the current shear wave elastography version, and the spatial resolution was 1 × 1 mm. The probe was aligned with either the muscle fiber direction (VL and VM) or the muscle shortening direction (RF). Five continuous elastography maps were acquired in 5 s. During each measurement, the square color box was adjusted accordingly depending on the muscle thickness.
Maximal isometric voluntary contraction (MIVC) test
After elastography measurements, MIVC of knee extension assessment was performed. The position of participants was the same as that of the passive stiffness measurement. Prior to measurement, the participants were instructed to perform two submaximal contractions for warm-up and to be familiar with the test procedure. Each participant conducted three 5-s maximal isometric voluntary contractions, with a 2-min interval between each contraction. The MIVC was assessed at three knee angles (30°, 60°, and 90° knee flexion; 0° = full extension). Due to the discomfort of the joint, the MIVC at 105° was not measured. The maximum peak torque of the three contractions captured at each angle was considered as MIVC torque at this angle.
Data reduction and analysis
The shear wave elastography data were exported in mp4 format and sequenced in png. Image processing was performed using a custom Matlab script (MathWorks, Natick, MA). Each image was then carefully inspected for artifacts. If artifacts were present in any image, the region of interest was reduced in size to remove the artifact. The colored 2D maps were converted into shear modulus values, and the shear modulus values were averaged over the map and over the five images for further analysis.
Statistical analysis was conducted using SPSS 21.0 software package (New York, USA). Independent sample t-test was performed to compare the difference of basic demographic information and MIVC between young and senior adults. The MIVC comparison between the two age groups was analyzed via a two-way repeated measures ANOVA with knee angle as within-subject factors and group (young and senior adults) as between-subject factors. A three-way repeated measures ANOVA with muscle heads and knee angle as within-subject factors and group (young and senior adults) as between-subject factors was used to compare the shear moduli of the two groups. Post hoc analyses were performed when appropriate using the Bonferroni method. The correlations between shear modulus and the MIVC of quadriceps femoris muscle were calculated using Pearson’s coefficient if the data were normally distributed; otherwise, Spearman correlation coefficients were calculated. Statistical significance level of p-value was set to be less than 0.05.
Results
The shear modulus values of the three muscles are presented in Fig. 2. A significant main effect was noted in terms of group, muscle, and angle; a significant interaction between muscle, angle, and group was also observed (all p-values < 0.001). Post hoc analysis revealed that the muscle shear moduli of RF and VL were significantly higher in the senior than the young adults when the knee was positioned at 60°, 90°, and 105° of knee flexion (all p-values < 0.006 for RF and < 0.041 for VL). Specifically, at 60°knee flexion, the shear moduli were 34% (p = 0.006) and 16% (p = 0.031) greater in the senior than the young adults for RF and VL, respectively. At 90° of knee flexion, the shear moduli were 56% (p < 0.001) and 13% (p = 0.031) greater in the senior adults for RF and VL, respectively. At 105° of knee flexion, the values reached to 68% (p < 0.001) and 13% (p = 0.041) for RF and VL, respectively. When considering the VM muscle, significant greater shear modulus of the senior adults was only observed at 105° of knee flexion, which was 11% greater than young adults (p = 0.020).

The shear moduli of VM, RF, and VL of the young and senior adults at different knee angles. Compared to young adults, the senior adults exhibited significantly greater shear moduli when assessed over 60° knee flexion for the RF and VL muscles and at 105° of knee flexion for the VM. *: p < 0.05, **: p < 0.01. VM: vastus medialis; RF: rectus femoris; VL: vastus lateralis
The MIVC of the senior adults was significantly lower than young adults at 60° and 90° of knee flexion (Table 2). The correlations between passive shear modulus and MIVC are depicted in Table 3 and Fig. 3. As regards the association between the shear modulus and muscle force, we have only detected moderate and negative association between the passive RF shear modulus at 60° of knee flexion and knee extension force in the senior adults (r = − 0.485, p = 0.030). In addition, there was a trend for a significant correlation in the young adults (r = − 0.395, p = 0.085). However, no significant association was found for other conditions both in the senior and young adults.

Relationship between passive shear modulus of the rectus femoris muscle and the maximal isometric voluntary contraction torque at 60° of knee flexion in senior adults
Discussion
In this study, we aimed to assess age-related changes in muscle stiffness in the female senior adults. We have chosen the quadriceps muscle for analysis because of its importance in daily function. Findings from this study were in accordance with our hypothesis that the senior adults had higher muscle stiffness at long muscle length. More importantly, RF muscle stiffness was negatively associated knee extension force.
In this study, we assessed passive muscle stiffness of the superficial heads of the quadriceps muscle at its shortened to lengthened positions by varying the knee positions. The senior female adults exhibited greater muscle stiffness at the RF and VL muscle heads when the muscles were positioned over 60° of knee flexion (knee extension = 0°) and at the VM muscle at a more lengthened position (105° of knee flexion). This present result is consistent with previous studies reporting higher passive stiffness of aged muscles at long muscle length in either animals [11, 28] or humans [16]. However, the difference was not observed when assessed at shortened muscle length [11, 16, 22]. In this connection, it is well known that change in muscle length affects passive muscle stiffness [26, 29, 30]. The muscle fiber will become taut and even reach to a cross-bridges detached length when the muscle is stretched beyond its slack length which is just over 40° of knee flexion for quadriceps muscle [26, 31].
The underlying mechanisms responsible for the observed increased passive stiffness in the senior female adults could be multifactorial. These include the increased extracellular matrix with aging, which, in turn, increases the muscle stiffness nearly three times in aged quadriceps femoris muscle [32, 33], the decrease of the amount of highly compliant element in the extracellular matrix such as elastin and collagen type III [28], as well as the changes in muscle fiber type and the declined level of estrogen in senior female adults. It has been reported that the area and the percentage of type I muscle fibers increase with age [34]. In response to passive stretch, the type I muscle fibers in isolated VL muscle exhibit greater passive stiffness than type II fibers [35]. The aged muscles with greater amount of type I muscle fibers might display greater passive stiffness than young muscles. Estrogen deficiency might be another factor that reduces passive muscle stiffness in females by means of mediating collagen content within the muscle [36]. Therefore, estrogen deficiency in senior female adults, in particular, after menopause, might contribute to the increase in passive muscle stiffness.
However, age-related increase in passive stiffness of VM was not identically observed as RF and VL over 60° knee flexion. The possible reason could be the imbalanced reduction in muscle mass with aging. It has been reported that the volume of VM was the least affected when it comes to aging among the four heads of quadriceps femoris muscle [37]. The loss of muscle mass with aging would then be replaced by fat and connective tissue, which, in turn, could induce increase in passive muscle stiffness [8]. The relative preservation of VM volume might be one of the reasons for the less increased stiffness at 60° and 90° knee flexion.
We have also detected a negative relationship between passive stiffness of RF and the maximal isometric extension force in senior adults. More specifically, individuals with stiffer RF would produce lower knee extensor force when measured at 60° of knee flexion. In older adults (mean age of 73), no association between the RF stiffness and quadriceps extension strength was observed [15].The authors used strain ratio as an index of passive muscle stiffness. The strain ratio was computed from the strain of the muscle to that of an acoustic coupler. More importantly, the strain of the muscle was measured at its shortened length (knee in extension). Similarly, we could not delineate a significant relationship between muscle stiffness of RF when measured at 30° of flexion and knee extension force.
As muscles with higher echo intensity displayed lower muscle shear modulus and greater muscle strength [38], and echo intensity of RF was negatively associated with maximal isometric force at 60° of knee flexion and functional performance [39, 40], it might be possible that muscle stiffness of RF is associated with knee extension force. However, further investigation is required to understand the underlying mechanism of this correlation.
This study had limitations. First, the results of this present study should be interpreted in regard to the characteristics of our population, i.e., females aged between 50 and 70 years old. This age group was examined because of its association with muscle atrophy that almost apparently occurred in all the skeletal muscles by 50 years old [41]. It is likely that the effect of aging on increased passive muscle stiffness would be more noticeable in more advanced age (> 70) as muscle shear modulus increased with age in person aged more than 60 years old [16], but it remains to be investigated. Second, only females were examined in this study. Thus, in the future, studies examining the age-related alteration on muscle stiffness in males and its effect on muscle force should be conducted. Another limitation is that a rest time of 30 s between passive shear modulus assessments at each angle might be inadequate for stable muscle stiffness after position change due to stress-relaxation effect. In addition, we measured active muscle torque from all the knee extensor groups, but only measuring stiffness from a single/superficial muscle group. Also, the deep quadriceps femoris muscle head (vastus intermedius) was not examined in this study due to its deep position.
Conclusions
This present study has demonstrated that senior females had greater passive stiffness at the superficial heads of the quadriceps muscle when measured at long muscle length. In the senior female adults, the passive stiffness of RF muscle was found to be negatively associated with the knee extensor force only at 60° knee flexion. No significant association was found for other angles and muscles. These findings highlight how aging affects muscle stiffness of the quadriceps muscle and possible strategies to keep muscle force for senior female adults.
Availability of data and materials
All data generated or analyzed in this study are included in this published article.
Abbreviations
- MIVC:
-
Maximal isometric voluntary contraction
- RF:
-
Rectus femoris
- VL:
-
Vastus lateralis
- VM:
-
Vastus medialis
References
Candow DG, Chilibeck PD. Differences in size, strength, and power of upper and lower body muscle groups in young and older men. J Gerontol A Biol Sci Med Sci. 2005;60(2):148–56.
Landers KA, Hunter GR, Wetzstein CJ, Bamman MM, Weinsier RL. The interrelationship among muscle mass, strength, and the ability to perform physical tasks of daily living in younger and older women. J Gerontol A Biol Sci Med Sci. 2001;56(10):B443-448.
Frontera WR, Suh D, Krivickas LS, Hughes VA, Goldstein R, Roubenoff R. Skeletal muscle fiber quality in older men and women. Am J Physiol Cell Physiol. 2000;279(3):C611-618.
Ditroilo M, Cully L, Boreham CA, De Vito G. Assessment of musculo-articular and muscle stiffness in young and older men. Muscle Nerve. 2012;46(4):559–65.
Miller MS, Callahan DM, Toth MJ. Skeletal muscle myofilament adaptations to aging, disease, and disuse and their effects on whole muscle performance in older adult humans. Front Physiol. 2014;5:369.
Callahan DM, Bedrin NG, Subramanian M, Berking J, Ades PA, Toth MJ, et al. Age-related structural alterations in human skeletal muscle fibers and mitochondria are sex specific: relationship to single-fiber function. J Appl Physiol (1985). 2014;116(12):1582–92.
Muraki S, Fukumoto K, Fukuda O. Prediction of the muscle strength by the muscle thickness and hardness using ultrasound muscle hardness meter. Springerplus. 2013;2:457.
Gajdosik RL. Passive extensibility of skeletal muscle: review of the literature with clinical implications. Clin Biomech (Bristol, Avon). 2001;16(2):87–101.
Marcucci L, Reggiani C. Increase of resting muscle stiffness, a less considered component of age-related skeletal muscle impairment. Eur J Transl Myol. 2020;30(2):8982.
Zhang Y, Chen JS, He Q, He X, Basava RR, Hodgson J, et al. Microstructural analysis of skeletal muscle force generation during aging. Int J Numer Method Biomed Eng. 2020;36(1):e3295.
Rosant C, Nagel MD, Perot C. Aging affects passive stiffness and spindle function of the rat soleus muscle. Exp Gerontol. 2007;42(4):301–8.
Alnaqeeb MA, Al Zaid NS, Goldspink G. Connective tissue changes and physical properties of developing and ageing skeletal muscle. J Anat. 1984;139(4):677–89.
Noonan AM, Mazara N, Zwambag DP, Weersink E, Power GA, Brown SHM. Age-related changes in human single muscle fibre passive elastic properties are sarcomere length dependent. Exp Gerontol. 2020;137:110968.
Lim JY, Choi SJ, Widrick JJ, Phillips EM, Frontera WR. Passive force and viscoelastic properties of single fibers in human aging muscles. Eur J Appl Physiol. 2019;119(10):2339–48.
Saito A, Wakasa M, Kimoto M, Ishikawa T, Tsugaruya M, Kume Y, et al. Age-related changes in muscle elasticity and thickness of the lower extremities are associated with physical functions among community-dwelling older women. Geriatr Gerontol Int. 2019;19(1):61–5.
Eby SF, Cloud BA, Brandenburg JE, Giambini H, Song P, Chen S, et al. Shear wave elastography of passive skeletal muscle stiffness: influences of sex and age throughout adulthood. Clin Biomech (Bristol, Avon). 2015;30(1):22–7.
Akagi R, Yamashita Y, Ueyasu Y. Age-related differences in muscle shear moduli in the lower extremity. Ultrasound Med Biol. 2015;41(11):2906–12.
Alfuraih AM, Tan AL, O’Connor P, Emery P, Wakefield RJ. The effect of ageing on shear wave elastography muscle stiffness in adults. Aging Clin Exp Res. 2019;31(12):1755–63.
Wang CZ, Li TJ, Zheng YP. Shear modulus estimation on vastus intermedius of elderly and young females over the entire range of isometric contraction. PLoS One. 2014;9(7):e101769.
Arda K, Ciledag N, Aktas E, Aribas BK, Kose K. Quantitative assessment of normal soft-tissue elasticity using shear-wave ultrasound elastography. Am J Roentgenol. 2011;197(3):532–6.
Domire ZJ, McCullough MB, Chen Q, An KN. Feasibility of using magnetic resonance elastography to study the effect of aging on shear modulus of skeletal muscle. J Appl Biomech. 2009;25(1):93–7.
Debernard L, Robert L, Charleux F, Bensamoun SF. Analysis of thigh muscle stiffness from childhood to adulthood using magnetic resonance elastography (MRE) technique. Clin Biomech (Bristol, Avon). 2011;26(8):836–40.
Lindemann I, Coombes BK, Tucker K, Hug F, Dick TJM. Age-related differences in gastrocnemii muscles and Achilles tendon mechanical properties in vivo. J Biomech. 2020;112:110067.
Hirata K, Yamadera R, Akagi R. Can static stretching reduce stiffness of the triceps surae in older men? Med Sci Sports Exerc. 2020;52(3):673–9.
Ando R, Suzuki Y. Positive relationship between passive muscle stiffness and rapid force production. Hum Mov Sci. 2019;66:285–91.
Xu J, Hug F, Fu SN. Stiffness of individual quadriceps muscle assessed using ultrasound shear wave elastography during passive stretching. J Sport Health Sci. 2018;7(2):245–9.
Giles LS, Webster KE, McClelland JA, Cook J. Can ultrasound measurements of muscle thickness be used to measure the size of individual quadriceps muscles in people with patellofemoral pain? Phys Ther Sport. 2015;16(1):45–52.
Stearns-Reider KM, D’Amore A, Beezhold K, Rothrauff B, Cavalli L, Wagner WR, et al. Aging of the skeletal muscle extracellular matrix drives a stem cell fibrogenic conversion. Aging Cell. 2017;16(3):518–28.
Hug F, Lacourpaille L, Maisetti O, Nordez A. Slack length of gastrocnemius medialis and Achilles tendon occurs at different ankle angles. J Biomech. 2013;46(14):2534–8.
Koo TK, Guo JY, Cohen JH, Parker KJ. Quantifying the passive stretching response of human tibialis anterior muscle using shear wave elastography. Clin Biomech. 2014;29(1):33–9.
Proske U, Morgan DL. Do cross-bridges contribute to the tension during stretch of passive muscle? J Muscle Res Cell Motil. 1999;20(5):433–42.
Lacraz G, Rouleau AJ, Couture V, Söllrald T, Drouin G, Veillette N, et al. Increased stiffness in aged skeletal muscle impairs muscle progenitor cell proliferative activity. PLoS One. 2015;10(8):e0136217.
Wood LK, Kayupov E, Gumucio JP, Mendias CL, Claflin DR, Brooks SV. Intrinsic stiffness of extracellular matrix increases with age in skeletal muscles of mice. J Appl Physiol (1985). 2014;117(4):363–9.
Lee WS, Cheung WH, Qin L, Tang N, Leung KS. Age-associated decrease of type IIA/B human skeletal muscle fibers. Clin Orthop Relat Res. 2006;450(450):231–7.
Noonan A, Zwambag D, Mazara N, Weersink E, Power GA, Brown SHM. Fiber type and size as sources of variation in human single muscle fiber passive elasticity. J Biomech Eng. 2020;142(8):081008.
Morse CI. Gender differences in the passive stiffness of the human gastrocnemius muscle during stretch. Eur J Appl Physiol. 2011;111(9):2149–54.
Barnouin Y, Butler-Browne G, Voit T, Reversat D, Azzabou N, Leroux G, et al. Manual segmentation of individual muscles of the quadriceps femoris using MRI: a reappraisal. J Magn Reson Imaging. 2014;40(1):239–47.
Bachasson D, Dubois GJR, Allenbach Y, Benveniste O, Hogrel JY. Muscle shear wave elastography in inclusion body myositis: feasibility, reliability and relationships with muscle impairments. Ultrasound Med Biol. 2018;44(7):1423–32.
Mota JA, Stock MS. Rectus femoris echo intensity correlates with muscle strength, but not endurance, in younger and older men. Ultrasound Med Biol. 2017;43(8):1651–7.
Rech A, Radaelli R, Goltz FR, da Rosa LH, Schneider CD, Pinto RS. Echo intensity is negatively associated with functional capacity in older women. Age (Dordr). 2014;36(5):9708.
Lexell J, Taylor CC, Sjostrom M. What is the cause of the ageing atrophy? Total number, size and proportion of different fiber types studied in whole vastus lateralis muscle from 15- to 83-year-old men. J Neurol Sci. 1988;84(2–3):275–94.
Acknowledgements
The authors are grateful to Mr. Siu Sik Cheung (Department of Rehabilitation Science, the Polytechnic University) for his help in the technique of data collection.
Funding
The shear wave Aixplorer was donated by Dr. and Mrs. Lui Che Woo.
Author information
Authors and Affiliations
Contributions
SNF and JX have taken part in the whole design of the study and statistical analysis. JX was responsible for the data collection and manuscript writing. FH edited the manuscript. All authors contributed to the interpretation of the results and the critical revision of the manuscript and agreed to the submission of this paper.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All methods were carried out in accordance with the Declaration of Helsinki. This study was approved by the Human Subject Ethics Subcommittee of the Department of Rehabilitation Sciences of Hong Kong Polytechnic University (HSEARS20150416003). All the participants volunteered for this study and provided informed written consent.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Xu, J., Fu, S.N. & Hug, F. Age-related increase in muscle stiffness is muscle length dependent and associated with muscle force in senior females. BMC Musculoskelet Disord 22, 829 (2021). https://doi.org/10.1186/s12891-021-04519-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12891-021-04519-8