
Abstract
Background
Wilms tumor is the most prevalent embryonal kidney malignancy in children worldwide. Previous genome-wide association study (GWAS) identified that LIM domain only 1 (LMO1) gene polymorphisms affected the susceptibility to develop certain tumor types. Apart from LMO1, the LMO gene family members also include LMO2-4, each of which has oncogenic potential.
Methods
We conducted this five-center case‒control study to assess the correlations between single nucleotide polymorphisms in LMO family genes and Wilms tumor susceptibility. Odds ratios and 95% confidence intervals were calculated to evaluate the strength of the association.
Results
We found LMO1 rs2168101 G > T and rs11603024 C > T as well as LMO2 rs7933499 G > A were significantly associated with Wilms tumor risk. Stratified analysis demonstrated a protective role of rs2168101 GT/TT genotypes against Wilms tumor in the subgroups of age ≤ 18 months, males and clinical stages I/II compared to the rs2168101 GG genotype. Nevertheless, carriers with the rs11603024 TT genotype were more likely to have an increased risk of Wilms tumor than those with rs11603024 CC/CT genotypes in age > 18 months. And the rs11603024 was identified as a protective polymorphism for reducing the risk of Wilms tumor in the sex- and gender- subgroup. Likewise, carriers with the rs7933499 GA/AA genotypes were at significantly elevated risk of Wilms tumor in age ≤ 18 months and clinical stages I/II.
Conclusion
Overall, our study identified the importance of LMO family gene polymorphisms on Wilms tumor susceptibility in Chinese children. Further investigations are needed to validate our conclusions.
Graphical Abstract

Similar content being viewed by others


Introduction
Wilms tumor (WT), or nephroblastoma, is mainly derived from abnormal renal development, especially the failure of metanephric precursor differentiation, and has classified triphasic histology [1]. Wilms tumor is considered a common kidney malignancy affecting young children under 15 years old, accounting for nearly 90% of pediatric renal tumors and approximately 7% of childhood cancers [2, 3]. The incidence and distribution of WT varies widely by race and geography, with the highest incidence of 10 cases per million children in populations of Western black ancestry and the lowest incidence of 3–4 cases per million children in East Asian populations [4, 5]. Currently, with the exception of high-risk, histologically unfavorable, bilateral and recurrent cases, more than 90% of WT patients can benefit from improved modern multimodality therapy and refined risk stratification [6]. However, a proportion of survivors still suffer from chronic adverse health problems and social outcomes [6]. Unlike adult tumors, which are more influenced by the environment and unhealthy lifestyle, pediatric tumors appear to be more susceptible to genetic variation [7]. As published, the genetic landscape of Wilms tumor is diverse and involves approximately 40 cancer genes [8]. Early information on Wilms tumor-associated genes were identified in patients with tumor predisposition syndromes, highlighting the vital roles of Wilms tumor 1 (WT1), WTX, the WNT pathway and so on [8,9,10]. Subsequently, several novel cancer-related genes underpinning the occurrence of Wilms tumor have been primarily deciphered, including microRNA processing genes (DROSHA, DICER1, DGCR8, XPO5 and TARBP2) [1, 8, 11, 12], transcription factors (SIX1 and SIX2) [1, 13], and epigenetic remodelers (SMARCA4 and ARID1A) [8, 14]. Despite significant efforts to study oncogenes in Wilms tumor, we must acknowledge that the genes identified to date could only explain a small part of the etiology of Wilms tumor.
A genome-wide association study (GWAS) is an extremely useful method to focus on the relationship between susceptible genes and complicated human diseases, including cancers [7, 15]. Previous GWAS and candidate gene approaches have discovered some susceptible genes correlated with the predisposition to Wilms tumor, such as METTL3 [16], ALKBH5 [17], BRAD1 [18], PHOX2B [19], and LMO1 [20]. The LMO gene family comprises four members, LMO1-4, which encode many transcriptional cofactors with a common structure of only two tandem LIM domains [21]. Accumulating evidence indicates that the LMO gene family is strongly linked to the occurrence and progression of several cancers [21]. For instance, LMO1 and LMO2 were first described in a chromosomal translocation event at different loci in T-cell leukemia [22]. LMO1 and LMO3 both contribute to the development of neuroblastoma [23, 24]. LMO4 was originally found to be overexpressed in a proportion of breast cancer patients and related to poor prognosis [21, 25].
Currently, there are no genetic association studies between other members of the LMO family gene polymorphisms and Wilms tumor susceptibility. In addition to a replication study of the LMO1 gene in a larger population, we also need further investigation to validate the associations between other underlying SNPs in the LMO1 gene and Wilms tumor susceptibility. With this in mind, we carried out this five-center case‒control study to comprehensively explore whether SNPs in the LMO gene family influence Wilms tumor susceptibility in a larger Chinese population Our current study was the first to explore the associations and contributed to understanding the role of LMO familial genetic variants in the mechanisms of Wilms tumor tumorigenesis, thus filling a gap in this field.
Materials and methods
Study population
A total of 414 Wilms tumor-bearing patients and 1199 cancer-free controls were recruited from five independent hospitals in this case‒control study, as reported previously (Table S1) [17, 26]. All patients were confirmed to have a diagnosis of Wilms tumor by histopathology. Matched to the cases by age, sex and race, the controls were randomly selected from healthy volunteers who received a physical examination at the same hospitals during the periods.
Genotyping
We collected approximately 2 ml of peripheral blood from each participant for DNA extraction and carefully screened for six potentially functional SNPs in the LMO family genes, including rs2168101 G > T and rs11603024 C > T in the LMO1 gene, rs2273799 C > T, rs3758640 A > G and rs7933499 G > A in the LMO2 gene, and rs3766019 G > A in the LMO4 gene. The LMO1 gene rs2168101 G > T was selected according to a previous publication [27], while the remaining five functional SNPs were chosen from the websites of the NCBI dbSNP database (http://www.ncbi.nlm.nih.gov/projects/SNP) and SNPinfo (http://snpinfo.niehs.nih.gov/snpfunc.html). The selection criteria for screening candidate SNPs have been described previously [28, 29]. In brief, we selected functional SNPs located in the 5′ and 3′ untranslated region (UTR), 5′ flanking region, exons, and introns of the LMO family genes. The minor allele frequencies (MAFs) of the chosen SNPs were at or above 5% in the Chinese Han population. Moreover, the linkage disequilibrium (LD) between selected SNPs was low (R2 < 0.8). We included other new LMO1, LMO2 and LMO4 SNPs with potential functions. The rs11603024 in the LMO1 gene, rs2273799 and rs3758640 in the LMO2 gene may affect the function of transcription factor binding site (TFBS), while LMO2 rs7933499 might affect the binding capacity of miRNA and the splicing process (Table S2). DNA was extracted from each subject’s peripheral blood sample using the Genomic DNA kit (Tian Gen Biotech Co. Ltd., Beijing, China), and genotyping of the LMO family gene polymorphisms was performed by the TaqMan real-time PCR assay. In addition, we randomly selected 10% of all samples for repeated assays to avoid the incorrect experimental results, and the results of regenotyping were in 100% concordance with the original results.
Statistical analysis
We then assessed the concordance of genotype frequencies with Hardy-Weinberg Equilibrium (HWE) for the control group through a χ2 goodness-of-fit test. A two-sided χ2 test was used to detect the differences in genotype frequencies and demographic variables between cases and healthy controls. To further investigate the relationship between the LMO family gene polymorphisms and Wilms tumor risk, we also applied unconditional logistic regression to calculate age- and sex-adjusted odds ratios (ORs), 95% confidence intervals (CIs), and two-sided P values. Besides, the Genotype-Tissue Expression (GTEx) (https://gtexportal.org) was applied to performing the expression quantitative trait locus (eQTL) analysis, assessing whether significant SNPs affecting the expression level of corresponding genes or nearby genes. The results were considered statistically significant if P < 0.05. All statistical analyses were performed rigorously with SAS 9.1 software (SAS Institute, Cary, NC, USA).
Results
Association between LMO family gene polymorphisms and Wilms tumor susceptibility
According to the current study, 403 cases and 1198 cancer-free controls were successfully genotyped for the six SNPs in the LMO family gene out of a total of 414 cases and 1199 controls. The correlation of LMO1 gene, LMO2 gene and LMO4 gene polymorphisms with Wilms tumor susceptibility is described in detail in Table 1. The genotype frequencies of the control groups were all consistent with HWE (P = 0.367 for rs2168101 G > T, P = 0.487 for rs11603024 C > T, P = 0.639 for rs2273799 C > T, P = 0.712 for rs3758640 A > G, P = 0.929 for rs7933499 G > A, and P = 0.599 for rs3766019 G > A). Regarding the LMO1 gene, rs2168101 G > T polymorphism was associated with Wilms tumor susceptibility. Specifically, the LMO1 rs2168101 GT/TT genotypes were found to decrease the risk of developing Wilms tumor in the dominant model (adjusted OR = 0.74, 95% CI = 0.59–0.93, P = 0.009). In contrast, subjects carrying the LMO1 rs2168101 TT genotype might have an increased risk of Wilms tumor in the recessive model (adjusted OR = 1.79, 95% CI = 1.25–2.57, P = 0.001). Moreover, another SNP in the LMO1 gene, rs11603024 C > T, was detected to increase Wilms tumor risk in a recessive model (adjusted OR = 4.29, 95% CI = 2.08–8.84, P < 0.0001). Regarding the LMO2 gene, among the three genotyped SNPs (rs2273799, rs3758640, and rs7933499), only the rs7933499 G > A was significantly associated with an increased risk of developing Wilms tumor (dominant model: adjusted OR = 1.37, 95% CI = 1.03–1.82, P = 0.032 and recessive model: adjusted OR = 4.91, 95% CI = 2.11–11.46, P = 0.0002). For the LMO4 gene, there were no obviously significant correlations between the rs3766019 G > A polymorphism and Wilms tumor susceptibility.
Stratification analysis
We then further performed a stratification analysis on some significant SNPs that were related to Wilms tumor by age, sex and clinical stage and explored the relationships between significant SNPs derived from LMO family genes and Wilms tumor risk. The obtained results are depicted in Table 2. We found that rs2168101 GT/TT genotypes played a protective role against the occurrence of Wilms tumor in the subgroups of age ≤ 18 months (adjusted OR = 0.59, 95% CI = 0.40–0.88, P = 0.008), males (adjusted OR = 0.69, 95% CI = 0.50–0.94, P = 0.020) and clinical stages I/II (adjusted OR = 0.74, 95% CI = 0.56–0.97, P = 0.031) compared to the rs2168101 GG genotype. Nevertheless, carriers with the rs11603024 TT genotype were significantly related to an increased risk of Wilms tumor than carriers with rs11603024 CC/CT genotypes among children aged over 18 months (adjusted OR = 5.49, 95% CI = 2.16–13.98, P = 0.0004), females (adjusted OR = 5.72, 95% CI = 1.70-19.25, P = 0.005), males (adjusted OR = 3.63, 95% CI = 1.45–9.07, P = 0.006), clinical stages I/II (adjusted OR = 2.67, 95% CI = 1.05–6.78, P = 0.040) and clinical stages III/IV (adjusted OR = 7.56, 95% CI = 3.23–17.67, P < 0.0001). Similarly, we found that carriers with rs7933499 GA/AA genotypes significantly increased the risk of Wilms tumor in cases aged ≤ 18 (adjusted OR = 1.74, 95% CI = 1.08–2.81, P = 0.023) and clinical stages I/II (adjusted OR = 1.49, 95% CI = 1.06–2.09, P = 0.021) compared to subjects with the rs7933499 GG genotype.
The eQTL analysis of LMO family gene
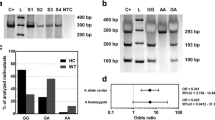
We further used the GTEx database to evaluate the effect of different genotypes of selected SNP on mRNA expression and the result was shown as in Fig. 1. As for rs3758640 A > G, compared to rs3758640 GG genotype, carriers with GA/AA genotypes were associated with reduced mRNA expression of the LMO2 gene in muscle-skeletal and whole blood.

The eQTL analysis of the rs3758640 polymorphism on gene expression using data from GTEx database. In muscle-skeletal (A) and whole blood (B), compared to rs3758640 GG genotype, carriers with GA/AA genotypes were associated with reduced RNA expression of the LMO2 gene
Discussion
As one of the most frequently diagnosed childhood renal malignancies, high-risk Wilms tumor continues to threaten the health of patients with recurrent and unfavorable histological cases. Therefore, with the aim of identifying new biomarkers for early diagnosis and better targeted therapies, we performed a five-center case‒control study to explore the association of LMO family gene polymorphisms and Wilms tumor risk in a Chinese Han population for the first time. The results indicated that LMO1 and LMO2 genetic variants were associated with Wilms tumor susceptibility, which may help identify additional genetic susceptibility loci as novel biomarkers for the diagnosis and treatment of Wilms tumor.
The LMO family genes consist of four members, LMO1-4, each of which has been established to be responsible not only for many normal developmental processes but also for the initiation and development of various human cancers, including T-cell acute lymphoblastic leukemia, neuroblastoma, lung cancer, colorectal cancer, and breast cancer [21, 22, 25, 30,31,32]. The products of LMO family genes share a common two-tandem LIM domain structure, a highly conserved and cysteine-rich zinc binding motif without additional domains, mediating protein‒protein interactions instead of binding to DNA directly [33]. Thus, LMO proteins were previously regarded as adapter molecules for the formation of new versatile multiprotein complexes, which are crucial for participation in various cellular processes, including cell proliferation, cell self-renewal, differentiation, and metastasis [21, 34]. Interestingly, LMO proteins are capable of modulating transcriptional events by forming transcription factor regulators, thereby affecting target gene expression and cell fate decisions [21], suggesting a potential possible mechanism for the involvement of LMO family genes in the pathogenesis and progression of several cancers. Nevertheless, the exact details of the mechanism remain to be fully investigated.
Currently, among the LMO family genes, genetic associations between LMO1 gene polymorphisms and tumor susceptibility have been most intensively studied. An increasing number of publications have identified that LMO1 plays a determinant role in cancer susceptibility. In a previous GWAS performed in participants of European descent, Wang et al. identified that four SNPs (rs110419 A > G, rs4758051 G > A, rs10840002 A > G and rs204938 A > G) in the LMO1 locus at 11p15.4 were strongly associated with the development of neuroblastoma, and subsequent replication studies successfully identified the same findings in different cohorts from the UK, USA and Italy [35]. Thereafter, the relationship between LMO1 polymorphisms and neuroblastoma risk was further determined in other epidemiological case‒control studies among subject populations of various ethnicities [23, 30, 36, 37]. In addition to neuroblastoma, genetic variants in the LMO1 gene also contribute to susceptibility to acute lymphoblastic leukemia (ALL). Beuten et al. discovered that the LMO1 gene rs442264 A > G polymorphism significantly corresponded to an increased risk of developing ALL [38]. Considering the critical effect of LMO1 gene polymorphisms on tumor risk, our team has spared no effort to investigate the relationship between LMO1 gene polymorphisms and Wilms tumor susceptibility.
In 2017, in a case‒control study of 145 Wilms tumor patients and 531 cancer-free controls recruited from southern China, our team successfully genotyped four SNPs in the LMO1 gene (rs110419 A > G, rs4758051 G > A, rs10840002 A > G and rs204938 A > G) and demonstrated for the first time that only rs110419 A > G had a protective effect against developing Wilms tumor [20]. Subsequently, we further explored the correlation between five other potentially functional LMO1 gene polymorphisms (rs2168101 G > T, rs1042359 A > G, rs11041838 G > C, rs2071458 C > A, and rs3750952 G > C) and Wilms tumor risk among the same southern Chinese population mentioned above. The results suggested that only rs2168101 G > T could significantly decrease the risk of Wilms tumor, particularly in GT/TT genotype carriers. And the eQTL analyses showed that GT/TT genotypes reduced the expression level of LMO1 gene in Muscle-Skeletal tissue compared to GG genotype. It is likely that rs2168101 G > T polymorphism participated in the development of Wilms tumor by regulating corresponding LMO1 gene expression level [27]. However, the sample size was too small to limit the statistical power for the conclusion. Additionally, given the structural similarity of the LMO gene family, we reasonably speculated that LMO2 and LMO4 gene polymorphisms might be correlated to Wilms tumor susceptibility as LMO1 gene did. Herein, we expanded the sample size to verify whether functional polymorphisms in the LMO1 gene (rs2168101 and rs11603024), LMO2 gene (rs2273799, rs3758640 and rs7933499) and LMO4 gene (rs3766019) were associated with Wilms tumor susceptibility in a five-center case‒control study for Chinese population.
The current results implied that rs2168101 GT/TT genotypes in the LMO1 gene were found to decrease the risk of developing Wilms tumor in the dominant model (adjusted OR = 0.74, 95% CI = 0.59–0.93, P = 0.009), which is consistent with the tendency for neuroblastoma and Wilms tumor in previous studies [27, 39, 40]. Oldridge et al. [39] observed that rs2168101 G > T is located in a super-enhancer of the first intron area within the LMO1 gene. The rs2168101 G allele constituted a highly conserved transcription factor-binding site of GATA, which helped to promote the expression of the LMO1 gene. When the rs2168101 G allele was mutated to the rs2168101 T allele, it abrogated the binding of GATA3 (P < 0.0001), thereby reducing the expression level of LMO1 [39]. This may account for the decreased risk of tumorigenesis for the rs2168101 G > T polymorphism, including Wilms tumor. When biologically functional SNP in LMO1 gene was mutated, the expression of corresponding genes might be affected and was related to the occurrence of Wilms tumor. In contrast, in the recessive model (adjusted OR = 1.79, 95% CI = 1.25–2.57, P = 0.001), carriers with the LMO1 rs2168101 TT genotype seemed to have an increased risk of Wilms tumor according to an adjusted OR of 1.79. This discovery was opposite to a meta-analysis publication that assessed the association between LMO1 gene polymorphism and neuroblastoma risk. In that meta-analysis, the researcher found that the rs2168101 TT polymorphism was more likely to reduce the risk of neuroblastoma than the rs2168101 GT/GG polymorphisms, with an OR of 0.48 in the recessive model (TT vs. GT + GG: OR = 0.48, 95% CI = 0.31–0.75, P = 0.001) [41]. The obvious differences were mainly attributed to the following reasons: the relatively small size of subject populations, selection bias for participants, chance of the experiments, etc. As a result, replication studies are needed to further validate our findings. Moreover, another SNP locus in the LMO1 gene, rs11603024 C > T, was detected to confer enhanced susceptibility to Wilms tumor in recessive model. The stratified analysis demonstrated that rs2168101 GT/TT genotypes played a protective role against the occurrence of Wilms tumor in the subgroups of age ≤ 18 months, males and clinical stages I/II compared to the rs2168101 GG genotype. Nevertheless, carriers with the rs11603024 TT genotype were significantly related to an increased risk of Wilms tumor than carriers with rs11603024 CC/CT genotypes among children aged over 18 months, females, males, clinical stages I/II and clinical stages III/IV.
Referring to the LMO2 gene, only the rs7933499 G > A was significantly associated with an enhanced risk of developing Wilms tumor. From the stratified analysis, we found that carriers with the rs7933499 GA/AA genotypes could significantly increase the risk of Wilms tumor in the subgroup of age ≤ 18 and clinical stages I/II compared to subjects with the rs7933499 GG genotype. Regarding the LMO4 gene, no significant association was observed between the rs3766019 G > A polymorphism and predisposition to Wilms tumor.
We further assessed the effect of different genotypes of selected SNPs on mRNA expression using the GTEx Portal. The eQTL analysis result of rs3758640 A > G showed that GA/AA genotype carriers were associated with reduced RNA expression of the LMO2 gene in muscle-skeletal and whole blood compared to rs3758640 GG genotype. This implied that the different genotypes of rs3758640 may result in altered mRNA expression of the corresponding genes, which is associated with Wilms tumor susceptibility. However, other polymorphisms were not detected significant associations with Wilms tumor susceptibility. In fact, data resources of the GTEx Portal were almost from Caucasian populations, while our study was conducted in Chinese Han children. Therefore, it is possible that our results could not be exactly explained by the eQTL analysis. Further functional experiments are necessary to validate the current results, and to elucidate the potential pathogenesis of Wilms tumor in the future.
The strength of our study is that we enlarged the sample size from five independent hospitals, and we are the first to comprehensively examine the relationship between LMO family gene polymorphisms and Wilms tumor risk. We used Power and Sample Size Calculation software to evaluate statistical power. Assuming that the MAF of selected SNP was between 0.05 and 0.40, with an α level of 0.05, we were able to detect the risk of Wilms tumor with an OR value of 1.38–1.87 with 80% certainty. However, there are still several limitations for the current study that cannot be ignored. First, even if we expand the sample size, the size of the subject population seems to be relatively small, which inevitably weakens the statistical power. Meanwhile, the lack of combination analysis also weakens the strength of the conclusions. Second, due to the various genetic backgrounds for different ethnicities and regions, the conclusions obtained cannot be applied to other populations until they are validated by replicated studies. Third, the drawbacks of retrospective studies cannot be avoided, so some valuable information, such as environmental factors, was unavailable for further investigations. Finally, we only concentrate on the effect of several functional SNPs in the LMO family genes on Wilms tumor susceptibility, whereas there are many unknown potential polymorphisms in LMO family genes that deserve extensive study. Importantly, there is a strong need for functional experiments to further explore the underlying biological mechanisms of how highly related SNPs in the LMO family genes affect the risk of Wilms tumor.
Conclusion
In brief, the current epidemiological genetic association study identified that several functional SNPs in the LMO family genes, namely, rs2168101 G > T and rs11603024 C > T in the LMO1 gene and rs7933499 G > A in the LMO2 gene, are significantly associated with susceptibility to Wilms tumor. Our findings are worth further validation in well-designed studies with large sample sizes among different races.
Data availability
No datasets were generated or analysed during the current study.
Abbreviations
- WT:
-
Wilms tumor
- WT1:
-
Wilms tumor 1
- GWAS:
-
genome-wide association study
- UTR:
-
untranslated region
- MAF:
-
minor allele frequency
- LD:
-
linkage disequilibrium
- TFBS:
-
transcription factor binding site
- HWE:
-
Hardy-Weinberg equilibrium
- OR:
-
odds ratio
- CI:
-
confidence interval
- GTEx:
-
Genotype-Tissue Expression
- eQTL:
-
expression quantitative trait locus
- ALL:
-
acute lymphoblastic leukemia
References
Hohenstein P, Pritchard-Jones K, Charlton J. The Yin and Yang of kidney development and Wilms’ tumors. Genes Dev. 2015;29(5):467–82.
Servaes SE, Hoffer FA, Smith EA, Khanna G. Imaging of Wilms tumor: an update. Pediatr Radiol. 2019;49(11):1441–52.
Breslow N, Olshan A, Beckwith JB, Green DM. Epidemiology of Wilms tumor. Med Pediatr Oncol. 1993;21(3):172–81.
Stiller CA, Parkin DM. International variations in the incidence of childhood renal tumours. Br J Cancer. 1990;62(6):1026–30.
Nakata K, Colombet M, Stiller CA, Pritchard-Jones K, Steliarova-Foucher E. Incidence of childhood renal tumours: an international population-based study. Int J Cancer. 2020;147(12):3313–27.
Dome JS, Graf N, Geller JI, Fernandez CV, Mullen EA, Spreafico F, Van den Heuvel-Eibrink M, Pritchard-Jones K. Advances in Wilms Tumor Treatment and Biology: progress through international collaboration. J Clin Oncol. 2015;33(27):2999–3007.
Maris JM, Knudson AG. Revisiting tissue specificity of germline cancer predisposing mutations. Nat Rev Cancer. 2015;15(2):65–6.
Treger TD, Chowdhury T, Pritchard-Jones K, Behjati S. The genetic changes of Wilms tumour. Nat Rev Nephrol. 2019;15(4):240–51.
Lee SB, Haber DA. Wilms tumor and the WT1 gene. Exp Cell Res. 2001;264(1):74–99.
Koesters R, Niggli F, von Knebel Doeberitz M, Stallmach T. Nuclear accumulation of beta-catenin protein in Wilms’ tumours. J Pathol. 2003;199(1):68–76.
Walz AL, Ooms A, Gadd S, Gerhard DS, Smith MA, Guidry Auvil JM, Meerzaman D, Chen QR, Hsu CH, Yan C, et al. Recurrent DGCR8, DROSHA, and SIX homeodomain mutations in favorable histology Wilms tumors. Cancer Cell. 2015;27(2):286–97.
Rakheja D, Chen KS, Liu Y, Shukla AA, Schmid V, Chang TC, Khokhar S, Wickiser JE, Karandikar NJ, Malter JS, et al. Somatic mutations in DROSHA and DICER1 impair microRNA biogenesis through distinct mechanisms in Wilms tumours. Nat Commun. 2014;2:4802.
Wegert J, Ishaque N, Vardapour R, Georg C, Gu Z, Bieg M, Ziegler B, Bausenwein S, Nourkami N, Ludwig N, et al. Mutations in the SIX1/2 pathway and the DROSHA/DGCR8 miRNA microprocessor complex underlie high-risk blastemal type Wilms tumors. Cancer Cell. 2015;27(2):298–311.
Hodges C, Kirkland JG, Crabtree GR. The many roles of BAF (mSWI/SNF) and PBAF complexes in Cancer. Cold Spring Harb Perspect Med. 2016;6(8):a026930.
Wang MH, Cordell HJ, Van Steen K. Statistical methods for genome-wide association studies. Semin Cancer Biol. 2019;55:53–60.
Lin A, Zhou M, Hua RX, Zhang J, Zhou H, Li S, Cheng J, Xia H, Fu W, He J. METTL3 polymorphisms and Wilms tumor susceptibility in Chinese children: a five-center case-control study. J Gene Med. 2020;22(11):e3255.
Hua RX, Liu J, Fu W, Zhu J, Zhang J, Cheng J, Li S, Zhou H, Xia H, He J, et al. ALKBH5 gene polymorphisms and Wilms tumor risk in Chinese children: a five-center case-control study. J Clin Lab Anal. 2020;34(6):e23251.
Fu W, Zhu J, Xiong SW, Jia W, Zhao Z, Zhu SB, Hu JH, Wang FH, Xia H, He J, et al. BARD1 gene polymorphisms Confer Nephroblastoma susceptibility. EBioMedicine. 2017;16:101–5.
Lin A, Fu W, Wang W, Zhu J, Liu J, Xia H, Liu G, He J. Association between PHOX2B gene rs28647582 T > C polymorphism and Wilms tumor susceptibility. Biosci Rep. 2019;39(10):BSR20192529.
Liu GC, Zhuo ZJ, Zhu SB, Zhu J, Jia W, Zhao Z, Hu JH, He J, Wang FH, Fu W. Associations between LMO1 gene polymorphisms and Wilms’ tumor susceptibility. Oncotarget. 2017;8(31):50665–72.
Matthews JM, Lester K, Joseph S, Curtis DJ. LIM-domain-only proteins in cancer. Nat Rev Cancer. 2013;13(2):111–22.
Boehm T, Foroni L, Kaneko Y, Perutz MF, Rabbitts TH. The rhombotin family of cysteine-rich LIM-domain oncogenes: distinct members are involved in T-cell translocations to human chromosomes 11p15 and 11p13. Proc Natl Acad Sci U S A. 1991;88(10):4367–71.
He L, Zhu J, Han F, Tang Y, Zhou C, Dai J, Wang Y, Zhou H, He J, Wu H. LMO1 gene polymorphisms reduce Neuroblastoma Risk in Eastern Chinese children: A Three-Center Case-Control Study. Front Oncol. 2018;8:468.
Aoyama M, Ozaki T, Inuzuka H, Tomotsune D, Hirato J, Okamoto Y, Tokita H, Ohira M, Nakagawara A. LMO3 interacts with neuronal transcription factor, HEN2, and acts as an oncogene in neuroblastoma. Cancer Res. 2005;65(11):4587–97.
Sutherland KD, Visvader JE, Choong DY, Sum EY, Lindeman GJ, Campbell IG. Mutational analysis of the LMO4 gene, encoding a BRCA1-interacting protein, in breast carcinomas. Int J Cancer. 2003;107(1):155–8.
Hua RX, Fu W, Lin A, Zhou H, Cheng J, Zhang J, Li S, Liu G, Xia H, Zhuo Z, et al. Role of FTO gene polymorphisms in Wilms tumor predisposition: a five-center case-control study. J Gene Med. 2021;23(8):e3348.
Li G, Jia W, Yin Z, Zhu J, Liu G, Xia H, He J, Fu W. LMO1 Super-enhancer rs2168101 G > T polymorphism reduces Wilms Tumor Risk. J Cancer. 2019;10(8):1808–13.
Chen YP, Liao YX, Zhuo ZJ, Yuan L, Lin HR, Miao L, Li X, Huang XK, Zhou JY, Bian J, et al. Association between genetic polymorphisms of base excision repair pathway and glioma susceptibility in Chinese children. World J Pediatr. 2022;18(9):632–5.
Guan Q, Lin H, Hua W, Lin L, Liu J, Deng L, Zhang J, Cheng J, Yang Z, Li Y, et al. Variant rs8400 enhances ALKBH5 expression through disrupting miR-186 binding and promotes neuroblastoma progression. Chin J Cancer Res. 2023;35(2):140–62.
Zhang J, Lin H, Wang J, He J, Zhang D, Qin P, Yang L, Yan L. LMO1 polymorphisms reduce neuroblastoma risk in Chinese children: a two-center case-control study. Oncotarget. 2017;8(39):65620–6.
Zhang Y, Yang J, Wang J, Guo H, Jing N. LMO1 is a novel oncogene in lung cancer, and its overexpression is a new predictive marker for anti-EGFR therapy. Med Oncol. 2014;31(8):99.
Liu J, Yan P, Jing N, Yang J. LMO1 is a novel oncogene in colorectal cancer and its overexpression is a new predictive marker for anti-EGFR therapy. Tumour Biol. 2014;35(8):8161–7.
Zhao GF, Du LQ, Zhang L, Jia YC. LIM domain only 1: an oncogenic transcription cofactor contributing to the tumorigenesis of multiple cancer types. Chin Med J (Engl). 2021;134(9):1017–30.
Bach I. The LIM domain: regulation by association. Mech Dev. 2000;91(1–2):5–17.
Wang K, Diskin SJ, Zhang H, Attiyeh EF, Winter C, Hou C, Schnepp RW, Diamond M, Bosse K, Mayes PA, et al. Integrative genomics identifies LMO1 as a neuroblastoma oncogene. Nature. 2011;469(7329):216–20.
Capasso M, Diskin SJ, Totaro F, Longo L, De Mariano M, Russo R, Cimmino F, Hakonarson H, Tonini GP, Devoto M, et al. Replication of GWAS-identified neuroblastoma risk loci strengthens the role of BARD1 and affirms the cumulative effect of genetic variations on disease susceptibility. Carcinogenesis. 2013;34(3):605–11.
Latorre V, Diskin SJ, Diamond MA, Zhang H, Hakonarson H, Maris JM, Devoto M. Replication of neuroblastoma SNP association at the BARD1 locus in African-americans. Cancer Epidemiol Biomarkers Prev. 2012;21(4):658–63.
Beuten J, Gelfond JA, Piwkham D, Pollock BH, Winick NJ, Collier AB 3rd, Tomlinson GE. Candidate gene association analysis of acute lymphoblastic leukemia identifies new susceptibility locus at 11p15 (LMO1). Carcinogenesis. 2011;32(9):1349–53.
Oldridge DA, Wood AC, Weichert-Leahey N, Crimmins I, Sussman R, Winter C, McDaniel LD, Diamond M, Hart LS, Zhu S, et al. Genetic predisposition to neuroblastoma mediated by a LMO1 super-enhancer polymorphism. Nature. 2015;528(7582):418–21.
He J, Zhang X, Zhang J, Zhang R, Yang T, Zhu J, Xia H, Zou Y. LMO1 super-enhancer polymorphism rs2168101 G > T correlates with decreased neuroblastoma risk in Chinese children. J Cancer. 2018;9(9):1592–7.
Hashemi M, Sarabandi S, Karami S, Smieja J, Moazeni-Roodi A, Ghavami S, Los MJ. LMO1 polymorphisms and the risk of neuroblastoma: Assessment of meta-analysis of case-control studies. J Cell Mol Med. 2020;24(2):1160–8.
Acknowledgements
Not applicable.
Funding
This study was funded by grants from the Natural Science Foundation of Guangdong Province (No: 2021A1515010860), Science and Technology Project of Guangzhou (No: 202102010170), National Natural Science Foundation of China (No: 82003523), and Natural Science Foundation of Zhejiang Province (No: LGF21H260012).
Author information
Authors and Affiliations
Contributions
All authors contributed significantly to this work. WF, JH, and GL designed the research study; WF, RXH, XY, JZ, HZ, CD, SL, JC, JR, JH, and GL performed the research study and collected the samples and clinical information; JH analyzed the data and prepared all the tables; WF, LD, RXH, and JH wrote the paper. All authors included in the final manuscript have read and approved the final version to be published.
Corresponding authors
Ethics declarations
Ethics approval
All operational processes in this study strictly obeyed the Declaration of Helsinki. The Institutional Review Board of Guangzhou Women and Children’s Medical Center (No. 202016601) approved this research, and the participants’ parents or their guardians signed written informed consent forms.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Fu, W., Deng, L., Yan, X. et al. LMO family gene polymorphisms and Wilms tumor susceptibility in Chinese children: a five-center case-control study. BMC Cancer 24, 772 (2024). https://doi.org/10.1186/s12885-024-12557-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12885-024-12557-3