
Abstract
Background
Colorectal cancer (CRC) is ranked as the third most commonly diagnosed cancer and the third cause of cancer related deaths. CRC is greatly attributed to genetic and epigenetic mutations and immune dysregulation. Tumor aberrant expression of Toll-like Receptors (TLRs) can contribute to tumorigenesis. Recent studies suggested that microRNAs act as direct ligands of TLRs altering their expression and signaling pathways.
Aim
To prove our concept that specific miRNA mimics may act as antagonists of their specific toll like receptors inhibiting their expression that could limit the release of pro-inflammatory and pro-tumorigenic cytokines leading to apoptosis of tumor cells.
Methods
From public microarray databases, we retrieved TLRs and miRNAs related to CRC followed by in silico docking of the selected miRNA ligands into the TLRs. Clinical validation after co-immunoprecipitation of TLRs and their interacting miRNA ligands was done. Expression of TLRs 1, 7,8 was determined by ELISA while miRNAs was measured by RT-qPCR. In addition, microRNA mimics of the down regulated miRNAs were transfected into human CRC cell lines.
Results
Our data demonstrate that TLRs 1, 7, 8 are up regulated in CRC compared to controls. Further, three miRNAs (-122, -29b and -15b) are relatively downregulated, while 4 miRNAs (-202, miRNA-98, -21 and -let7i) are upregulated in CRC patients compared to those with benign tumor and healthy controls. Transfection of down regulated miRNA mimics into CRC cell lines resulted in a significant reduction of the number and viability of cells as well as down regulating the expression of TLRs 1, 7 and 8 with ultimate reduction of downstream effector IL6 protein, suggesting that these miRNAs are negative regulators of carcinogenesis.
Conclusion
MicroRNAs could act as antagonistic ligands of TLRs limiting the inflammatory tumor microenvironment.
Key points
• Specific miRNAs were identified by in silico docking as signaling antagonists of 3 TLRs and were validated in clinical samples.
• TLRs 1, 7, 8 were upregulated while their miRNAs (-122, -29b and -15b)were down regulated in CRC.
• Transfection of downregulated miRNA mimics into CRC cell lines resulted in downregulation of TLRs that was associated with reduction in IL6 protein and tumor growth.
• The present study proposed a novel approach that enables a reliable integration of Toll like receptors and miRNAs in CRC pathogenesis.
Similar content being viewed by others


Introduction
Colorectal cancer (CRC) is the third most commonly diagnosed cancer worldwide. It represents the third leading cause of cancer related mortality in men and women as well [1]. Many factors are implicated in the pathogenesis of CRC including gene mutations, epigenetic alterations, local inflammatory responses, and immune system deregulation [2]. However, it has become increasingly clear that chronic inflammation of the intestine is a major carcinogenic process. The resulting release of inflammatory cytokines and the ensuing immune reaction result in epigenetic changes, recruitment of immune cells as well as pro-tumorigenic signals that contribute to initiation and progression of tumor growth [3].
Toll-like receptors (TLRs) are a diverse family of Pattern Recognition Receptors (PRR) expressed by intestinal epithelial cells, tumor cells and immune cells from both the innate and adaptive arms of the immune system [4]. Several studies highlighted the dual role of certain TLRs in cancer progression as well as anti-cancer immune responses. It was found that TLRs on the surface of tumor cells with their ligands can activate subsequent signaling cascades and recruit more immune cells in the tumor microenvironment to enhance inflammation and this TLR-induced inflammation might stimulate tumor growth and spread into the microenvironment [5]. On the other hand, aberrant expression of certain TLRs by tumor cells correlated with growth inhibition and antitumor response [6].
MicroRNAs (miRNAs) are a class of small non-coding RNAs with an average 22 nucleotides in length. Although the conventional role of microRNAs is regulation of gene expression, recent studies suggested that microRNAs can act as direct ligands of TLRs [7]. For example, miRNA-21 released from cancer cells exosomes was found to bind to TLR7 and ultimately led to TLR-mediated tumor metastasis and growth [8]. Also, studies found that cancer cell-derived exosomal miRNAs can bind to TLRs in macrophages and intestinal cells and stimulate NF-κB pathway with subsequent release of the pro-inflammatory and pro-metastatic cytokines IL-6 and TNF-α which stimulate tumor growth and metastasis [9]. MicroRNA molecules are now at the center of molecular oncology, with applications for diagnosis and therapy starting to be proposed. The ability of miRNAs to regulate important cellular processes by concurrently regulating multiple targets illustrates their potential as a viable therapeutic tool [10]. miRNA have crucial role in GIT malignancy [11] including CRC [12].
The development of miRNA-based therapy for cancer treatment is based on the premise that aberrantly expressed miRNAs play a crucial role in the development of cancer and therapeutic response to anticancer drugs [13]. Therefore, correcting the miRNA deficiency or restoring the miRNA function could be used as a novel strategy in cancer treatment [14]. Based on the fact that some miRNAs have dual role in regulation of gene expression; a)the tumor suppressive role of the selected miRNAs through binding to Argonaute proteins and guide the silencing of target genes; b) on the other hand, they act independently of Argonaute proteins by interacting directly with TLRs as an antagonist thus representing a possible target for cancer treatment [15]. So, we hypothesize that specific miRNA mimics may act as antagonists of their specific toll like receptors inhibiting their expression that could limit the release of pro-inflammatory and pro-tumorigenic cytokines leading to apoptosis of tumor cells.
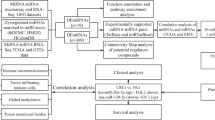
To prove this hypothesis, we identified a panel of miRNA ligands that interact with specific toll like receptors to regulate their expression and signaling in CRC using bioinformatics methods. Then, we confirmed their presence in patient’s samples and cell lines, lastly, we investigated the efficacy of specific miRNA mimics in CRC growth inhibition A summary of the of the study experimental procedures is presented in (Fig. 1A).

A flow chart of the experimental procedures performed to validate the study hypothesis. B Rationale for bioinformatics based selection of miRNA-TLR specific to CRC pathogenesis
Materials and methods
In-silico analysis
Bioinformatics based selection of miRNA-TLR specific to CRC pathogenesis is provided in (Fig. 1b). The COSMIC database v78 (http://cancer.sanger.ac.uk/cosmic) was downloaded and interrogated for TLR1,7 & 8 mutations. Mutations were counted by cancer type as defined by the filters for tissue type and described in S1&2 Table and figure S1. Public microarray databases were used to examine the expression of candidate TLRs in CRC tissues and normal tissues using updated GENT2 Gene expression database [16] (Available at: http://gent2.appex.kr/gent2/, Accessed on: 18/9/2022). Three TLRs; TLR1, TLR7 and TLR8 were selected based on high ranking score of their differential expression in CRC compared to controls by Two-sample T-test analysis (p value < 0.001 and Log2FC = -0.309 for TLR1, p value < 0.001 and Log2FC = -0.806 for TLR7 and with p value = 0.004 and Log2FC = -0.157 for TLR8. The chosen CRC specific TLR 1,7 and 8 were verified in previous literatures [14] and other databases e g. Expression atlas, Disease Gene and colorectal cancer gene databases to decrease the false discovery rate. Then, gene ontology tracking of the three TLRs was done through Princeton Gene ontology tools database and European Bioinformatics institute Quick GO annotations database [17].
Moreover, seven miRNAs were identified from public databases (Colon cancer microRNA, Human miRNA Tissue atlas expression database, miRo: a miRNA knowledge base and dbDEMC: database of differentially expressed miRNAs in human cancers [18]). Hsa-miR122, hsa-miR29b, hsa-miR 15b, hsa-miR202, hsa-miR 98, hsa-miR 21 and hsa-miR let 7i were selected according to their high ranking score of their differential expression in CRC compared to control. The results were verified in previous literatures [15,16,17,18,19,20,21]. Then, pathway enrichment analysis of the seven miRNAs using DIANA tools database was carried out. Lastly, we examined the direct interaction between selected miRNAs and the TLRs via the in silico analysis. Discovery studio 3.5 software (DS 3.5; Accelrys Co. Ltd, San Diego, California, USA) was used for homology modelling and docking of miRNAs into TLRs [19, 20]. ZDOCK predicts all possible binding poses between the miRNA ligand and TL receptor and evaluates each pose using ZDOCK score, an energy-based scoring function which is a combination of pairwise shape complementarity, desolvation and electrostatics [21, 22] (Further details of docking methodology is provided in supplementary Figure S17-23). ZDOCK is a docking program that predicts all possible binding poses in the translational and rotational space between the ligand and receptor and evaluates each pose using ZDOCK score, an energy-based scoring function [23]. ZDOCK generated 60 clusters containing 2000 structures and ranked them according to their ZDOCK score. While, RDOCK is a program designed to refine and re-rank top predictions from ZDOCK using energy minimization algorithm [22].
Patient population
This study was approved by the Ethical Committee of the Faculty of Medicine, Ain Shams University (FMASU MD 379/2018) and all experimental procedures were in compliance with the Declaration of Helsinki and informed consents were obtained from all study participants. CRC cases were diagnosed according to The American Cancer Society (ACS) guidelines for diagnosis of CRC [24]. Before any intervention such as radiotherapy, chemotherapy or even surgery, 32 paired serum and tissue samples were initially collected from colonoscopy unit at Ain Shams University hospitals in the period from December 2019 till the end of 2021. Samples were classified as 16 CRC, 8 benign colorectal neoplasms and 8 normal samples from patients with normal colon mucosa.
Tissues were stored at -80 °C till further analysis. Then, other 100 sera samples were collected from patients at the oncology Clinic, General Surgery Department, Ain Shams University Hospitals to be added to the previous collected samples from colonoscopy unit. Altogether, serum samples included 50 samples from CRC patients, and 25 from patients with benign colorectal neoplasm. Moreover, 25 sera were obtained from healthy volunteers with matching age and sex to the patients’ groups.
All tumors were generally characterized as primary moderately differentiated adenocarcinomas via colonoscopy. The clinical stages of the CRC patients were determined according to the latest TNM staging system of the American Joint Committee on Cancer (AJCC) [25] where 30%-37% patients were in early stage (stage 0, 1) and 63%-70% patients were in late stage of CRC (stage 2, 3, 4). The clinical data for every patient were available in details. The serum levels of Carbohydrate Antigen (CA19.9) and Carcino-embroyonic Antigen (CEA), were assessed by.
Immunoradiometric method according to manufacturer’s instructions.
Sample processing
For colon tissue lysis, the full thickness colon tissue was homogenized in 1 mL cell lysis buffer containing protease inhibitor cocktails (Roche, 05056489001). The homogenate was incubated on ice for 30 min and finally the samples were centrifuged at 13,000 X g for 20 min at 4°C. Supernatants were later subjected to immunoprecipitation with the indicated antibodies in protein A/G beads.
All study samples (sera and tissue supernatant)were diluted in RIPA buffer (10 mM Tris (pH 7.4), 150 mM NaCl, 1% Triton X100, 1% Na. deoxycholate; dilution 1:1), supplemented with protease inhibitors cocktail (Sigma- Aldrich, Milano, Italy) to further proceed to Co-immunoprecipitation.
Co-immunoprecipitation
Co-IP was performed to identify the TLR-miRNA ligand interaction using the Pierce Co-Immunoprecipitation kit (Pierce, Thermo Fisher Scientific, Illinois, USA) according to manufacturer’s instructions. Antibodies used for immunoprecipitation were TLR1 monoclonal antibody purchased from (abcam,MA,USA#ab68158), TLR7 monoclonal antibody purchased from (Cell signaling technology, Beverly, MA, USA #5632s) and TLR8 monoclonal antibody purchased from Santa-Cruz Biotechnology (Santa Cruz Biotechnology, California, USA #SC-135584). Briefly, a 50-μl aliquot of the supplied antibody-coupling gel slurry (AminoLink Plus gel) was added to the pierce spin columns and washed with the coupling buffer supplied in the kit followed by conjugation with 50 μg of primary antibodies;TLR1-ab,TLR7-ab and TLR8-ab. The primary antibodies were covalently linked to the coupling gel by the addition of sodium cyanoborohydride and left for 2 h incubation at room temperature with constant agitation. Samples were then diluted with IP Lysis/ Wash Buffer (supplied within the kit; dilution 1:1 according to manufacturer’s instructions) then added to the antibody coupled resin with a maximum of 500 μl per spin column and incubated overnight with gentle mixing at 4°C.The spin columns were then washed and eluted in 50 µl elution buffer supplied within the kit. The CO-IP elute was then divided into two halves: a half for TLR protein estimation by ELISA and the other half for miRNA ligand expression by RT-qPCR.
ELISA measurement of TLRs protein in Co-IP eluate
The total protein in Co-IP eluate was measured using the BioRad Bradford protein assay (Biorad, CA, USA). Quantitative detection of TLRs 1,7 and 8 protein was done using (EIAab® TLR1, TLR7, TLR8 ELISA, EIAab Science Inc, Wuhan, China) kits according to manufacturer’s instructions. Luminescence signal was detected using BioTek ELX800 microplate luminescence reader (Bio-Tek Instr., Winooski, VT, USA).
MicroRNA extraction and Protein digestion of Co-IP elute
Total RNA (including miRNAs) was isolated from Co-IP elute using reagents supplied within the miRNeasy Mini kit (Qiagen, CA, USA) in addition to proteinase K buffer (Qiagen, CA, USA) for protein digestion during RNA preparation. Briefly the was re-suspended in 150 μl proteinase K buffer and incubated at 55 °C for 30 min with shaking. After the incubation, the supernatant was transferred into 250 μl RIPA Buffer (Thermo Fisher Scientific, Illinois, USA) and 400 μl of Qiazol Lysis reagent (Qiagen, CA, USA) was added to each tube. After centrifugation, the aqueous phase was removed into a new tube, mixed with 400 μl chloroform and vortexed briefly then centrifuged to separate phases. The aqueous phase was removed into a new tube. 50 μl of RWT buffer and 30 μl of RPE buffer (Buffers of miRNeasy Mini kit, Qiagen, CA, USA) were added to enhance the precipitation of RNA at -80 °C for overnight. After centrifugation, the pellet was washed by 80% ethanol and re-suspended in 15 μl of RNase-free water as illustrated in the protocol mentioned by Wang et al., (2018) [26]. The concentration and purity of the extracted RNA were determined by Thermo Scientific NanoDrop Spectrophotometer ( Thermo Scientific, Wilmington, USA) where the RNA purity was found to be ≥ 1.8.
Reverse Transcription-quantitative Polymerase Chain Reaction (RT-qPCR)
The extracted total RNA was reverse transcribed into cDNA using the miRCURY LNA RT kit (Qiagen, Hilden, Germany# cat no. 339340) according to manufacturer’s instructions where the thermal cycler; Thermo Hybaid PCR express (Thermo Fisher Scientific, Illinois, USA) was programmed for 60 min at 42 degrees then 5 min at 95 degrees to inactivate the transcriptase enzyme and finally cooled at 4 degrees.
The expression of 7 miRNAs was analyzed; hsa-miR-122-3p, hsa-miR-29b-3p, hsa-let-7i-3p, hsa-miR-15b-3p, hsa-miR-98-3p, hsa-miR-202-3p, hsa-miR-21-3p against endogenous control cel-mir-39 in a micro RNA array using miRCURY LNA miRNA Custom PCR Panel (Qiagen, Hilden, Germany# cat no. 339330) and miRCURY LNA SYBR Green PCR kit (Qiagen, Hilden, Germany #cat no.339345) according to manufacturer’s instructions. Due to Locked Nucleic Acid (miRCURY LNA) technology used in the primer synthesis and covered by patents owned by Qiagen, the sequences of the primers were not provided. The reaction was run on the ABI 7500 Real –Time PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc., Illinois, USA). qPCR was conducted as follows: 95˚C for 2 min, 40 cycles at 95˚C for 10 s, 56˚C for 60 s and melting curve analysis at 60 ˚C -95 ˚C. Relative gene expression levels were normalized to a housekeeping gene (SNORD 68). The 2−∆∆Ct equation was used to calculate the expression of the selected RNA-based candidate gene panel using the Applied Biosystems 7500 software v2.3 (Applied Biosystems; Thermo Fisher Scientific, Inc., Illinois, USA). In this study, appropriate standardization strategies were implemented according to MIQE guidelines [27].
CRC cell lines and culture
Human CRC cell lines LS174T and HT29 were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). The cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen, CA, USA) as per the manufacturer’s recommendations supplemented with 10% fetal bovine serum (FBS; Invitrogen, CA, USA) and 1% streptomycin in a humidified atmosphere of 5% CO2 at 37 °C. Cells were passaged at 75% confluence with 0.02% EDTA/0.25% trypsin. Cell culture pellet would be used for Co-immunoprecipitation then TLRs protein detection by ELIZA and miRNAs by Rt-PCR as above mentioned.
Measurement of IL6 by ELISA according to the manufacturer’s instructions, the concentrations of interleukin ( IL-6), in the cell culture pellets were analyzed using an ELISA kit( Invitrogen, USA) with measurement of the optical density (OD) at 450 nm on An automated microplate reader. And the results were expressed per mg protein.
Transfection of miRNA mimics
Three miRNA mimics of the downregulated miRNAs; miRCURY LNA miRNA mimic hsa-miR-122-3p (5'AACGCCAUUAUCACACUAAAUA3’# YM00470503), miRCURY LNA miRNA mimic hsa-miR-29b-3p (5'UAGCACCAUUUGAAAUCAGUGUU3’ # YM00473486) and miRCURY LNA miRNA mimic hsa-miR-15b-3p. (5'CGAAUCAUUAUUUGCUGCUCUA3’ # YM00472191) in addition to one negative control; miRCURY LNA miRNA mimic Negative control (UCACCGGGUGUAAAUCAGCUUG # YM00479902) were all purchased from (Qiagen, CA,USA).The tubes were centrifuged prior to opening and re-suspended by adding 300 μl RNAase-free water to the 20 nmol miRNA mimics and negative control to reach a final concentration of 20 μM.
LS174T and HT29 cells were seeded in 24-well plates at 5 × 104 cells per well in triplets. After 24 h, cells were transiently transfected by using HiPerFect Transfection reagent (Qiagen, CA, USA) according to the manufacturer’s instructions. After 36 h, a second transfection was done reintroducing the same transfection complexes to the cells in order to ensure good transfection of the cells. After 72 h post the second transfection, cells were harvested and washed with PBS (Gibco; Thermo Fisher Scientific, MA, USA) and briefly centrifuged to precipitate pellet for Co-immunoprecipitation then miRNA expression analysis by quantitative RT-qPCR and supernatant for TLRs and IL6 expression by ELISA. Details of miRNA extraction, reverse transcription, RT-qPCR and ELISA methodology are as previously mentioned.
Cellular viability was determined by the MTT assay. For that purpose, both control miRNA- and targeted mimic-transfected clones were cultured in 96-well plates (5,000 cells/well) in a final volume of 100 μL to a confluence of 50%. The culture media was replaced 72 h later by the MTT reagent, which was incubated for 4 h at 37 ℃. Finally, equal volumes of dimethyl sulfoxide were added to each well to dissolve the crystals and the absorbance was measured at 570 nm, the reference wavelength being 690 nm.
Statistical analysis
Statistical analysis was performed using the IBM SPSS Statistics for Windows, Version 25 (IBM SPSS Statistics for Windows, IBM Corporation, Armonk, NY, USA). Data were presented as the means ± standard deviation (SD) repeated in triplicates in tissue culture. A Student’s t-test was used for the statistical comparison of data. The nonparametric Kruskal–Wallis test was used for statistical comparison of the variables among the various groups where ranks were assigned. Chi-square test was also used to find out the relation between various qualitative data. Receiver‐operating characteristic (ROC) curves and the area under the ROC curve (AUC) were used to evaluate the specificity, diagnostic ability, and sensitivity of each gene to discriminate between CRC versus control. Youden index (calculated as J = Sensitivity + Specificity − 1) was used to determine the best effective diagnostic panel with the highest sensitivity and specificity [28]. Statistical differences were considered significantly if a P-value was less than 0.05 and highly significant if less than 0.01.
Results
Bioinformatics retrieval of CRC specific TLRs and their suspected miRNAs ligands
Applying bioinformatics analysis, three TLRs; TLR1, TLR7 and TLR8 and seven miRNAs (Hsa-miR122,hsa-miR29b,hsa-miR 15b, hsa-miR202, hsa-miR 98, hsa-miR 21 and hsa-miR let 7i) were selected from public data bases, based on high ranking score of their differential expression in CRC compared to controls. The 3 selected TLRs were found to be related to cancer and immunomodulatory pathways through their implication in positive regulation of tumor necrosis factor biosynthesis pathway, immune response activating and regulating pathway, leucocyte activation pathway involved in inflammatory response, regulation of cytokine biosynthetic pathway and pattern recognition receptor signaling pathway.
Pathway enrichment analysis of the seven miRNAs using DIANA tools database for all miRNAs [29] showed their implication in CRC and multiple cancers such as endometrial, prostate, pancreatic, Renal cell carcinoma and lung cancer. The selected miRNAs were also implicated in immunomodulatory pathways such as Transforming Growth Factor B (TGF-B) signaling pathway, FOXO signaling pathway, P53 signaling pathway, Hippo signaling pathway and AMPK signaling pathway.
Then, we searched for the possible clusters formation by the selected miRNAs in order to predict potential miRNA-CRC associations. We scanned Transcription factor and miRNA regulatory cascades in human diseases database that predicts miRNA clusters in a certain disease [30]. Our data revealed that three of the selected miRNAs; miRNA-21, miRNA-29b and miRNA-let7i could form clusters and thus exhibit high potentiality of being associated in CRC.
Using the ZDOCK score, we performed several Docking runs; miRNA-21, miRNA-122 and miRNA-15b were docked into TLR1 then miRNA-21 and miRNA-202 were docked into TLR7 and finally miRNA-21 and miRNA-29b were docked into TLR8. The interaction analysis using ZDock and RDock revealed that miRNAs could bind to TLRs as direct ligands with high affinity in the form of high ZDock score. Specifically, TLR1 bind to miRNA-21 with highest affinity (ZDock score = 22.9, Fig. 2A), miRNA-15b (ZDock score = 21.66, Fig. 2B) and miRNA-122 with the least ZDock score (ZDock score = 20.56, Fig. 2C). On the other hand, TLR7 bind to miRNA-20 with highest affinity (ZDock score = 25.96, Fig. 2D) followed by miRNA-21 (ZDock score (ZDock score = 25.12, Fig. 2E). Notably, we found that miRNA-21 could also bind to TLR8 with higher affinity than that for TLR7 (ZDock score = 25.82, Fig. 2F). TLR8 binds with high affinity to miRNA-29b (ZDock score = 29.74, Fig. 2G). Further analysis indicated that miRNA-TLR complexes are relatively stable, and the interacting amino acids of TLRs exist at the binding interfaces with miRNAs. (More figures for Bioinformatics analysis and further details of docking interfaces are provided in Fig.S1-Fig.S24).

Docking of miRNA ligands into TLRs using ZDock protocol in silico (A) miRNA-21 was docked into TLR1 with ZDock score 22.9, B miRNA-15b was docked into TLR1 with ZDock score 21.66, C miRNA-122 was docked into TLR1 with ZDock score 20.56, D miRNA-202 was docked into TLR7 with ZDock score 25.96, E miRNA-21 was docked into TLR7 with ZDock score 25.12, F miRNA-21 was docked into TLR8 with ZDock score 25.82, G miRNA-29b was docked into TLR8 with ZDock score 29.74
Clinical, laboratory and pathological factors among the three study groups
Results of paired tissue and serum samples which were collected from patients with CRC, benign tumors and normal colonic mucosa, show significant statistical difference (p < 0.05) in Body Mass Index (BMI), smoking, CEA, CA19.9 the classical tumor markers of CRC, colonoscopy appearance of tumor, histological type of the tumor, Table S3.
Differential expression of TLRs and their potential miRNA ligands in investigated groups
The protein level of TLR1, TLR7 and TLR8 in immunoprecipitated serum and tissue samples from all groups was measured using ELISA. In tissue samples, we found that TLR 1. TLR7. and TLR8 s expression is upregulated in CRC group compared to control groups (~ 6, 3, and 3 folds higher, respectively) with a highly significant statistical difference among the three groups (p < 0.01), Tables S4,S5.
We observed strong correlation between tissue and serum levels of all investigated TLRs and miRNAs. Altogether, these results suggest frequent communication between blood and corresponding CRC tissues and potential miRNA signature in blood, which are not random but have regular patterns. Thus, serum miRNAs could be a non-invasive biomarker for facilitating the diagnosis and prognosis of CRC.
In 100 serum samples, expression of TLR1, TLR7, and TLR8 was found to be upregulated in CRC group compared to control groups (~ 12, 11 and 15 folds higher, respectively) (p < 0.01).(Tables S4 & S5, Fig. 3). ANOVA post-hoc test revealed a highly significant difference between CRC and benign groups in the three TLRs (p = 0.000), but a non-significant difference between healthy control and benign groups.

Simple BOXPLOT graph represents differential expression of TLRs in IP of tissue and serum samples of CRC versus benign and normal control cases determined by ELISA (A-F). Serum CRC tumor markers CEA (G) and CA19.9 in the investigated groups. The line inside the box is the median. The top and bottom lines of the box are the first and third quartiles, respectively. The top and bottom whiskers are the 5th and 95th percentiles, respectively
We also analyzed the miRNA expression of 7 miRNAs suspected to act as TLR ligands by RT-qPCR in the immunoprecipitate of both tissue and serum samples of the three study groups. In the initial screening set of tissue samples, three of the miRNAs (miRNA-122, miRNA-29b: and miRNA-15b) were found to be relatively downregulated in CRC group compared to Benign and normal control groups. The expression level of RQ of miRNA-122, miRNA-29b and miRNA-15b was 12, 10, 13 folds, respectively, lower in CRC group compared to control with a highly significant statistical difference among the three groups (p < 0.01). As for miRNAs miRNA-202, miRNA-98, miRNA 21 and miRNA-let7i, they were found to be relatively upregulated in CRC group compared to benign and normal control groups. Their fold change of expression RQ was increased by 15, 806, and 189, 88.5 times respectively in CRC group compared to control group (Tables S4 & S5, Fig. 4A).

A-I Clustered BOXPLOT graph represents serum miRNA-122, miRNA-29b, miRNA-15b differential expression as determined by RT-qPCR among CRC, Benign adenoma and healthy control groups in IP of serum. A-II Clustered BOXPLOT graph represents serum miRNA-98, miRNA-202, miRNA-21, miRNA-let7i differential expression as determined by RT-qPCR among CRC, Benign adenoma and healthy control groups in IP of serum. The line inside the box is the median. The top and bottom lines of the box are the first and third quartiles, respectively. The top and bottom whiskers are the 5th and 95th percentiles, respectively. B Simple BOXPLOT graph represents differential expression of investigated miRNAs suggested to act as TLRs ligands in IP of tissue samples of CRC versus benign and normal control cases determined by RT-qPCR. The line inside the box is the median. The top and bottom lines of the box are the first and third quartiles, respectively. The top and bottom whiskers are the 5th and 95th percentiles, respectively
In the clinical validation set of 100 serum samples, three of the miRNAs (miRNA-122, miRNA-29b: and miRNA-15b) were found to be relatively down regulated in CRC group compared to Benign and normal control groups where their fold change of expression RQ was decreased in CRC group 13 times compared to control with a highly significant statistical difference among the three groups (p < 0.01). While four miRNAs (miRNA-202, miRNA-98, miRNA 21 and miRNA-let7i) were found to be relatively upregulated in CRC group compared to benign and normal control groups. The fold change of expression RQ of miRNA-98 and miRNA-let7i was increased 21 times in CRC group compared to control group, while fold change of expression RQ was increased 29 times for miRNA-202 and miRNA-21 in CRC group compared to control group with a highly significant statistical difference among the three groups (p < 0.01). (Tables S4 & S5, Fig. 4B). ANOVA post-hoc test revealed a significant difference between healthy control and benign groups in miRNA-122(p = 0.038), and a highly significant difference between CRC and benign groups in miRNA-15b (p = 0.006), while a significant difference between CRC and benign groups in miRNA-202(p = 0.036) and miRNA-21 (p = 0.033).
The discriminatory power of the investigated TLRs and their miRNA ligands
To investigate the discriminatory power of investigated serum TLRs and their miRNA ligands in CRC patients from patients with benign colorectal adenoma and healthy volunteers, ROC curves were analyzed and AUC values were obtained (Fig. 5). The discriminatory power was found to be highest in TLR1 (AUC = 0.954, best cut off value = 0.237) followed by TLR8 (AUC = 0.937, best cut off value = 0.521), TLR7 (AUC = 0.934, best cut off value = 0.397), miRNA-122 (AUC = 0.898, best cut off value = 0.9814), miRNA-29b (AUC = 0.885, best cut off value = 1.275), miRNA-21(AUC = 0.885, best cut off value = 1.255), miRNA-15b (AUC = 0.880, best cut off value = 0.966), miRNA-98 (AUC = 0.807, best cut off value = 1.118), miRNA-let7i (AUC = 0.797, best cut off value = 1.019), miRNA-202 (AUC = 0.735, best cut off value = 1.185) and at last the classical CRC biomarkers; CEA (AUC = 0.715, best cut off value = 3.05) and CA19.9 (AUC = 0.677, best cut off value = 4.86). Hence, these optimal cutoff values of the investigated biomarkers could be used to discriminate between CRC from benign adenoma patients and healthy participants suggesting their use as powerful diagnostic biomarkers of CRC surpassing the discriminatory power of the classical biomarkers CEA and CA19.9.

Receiver operator characteristics (ROC) curves representing the investigated serum TLRs-miRNA network discriminatory power of CRC from benign neoplasm patients and healthy volunteers. A TLR1, B TLR7, C TLR8, D miRNA-122, E miRNA-29b, F miRNA-15b, G miRNA-98, H miRNA-202, I miRNA-21, J miRNA-let7i, K CEA, and L CA19.9
Correlation between investigated serum TLRs and their potential ligands of miRNAs
A highly significant negative correlation was found between TLR1, TLR7, TLR8 and miRNA-122, miRNA-29b and miRNA-15b (the down regulated miRNAs) while a highly significant positive correlation was found between TLR1, TLR7, TLR8 and miRNA-98, miRNA-202, miRNA-21, miRNA-let7i (the upregulated miRNAs) (p < 0.01) (Table S6). This implies the high possibility of miRNAs acting as direct ligands of TLRs as proposed by bioinformatics analysis described above.
Effects of miRNA mimics of miRNA-122, miRNA-29b, miRNA-15b on CRC cell count
To further examine the role of differentially expressed miRNA in CRC pathogenesis, we transfected human CRC cell lines with three miRNA; miRNA-122, miRNA-29b and miRNA-15b, which were aberrantly downregulated in CRC tissue lysates compared to controls. Briefly, human CRC cell lines; LS174T and HT29 were successfully transfected with Hiperfect Transfection reagent as described under M&M and we examined the following 6 groups: untransfected (untreated) group, Mock group (only transfection reagent added to assess its cytotoxicity), group transfected with miRNA mimic negative control, group transfected with miRNA mimic of miRNA-122, group transfected with miRNA mimic of miRNA-29b and group transfected with miRNA mimic of miRNA-15b. Notably, our results indicated that higher expression of these mimics in both CRC cell lines significantly reduced cellular proliferation.
(LS174T cell line shown in Fig. 6A1-A6) and (HT-29 cell line shown in Fig. 6B1-B6) as well as viability (Fig. 6A7 in LS174T cell line; Fig. 6B7 in HT-29 cell line). The cell count was decreased in groups transfected with miRNA mimics by 2 folds when compared to mock control group (= p < 0.01), The percentage of viable cells was reduced by ~ twofold s in groups transfected with miRNA mimics compared to the mock group (p < 0.01). However, there was no significant difference as regards count between group transfected with miRNA mimic negative control and mock group (p = 0.108 and 0.360 respectively and between untransfected group and mock group there was no significant difference as well (p = 0.211 and 0.129 for count.

A LS174T CRC cell count and viability analysis 72 h post transfection showing; (A1) high cell count in the untransfected group,(A2) high cell count (minimal cytotoxicity of transfection reagent) in the mock group, (A3) high cell count in the miRNA mimic-transfected negative control group, (A4) marked reduction in cell count in the miRNA mimic-122 transfected group, (A5) marked reduction in cell count in the miRNA mimic-29b transfected group, (A6) marked reduction in cell count in the miRNA mimic-15b transfected group, (A7) Simple BOXPLOT graph represents the difference in the viability % of LS174T CRC cells among the study groups. B HT29 CRC cell count and viability analysis 72 h post transfection showing; (B1) high cell count in the untransfected group,(B2) high cell count (minimal cytotoxicity of transfection reagent) in the mock group, (B3) high cell count in the miRNA mimic-transfected negative control group, (B4) marked decrease in cell number in the miRNA mimic-122 transfected group, (B5) pronounced lowering in cell number in the miRNA mimic-29b transfected group, (B6) marked reduction in cell count in the miRNA mimic-15b transfected group, (B7) Simple BOXPLOT graph represents the difference in the viability % of HT29 CRC cells among the study groups
Effects of miRNA mimics of miRNA-122, miRNA-29b, miRNA-15b on the viability of CRC cells
The percentage of viable cells was reduced by ~ twofold s in groups transfected with miRNA mimics compared to the mock group (p < 0.01). However, there was no significant difference as regards viability of cells between group transfected with miRNA mimic negative control and mock group (p = 0.869 and 0.567) and between untransfected group and mock group there was no significant difference as well (p = 0.211 and (p = 0.092 and 0.072).
Effects of miRNA mimics of (miRNA-122, -29b, -15b) transfection on the expression of miRNA-TLR –IL6 in CRC cell line
Both cell lines LS174T and HT29 were successfully transfected with miRNA mimics and the expression of the investigated miRNAs that were previously downregulated in IP of serum and tissue lysates of CRC were increased by 4 times in cells transfected with miRNA mimics; miRNA-122, miRNA-29b and miRNA-15b with a highly significant difference compared to the mock group (p < 0.01) in LS174T and HT-29 cell lines. Their relative expression was measured in the cell pellet of harvested cells 72 h post second transfection by RT-qPCR.
As for the TLRs; TLR1, TLR7 and TLR8 that were previously significantly upregulated in IP of serum and tissue lysates of CRC were found to be significantly decreased by 19 times for TLR1 as well as TLR8 and 57 times for TLR7 in cells transfected with miRNA mimics; miRNA-122, miRNA-29b and miRNA-15b with a highly significant difference compared to the mock group (p < 0.01) in LS174T and HT-29 cell lines. Their relative expression was measured in the supernatant of harvested cells 72 h post second transfection by ELISA (Fig. 7). On the other hand, IL6 protein expression -the target effector of this signal pathway- was significantly lowered in both cell lines after miRNA transfection (Table 1).

A graphical comparison among the studied groups in CRC cell line; untransfected group, mock group, negative control group, group transfected with miRNA mimic 122, group transfected with miRNA mimic 29b and group transfected with miRNA mimic 15b as regards (A) mean of cell count in LS-174 T CRC cell line, B mean of cell count in HT-29 CRC cell line, C cell viability percentage in LS-174 T cell line, D cell viability percentage in HT-29 cell line, E Fold change RQ of the three investigated miRNAs; miR-122, miR-29b, miR-15b by RT-qPCR in LS-174 T cell line, F Fold change RQ of the three investigated miRNAs; miR-122, miR-29b, miR-15b by RT-qPCR in HT-29 cell line, G Relative protein expression of TLR1 by ELISA in LS-174 T cell line, H Relative protein expression of TLR7 by ELISA in LS-174 T cell line, I Relative protein expression of TLR8 by ELISA in LS-174 T cell line, J Relative protein expression of TLR1 by ELISA in HT-29 cell line, K Relative protein expression of TLR7 by ELISA in HT-29 cell line, L Relative protein expression of TLR8 by ELISA in HT-29 cell line
Discussion
Increasing evidence has suggested the new role of miRNAs as direct ligands of TLRs besides their classical role of regulation of gene expression [11]. However, the effects of miRNA-TLRs signaling on CRC pathogenesis are still poorly understood. In the present study, our in silico analysis identified, for the first time, three TLRs (TLR1, TLR7 and TLR8) and seven miRNAs (hsa-miR-122-3p, hsa-miR-29b-3p, hsa-let-7i-3p, hsa-miR-15b-3p, hsa-miR-98-3p, hsa-miR-202-3p, hsa-miR-21-3p) as differentially expressed molecules in CRC compared to controls. We have also assessed the possibility of binding between the selected TLRs and miRNAs as direct ligands through insilico molecular docking ZDock protocol.
Recently, computational strategies helped greatly in novel drug design of which molecular docking; a well-established computational method of drug discovery; is widely used to predict how a ligand interacts with its receptor and how tightly it binds revealing the electrostatic and steric complementarity between the protein and the ligand [31]. Also, the Prediction of miRNA ligand interaction with receptor protein is highly desirable to design miRNA-based therapeutics [32]. In this study, we introduced ZDock protocol approach of molecular docking of miRNAs into TLRs protein predicting the direct interaction between them. After performing several runs for docking the selected miRNAs into the selected TLRs; we found that relatively stable complexes could be formed by miRNA ligand binding to TLR with relatively high affinity as per high ZDock score.
Our in insilico analysis was consistent with in vivo expression of these TLRs and miRNA in serum and tissue samples from CRC patients.
We found the protein expression of TLR1, TLR7 and TLR8 to be aberrantly upregulated in CRC cases compared to their protein expression in benign neoplasms cases and healthy control samples in agreement to the studies of [33] who reported the up regulation of TLR1, TLR7 and TLR8 in CRC patients in comparison to control individuals [14]. On the other hand, we measured the expression of 7 miRNAs proposed or hypothesized to act as TLRs ligands by RT-qPCR and found that three of them (miRNA-122, miRNA-29b and miRNA-15b) were downregulated in CRC cases compared to benign cases and healthy normal control, while the other four (miRNA-202, miRNA-98, miRNA 21, miRNA-let7i) were found to be upregulated.
Several studies support our findings [15,16,17]. Although previous reports confirmed the differential expression of miRNA 122 in colorectal cancer versus normal control which was proved in our study, Sun et al., [34] found that exosomal miRNA-122 was significantly overexpressed in CRC patients especially in those with liver metastasis [34]. Also, Maierthaler et al., [35] defined a clear association of miR‐122 levels to the occurrence of liver metastasis in CRC patients [35]. Moreover, Pan et al., [36] demonstrated the upregulation of miRNA-15b in CRC cases in comparison to normal control [36]. This discrepancy is mostly attributed to the presence of liver metastasis associated with CRC patients in these previous reports since miR-122 is a liver specific miRNA that is highly enriched within hepatocytes. Also, liver metastasis with other types of cancer e.g. gastric cancer is associated with overexpression of miRNA 122 which has been a reported as a useful marker to differentiate colorectal with liver metastasis versus those without [34, 37]. It could also be also attributed to population differences that were observed for complex traits which may be due to the combined effect of socioeconomic, environmental, genetic and epigenetic factors [38]. Moreover, variations in biological samples type (exosome, serum, tissue), samples processing and RNA quantitation methods are responsible for discrepancy in miRNAs expression between different studies [39].
Accumulating evidence reported the upregulated expression of (miRNA-202, miRNA-98, miRNA 21, miRNA-let7i) in CRC cases compared to benign cases and healthy normal control [18,19,20,21].In contrast, Lin et al. [40] Zhu et al. [41], and Song et al. [42], reported the downregulated expression of miR-202, miR-98 and let7i in CRC tissues compared to adjacent colon tissues respectively [40,41,42].
In the light of previous findings We tend to focus on miRNAs role in CRC beyond the post-transcriptional level by serving as ligands of TLRs [8, 43]. Based on our study docking results, that verified the stability of miRNA-TLR complexes, we postulated that miRNA-122, -29b, and-15b can act as a signaling antagonist of TLR 1and -8 because the interacting amino acids of TLRs exist at the binding interfaces with miRNAs sequence in agreement with Heil approach [44]. These insilico results were validated by coimmunoprecipitation of TLRs-miRNA complexes from serum and tissue samples. Thus, synthetic miRNA-122, -29b, and-15b may act both as paracrine antagonists and transcription suppressors of TLRs 1,7 and 8 receptor protein inhibiting macrophages via suppressing MyD88 thus inhibiting the formation of a complex with TRAF6, IRAK1, IRAK4 that leads to NF- kappa B suppression [45], and inhibiting proinflammatory cytokines secretion (e.g., TNF-α and IL-6). The major sources of IL-6 in CRC are tumor-associated mesenchymal stem cells, macrophages and colon cancer-associated stromal fibroblasts. TLR1,7,8 inhibition ultimately leads to cessation of Epithelial to Leucocytic transition. Epithelial to leucocytic transition (ELT) is the acquisition of immune properties by tumor cells. Epithelial tumor cells can make transition to a mesenchymal phenotype which increases local motility and remodeling of the extracellular matrix This will leads to tumor growth inhibition that go in hand with Luddy and his colleagues [46].
Conclusion
The present study proposed a novel approach that enables a reliable integration of Toll like receptors and miRNAs in CRC management. We hypothesized that miRNAs act as TLR ligands where they block or inhibit pro-tumorigenic TLR1/7/8-mediated tumor growth and survival in CRC (Fig. 8).

Proof of concept of study hypothesis. A CRC patients showed upregulation of oncogenic miRNA-202,-98, 21, and let7i and downregulation of tumor suppressive miRNA-122, -29b, and-15b with subsequent activation of TLR1,7, 8 that resulted in CRC pathogenesis by epithelial to leukocytes transition is the acquisition of immune properties by tumor cells (1) evade the immune system at the primary tumor site, (2) access the lymphatic system, (3) circulate through the vasculature, home to favorable sites of metastasis, and extravasate into a metastatic niche, and (4) avoid destruction by the immune system at the site of metastasis. B Proof of concept of study hypothesis: miRNA-122, -29b, and-15b act as antagonists of TLR 1,7 ,8 with suppressing MyD88 and inhibiting the formation of a complex with TRAF6, IRAK1, IRAK4 that leads to NF- kappa B and TNFα suppression with CRC tumor growth inhibition
Availability of data and materials
Data is provided within the manuscript or supplementary information files.
References
Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer statistics, 2022. CA Cancer J Clin. 2022;72(1).
Jung G, Hernández-Illán E, Moreira L, Balaguer F, Goel A. Epigenetics of colorectal cancer: biomarker and therapeutic potential. Nat Rev Gastroenterol Hepatol. 2020;17(2):111–30.
Greten FR, Grivennikov SI. Inflammation and cancer: triggers, mechanisms, and consequences. Immunity. 2019;51(1):27–41.
Vijay K. Toll-like receptors in immunity and inflammatory diseases: Past, present, and future. Int Immunopharmacol. 2018;1(59):391–412.
Skorka K, Wlasiuk P, Karczmarczyk A, Giannopoulos K. Aberrant Expression of TLR2, TLR7, TLR9, Splicing Variants of TLR4 and MYD88 in Chronic Lymphocytic Leukemia Patients. J Clin Med. 2021;10(4):867.
Javaid N, Choi S. Toll-like receptors from the perspective of cancer treatment. Cancers. 2020;12(2):297.
Xie Z, Huang G, Wang Z, Luo S, Zheng P, Zhou Z. Epigenetic regulation of Toll-like receptors and its roles in type 1 diabetes. J Mol Med. 2018;96(8):741–51.
Fabbri M, Paone A, Calore F, Galli R, Gaudio E, Santhanam R, Lovat F, Fadda P, Mao C, Nuovo GJ. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc Natl Acad Sci U S A. 2012;109:E2110–6.
Bayraktar R, Bertilaccio MT, Calin GA. The interaction between two worlds: microRNAs and toll-like receptors. Front Immunol. 2019;14(10):1053.
Shah MY, Ferrajoli A, Sood AK, Lopez-Berestein G, Calin GA. microRNA therapeutics in cancer—an emerging concept. EBioMedicine. 2016;1(12):34–42.
Nedaeinia R, Manian M, Jazayeri MH, Ranjbar M, Salehi R, Sharifi M, Mohaghegh F, Goli M, Jahednia SH, Avan A, Ghayour-Mobarhan M. Circulating exosomes and exosomal microRNAs as biomarkers in gastrointestinal cancer. Cancer Gene Ther. 2017;24(2):48–56. https://doi.org/10.1038/cgt.2016.77. (Epub 2016 Dec 16 PMID: 27982021).
Nedaeinia R, Sharifi M, Avan A, Kazemi M, Nabinejad A, Ferns GA, Ghayour-Mobarhan M, Salehi R. Inhibition of microRNA-21 via locked nucleic acid-anti-miR suppressed metastatic features of colorectal cancer cells through modulation of programmed cell death 4. Tumour Biol. 2017;39(3):1010428317692261. https://doi.org/10.1177/1010428317692261. (PMID: 28347230).
Shah MY, Calin GA. MicroRNAs as therapeutic targets in human cancers. Wiley Interdisciplinary Reviews: RNA. 2014;5(4):537–48.
To KK, Tong CW, Wu M, Cho WC. MicroRNAs in the prognosis and therapy of colorectal cancer: From bench to bedside. World J Gastroenterol. 2018;24(27):2949.
Chen X, Liang H, Zhang J, Zen K, Zhang CY. microRNAs are ligands of Toll-like receptors. RNA. 2013;19(6):737–9. https://doi.org/10.1261/rna.036319.112. Epub 2013 Apr 3. PMID: 23554231; PMCID: PMC3683908.
Park SJ, Yoon BH, Kim SK, Kim SY. GENT2: an updated gene expression database for normal and tumor tissues. BMC Med Genomics. 2019;12(5):1–8.
Brooksbank C, Cameron G, Thornton J. The European Bioinformatics Institute’s data resources. Nucleic Acids Res. 2010;38(Database issue):D17–25.
Yang Z, Wu L, Wang A, Tang W, Zhao Y, Zhao H, Teschendorff AE. dbDEMC 2.0: updated database of differentially expressed miRNAs in human cancers. Nucleic Acids Res. 2017;45(D1):D812–8.
Wiehe K, Pierce B, Mintseris J, Tong WW, Anderson R, Chen R, Weng Z. ZDOCK and RDOCK performance in CAPRI rounds 3, 4, and 5. Proteins. 2005;60(2):207–13.
Pierce BG, Hourai Y, Weng Z. Accelerating protein docking in ZDOCK using an advanced 3D convolution library. PLoS ONE. 2011;6(9): e24657.
Li HL, Ma Y, Ma Y, Li Y, Chen XB, Dong WL, Wang RL. The design of novel inhibitors for treating cancer by targeting CDC25B through disruption of CDC25B-CDK2/Cyclin A interaction using computational approaches. Oncotarget. 2017;8(20):33225–40.
Li L, Chen R, Weng Z. RDOCK: refinement of rigid-body protein docking predictions. Proteins. 2003;53(3):693–707.
Sohn R, Parks JM, Buhrman G, Brown P, Kristjansdottir K, Safi A, Edelsbrunner H, Yang WT, Rudolph J. Experimental validation of the docking orientation of Cdc25 with its Cdk2-CycA protein substrate. Biochemistry. 2005;44:16563–73. https://doi.org/10.1021/bi0516879.
Wolf AM, Fontham ET, Church TR, Flowers CR, Guerra CE, LaMonte SJ, Smith RA. Colorectal cancer screening for average-risk adults: 2018 guideline update from the American Cancer Society. CA Cancer J Clin. 2018;68(4):250–81.
Weiser MR. AJCC 8th edition: colorectal cancer. Ann Surg Oncol. 2018;25(6):1454–5.
Wang X, Yu H, Sun W, Kong J, Zhang L, Tang J, Zhang H. The long non-coding RNA CYTOR drives colorectal cancer progression by interacting with NCL and Sam68. Mol Cancer. 2018;17(1):110.
Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL, Vandesompele J. The MIQE Guidelines: M inimum I nformation for Publication of Q uantitative Real-Time PCR E xperiments. Clin Chem. 2009;55(4):611–22.
DeLong ER, DeLong DM, Clarke-Pearson DL. Comparing the areas under two or more correlated receiver operating characteristic curves: a nonparametric approach. Biometrics. 1988;44:837–45.
Vlachos IS, Zagganas K, Paraskevopoulou MD, Georgakilas G, Karagkouni D, Vergoulis T, Hatzigeorgiou AG. DIANA-miRPath v3. 0: deciphering microRNA function with experimental support. Nucleic Acids Res. 2015;43(W1):W460–6.
Wang S, Li W, Lian B, Liu X, Zhang Y, Dai E, Li X. TM REC: A Database of Transcription Factor and MiRNA Regulatory Cascades in Human Diseases. PLoS One. 2015;10(5).
Sousa SF, Ribeiro AJ, Coimbra JTS, Neves RPP, Martins SA, Moorthy NSHN, Ramos MJ. Protein-ligand docking in the new millennium–a retrospective of 10 years in the field. Curr Med Chem. 2013;20(18):2296–314.
Agrawal P, Singh H, Srivastava HK, Singh S, Kishore G, Raghava GP. Benchmarking of different molecular docking methods for protein-peptide docking. BMC Bioinformatics. 2019;19(13):105–24.
Lu CC, Kuo HC, Wang FS, Jou MH, Lee KC, Chuang JH. Upregulation of TLRs and IL-6 as a marker in human colorectal cancer. Int J Mol Sci. 2014;16(1):159–77. https://doi.org/10.3390/ijms16010159.
Sun L, Liu X, Pan B, Hu X, Zhu Y, Su Y, Guo Z, Zhang G, Xu M, Xu X, Sun H, Wang S. Serum exosomal miR-122 as a potential diagnostic and prognostic biomarker of colorectal cancer with liver metastasis. J Cancer. 2020;11(3):630–7.
Maierthaler M, Benner A, Hoffmeister M, Surowy H, Jansen L, Knebel P, Burwinkel B. Plasma miR-122 and miR-200 family are prognostic markers in colorectal cancer. Int J Cancer. 2017;140(1):176–87.
Pan C, Yan X, Li H, Huang L, Yin M, Yang Y, Qin H. Systematic literature review and clinical validation of circulating microRNAs as diagnostic biomarkers for colorectal cancer. Oncotarget. 2017;8(40):68317.
Coulouarn C, Factor VM, Andersen JB, Durkin ME, Thorgeirsson SS. Loss of miR-122 expression in liver cancer correlates with suppression of the hepatic phenotype and gain of metastatic properties. Oncogene. 2009;28(40):3526–36. https://doi.org/10.1038/onc.2009.211. Epub 2009 Jul 20. PMID: 19617899; PMCID: PMC3492882.
Huang RS, Gamazon ER, Ziliak D, Wen Y, Im HK, Zhang W, Wing C, Duan S, Bleibel WK, Cox NJ, Dolan ME. Population differences in microRNA expression and biological implications. RNA Biol. 2011;8(4):692–701. https://doi.org/10.4161/rna.8.4.16029. Epub 2011 Jul 1. PMID: 21691150; PMCID: PMC3225983.
Cervena K, Novosadova V, Pardini B, Naccarati A, Opattova A, Horak J, Vodenkova S, Buchler T, Skrobanek P, Levy M, Vodicka P, Vymetalkova V. Analysis of MicroRNA Expression Changes During the Course of Therapy In Rectal Cancer Patients. Front Oncol. 2021;2(11): 702258. https://doi.org/10.3389/fonc.2021.702258.PMID:34540669;PMCID:PMC8444897.
Lin Y, Chen Z, Lin S, Zheng Y, Liu Y, Gao J, Chen S. MiR-202 inhibits the proliferation and invasion of colorectal cancer by targeting UHRF1. Acta Biochim Biophys Sin. 2019;51(6):598–606.
Zhu W, Huang Y, Pan Q, Xiang P, Xie N, Yu H. MicroRNA-98 suppress Warburg effect by targeting HK2 in colon cancer cells. Dig Dis Sci. 2017;62(3):660–8.
Song J, Wang L, Ma Q, Yang Y, Yang Z, Wang B, He N. Let-7i-5p inhibits the proliferation and metastasis of colon cancer cells by targeting kallikrein-related peptidase 6. Oncol Rep. 2018;40(3):1459–66.
Meng F, Henson R, Wehbe-Janek H, Ghoshal K, Jacob ST, Patel T. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology. 2007;133:647–58.
Heil F, Hemmi H, Hochrein H, Ampenberger F, Kirschning C, Akira S, Lipford G, Wagner H, Bauer S. Species-specific recognition of single-stranded RNA via Toll-like receptor 7 and 8. Science. 2004;303:1526–9.
Karin M, Greten FR. NF-kappaB: linking inflammation and immunity to cancer development and progression. Nat Rev Immunol. 2005;5:749–59. https://doi.org/10.1038/nri1703.
Luddy KA, Robertson-Tessi M, Tafreshi NK, Soliman H, Morse DL. The role of toll-like receptors in colorectal cancer progression: evidence for epithelial to leucocytic transition. Front Immunol. 2014;20(5):429. https://doi.org/10.3389/fimmu.2014.00429.PMID:25368611;PMCID:PMC4202790.
Acknowledgements
N-A.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). This work was supported by STDF-Science and Technology Development Fund for accredited centers of scientific excellence, Cairo, Egypt (Grant no. 4118 51).
Author information
Authors and Affiliations
Contributions
M.M. shared in conceptualization, methodology, supervision, and reviewing and editing. All authors have read and agreed to the published version of the manuscript. N.H. and D.F. shared in Bioinformatic and docking analysis, validation, original drafting, methodology. M.H. shared in investigation. H.S and M.A. shared in data analysis and original drafting, N. I. shared in data analysis and draft reviewing. S.E. participated in funding acquisition and resources, supervision, and original drafting—reviewing and editing. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Informed consent was obtained from all subjects involved in the study. This study was approved by the Ain Shams Faculty of Medicine IRB (Approval no.:FWA0000017585), and the work described has been carried out in accordance with code of ethics of the world medical association (Declaration of Helsinki).
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Matboli, M., Hossam, N., Farag, D. et al. miRNAs: possible regulators of toll like receptors and inflammatory tumor microenvironment in colorectal cancer. BMC Cancer 24, 824 (2024). https://doi.org/10.1186/s12885-024-12417-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12885-024-12417-0