
Abstract
Background
Stonin1 (STON1) is an endocytic protein but its role in cancer remains unclear. Here, we investigated the immune role of STON1 in kidney renal clear cell carcinoma (KIRC).
Methods
We undertook bioinformatics analyses of the expression and clinical significance of STON1 in KIRC through a series of public databases, and the role of STON1 in the tumor microenvironment and the predictive value for immunotherapy and targeted treatment in KIRC were identified with R packages. STON1 expression was validated in clinical KIRC tissues as well as in KIRC and normal renal tubular epithelial cells.
Results
Through public databases, STON1 mRNA was found to be significantly downregulated in KIRC compared with normal controls, and decreased STON1 was related to grade, TNM stage, distant metastasis and status of KIRC patients. Compared with normal controls, STON1 was found to be downregulated in KIRC tissues and cell lines. Furthermore, OncoLnc, Kaplan–Meier, and GEPIA2 analyses also suggested that KIRC patients with high STON1 expression had better overall survival. The high STON1 group with enriched immune cells had a more favorable prognosis than the low STON1 group with decreased immune cells. Single sample Gene Set Enrichment Analysis and Gene Set Variation Analysis indicated that STON1 creates an immune non-inflamed phenotype in KIRC. Moreover, STON1 was positively associated with mismatch repair proteins and negatively correlated with tumor mutational burden. Furthermore, Single sample Gene Set Enrichment Analysis algorithm and Pearson analysis found that the low STON1 group was more sensitive to immune checkpoint blockage whereas the high STON1 group was relatively suitable for targeted treatment.
Conclusions
Decreased STON1 expression in KIRC leads to clinical progression and poor survival. Mechanically, low STON1 expression is associated with an aberrant tumor immune microenvironment. Low STON1 is likely to be a favorable indicator for immunotherapy response but adverse indicator for targeted therapeutics in KIRC.
Similar content being viewed by others


Introduction
Renal cell carcinoma (RCC) is a type of cancer generally derived from renal tubular epithelial cells, with 73,820 new cases and 14,770 deaths reported in 2019 [1]. Most cases occur in developed countries, and the morbidity rate in men is two to three times that in women [2]. There are various subtypes of RCC according to the 2016 World Health Organization (WHO) classification of tumors of the urinary system, such as kidney renal clear cell carcinoma (KIRC), chromophobe RCC, papillary RCC, and collecting duct and renal medullary carcinoma [3]. KIRC accounts for 70% of cases of RCC [4].
The current treatment strategy for RCC includes partial nephrectomy, radical nephrectomy, ablative therapies, and targeted therapeutics [5], and its pathogenesis has been widely explored; however, 25% of RCC patients still undergo disease progression or metastasis [6]. Recently, immune checkpoint blockage (ICB) therapy was developed. Monoclonal antibodies against immune checkpoint blockade molecules—such as CTLA-4 (cytotoxic T lymphocyte antigen 4), PD-1 (programmed cell death 1), and PD-L1 (programmed cell death ligand 1)—target the tumor microenvironment (TME) with an obvious objective response rate for KIRC [7]. Nevertheless, the absence of economical and effective indicators for predicting response to ICB treatment remains a hindrance. Thus, there is an urgent need to identify novel biomarkers to guide prognosis, ICB treatment prediction, and clinical management.
Stonins are an evolutionally conserved family that mediate the recovery and circulation of vesicles at neuromuscular junctions, and consist of Stonin1 (STON1, also known as germ-line-specific transcription factor) and Stonin2 (STON2) [8]. The human STON1 protein consists of 735 amino acids and has a predicted molecular mass of 83 kDa [9]. Due to the alternative splicing, another two STON1 isoforms with molecular mass of 127 kDa and 129 kDa were displayed in uniport database. Additionally, the level of STON1 gene methylation is a prognostic marker for the progression and personalized treatment of obesity [10]. Focal adhesion (FA) plays a crucial role in tumor cell motility which are orchestrated by signals cells receive from outside via cell surface receptors. STON1, as a regulator of FA dynamics and cell motility, can facilitates the internalization of the oncogenic proteoglycan neuron-glial antigen 2 (NG2), an FA-associated transmembrane protein serving as a promoter of cellular motility and tumor growth [11], but the potential functions of STON1 in mediating specific molecular mechanisms in carcinomas remain entirely unexplored.
In our current study, the expression of STON1 in KIRC was explored using online databases combined with clinical tissue array and cell lines. The prognostic value of STON1 in KIRC was explored using bioinformatics analysis. Additionally, the relationship between STON1 expression and the tumor microenvironment was primarily presented with Kaplan–Meier survival analysis. Our multidimensional analysis ultimately demonstrated that high STON1 expression contributed to a non-inflamed KIRC phenotype, and that cases with low expression of STON1 seem to be sensitive to ICB, whereas high STON1 cases are more suitable for targeted treatment. Our current research has revealed the potential new role of STON1 in cancer, special for KIRC.
Methods
Expression of STON1 in KIRC
The Tumor Immune Estimation Resource (TIMER) database (TIMER2.0, http://timer.comp-genomics.org/) is a comprehensive web server for systematical analysis of immune infiltrates across diverse cancer types. We used Diff Exp module to explore STON1 mRNA expression between pan-cancer and normal tissue, including KIRC. The statistical significance computed by the Wilcoxon test [12]. The independent KIRC cohorts were downloaded from the Gene Expression Omnibus (GEO) database, including GSE16441 (platform: GPL6480), GSE16449 (platform: GPL6480), and GSE71963 (platform: GPL6480), with the “GEO2R” online tool [13] (https://www.ncbi.nlm.nih.gov/geo/). STON1 protein expression was assessed using the UALCAN database (http://ualcan.path.uab.edu/index.html) which provides protein expression analysis option using data from Clinical Proteomic Tumor Analysis Consortium (CPTAC) [14].
Tissue microarray and immunohistochemical staining (IHC)
The tissue microarray data (Cat: HKidE180Su02) was obtained from Shanghai Outdo Biotech Co., LTD (Shanghai, China), which enrolled 30 normal kidney tissues and 150 KIRC tissues. The clinicopathological parameters of the KIRC patients in the tissue microarray are presented in Supplementary Table 1. The IHC was performed using a DAKO automatic immunohistochemistry instrument with the programs of the “Autostainer Link 48 Usage Guide”. The array was incubated with primary antibodies against STON1 (Cat: ABP53586, Abbkine, Wuhan, China) at 1:200 dilution overnight at 4 °C. Non-immune IgG was used as a negative control. Antigenic sites were visualized using a UltraSensitive SP kit (Cat: KIT-9709, Maxin, Fuzhou, China) and DAB kit (Cat: GK600511, GeneTech, Shanghai, China). The STON1 scores were calculated as follows: 0, negative; 1, weak; 2, moderate; and 3, strong. The percentage of positive cells was scored as follows: 1, 0–25% positive cells; 2, 26–50% positive cells; 3, 51–75% positive cells, and 4, 76–100% positive cells. The total immunoreactive scores were determined by the nuclear staining score plus the cytoplasm membrane staining score.
Analysis of the relationships between STON1 and clinical phenotype and prognosis in KIRC
RNA sequencing data and the related clinicopathological data were downloaded from TCGA using UCSC Xena (https://xena.ucsc.edu/), an online explorer that allows users to explore functional genomic data sets for correlations between genomic and phenotypic variables. A total of 491 patients obtained from UCSC Xena were enrolled in the cohort after exclusion criteria were applied. The exclusion criteria were as follows: 1) patients without follow-up records; 2) patients without a diagnosis of KIRC; 3) the primary tumor could not be evaluated; and 4) the stage and grade were not reported. Finally, 491 patients were divided into high and low STON1 expression groups according to the median expression value of STON1. The OncoLnc database (http://www.oncolnc.org/) is a platform for survival analysis with TCGA data [15]. Kaplan–Meier Plotter (https://kmplot.com/analysis/) is an accessible online website with the purpose of identifying survival biomarkers. The prognosis value of each marker based on different immune cells backgrounds was also assessed [16]. We applied OncoLnc and Kaplan–Meier to identify the independent prognostic value of STON1 in KIRC. We also analyzed the prognostic value of STON1 expression in KIRC patients using the Gene Expression Profiling Interactive Analysis (GEPIA) browser (http://gepia2.cancer-pku.cn/#survival) [17].
The immunological role of STON1 in the KIRC tumor microenvironment (TME)
We first identified the infiltration score of 35 immune cells by exploring Assistant for Clinical Bioinformatics (https://www.aclbi.com), which consists of fifteen modules, as well as the immune score, stroma score, and microenvironment score. After selecting the Immunity module, we divided the TCGA-KIRC cohort into low and high STON1 groups according to the median value of STON1, excluding cases where the metastasis and grade (Mx and Gx) could not be evaluated. The statistical difference of two groups was compared through the Wilcoxon test and the consequence was presented with box plot by using Xcell algorithm. The cBio Cancer Genomics Portal (http://cbioportal.org) is an open-access resource for interactive exploration of multidimensional cancer genomics data sets [18]. Kidney was selected as the primary site in the query module of the database, which contained 17 data sets with different sample quantity. Then, KIRC dataset containing 538 tumor samples (Kidney Renal Clear Cell Carcinoma-TCGA, Firehose Legacy) was screened for further analysis, and input the gene symbol STON1. Finally, the co-expression module was used to analyze the correlation between STON1 and expression of immune cell biomarker. The anti-tumor immune response is a multistep coordinated process called the cancer-immunity cycle, which determines the final direction of immune activation or suppression [19]. The cancer-immunity cycle, including seven steps based on specific marker gene sets (Supplemental Table 2), was obtained from the TIP (Tracking Tumor Immunophenotype) database (http://biocc.hrbmu.edu.cn/TIP/) [20]. The enrichment score, indicating anti-cancer immunity, was calculated with the Single sample Gene Set Enrichment Analysis (ssGSEA) algorithm using the R package “GSEABase” and was presented with a boxplot by employing the online tool SangerBox 3.0 (http://vip.sangerbox.com/home.html). The correlation heatmap between STON1 expression and the enrichment score of the cancer-immunity cycle pathways was presented with the R package “ggcorrplot”. In addition, we selected four stromal pathways with immunosuppressive effects from the previous literature, including epithelial–mesenchymal transition (EMT) markers and the pan-fibroblast TGF-β response signature (Pan-FTBRS) [21]. The Gene Set Variation Analysis (GSVA) algorithm was performed to calculate the enrichment score of these signatures.
Correlation of tumor mutational burden (TMB), microsatellite instability, copy number variation (CNV), tumor mRNA stemness index (mRNAsi) and STON1
Spearman correlation analysis was conducted to identify the correlation between STON1 expression and microsatellite instability and a marginal scatter plot was acquired by the “ggpubr” R package for the KIRC mRNA profile. The relationship between the expression of STON1, the tumor mutational burden (TMB) score and the tumor mRNA stemness index (mRNAsi) was displayed by the Assistant for Clinical Bioinformatics with TCGA module and CSCs module, an open integrated database described above (https://www.aclbi.com/). The TIMER database with SCNA module was employed to display the correlation between copy number variants of STON1 and immune cells using a two-sided Wilcoxon rank-sum test [22].
Analysis of the value of STON1 in predicting immunotherapy and targeted therapy response in KIRC
The enrichment score of 19 ICB-response-associated gene signatures downloaded from the previous literature were calculated with the ssGSEA algorithm [21]. These signatures contain 18 positive pathways and one negative pathway (Supplemental Table 3). We further verified the correlation between STON1 and the expression of each immune checkpoint biomarkers with Gene_Corr module in the TIMER2.0 database. The different IC50 values between the low and the high STON1 group were calculated using the Assistant for Clinical Bioinformatics (https://www.aclbi.com/) with the IC50 module.
Validation STON1 expression in normal renal tubular epithelial cell line and KIRC cell lines
Normal renal tubular epithelial cell line HK-2, KIRC cell line A498, ACHN and 786-O were presented by the Department of Urology, Affiliated Hospital of Guangdong Medical University. All cell lines were maintained in Dulbecco's modified eagle medium (ThermoFisher Scientific China, Shanghai) supplemented with 10% fetal bovine serum (ThermoFisher Scientific China), and cells were cultured at 37 °C, 5% CO2 in saturation humidity. qRT-PCR and western blotting were used to detect the expression of STON1. For qRT-PCR, the following primers pairs were used, STON1 (NM_001198595.2) upstream, 5’-GCCCAAATATTTCCTGCAGAGTC-3’, STON1 downstream, 5’-CTGAGGCCAGGAAGGTTCAG-3’; GAPDH (NM_002046.7) upstream, 5’-TCGGAGTCAACGGATTTGGT-3’, GAPDH downstream, 5’-TTCCCG TTCTCAGCCTTGAC-3’. For western blotting, the primary antibody against STON1 (Cat: Abp53586, Abbkine) and GAPDH (Cat: 2118, CST China, Shanghai) were used.
Statistical analysis
All mRNA expression data from public databases was normalized by log2 transition. A columnar scatter plot was created by GraphPad Prism (Version 8, GraphPad Software, CA, USA). The immunoreactive scores of STON1 in tissue arrays were calculated using the Wilcoxon signed-rank test. Unpaired t-test was adopted to compare STON1 mRNA levels in cell lines using GraphPad Prism (Version 8). The chi-squared test was performed using IBM SPSS Statistics (version 25) to analyze the association between the STON1 mRNA level and clinicopathological parameters. The correlation analysis between STON1, the mismatch repair system, and the TMB was performed using Pearson’s test in RStudio software (version 4.0.3). P < 0.05 was considered statistically significant.
Results
STON1 expression was significantly reduced in KIRC
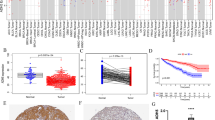
We first identified the transcription level of STON1 in different TCGA tumors with the TIMER2.0 database. A total of 33 human cancer types were explored. We found that most cancers including KIRC, kidney renal papillary cell carcinoma (KIRP), kidney chromophobe (KICH), breast invasive carcinoma (BRCA), colon adenocarcinoma (COAD), lung adenocarcinoma (LUAD), lung squamous cell carcinoma (LUSC), prostate adenocarcinoma (PRAD), stomach adenocarcinoma (STAD), thyroid carcinoma (THCA), and uterine corpus endometrial carcinoma (UCEC)—showed a decreased STON1 mRNA level, while head and neck squamous cell carcinoma (HNSC) displayed an upregulated STON1 mRNA level (Fig. 1a). Moreover, results from three GEO datasets, GSE16441, GSE16449, and GSE71963, also revealed aberrantly downregulated STON1 mRNA in KIRC (P < 0.01, Fig. 1b). Finally, the protein level of STON1 in KIRC was also found to be reduced using the CPTAC database (P < 0.0001) (Fig. 1c). We further validated the protein expression with our tissue microarray, and the results showed that KIRC samples displayed significantly lower immunoreactive scores compared with paired adjacent normal tissue (P < 0.0001, Fig. 2a, b). According to the results of in vitro cytological experiments, STON1 was again found to be downregulated in KIRC cell lines compared with normal renal tubular epithelial cell HK-2 (Fig. 2c, d, Supplementary Fig. 1).

STON1 expression is downregulated in KIRC. A STON1 mRNA expression in tumor and normal tissues of different human cancer types in TCGA database (the Wilcoxon test). B STON1 mRNA level in KIRC tumors and adjacent normal tissue in GSE16441, GSE16449, and GSE71963 from GEO database. C Protein expression of STON1 in CPTAC samples. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001

Expression of STON1 in KIRC tissue microarray and cell lines. A Representative images of STON1 protein in clinical KIRC and control tissues. B The immunoreactive score of STON1 presented by boxplot with t-test. ****P < 0.0001. C Expression of STON1 mRNA in normal renal tubular epithelial cells and KIRC cell lines. qRT-PCR was used to detect the expression of STON1 mRNA in normal renal tubular epithelial cell HK-2 and KIRC cell line A498, ACHN and 786-O. ****P < 0.0001. D Representative western blotting bands for STON1 protein in normal renal tubular epithelial cell HK-2 and KIRC cell line A498, ACHN and 786-O. For the original bands, please see Additional file 4
Correlation between STON1 expression and clinicopathological characteristics of KIRC patients
The KIRC GDC Expression Matrix was downloaded from the UCSC Xena, which is one of the most comprehensive clinical oncology databases [23]. A total of 491 KIRC patients were included in the cohort after excluding non-KIRC diagnoses and those with missing clinical information. They were classified into either a high expression group (n = 246) or a low expression group (n = 245) according to the median value of STON1. Notably, the STON1 mRNA levels were related to grade (P = 0.022), TNM stage (P = 0.039), distant metastasis (P = 0.019), and vital status (P = 5.46 × 10−6). However, no correlation was found between STON1 mRNA levels and age (P = 0.136), gender (P = 0.949), invasion depth (P = 0.074), and lymph node metastasis (P = 0.385) (Table 1). To further analyze the prognostic value of STON1, we performed a survival analysis using three oncological databases. The results indicated that STON1 expression levels were strongly associated with overall survival (OS) in KIRC. Patients with high expression of STON1 had a better outcome compared with those with low levels of STON1 in the OncoLnc database (logrank P = 9.78 × 10−8) (Fig. 3a). Similar results were acquired from the Kaplan–Meier database, and GEPIA2 with TCGA data (Fig. 3b–c).

High expression of STON1 indicated a better outcome in KIRC patients. Kaplan–Meier curves of overall survival according to high and low expression of STON1 in TCGA data in KIRC patients by exploring the OncoLnc database (A), Kaplan–Meier database (B), and GEPIA2 database (C)
The prognostic value of STON1 based on the levels of infiltration of various immune cells
Kaplan–Meier survival curves of the differential expression of STON1 between KIRC cohorts either enriched with immune cells or with decreased immune cells were explored. The high STON1 groups of KIRC cohorts with decreased CD4 + memory T cells (HR = 0.45, P = 0.0084), natural killer T cells (HR = 0.66, P = 0.031), regulatory T cells (Treg cell, HR = 0.48, P = 0.00011), type 1 T-helper cells (HR = 0.55, P = 0.00025), and type 2 T-helper cells (HR = 0.53, P = 0.00024), indicating favorable prognosis (Fig. 4). Similarly, high STON1 levels in the KIRC cohorts enriched with B cells (HR = 0.44, P = 9.8 × 10−5), CD4 + memory T cells (HR = 0.49, P = 0.00016), CD8 + T cells (HR = 0.46, P = 1.4 × 10−5), and macrophages (HR = 0.45, P = 4.4 × 10−6) exhibited a better OS, except natural killer T cells and type 1 T-helper cells (Fig. 5). Interestingly, high STON1 expression in KIRC with enriched type 1 T-helper cells displayed a poor OS, although it was not statistically significant (HR = 2.55, P = 0.067) (Fig. 5).

The prognostic value of STON1 in cases with decreased numbers of eight immune cells

The prognostic value of STON1 in cases enriched with eight immune cells
Low STON1 expression forms an immune inflamed phenotype in KIRC
Given that STON1 play totally different role in certain immune environment, we further analyzed the potential association between STON1 and immune cells. The majority of the cancer-related immune cell scores for B cells, CD4 + effector/memory T cells, CD8 + T cells, CD8 + effector/memory T cells, class-switched memory B cells, macrophages, M1 macrophages, M2 macrophages, memory B cells, monocytes, native B cells, natural killer (NK) T cells, plasmacytoid dendritic cells, plasma B cells, CD4 + T-helper 1 (Th1) cells and CD4 + T-helper 2 (Th2) cells were lower in the high STON1-expressing KIRC group (Fig. 6a–d). Additionally, we noted the immune score was higher in the low STON1 group, whereas the stroma score was relatively downregulated (Fig. 6d). To further confirm the relationship between STON1 expression and immune cell infiltration levels in KIRC, we used cBioportal database to explore the correlations between STON1 expression and various immune infiltration associated markers [24]. Our results showed there was a significant correlation between STON1 expression and the most of biomarker sets of CD8 + T cell, T cell (general), B cell, TAM, M1 macrophage, M2 macrophage, Th1, Th2, Tfh, Th17, Treg, T cell exhaustion (Supplemental Table 4). Cancer-Immunity Cycle is a multistep fine-regulated network, so we explored the function of STON1 in anti-tumor immune activities to compare the difference between the low and high STON1 groups. As shown in Fig. 7a, the low STON1 group manifested an intensive effect of anti-cancer immunity activities, especially priming and activation, the recruiting of B cells, CD8 T cells, dendritic cells, macrophages, myeloid derived suppressor cells (MDSCs), neutrophils, NK cells, T cells, and Th1 cells, and the recognition of cancer cells by T cells. The heatmap also indicated that STON1 expression was obviously negatively related to most of these cancer-immunity cycle pathway signatures (Fig. 7b, Supplemental Table 5). These results imply that low STON1 prompts an immune inflamed phenotype in KIRC. Furthermore, a previous report had proposed that activation of stromal pathways results in immunosuppressive effects of anti-cancer immunity [21]. Consistent with the above results, the enrichment scores of the EMT1 pathway, the EMT3 pathway, and the pan-F-TBRS pathway were significantly higher in the high STON1 group (Fig. 7c).

The correlation between STON1 and the tumor microenvironment. A–D The percentage abundance of tumor-infiltrating immune cells in each sample, with different colors and types of immune cells. (The ordinate represents the percentage of immune cell content in a single sample; high, high expression of STON1 group; low, low expression of STON1 group; the Wilcoxon test). *P < 0.05, **P < 0.01, ***P < 0.001; ns, not statistically significant

The different immune phenotypes between the low and high STON1 groups. A Enrichment score of cancer-immunity cycle steps between the two groups. (High, high expression of the STON1 group; low, low expression of the STON1 group; ssGSEA analysis and the Wilcoxon test; the abscissa represents different anti-tumor immune cycle steps and the ordinate represents the enrichment score based on special gene signatures; **P < 0.01; ***P < 0.001; ****P < 0.0001; ns, not statistically significant). B The correlation between STON1 expression and the enrichment score of cancer-immunity cycle. C Enrichment score of four stromal pathways between the two groups. (High, high expression of STON1 group; low, low expression of STON1 group; GSVA analysis and the Wilcoxon test; **P < 0.01; ****P <0.0001; ns, not statistically significant)
Correlations of STON1 levels with TMB, mismatch repair genes, CNV, and tumor stem index in KIRC
Given the sensitive connection between TMB, microsatellite instability (MSI), and immunotherapeutic response, we focused on their relationship with STON1 expression levels. Marginal scatter plots revealed that alterations in STON1 expression were always accompanied by changes in mismatch repair genes (MLH1, PMS2, MSH2, MSH6, and EpCAM). Moreover, the four mismatch genes seemed to be positively associated with STON1 (Fig. 8a, P < 2.2 × 10−16). The STON1 mRNA level was weakly negatively associated with the TMB score (P = 0.012) (Fig. 8b). In addition, the infiltration level of six immune cells was distinctly downregulated by arm-level deletion of STON1 (Fig. 9a). Cancer stem cells (CSCs) are characterized by the ability to generate all cell types in specific cancer samples. Compared with non-tumor stem cells, CSCs have self-protection mechanisms, such as DNA damage repair and inhibition of apoptotic pathways, that in turn lead to tumor progression, metastasis, drug resistance and increased self-renewal [25]. We found that the low STON1 group displayed a higher mRNAsi score (Fig. 9b), a marker associated with malignant biological processes in CSCs and more tumor dedifferentiation [26].

Correlation between STON1 and the mismatch system and tumor mutational burden (TMB). A The STON1 mRNA level was positively associated with MLH1, PMS2, MSH2, MSH6, and EpCAM, as presented in marginal scatter plots. B Relationship between the STON1 mRNA level and TMB in marginal scatter plots. The expression values of all genes are presented as logarithmic values

Association between copy number variants (CNV) of STON1, immune cells, and cancer stem index. A The infiltration level of six immune cell subtypes related to STON1 CNV. B The stem characteristics of tumor cells between the low STON1 group and the high STON1 group. *P < 0.05, ***P < 0.001, ****P < 0.0001
Decreased STON1 predicted a better response to ICB
The immune inflamed tumor phenotype is essential for KIRC patient response to ICB [27, 28]. We further analyzed the different ICB response between the low and high STON1 groups based on the efficacy of ICB-response-related signatures. The enrichment score for pathways related to a positive response to ICB was higher in the low STON1 group, such as RNA degradation, spliceosome, DNA replication, mismatch repair, and nucleotide excision repair (Fig. 10a). In contrast, the enrichment score for cytokine–cytokine receptor interaction, giving a negative response to ICB, was upregulated in the high STON1 group (Fig. 10a). Moreover, correlation analysis showed that STON1 expression was negatively correlated with the enrichment score of most of the eighteen ICB-response-related pathways (Fig. 10b, Supplemental Table 6). Given that immune checkpoints play an indispensable role in immunotherapy [29], our study found that STON1 expression was negatively correlated with the expression of immune checkpoints such as LAG3 (Lymphocyte activating 3), LGALS3 (Galectin 3), PDCD1 (Programmed cell death 1), and CTLA4 (Cytotoxic T-lymphocyte associated protein 4) (Fig. 11).

Sensitivity difference to ICB response between two groups. A The heatmap of the enrichment score of 19 special signatures for ICB response between the two groups. (High, high expression of STON1 group; low, low expression of STON1 group; ssGSEA analysis and the Wilcoxon test; the bar plot on the right represents the enrichment score based on special gene signatures). B The correlation between STON1 expression and the enrichment score of 19 special signatures for ICB response

The relationship between STON1 expression and four immune checkpoints
The high STON1 group was more sensitive to targeted treatment of KIRC
Currently, molecular targeting drugs for metastatic KIRC mainly include inhibitors for vascular endothelial growth factor (VEGF) signaling (Sunitinib, Pazopanib, Axitinib, Sorafenib) and the mammalian target of rapamycin (mTOR) inhibitor (Temsirolimus) [30,31,32]. We therefore employed the disparate sensitivity of these agents between the low and high STON1 groups. We found that the half maximal inhibitory concentration (IC50) score of the low STON1 group was higher than that of the high STON1 group in KIRC patients treat with temsirolimus (highmean vs lowmean = -2.08 vs -2.01, P < 0.001), which indicated that the high STON1 group was more sensitive to this drug, as well as sunitinib (highmean vs lowmean = 2.48 vs 2.59, P < 0.0001), axitinib (highmean vs lowmean = 1.98 vs 2.12, P < 0.001), pazopamib (highmean vs lowmean = 3.28 vs 3.30, P < 0.001), sorafenib (highmean vs lowmean = 2.12 vs 2.19, P < 0.001) (Fig. 12, Supplemental Table 7).

The sensitivity for five targeted agents among the two groups in KIRC. High, high expression of STON1 group; low, low expression of STON1 group; the vertical axis represents the distribution of IC50 scores; ***P < 0.001, ****P < 0.0001
Discussion
Although early-stage KIRC patients can be cured by partial or complete nephrectomy with a better outcome, approximately 25% of KIRC patients still suffer from recurrence or metastasis [1, 5]. Recently, immune checkpoint blockers for cancer therapy, such as combination immunotherapy of nivolumab plus ipilimumab, have shown promise in KIRC patients with rapid progression in clinical individualized treatment [33], but some patients displayed a low or no immune response [34]. Therefore, further identification of novel biomarkers to predict the clinical therapy response and survival of KIRC patients remains challenging. In our current study, STON1, a protein-coding gene involved in vesicle transport, was identified as a target gene at the rs13405728 locus in polycystic ovary syndrome [35]. A previous study reported that alterations in focal adhesion dynamics, cellular motility, and signaling were induced by the absence of STON1 [11]. The above processes are closely correlated with tumor migration. However, to date, no reports have published the underlying functions of STON1 in cancer, especially immune functions. To our knowledge, this is the first report to reveal that STON1 is decreased in most human cancer types, including KIRC, and is strongly associated with grade, stage, distant metastasis, and vital status in KIRC. IHC analysis and western blotting further indicated the lower expression level of STON1 protein in KIRC tissues and cell lines. Furthermore, STON1 overexpression favored prognosis in KIRC. The above results indicate that STON1 may act as a tumor suppressor in KIRC.
Tumor immunotherapy is a prominent milestone of the oncotherapy era. Currently, many indicators have been used to predict the ICB treatment response, such as the number of tumor-infiltrating lymphocytes (TIL), the proportion of CD28 + , CD39 + , and CD96 + TILs, PD-L1 expression, PD-1 expression, the gut microbiome, the TMB, and MSI [19]. However, these indicators are not economical or effective to predict the immunotherapy response because of the complex detection process. With the advent of ICBs, there is an urgent need to determine novel markers to predict immunotherapy responses in clinical practice. In our study, STON1 was identified as a promising indicator to predict the ICB treatment response and prognosis for KIRC patients.
We preliminarily analyzed the prognostic value of STON1 enriched or decreased with eight different immune cells. Compared with KIRC cohorts with decreased immune cells, the high STON1 group enriched with the anti-cancer immune cells B cells, CD4 + T cells, CD8 + T cells, and macrophages demonstrated a favorable prognosis. In line with the above results, the high STON1 group with decreased levels of Treg or Th2 cells—which are considered immunosuppressors [36, 37]—had a more significant P value than the high STON1 group enriched with Treg cells or Th2 cells. Interestingly, the high STON1 group enriched with Th1 cells showed a poor OS, although the P value was not statistically significant. We speculate that this was because of inadequate samples of KIRC patients. Thus, we were interested in the correlation between STON1 and TME in KIRC.
CD4 + T cells and CD8 + T cells that contribute to an inflammatory environment act as guardians in anti-tumor immunity [38]. Interferon-γ producing Th1 cells secrete TNF, resulting in tumor destruction. Th1 cells can not only recruit NK cells and macrophages near to the tumor site to exert anti-tumor effects, but also act as enhancers to promote tumor-specific cytotoxic T-lymphocyte (CTL) responses [39]. In contrast, a previous report showed that CD4 + T lymphocytes in a Th2-type tumor microenvironment can promote metastasis by regulating the pro-tumor properties of tumor-associated macrophages (TAMs), as opposed to limiting or eradicating malignant cells by engaging cytotoxic mechanisms [37]. We further analyzed the different infiltration levels of 35 immune cell subtypes between the low STON1 and high STON1 groups. Compared with the high group, most infiltration levels of the 35 immune cells were higher in the low STON1 group, especially for CD8 + T cells, NK T cells, and CD4 + Th1 T cells, as well as the immune score. Furthermore, tumors infiltrated with more activated immune cells (such as CD8 + T and Th1 cells) could have a better response to immunotherapy [40,41,42]. In the current study, we primarily speculated that the low STON1 group enriched with immune cells may be a good indicator for ICB treatment. Meanwhile, arm-level deletion of STON1 also significantly affected the abundance level of immune cells. Additionally, the activities of the cancer-immunity cycle were intense in the low STON1 group, including priming and activation, recruitment of B cells, CD8 T cells, dendritic cells, and macrophages, and the infiltration of immune cells into tumors. In contrast, the epithelial–mesenchymal translation pathways and the pan-fibroblast TGF-b response signature (Pan-FTBRS) with immunosuppressive effects [21] were significantly activated in the high STON1 group. In brief, low STON1 expression shapes an immune inflamed phenotype in KIRC which is essential for immunotherapy.
Mismatch repair genes are related to the human mismatch repair response, which is responsible for the repair of base mismatches that occur during DNA replication [43]. TMB is a novel indicator of mutation quantity [44]. Tumors with mismatch repair possess the capacity to increase the mutational burden, resulting in increased immune checkpoint protein expression, including PD-1 and PD-L1. Furthermore, the immunogenic neoepitopes generated by the imbalance of the mismatch system significantly improved the immunotherapy response rate [45]. Interestingly, the mRNA level of STON1 was positively associated with mismatch repair (MMR) genes in our current study. As the STON1 expression decreased, the TMB score increased. Hence, STON1 may be a candidate prognostic biomarker for evaluating the efficacy of immunotherapy responses and further experiments should be conducted to confirm this theory.
KIRC is one of the most immune-infiltrated cancers of all human solid cancer types [46]. The emergence of immune checkpoint targets provides a new direction for tumor immunotherapy. In recent years, CTLA4 inhibitors, PD-l inhibitors, PD-L1 inhibitors, and PD-L1 inhibitors plus tyrosine kinase inhibitors (TKI) in progressed KIRC patients have been approved by the FDA and achieved great curative effect [47]. We conducted a comprehensive bioinformatics analysis to reveal the detail of STON1 levels as an indicator for ICB response in KIRC. A total of 19 ICB-response-related signatures were employed to predict different response activities between the two groups. As expected, STON1 was significantly negatively correlated with 18 pathways that reflect a positive response to ICB and was positively associated with the cytokine–cytokine receptor interaction that is negative for ICB therapy. In addition, the immune checkpoint is necessary for the response to immunotherapy. CTLA4 inhibits T-cell activation to some extent, thus, anti-CTLA4 antibody immunotherapy can enhance the anti-tumor immune effects of T cells [48]. An anti-PD1 (PDCD1) agent series is approved by the FDA and used in various carcinomas with better immune response [49]. Our results indicated that low STON1 expression correlated with the high expression of these immune checkpoints. Furthermore, we focused on several targeted agents, including temsirolimus, sunitinib, axitinib, pazopamib, sorafenib and found that the IC50 score of these drugs were significantly higher in the high STON1 group which indicated that targeted therapy could be a treatment option for the high STON1 group. In summary, ICB application is more suitable for the low STON1 group, whereas cases with high expression of STON1 are more sensitive to targeted treatment.
We attempted to validate the expression and prognostic value of STON1 in our clinical KIRC cohorts and cell lines. Our results also showed that STON1 protein was downregulated in KIRC tissues compared with normal controls. In vivo experiments, we also found that STON1 was down-regulated in KIRC cell lines compared to normal renal tubular epithelial cells both at mRNA and protein levels. However, because of the incomplete clinical parameters in our tumor cohort, especially the small number of samples in the death group in the follow-up data, we could not obtain satisfactory statistical results. Therefore, a clinical cohort with a larger sample size is required to validate these consequences, and further cytological functional studies need to verify the specific role and molecular mechanism of STON1 in KIRC in future. Of notable, we detected STON1 protein with the molecular mass between 100 to 130 kDa but not the classical 83 kDa in the KIRC cell lines, the related spicing mechanism need illuminate in future.
Conclusions
Collectively, lower STON1 expression in KIRC indicates tumor progression and worse survival outcome. Our current study provides new insights into the function of STON1 in the tumor microenvironment. High STON1 expression may shape an immune non-inflamed phenotype. Importantly, STON1 is relatively suitable as an indicator of ICB and targeted therapy response.
Availability of data and materials
The datasets used and/or analyzed during the current study are available in the following repositories, GEO database (https://www.ncbi.nlm.nih.gov/geo/), accession numbers: GSE16441, GSE16449 and GSE71963, UCSC Xena (https://xena.ucsc.edu/), TCGA database (https://portal.gdc.cancer.gov/).
Abbreviations
- CNV:
-
Copy number variant
- EMT:
-
Epithelial–mesenchymal transition
- GEO:
-
Gene Expression Omnibus
- GEPIA:
-
Gene Expression Profiling Interactive Analysis
- GSVA:
-
Gene Set Variation Analysis
- ICB:
-
Immune checkpoint blockage
- IHC:
-
Immunohistochemical staining
- KIRC:
-
Kidney renal clear cell carcinoma
- MMR:
-
Mismatch repair
- mRNAsi:
-
mRNA stemness index
- NG2:
-
Neuron-glial antigen 2
- OS:
-
Overall survival
- Pan-FTBRS:
-
Pan-fibroblast TGF-β response signature
- RCC:
-
Renal cell carcinoma
- ssGSEA:
-
Single sample Gene Set Enrichment Analysis
- STON1:
-
Stonin 1
- TCGA:
-
The Cancer Genome Atlas
- TIMER:
-
The Tumor Immune Estimation Resource
- TMB:
-
Tumor mutational burden
- TME:
-
Tumor microenvironment
References
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69(1):7–34.
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424.
Moch H, Cubilla AL, Humphrey PA, Reuter VE, Ulbright TM. The 2016 WHO Classification of Tumours of the Urinary System and Male Genital Organs-Part A: Renal, Penile, and Testicular Tumours. Eur Urol. 2016;70(1):93–105.
Prasad SR, Humphrey PA, Catena JR, Narra VR, Srigley JR, Cortez AD, et al. Common and uncommon histologic subtypes of renal cell carcinoma: imaging spectrum with pathologic correlation. Radiographics. 2006;26(6):1795–806 (discussion 1806-1710).
Hsieh JJ, Purdue MP, Signoretti S, Swanton C, Albiges L, Schmidinger M, et al. Renal cell carcinoma. Nat Rev Dis Primers. 2017;3:17009.
Jonasch E, Gao J, Rathmell WK. Renal cell carcinoma. Bmj. 2014;349:g4797.
Barata PC, Rini BI. Treatment of renal cell carcinoma. Current status and future directions. CA Cancer J Clin. 2017;67(6):507–24.
Maritzen T, Podufall J, Haucke V. Stonins–specialized adaptors for synaptic vesicle recycling and beyond? Traffic. 2010;11(1):8–15.
Martina JA, Bonangelino CJ, Aguilar RC, Bonifacino JS. Stonin 2: an adaptor-like protein that interacts with components of the endocytic machinery. J Cell Biol. 2001;153(5):1111–20.
Almén MS, Jacobsson JA, Moschonis G, Benedict C, Chrousos GP, Fredriksson R, et al. Genome wide analysis reveals association of a FTO gene variant with epigenetic changes. Genomics. 2012;99(3):132–7.
Feutlinske F, Browarski M, Ku MC, Trnka P, Waiczies S, Niendorf T, et al. Stonin1 mediates endocytosis of the proteoglycan NG2 and regulates focal adhesion dynamics and cell motility. Nat Commun. 2015;6:8535.
Li T, Fu J, Zeng Z, Cohen D, Li J, Chen Q, et al. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020;48(W1):W509-w514.
Barrett T, Wilhite SE, Ledoux P, Evangelista C, Kim IF, Tomashevsky M, et al. NCBI GEO: archive for functional genomics data sets–update. Nucleic Acids Res. 2013;41(Database issue):D991-995.
Chandrashekar DS, Bashel B, Balasubramanya SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi B, et al. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia. 2017;19(8):649–58.
Liang L, Cen H, Huang J, Qin A, Xu W, Wang S, et al. The reversion of DNA methylation-induced miRNA silence via biomimetic nanoparticles-mediated gene delivery for efficient lung adenocarcinoma therapy. Mol Cancer. 2022;21(1):186.
Nagy Á, Munkácsy G, Győrffy B. Pancancer survival analysis of cancer hallmark genes. Sci Rep. 2021;11(1):6047.
Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(W1):W98-w102.
Cerami E, Gao J, Dogrusoz U, Gross BE, Sumer SO, Aksoy BA, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012;2(5):401–4.
Chen DS, Mellman I. Oncology meets immunology: the cancer-immunity cycle. Immunity. 2013;39(1):1–10.
Xu L, Deng C, Pang B, Zhang X, Liu W, Liao G, et al. TIP: A Web Server for Resolving Tumor Immunophenotype Profiling. Cancer Res. 2018;78(23):6575–80.
Mariathasan S, Turley SJ, Nickles D, Castiglioni A, Yuen K, Wang Y, et al. TGFβ attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature. 2018;554(7693):544–8.
Li T, Fan J, Wang B, Traugh N, Chen Q, Liu JS, et al. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017;77(21):e108–10.
Goldman MJ, Craft B, Hastie M, Repečka K, McDade F, Kamath A, et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nat Biotechnol. 2020;38(6):675–8.
Pan JH, Zhou H, Cooper L, Huang JL, Zhu SB, Zhao XX, et al. LAYN Is a Prognostic Biomarker and Correlated With Immune Infiltrates in Gastric and Colon Cancers. Front Immunol. 2019;10:6.
Walcher L, Kistenmacher AK, Suo H, Kitte R, Dluczek S, Strauß A, et al. Cancer Stem Cells-Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Front Immunol. 2020;11:1280.
Zhang Y, Tseng JT, Lien IC, Li F, Wu W, Li H. mRNAsi Index: Machine Learning in Mining Lung Adenocarcinoma Stem Cell Biomarkers. Genes (Basel). 2020;11(3):257.
Gajewski TF, Corrales L, Williams J, Horton B, Sivan A, Spranger S. Cancer Immunotherapy Targets Based on Understanding the T Cell-Inflamed Versus Non-T Cell-Inflamed Tumor Microenvironment. Adv Exp Med Biol. 2017;1036:19–31.
Ji RR, Chasalow SD, Wang L, Hamid O, Schmidt H, Cogswell J, et al. An immune-active tumor microenvironment favors clinical response to ipilimumab. Cancer Immunol Immunother. 2012;61(7):1019–31.
Safarzadeh A, Alizadeh M, Beyranvand F, FalavandJozaaee R, Hajiasgharzadeh K, Baghbanzadeh A, et al. Varied functions of immune checkpoints during cancer metastasis. Cancer Immunol Immunother. 2021;70(3):569–88.
Méjean A, Ravaud A, Thezenas S, Colas S, Beauval JB, Bensalah K, et al. Sunitinib Alone or after Nephrectomy in Metastatic Renal-Cell Carcinoma. N Engl J Med. 2018;379(5):417–27.
Escudier B, Szczylik C, Hutson TE, Demkow T, Staehler M, Rolland F, et al. Randomized phase II trial of first-line treatment with sorafenib versus interferon Alfa-2a in patients with metastatic renal cell carcinoma. J Clin Oncol. 2009;27(8):1280–9.
Hutson TE, Lesovoy V, Al-Shukri S, Stus VP, Lipatov ON, Bair AH, et al. Axitinib versus sorafenib as first-line therapy in patients with metastatic renal-cell carcinoma: a randomised open-label phase 3 trial. Lancet Oncol. 2013;14(13):1287–94.
Motzer RJ, Tannir NM, McDermott DF, ArenFrontera O, Melichar B, Choueiri TK, et al. Nivolumab plus Ipilimumab versus Sunitinib in Advanced Renal-Cell Carcinoma. N Engl J Med. 2018;378(14):1277–90.
Signoretti S, Flaifel A, Chen YB, Reuter VE. Renal Cell Carcinoma in the Era of Precision Medicine: From Molecular Pathology to Tissue-Based Biomarkers. J Clin Oncol. 2018;36(36):JCO2018792259.
Cao CH, Wei Y, Liu R, Lin XR, Luo JQ, Zhang QJ, et al. Three-Dimensional Genome Interactions Identify Potential Adipocyte Metabolism-Associated Gene STON1 and Immune-Correlated Gene FSHR at the rs13405728 Locus in Polycystic Ovary Syndrome. Front Endocrinol (Lausanne). 2021;12:686054.
Tanaka A, Sakaguchi S. Targeting Treg cells in cancer immunotherapy. Eur J Immunol. 2019;49(8):1140–6.
DeNardo DG, Barreto JB, Andreu P, Vasquez L, Tawfik D, Kolhatkar N, et al. CD4(+) T cells regulate pulmonary metastasis of mammary carcinomas by enhancing protumor properties of macrophages. Cancer Cell. 2009;16(2):91–102.
Chen DS, Mellman I. Elements of cancer immunity and the cancer-immune set point. Nature. 2017;541(7637):321–30.
Kennedy R, Celis E. Multiple roles for CD4+ T cells in anti-tumor immune responses. Immunol Rev. 2008;222:129–44.
Desbois M, Udyavar AR, Ryner L, Kozlowski C, Guan Y, Dürrbaum M, et al. Integrated digital pathology and transcriptome analysis identifies molecular mediators of T-cell exclusion in ovarian cancer. Nat Commun. 2020;11(1):5583.
Zhang B, Wu Q, Li B, Wang D, Wang L, Zhou YL. m(6)A regulator-mediated methylation modification patterns and tumor microenvironment infiltration characterization in gastric cancer. Mol Cancer. 2020;19(1):53.
Hornburg M, Desbois M, Lu S, Guan Y, Lo AA, Kaufman S, et al. Single-cell dissection of cellular components and interactions shaping the tumor immune phenotypes in ovarian cancer. Cancer Cell. 2021;39(7):928-944.e926.
Luchini C, Bibeau F, Ligtenberg MJL, Singh N, Nottegar A, Bosse T, et al. ESMO recommendations on microsatellite instability testing for immunotherapy in cancer, and its relationship with PD-1/PD-L1 expression and tumour mutational burden: a systematic review-based approach. Ann Oncol. 2019;30(8):1232–43.
Jardim DL, Goodman A, de Melo GD, Kurzrock R. The Challenges of Tumor Mutational Burden as an Immunotherapy Biomarker. Cancer Cell. 2021;39(2):154–73.
Dudley JC, Lin MT, Le DT, Eshleman JR. Microsatellite Instability as a Biomarker for PD-1 Blockade. Clin Cancer Res. 2016;22(4):813–20.
Vuong L, Kotecha RR, Voss MH, Hakimi AA. Tumor Microenvironment Dynamics in Clear-Cell Renal Cell Carcinoma. Cancer Discov. 2019;9(10):1349–57.
Hirsch L, Flippot R, Escudier B, Albiges L. Immunomodulatory Roles of VEGF Pathway Inhibitors in Renal Cell Carcinoma. Drugs. 2020;80(12):1169–81.
Egen JG, Ouyang W, Wu LC. Human Anti-tumor Immunity: Insights from Immunotherapy Clinical Trials. Immunity. 2020;52(1):36–54.
Sharpe AH, Pauken KE. The diverse functions of the PD1 inhibitory pathway. Nat Rev Immunol. 2018;18(3):153–67.
Acknowledgements
We thank Catherine Perfect, MA (Cantab), from Liwen Bianji (Edanz) (www.liwenbianji.cn/), for editing the English text of a draft of this manuscript.
Funding
This work was supported by grants from the YangFan Plan of Guangdong Province (4YF16007G). The funders had no role in the design of the study, collection, analysis, and interpretation of data, or writing the manuscript.
Author information
Authors and Affiliations
Contributions
Conception and design: WJ, BTL and ZHS, Administrative support: WJ, BTL and ZHS, Provision of study materials patients: JRB, YPH, Collection and assembly of data: AXZ, YPY, YHF, Data analysis and interpretation: AXZ, JRB, MHL, YDL, ZHS, Manuscript writing: All authors, Final approval of manuscript: All authors.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplemental Table 1.
The clinicopathological data of tissue microarray.
Additional file 2: Supplemental Table 2.
The gene sets for the cancer-immunity cycle analysis.
Additional file 3: Supplemental Table 3.
The signatures related to response of immunotherapy.
Additional file 4: Supplemental Figure 1.
Original bands for western blotting of STON1 and GAPDH in cell lines.
Additional file 5: Supplemental Table 4.
Correlation analysis between STON1 and relate genes and markers of immune cells in KIRC by cBioportal database.
Additional file 6: Supplemental Table 5.
Correlations between STON1 and anti-cancer immunity in TCGA cohort.
Additional file 7: Supplemental Table 6.
Correlations between STON1 and immunotherapy associated signatures in TCGA cohort
Additional file 8: Supplemental Table 7.
The IC50 score between high and low expression of STON1 groups in KIRC.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zheng, A., Bai, J., Ha, Y. et al. Integrated analysis of the relation to tumor immune microenvironment and predicted value of Stonin1 gene for immune checkpoint blockage and targeted treatment in kidney renal clear cell carcinoma. BMC Cancer 23, 135 (2023). https://doi.org/10.1186/s12885-023-10616-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12885-023-10616-9