
Abstract
Introduction
Difference in clinical responses to cancer therapy in each patient is from several factors. Gastrointestinal microbiota is one of the reasons. However, this correlation remains unknown. This study aims to explore correlation between gastrointestinal microbiota profile and clinical outcomes in Thai advanced non-small cell lung cancer (NSCLC) according to epidermal growth factor receptor (EGFR) status.
Methods
We enrolled 13 patients with advanced EGFR–wild-type (WT) NSCLC who received chemotherapy and 15 patients with EGFR-mutant NSCLC who received EGFR tyrosine kinase inhibitors. We collected fecal samples at baseline and first disease evaluation and performed 16S rRNA gene sequencing by NGS to assess microbiota profile. The correlations between gastrointestinal microbiota and clinical variables were studied.
Results
The clinical characteristics were balanced between the cohorts, excluding significantly higher albumin levels in the EGFR-mutant group. Albumin was the only significant clinical factor affecting the treatment response in multivariate analysis (ORR 15.6%, P = 0.03). Proteobacteria counts were higher in the EGFR-WT group, whereas Bacteroidetes and Firmicutes counts were higher in the EGFR-mutant group. The alpha diversity of the gastrointestinal microbiome was significantly higher in the EGFR-mutant group (Shannon index: 3.82 vs. 3.25, P = 0.022). Following treatment, Proteobacteria counts were lower and Bacteroidetes and Firmicutes counts were higher in both cohorts; the changes were more prominent in the EGFR-WT cohort. No significant correlation between microbiota profile and treatment response were demonstrated in our study. However, beta diversity was significantly different according to severity of adverse events. Enrichment of Clostridia and Bacteroidia was associated with higher adverse event risk in the EGFR-WT cohort.
Conclusions
Proteobacteria was dominant in Thai lung cancer patients both EGFR-WT and EGFR-mutant, and this phylum maybe associate with lung cancer carcinogenesis. Chemotherapy altered the gastrointestinal microbiota, whereas EGFR-TKIs had less effects. Our findings highlight the potential predictive utility of the gastrointestinal microbiota for lung cancer carcinogenesis. Studies with larger cohorts and comparison with the healthy Thai population are ongoing to validate this pilot study.
Similar content being viewed by others


Introduction
Non-small cell lung cancer (NSCLC) accounts for approximately 80–85% of all lung cancers worldwide [1]. In Thailand, NSCLC is the most common cancer and the second most common cause of cancer-related death after biliary tract cancer [2, 3]. Although advancements in NSCLC treatment have prolonged survival, platinum-based doublet chemotherapy, which is a mainstay of treatment for patients with advanced NSCLC in developing countries [4], carries an overall response rate (ORR) of approximately 30–40% and overall survival (OS) of 8–12 months [5]. Furthermore, Asian patients with lung adenocarcinoma have a high prevalence of epidermal growth factor receptor (EGFR) mutation (50%) [6]. In tyrosine kinase inhibitor (TKI)-sensitive EGFR-mutant NSCLC, EGFR-TKIs significantly improve survival and ORRs (60–70%) [7, 8]. However, some patients with TKI-sensitive EGFR-mutant NSCLC do not respond to EGFR-TKIs. In addition, patients with EGFRexon 19 deletion appear to have a longer duration of response and higher ORRs [7, 8]. To date, no predictive biomarkers of the efficacy and toxicity of chemotherapy and EGFR-TKIs in the treatment of NSCLC have been identified.
The human microbiota, which consists of more than 1000 species, approximately 97% of which reside in the intestine, is a key focus of global research [9]. Research on bacterial 16S ribosomal RNA using metagenomics technology including next-generation sequencing (NGS) has provided several insights into the human microbiota. The gastrointestinal microbiota can affect host immune responses in various ways. Previous research reported that the gastrointestinal microbiota plays an important role in drug efficacy and toxicity in response to therapy [10,11,12,13,14]. Conversely, dysbiosis can affect patient outcomes. Several factors can alter the microbiome composition such as medications, diet, prebiotics, probiotics [15,16,17,18,19,20,21], and cancer therapies. Several studies found that chemotherapies, such as oxaliplatin and cyclophosphamide, can directly cause epithelial damage. Translocation of gastrointestinal microbes and their biologic products across the gastrointestinal epithelium can stimulate T cell expansion in lymphoid tissue [9, 22, 23] and result in different immune responses that might affect cancer. Inhibition of EGFR in the gastrointestinal tract leads to changes in intestinal architecture, thereby resulting in microbiota changes by increasing in bowel movement frequency [24]. Understanding the interactions among the gastrointestinal microbiota, therapies, and cancer response is necessary for the development of microbiota-targeting approaches to improve treatment efficacy.
Most previous studies [12,13,14] in various tumor types involved patients who received immunotherapy. In these studies, certain microbes and high microbial diversity were related to different clinical outcomes. There is limited knowledge regarding the gastrointestinal microbiota profile of patients with advanced EGFR-WT and EGFR-mutant lung cancer who received chemotherapy and targeted therapy.
This study explored the gastrointestinal microbiota profiles of patients with advanced EGFR-WT and EGFR-mutant NSCLC who received chemotherapy and EGFR-TKIs, respectively. We analyzed gastrointestinal microbiota profiles before and after treatment initiation and their associations with clinical features, especially treatment outcomes.
Materials and methods
Patients and study design
This prospective study enrolled treatment-naïve patients with stage III–IV EGFR-WT or EGFR-mutant NSCLC in Ramathibodi Hospital, Mahidol University (Bangkok, Thailand) between June 2020 and January 2021. The eligibility criteria were an age of at least 18 years and Eastern Cooperative Oncology Group performance status (ECOG PS) of 0–2. The diagnosis of NSCLC was on the basis of histopathological analysis, and stage was classified according to the TNM classification (AJCC 8th edition). The EGFR status was confirmed in all patients using tissue samples by real-time PCR (Therascreen, Qiagen). Patients underwent stool collection after receiving an explanation of the process. All participants had measurable disease according to computed tomography or magnetic resonance imaging using Response Evaluation Criteria in Solid Tumors version 1.1 (RECIST 1.1). This study and all experimental protocols were approved by The Human Research Ethics Committee of Ramathibodi Hospital, Mahidol University, Bangkok, Thailand with the IRB number of COA. MURA2021/305. All methods were carried out in accordance with relevant guidelines and local regulations. Informed consent was obtained from all patients in this study.
Patients were classified into two cohorts: the EGFR-WT cohort, in which patients were treated with standard platinum-based doublet chemotherapy regimens every 3 weeks for 4–6 cycles or until unacceptable toxicity; and the EGFR-mutant cohort, in which patients received any-generation EGFR-TKIs until progression or until unacceptable toxicity. The response to treatment was evaluated 8–12 weeks after treatment initiation. In this study, patients were categorized as responders and non-responders. In the EGFR-WT cohort, responders comprised patients with complete responses (CRs), partial responses (PRs), and stable disease (SD), and patients with progressive disease (PD) were classified as non-responders. In the EGFR-mutant cohorts, responders included patients with CR or PR, whereas those with SD or PD were classified as non-responders.
Procedures
Informed consent was received from all eligible patients. Paired fecal samples were collected from each patient at baseline (before treatment initiation) and at the first disease evaluation (2–3 months after treatment initiation). The proper size of fecal samples was a grain of rice (50–100 mg/time point), and samples were collected using StoolFiX™ (Isohelix, Cell Projects) containing 500 μL of lysis buffer and 25 μL of proteinase K and stored at room temperature until DNA extraction.
Clinical data (age, sex, underlying disease, smoking status, performance status, body weight, body mass index, type of lung cancer, type of EGFR mutation, metastatic site, laboratory results, and history of gastrointestinal tract surgery) were collected at baseline by reviewing the Electronic Medical Record (EMR). We also interviewed the patients in person for the history of antibiotics, proton pump inhibitors, prebiotics, or probiotics usage within 1 month prior starting the treatment. During the treatment, we collected the response and adverse events (AEs) of treatment of each patient. AEs were assessed and graded according to the Common Terminology Criteria for Adverse Events (CTCAE) version 4. CTCAE grading ≥ 2 AEs were categorized as severe AEs, and CTCAE grading 0–1 were classified as mild AEs.
Bacterial fecal DNA extraction
Bacterial fecal DNA extraction was performed at the Center for Medical Genomics, Faculty of Medicine, Ramathibodi Hospital, Mahidol University. The fecal sample was mixed with 20 μL of PK solution and incubated in a 60◦C water bath for 1 h. Then, the sample tubes were centrifuged at 12,000 × g for 10 min. The supernatant was removed, and then the tubes were briefly re-spun to remove any remaining supernatant. Next, 100 μL of elution buffer were added. The subsequent extraction procedure was performed according to the manufacturer’s instruction. Metagenomic DNA was extracted using a QIAamp PowerFecal DNA Kit (QIAGEN, Germany). DNA quality and quantity were assessed using a NanoDrop One spectrophotometer (Thermo Fisher Scientific, USA) and Qubit® 3.0 fluorometer (Thermo Fisher Scientific).
Polymerase chain reaction (PCR) and sequencing
To determine the bacterial composition in feces, construction of an Illumina metagenomic sequencing library was performed via two-step PCR. The first PCR was performed to amplify the V3–V4 hypervariable region of the bacterial 16S rRNA gene using the primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). The second PCR was performed to add the index sequences for the Illumina sequencer with the barcode sequence using a Nextera XT Index kit (Illumina, San Diego, CA, USA). The prepared libraries were subjected to sequencing of the paired-end 2 × 250 bases using a MiSeq Reagent Kit v2 on the MiSeq (Illumina).
Bioinformatics analysis
Raw sequencing data were cleaned using FLASH software and analyzed using Quantitative Insights into Microbial Ecology 2 (QIIME 2) [25]. Demultiplexed cleaned sequence data were denoised using DADA2, and then the amplicon sequence variant (ASVs) counts per sample was generated and grouped by different taxonomic levels (phylum, class, order, family, genus, species). Taxonomic assignment was based on the SILVA 138 ribosomal (rRNA) database.
The diversity within the gastrointestinal community was assessed via alpha and beta diversity analyses. To assess alpha diversity, Shannon’s index, which reflects the complexity of species diversity within samples, was calculated. To assess beta diversity, principal coordinate analysis (PCoA) was performed using the Bray–Curtis index to compare community dissimilarity. Linear discriminant analysis with effect size (LEfSe) was used to compared our groups of interest to identify biomarkers and visualize the result using a taxonomic bar chart [26].
Statistical analysis
The patients’ baseline characteristics were reported using descriptive statistics. Comparisons between the EGFR-WT and EGFR-mutant cohorts were performed using the Chi-squared or Fisher’s exact test for categorical variables and Student’s t-test or the Mann–Whitney U test for continuous variables as appropriate. Progression-free survival (PFS) was estimated using the Kaplan–Meier method as the time from diagnosis to disease progression or death from any cause. The associations of clinical variables with alpha diversity were explored using univariate and multivariate linear regression analysis. The associations of clinical variables including alpha diversity with the response to treatment were analyzed using univariate and multivariate logistic regression analysis and presented as odds ratios (ORs) with 95% confidence intervals (CIs) and P-values. All statistical analyses were performed using Stata version 16. P < 0.05 indicated statistical significance.
Results
Patient characteristics and clinical outcomes
From June 2020 to January 2021, 42 patients were screened. Twenty-eight patients (13 with EGFR-WT NSCLC and 15 with EGFR-mutant NSCLC) met the study’s criteria and finally included in this analysis. (Supplemental Fig. 1 ).

Relative abundance of gastrointestinal microbiota phyla at baseline in the EGFR-WT and EGFR-mutant cohorts. EGFR-wt: epidermal growth factor receptor wild-type; EGFR-mut: epidermal growth factor receptor mutant
In the EGFR-WT cohort, the median patient age was 63 years. Most patients were women (69.2%) and never-smokers (76.9%). Nearly half of the patients had normal BMI (46.1%), and 92.2% had an ECOG PS of 0–1 (Table 1). Hypertension (46.1%) was the most common underlying disease, followed by diabetes mellitus (17.8%). Adenocarcinoma was the most common histopathologic subtype (84.6%), whereas two patients had squamous histopathology. All patients received carboplatin/paclitaxel as the first-line chemotherapy.
In the EGFR-mutant cohort, the median patient age was 64 years. Most patients were men (66.6%) and never-smokers (66.6%). Nearly half of the patients had normal BMI (46.6%), and 14 (93.3%) patients had an ECOG PS of 0–1. Most patients had adenocarcinoma (93.3%), and one patient had adenosquamous histology. Hypertension (40.0%) was the most common underlying disease, followed by dyslipidemia (40.0%). Exon 19 deletion and exon 21 L858R mutations were the most common EGFR mutations in this population (86.6%). The remaining patients had uncommon EGFR mutations. Half of the patients (53.2%) received first-generation EGFR-TKIs, whereas 13.3% received third-generation EGFR-TKIs.
The most common metastatic sites in both cohorts were the pleura and lungs. The EGFR-mutant cohort had a significantly better nutritional status (albumin ≥ 3.2 g/dL) than the EGFR-WT cohort (80.0% vs. 30.7%, P = 0.02). Other clinical factors were similar between the two cohorts (Table 1).
The response to treatment in the EGFR-WT cohort was as follows: PR, 7.6%; SD, 38.4%; and PD, 53.8%. Meanwhile, the response in the EGFR-mutant cohort was as follows: PR, 66.6%; SD, 26.6%; and PD, 6.67% Thus, the response rates in the EGFR-WT and EGFR-mutant cohorts were 7.6 and 66.6%, respectively (Supplemental Table 1). Furthermore, median PFS was 3.5 months in the EGFR-WT cohort, versus 7.9 months in the EGFR-mutant cohort.
Albumin was significantly predictive of the response of treatment in both univariate and multivariate analyses. Patients with albumin ≥ 3.2 g/dL were significantly more likely to respond to treatment than those with albumin < 3.2 g/dL (ORR = 15.7, 95% CI = 1.3–182, P = 0.028, Table 2). In univariate analysis, patients with EGFR-mutant NSCLC were also more likely to respond to treatment than those with EGFR-WT NSCLC (ORR = 24.0, 95% CI = 2.4–240.6, P = 0.0007), whereas the small size of the EGFR-WT cohort precluded multivariate analysis.
Most patients in the EGFR-WT cohort (61.5%) experienced severe AEs (Common Terminology Criteria for Adverse Events: CTCAE ≥ 2), and 53.3% of patients in the EGFR-mutant cohort experienced all grades AEs. The most common AEs in the EGFR-WT cohort were anemia (92.3%) and vomiting (25.0%), whereas those in the EGFR-mutant cohort were rash (60.0%), anemia (53.0%), and diarrhea (20%). All patients in EGFR-mutant cohort developed mild degree of adverse events (CTCAE < 2), there was no severe AEs. There were no significant differences in antibiotic, proton pump inhibitor, and laxative use between the cohorts.
Gastrointestinal microbiota in patients with lung cancer at baseline
In the entire population, the five most abundant bacteria at the phylum level at baseline were Proteobacteria (53.9%), Bacteroidetes (23.7%), Firmicutes (16.4%), Fusobacteriota (2.5%), and Verrucomicrobia (1.5%) (Fig. 1). The relative abundance of these bacteria was similar between the groups, but Proteobacteria counts were higher in the EGFR-WT cohort whereas Bacteroidetes and Firmicutes counts were higher in the EGFR-mutant cohort, albeit without significance.
Compositional differences of the gastrointestinal microbiota according to the response to treatment
We found some alterations in the microbiota composition between before and after treatment. After patients with EGFR-WT NSCLC received chemotherapy, the relative abundance of Proteobacteria decreased (46.2%), whereas that of Bacteroidetes (25.5%) and Firmicutes (24.1%) increased. There was no distinct change in the microbiota composition in the EGFR-mutant cohort (Fig. 2).

Comparison of relative abundance of gut microbiota phyla before treatment (T1) and after treatment (T2). A. EGFR-WT cohort B. EGFR-mutant cohort
To investigate whether the microbiota composition was associated with the response to chemotherapy or EGFR-TKIs, we descriptively compared the relative abundance of gastrointestinal bacteria at the phylum level between responders and non-responders. The results revealed no significant differences between responders and non-responders in both cohorts (Supplement Fig. 2A and B). However, we observed that Desulfobacterota was enriched (21.1%) among partial responders in the EGFR-WT cohort, whereas Actinobacteria was enriched in patients with PD in the EGFR-mutant cohort (Supplemental Fig. 3A and B).

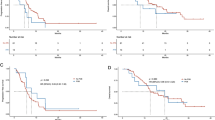
Comparison of alpha diversity in EGFR-WT and EGFR-mutant cohorts. A Shannon index in every patient at baseline B. Shannon index between 2 cohorts at baseline. B Shannon index in every patient after treatment D. Shannon index between 2 cohorts after treatment. EGFR-wt: epidermal growth factor receptor wild-type; EGFR-mut: epidermal growth factor receptor mutant
Differences of gastrointestinal microbiota diversity
Shannon index (alpha diversity) was calculated to show species abundance in this population. The results revealed that the alpha diversity of the gastrointestinal microbiota was significantly higher in the EGFR-mutant cohort than that in the EGFR-WT cohort at baseline (3.85 vs. 3.25, P = 0.022, Fig. 3). Consistent with the results of multivariate analysis, the EGFR mutation status was significantly associated with the Shannon index (Table 3). Univariate linear regression analysis between clinical factors and the Shannon index revealed that gender and the EGFR mutation status were significantly associated with the Shannon index. Specifically, women had a lower Shannon index than men (3.21 ± 0.63 vs. 3.86 ± 0.55, P = 0.008), whereas patients with EGFR-mutant NSCLC had a higher Shannon index than those with EGFR-WT NSCLC. After adjustment for potential confounding factors in multivariate linear regression analysis, only the EGFR status remained significantly associated with the Shannon index (3.79 ± 0.59 vs. 3.25 ± 0.65, P = 0.047).
We also explored the potential of the Shannon index to predict treatment responses, but no association was identified in univariate or multivariate logistic regression analysis.
At the time of disease evaluation, we observed a non-significant increase of the Shannon index in the EGFR-WT cohort, whereas the index was similar between baseline and the time of disease evaluation in the EGFR-mutant cohort (Fig. 3). Following treatment, the difference in the Shannon index between the two cohorts had dissipated (P = 0.495, Supplement Fig. 4 ). Beta diversity was similar between the two cohorts at both baseline and the time of disease evaluation. We further investigated the microbiota diversity between responders and non-responders and found no significant differences in alpha or beta diversity in both cohorts.

Relative abundance of gut microbiota phyla between less severe (CTCAE grading < 2) and severe adverse events (CTCAE grading ≥ 2). A EGFR-WT cohort B EGFR-mutant cohort. CTCAE: Common Terminology Criteria for Adverse Events; EGFR: epidermal growth factor receptor; WT: wild-type
Association between the gastrointestinal microbiota profile and AEs
We next examined the potential association of the microbiota composition with AEs during treatment. The microbiota profile was compared between patients with mild and severe AEs at both baseline and the time of disease evaluation. Differences in the relative abundance of microbes were observed in both groups at baseline, but the differences were more prominent in the EGFR-WT group. Proteobacteria were enriched in patients with mild AEs, whereas Bacteroidetes and Firmicutes were enriched in patients with severe AEs (Fig. 4). The Shannon index was numerically but not significantly higher in patients with severe AEs. We further conducted PCoA to assess beta diversity. The results revealed significant differences in microbial communities in patients with mild and severe AEs in the EGFR-WT cohort (Fig. 5).

Comparison of alpha and beta diversity (Principal Coordinates Analysis plot; PCoA plot) between CTCAE grading < 2 and CTCAE grading ≥ 2 in EGFR-WT cohort. A Shannon index in each sample B Shannon index between CTCAE grading < 2 and CTCAE grading ≥ 2. C. Beta diversity (PCoA plot). CTCAE: Common Terminology Criteria for Adverse Events; EGFR-WT: epidermal growth factor receptor wild-type; PCoA plot: Principal Coordinates Analysis plot
We performed LEfSE analysis to identified biomarkers for AEs at the class level. In the EGFR-WT group, the abundance of Gammaproteobacteria was higher in patients with mild AEs (LDA score = 6), whereas those of Clostridia (LDA score = − 5.8) and Bacteroidia (LDA score = − 6) were higher in patients with severe AEs (Fig. 6). The fecal microbiota profile was similar between patients with mild and severe AEs in the EGFR-mutant cohort.

Linear discrimination analysis (LDA) identify significant microbiota between CTCAE grading < 2 and CTCAE grading ≥ 2 in EGFR-WT cohort A. Phylum level B. Class level. CTCAE: Common Terminology Criteria for Adverse Events; EGFR-WT: epidermal growth factor receptor wild-type; LDA: Linear discrimination analysis
Discussion
Our study investigated the gastrointestinal microbiota in Thai patients with advanced EGFR-WT (received platinum-based doublet chemotherapy) and EGFR-mutant NSCLC (received EGFR-TKIs). First, we explored the composition of the gastrointestinal microbiota at baseline. Generally, the gastrointestinal tract is dominated by members of four bacterial phyla: Bacteroidetes, Firmicutes, Proteobacteria, and Actinobacteria [27,28,29]. These data correspond with those in healthy Thai people [30, 31]. Our study found that Proteobacteria was the most abundant phylum in Thai lung cancer patients, whereas the abundance of Actinobacteria was diminished in both the EGFR-WT and EGFR-mutant cohorts. Alteration of the gastrointestinal microbiota composition has been linked to the pathogenesis of several metabolic and inflammatory diseases including cancer. Shin et al. [29] also observed a higher abundance of Proteobacteria in patients with inflammatory conditions and cancer. In mouse models, abnormal expansion of Proteobacteria is associated with dysregulation of both innate and adaptive immune responses because of interference with interleukin and CD4 + T cell function. Dysbiosis in mice increases the risks of inflammatory bowel disease and colitis-associated colorectal cancer. Moreover, previous data revealed that Firmicutes promotes colonic luminal short-chain fatty acid and modulates inflammation in mice and humans. Decreases in the abundance of Firmicutes may be associated with lung cancer development [32]. Actinobacteria, which comprises a large proportion of the healthy human microflora, has potent cancer-suppressing activity because of its metabolites [32]. Thus, the increased abundance of Proteobacteria and decreased abundance of Firmicutes or Actinobacteria at the phylum level relative to those in lung cancer patients could be emerged as a potential biomarkers of lung carcinogenesis, but we need to have the further exploration in the larger cohort, together with the Thai healthy people cohort in the future. Furthermore, we also found that the abundance of Proteobacteria was decreased and that of Bacteroidetes and Firmicutes was increased after treatment in both cohorts, although the changes were stronger in patients in the EGFR-WT cohort, who received chemotherapy. It has been reported that chemotherapy can alter the gastrointestinal microbiota of patients with cancer by disrupting the gastrointestinal barrier and promoting subsequent bacterial translocation [33]. Consistently, our results revealed distinctive changes in the microbiota composition after treatment compared with the findings before treatment using fecal samples. The EGFR-TKI, osimertinib, did not significantly change the relative abundance of gastrointestinal microbiota in a previous study [34], in line with our findings in the EGFR-mutant cohort. This result is probably attributable to the lack of a direct cytotoxic effect of EGFR-TKIs on the gastrointestinal even though one of their common AEs is diarrhea. Only three patients (20%) in our EGFR-mutant cohort had diarrhea (one patient with severe diarrhea), indicating a lack of microbial composition changes in the EGFR-mutant cohort.
Dysbiosis predisposes people to cancer by causing direct DNA damage via toxins, altering the regulation of immune and pro-inflammatory pathways, and stimulating the production of carcinogenic metabolites, dysbiosis also can potentially affect the response to cancer therapy [35]. In our study, alpha diversity at baseline significantly differed between the cohorts. A lower alpha diversity index suggests the presence of fewer species in the gastrointestinal tract in the EGFR-WT cohort, consistent with the differential microbiota composition between the two cohorts (Fig. 6B). Beta diversity as visualized using PCoA also revealed some differences in the microbiota between the EGFR-WT and EGFR-mutant cohorts, albeit without significance. Previous research revealed no significant reduction in the alpha diversity of the gastrointestinal in patients with lung cancer compared to healthy individuals. However, they did not show the difference between EGFR-WT and EGFR-mutant cancer, as shown in our study [34, 36]. This result is important for explaining the differences in biology between these lung cancer types.
Previous studies described a relationship between high microbiota diversity and better clinical responses to treatment with immune checkpoint inhibitors. The improved response was associated with increases of memory CD8 + T cell and natural killer cell counts in patients with in high microbiota diversity [12, 13]. However, there were no significant differences in alpha and beta diversity between responders and non-responders in both cohorts in our study, probably because of the small numbers of patients in each group.
The most common AEs in the EGFR-WT cohort were anemia (92.3%) and vomiting (25.0%). Interestingly, we observed a high abundance of Clostridia and Bacteroidia at baseline in patients who experienced severe AEs in the EGFR-WT cohort. Previous studies found Bacteroidia and Clostridia species as potentially potential enteropathogenic bacteria usually associated with diarrhea and other gastrointestinal toxicities [37, 38]. Thus, these two classes of bacteria are likely associated with the treatment toxicity, and may be used as biomarkers of chemotherapy-induced toxicity, although further research is needed.
In addition, our study explored the clinical factors associated with the response to treatment. Interestingly, we found that serum albumin level, which reflects the nutritional status of patients, affected the outcome of treatment. Patients with serum albumin level ≥ 3.2 g/dL had significantly higher treatment response rates than those with low albumin. This was consistent with previous studies identifying that the pre-treatment serum albumin level was a prognostic biomarker in cancer treatment [39]. However, most EGFR-mutant patients have higher serum albumin level, thus, EGFR status may be a confounding factor for analyzing correlation of clinical factors and response to treatment.
Smoking is another confounding factor in this study. 20–30% of populations have prior smoking history. Toxic chemical exposure such as nicotine, aldehydes, polycyclic aromatic hydrocarbons result in intestinal irritation and impaired mucosal immune responses. Proteobacteria increase in number with high number of pack-years of cigarette smoking in systematic review [40].
One limitation of this study was its small sample size, which might explain the findings concerning the associations of the gastrointestinal microbiota with treatment responses and AEs. More participants are needed to confirm whether the gastrointestinal microbiota can predict treatment response or toxicity.
In conclusion, this is the first study of the gastrointestinal microbiota in patients with EGFR-WT and EGFR-mutant lung cancer. Our study indicated that lung cancer disease might affect the gastrointestinal microbiota at baseline in composition and diversity. Proteobacteria was dominant in our cohort, and this phylum maybe associate with lung cancer carcinogenesis. Chemotherapy altered the gastrointestinal microbiota, whereas EGFR-TKIs had less effects. The findings of our pilot study highlight the potential predictive utility of the gastrointestinal microbiota for lung cancer carcinogenesis, particularly concerning cancer prevention and treatment. Studies with larger cohorts and comparison with the healthy Thai population are ongoing to confirm and further explore the gastrointestinal microbiota and its association with lung cancer.
Availability of data and materials
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Society TAC. Key Statistics for Lung Cancer 2020. https://www.cancer.org/cancer/lung-cancer/about/key-statistics.html.
Reungwetwattana T, Oranratnachai S, Puataweepong P, Tangsujaritvijit V, Cherntanomwong P. Lung cancer in Thailand. J Thorac Oncol. 2020;15(11):1714–21.
Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71:209.
National Comprehensive Cancer Network. Non-small cell lung cancer (version 4.2021). https://www.nccn.org/professionals/physician_gls/pdf/nscl.pdf. Accessed 3 Mar 2021.
Schiller JH, Harrington D, Belani CP, et al. Comparison of four chemotherapy regimens for advanced non-small-cell lung cancer. N Engl J Med. 2002;346(2):92–8.
Shi Y, Au JS, Thongprasert S, et al. A prospective, molecular epidemiology study of EGFR mutations in Asian patients with advanced non-small-cell lung cancer of adenocarcinoma histology (PIONEER). J Thorac Oncol. 2014;9(2):154–62.
Mok TS, Wu YL, Thongprasert S, et al. Gefitinib or carboplatin-paclitaxel in pulmonary adenocarcinoma. N Engl J Med. 2009;361(10):947–57.
Ramalingam SS, Vansteenkiste J, Planchard D, et al. Overall survival with osimertinib in untreated, EGFR-mutated advanced NSCLC. N Engl J Med. 2020;382(1):41–50.
Locey KJ, Lennon JT. Scaling laws predict global microbial diversity. Proc Natl Acad Sci U S A. 2016;113(21):5970–5.
Alexander CMO, Cody GD, De Gregorio BT, Nittler LR, Stroud RM. The nature, origin and modification of insoluble organic matter in chondrites, the possibly interstellar source of Earth’s C and N. Chem Erde. 2017;77(2):227–56.
Cong J, Zhu J, Zhang C, et al. Chemotherapy alters the phylogenetic molecular ecological networks of intestinal microbial communities. Front Microbiol. 2019;10:1008.
Gopalakrishnan V, Spencer CN, Nezi L, et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science. 2018;359(6371):97–103.
Jin Y, Dong H, Xia L, et al. The diversity of gut microbiome is associated with favorable responses to anti-programmed death 1 immunotherapy in Chinese patients with NSCLC. J Thorac Oncol. 2019;14(8):1378–89.
Routy B, Le Chatelier E, Derosa L, et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science. 2018;359(6371):91–7.
Bajaj JS. Alcohol, liver disease and the gut microbiota. Nat Rev Gastroenterol Hepatol. 2019;16(4):235–46.
Hakozaki T, Okuma Y, Omori M, Hosomi Y. Impact of prior antibiotic use on the efficacy of nivolumab for non-small cell lung cancer. Oncol Lett. 2019;17(3):2946–52.
Hasan N, Yang H. Factors affecting the composition of the gut microbiota, and its modulation. PeerJ. 2019;7: e7502.
Hemarajata P, Versalovic J. Effects of probiotics on gut microbiota: mechanisms of intestinal immunomodulation and neuromodulation. Therap Adv Gastroenterol. 2013;6(1):39–51.
Imhann F, Bonder MJ, Vich Vila A, et al. Proton pump inhibitors affect the gut microbiome. Gut. 2016;65(5):740–8.
Takagi T, Naito Y, Inoue R, et al. The influence of long-term use of proton pump inhibitors on the gut microbiota: an age-sex-matched case-control study. J Clin Biochem Nutr. 2018;62(1):100–5.
Venkataraman A, Sieber JR, Schmidt AW, Waldron C, Theis KR, Schmidt TM. Variable responses of human microbiomes to dietary supplementation with resistant starch. Microbiome. 2016;4(1):33.
Almeida A, Mitchell AL, Boland M, et al. A new genomic blueprint of the human gut microbiota. Nature. 2019;568(7753):499–504.
Sepich-Poore GD, Zitvogel L, Straussman R, Hasty J, Wargo JA, Knight R. The microbiome and human cancer. Science. 2021;371(6536):eabc4552.
Secombe KR, Van Sebille YZA, Mayo BJ, Coller JK, Gibson RJ, Bowen JM. Diarrhea Induced by Small Molecule Tyrosine Kinase Inhibitors Compared With Chemotherapy: Potential Role of the Microbiome. Integr Cancer Ther. 2020;19:1534735420928493.
Bolyen E, Rideout JR, Dillon MR, et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol. 2019;37(8):852–7.
Chong J, Liu P, Zhou G, Xia J. Using Microbiome Analyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat Protoc. 2020;15(3):799–821.
Budden KF, Gellatly SL, Wood DL, et al. Emerging pathogenic links between microbiota and the gut-lung axis. Nat Rev Microbiol. 2017;15(1):55–63.
Rajilic-Stojanovic M, de Vos WM. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS Microbiol Rev. 2014;38(5):996–1047.
Shin NR, Whon TW, Bae JW. Proteobacteria: microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015;33(9):496–503.
Gruneck L, Kullawong N, Kespechara K, Popluechai S. Gut microbiota of obese and diabetic Thai subjects and interplay with dietary habits and blood profiles. PeerJ. 2020;8: e9622.
La-Ongkham O, Nakphaichit M, Nakayama J, Keawsompong S, Nitisinprasert S. Age-related changes in the gut microbiota and the core gut microbiome of healthy Thai humans. 3 Biotech. 2020;10(6):276.
Zheng Y, Fang Z, Xue Y, et al. Specific gut microbiome signature predicts the early-stage lung cancer. Gut Microbes. 2020;11(4):1030–42.
Alexander JL, Wilson ID, Teare J, Marchesi JR, Nicholson JK, Kinross JM. Gut microbiota modulation of chemotherapy efficacy and toxicity. Nat Rev Gastroenterol Hepatol. 2017;14(6):356–65.
Cong J, Zhang Y, Xue Y, et al. A Pilot Study: Changes of Intestinal Microbiota of Patients With Non-small Cell Lung Cancer in Response to Osimertinib Therapy. Front Microbiol. 2020;11: 583525.
Parida S, Sharma D. The power of small changes: Comprehensive analyses of microbial dysbiosis in breast cancer. Biochim Biophys Acta Rev Cancer. 2019;1871(2):392–405.
Zhuang H, Cheng L, Wang Y, et al. Dysbiosis of the gut microbiome in lung cancer. Front Cell Infect Microbiol. 2019;9:112.
Stringer AM, Gibson RJ, Logan RM, Bowen JM, Yeoh AS, Keefe DM. Faecal microflora and beta-glucuronidase expression are altered in an irinotecan-induced diarrhea model in rats. Cancer Biol Ther. 2008;7(12):1919–25.
van Vliet MJ, Tissing WJ, Dun CA, et al. Chemotherapy treatment in pediatric patients with acute myeloid leukemia receiving antimicrobial prophylaxis leads to a relative increase of colonization with potentially pathogenic bacteria in the gut. Clin Infect Dis. 2009;49(2):262–70.
Gupta D, Lis CG. Pretreatment serum albumin as a predictor of cancer survival: a systematic review of the epidemiological literature. Nutr J. 2010;9:69.
Antinozzi M, Giffi M, Sini N, Galle F, Valeriani F, De Vito C, et al. Cigarette Smoking and Human Gut Microbiota in Healthy Adults: A Systematic Review. Biomedicines. 2022;10(2):510.
Acknowledgements
We thank Division of Medical Oncology, Department of Medicine and Center for Medical Genomics, Faculty of Medicine Ramathibodi Hospital, Mahidol University, Bangkok, Thailand for providing all samples and all tests for this research. We would like to thank all patients whom participated in this study.
Funding
This study was supported by the Thailand Center of Excellence for Life Science (TCELS).
Author information
Authors and Affiliations
Contributions
W.S. performed methodology, formal analysis, interpretation, investigation, data curation, writing-original draft, visualization. I.S. performed formal analysis, investigation, interpretation, resources, data curation. N.T. performed the validation, investigation, resources, project administration, analysis, interpretation. S.O. performed methodology, formal analysis, interpretation, investigation, data curation, writing-original draft. A.C. performed, validation, resources, project administration. C.R. performed investigation. N.M. performed collecting the samples. W.S. (Sukkasem) performed investigation, writing-review & editing, resources. P.I. perform the investigation, writing-review & editing, resources. T.S. performed the investigation, writing-review & editing, resources. P.C. performed study design, analysis, interpretation, conclusion, suggestion. T.A. performed writing-review & editing, resources. R.P. performed investigation, writing-review & editing, resources. W.C. performed supervision, funding acquisition, writing-review & editing. T.R. designed the study and performed the conceptualization, study design, methodology, formal analysis, interpretation, investigation, resources, data curation, writing-review & editing, visualization, supervision, funding acquisition. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study and all experimental protocols were approved by The Human Research Ethics Committee of Ramathibodi Hospital, Mahidol University, Bangkok, Thailand with the IRB number of COA. MURA2021/305. All methods were carried out in accordance with relevant guidelines and local regulations. Informed consent was obtained from all patients in this study.
Consent for publication
I am Thanyanan Reungwetwattana, the corresponding author of this study “Gastrointestinal Microbiota Profile and Clinical Correlations in Advanced EGFR-WT and EGFR-Mutant Non-Small Cell Lung Cancer”. All of the figures in this manuscript could not lead to identification of a study participants. Furthermore, on behalf of our research team, I would like to give my consent for information of this study to be published in BMC cancer. I understand that the text and any pictures published in the article will be freely available on the internet and may be seen by the general public.
Competing interests
All authors declare no competing interests to this study.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplemental Table 1.
Response of treatment. Supplemental Figure 1. Consort Diagram. Supplemental Figure 2. Comparison of relative abundance of gut microbiota phyla between responder (R) and non-responder (NR) A. EGFR-WT cohort B. EGFR-mutant cohort. Supplemental Figure 3. Bar chart of Phylogenetic composition of each patient according to response of treatment A. EGFR-WT cohort B. EGFR-mutant cohort. Supplemental Figure 4. Comparison of alpha diversity in responders (R) and non-responders (NR) in both cohorts A. EGFR-WT cohort B. EGFR-mutant cohort
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Saifon, W., Sensorn, I., Trachu, N. et al. Gastrointestinal microbiota profile and clinical correlations in advanced EGFR-WT and EGFR-mutant non-small cell lung cancer. BMC Cancer 22, 963 (2022). https://doi.org/10.1186/s12885-022-10050-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12885-022-10050-3