
Abstract
Background
The ability of Staphylococcus aureus to invade tissues and cause an infectious disease is the result of a multi-factorial process supported by the huge number of virulence factors inherent to this microorganism tightly regulated by the accessory gene regulator (agr). During antimicrobial therapy bacteria may be exposed to sub-inhibitory concentrations (subMICs) of antibiotics that may trigger transcriptional changes that may have an impact on the pathogenesis of infection. The objective of this study was to investigate the effect of oxacillin sub-MICs on agr system expression as the key component in the regulation of virulence in methicillin-susceptible (MSSA) and -resistant S. aureus (MRSA) strains. Furthermore, we studied the genetic basis of the agr locus and their potential association with the expression levels.
Methods
We have examined the expression of RNAIII and agrA mRNA as biomarkers for agr expression in the presence and absence of oxacillin subMICs in 10 MSSA and 4 MRSA clinical strains belonging to 5 clonal complexes (CC45-agrI, CC8-agrI, CC5-agrII, CC15-agrII and CC30-agrIII) causing endovascular complications. The DNA sequences of agr locus were obtained by whole genome sequencing.
Results
Our results revealed that exposure to subMICs of oxacillin had an impact on agr locus expression modifying the relative levels of expression with increases in 11 strains and with decreases in 3 strains. Thereby, the exposure to subMICs of oxacillin resulted in higher levels of expression of agr in CC15 and CC45 and lower levels in CC30. We also observed the presence of mutations in agrC and agrA in 13/14 strains with similar mutation profiles among strains within individual CCs except for strains of CC5. Although, agr expression levels differed among strains within CCs, the presence of these mutations was associated with differences in agr expression levels in most cases.
Conclusions
Changes in agr expression induced by exposure to oxacillin subMICs should be considered because they could lead to changes in the virulence modulation and have an adverse effect on the course of infection, especially in certain clonal complexes.
Similar content being viewed by others


Background
Staphylococcus aureus remains the major cause of endovascular infections with high morbidity and mortality rates that may reach 40% in case of infective endocarditis [1]. Even though most S. aureus genotypes exhibit the capacity to cause invasive disease, several studies have reported the association of particular clonal complexes (CC5, CC15 and CC30) with endovascular complications such as infective endocarditis [2,3,4].
The ability of S. aureus to produce infection involves a wide variety of virulence determinants, such as exotoxins, adhesins and immune evasion genes, constituting a gene framework that is under the control of global regulators, most notably the quorum-sensing accessory gene regulator (agr). Under conditions of high cell density, agr is responsible for the increased expression of many toxins and degradative exoenzymes, and decreased expression of several colonization factors. This regulation is important for the timing of virulence factor expression during infection [5]. Thereby, when adhesion to host tissue is crucial, cell density and agr expression are low, resulting in an increase in the surface virulence factors that are required for the colonization process. Once infection is established, the increased cell density results in higher agr expression that leads to production of toxins which trigger the host inflammatory response [6, 7]. The agr system up-regulates the production of secreted virulence factors such as exotoxins, and down-regulates the production of cell associated virulence factors [5, 8,9,10].
The agr locus consists of two divergent transcriptional loci, RNAII and RNAIII, driven by promoters P2 and P3, respectively [8, 9]. The P2 operon encodes four genes, agrB, agrD, agrC and agrA. The agrA and agrC genes constitute a two-component signaling system, of which agrC is the receptor, and agrA is the response regulator [11, 12]. Upon activation by agrC-dependent phosphorylation, agrA binds to the P2 promoter region for RNAII and the P3 promoter region for RNAIII, the effector of target gene regulation that also encodes delta-haemolysin, the expression of which serves as a surrogate for agr functionality [13, 14]. Moreover agrA directly up-regulates transcription of the phenol-soluble modulins operon (psmα and psmβ) whose role in the pro-inflammatory response has already been demonstrated [6, 7, 12, 15]. Several studies have reported that different mutations in the agr operon, mainly within the agrC or agrA genes, lead to decreased RNAIII expression and reduced virulence [9, 16,17,18]. However, it is known that strains with dysfunctional agr are more likely to cause persistent infection and be associated with poor outcome [19,20,21].
Despite the emergence of methicillin resistant S. aureus (MRSA), beta-lactams and specifically antistaphylococcal penicillins (nafcillin, oxacillin, cloxacillin and dicloxacillin), remain the usual treatment for the management of invasive infections caused by methicillin-susceptible S. aureus (MSSA). However, because of impaired diffusion and distribution of antibiotic, not all bacteria are exposed to the required lethal concentrations of bactericidal agents and therefore are likely subjected to sub-MIC antibiotic effects on the expression of virulence factors [7, 22,23,24,25,26,27,28,29,30]. Thereby, the effect of subMICs oxacillin on phenol-soluble modulins and RNAIII have been studied [27, 28], showing a decrease in mRNA level in both virulence determinants in USA300 strain. However, increase in spa and lukE mRNA levels under exposure to subMICs of oxacillin, cephalotin and penicillin were reported by Subrt et al. [29]. Moreover, Rasigade et al. revealed that subMIC of oxacillin, moxifloxacin and linezolid led to increase fnbA/B mRNA levels [30]. Collectively all this studies add more evidence that, that suboptimal concentrations of different antibiotics modulate the expression of virulence factors in S. aureus, and therefore may exert an influence over the pathogenesis of infection [23]. Nevertheless, limited data exist showing the effect of oxacillin in MSSA clinical strains. Thereby, the aim of the present study was to investigate the impact of sub-inhibitory concentrations (subMICs) of oxacillin on the agr operon expression, as a key element of virulence regulatory network, in a collection of S. aureus clinical strains belonging to the main clonal complexes related with invasive diseases. In addition, we analyzed whether the presence of mutations in agr locus could be associated with differences in agr gene expression.
Methods
S. aureus strains collection
A total of 14 S. aureus strains (10 MSSA and 4 MRSA) from two multi-center investigations of infective endocarditis [31] and catheter-related bacteremia [32] were included in the present study. The strains were selected as representatives of the CCs that were most frequently detected in the original investigations: CC5 (ST5 and ST125), CC15, CC30, CC45 and CC8 (ST8 and ST72). We randomly selected two MSSA strains of each CC, as well as methicillin resistant (MRSA) strains that belonged to the same CCs if they were available (Table 1).
Sub-inhibitory concentrations of oxacillin and growth curves
Strains were exposed to subMICs of oxacillin in order to observe agr gene expression under this condition. To select the optimal subMIC for each strain, we monitored growth by measuring the optical density of broth cultures exposed to ½, ¼ and 1/8 of the respective MIC for each strain as previously determined by E-test (bioMérieux). We defined the subMIC as the highest concentration below the MIC that showed no major growth defects over the entire growth curve relative to a control culture without oxacillin. [27] For each strain, the MIC and subMIC of oxacillin are shown in Table 1. Oxacillin was added to cultures at the time of inoculation (1:100 from pre-cultures) into 10 ml of tryptic soy broth (TSB) and incubated at 37 °C in a shaking incubator at 250 rpm. Growth under the same conditions was monitored by determining the optical density at 600 nm (OD600) and it was measured at the end of the incubation to determine the impact of antibiotic exposure over bacterial density (Additional file 1: Figure S1).
RNA isolation
Isolated colonies were resuspended in TSB and adjusted to 0.5 McFarland and incubated at 37 °C overnight. Fresh TSB (10 ml in 50-ml conical flask) was inoculated with 100 μl of overnight broth cultures (1:100 dilution) and incubated at 37 °C in a shaking incubator at 250 rpm in the presence or absence of the appropriate subMIC of oxacillin. Growth was monitored by determining the optical density (OD600). RNA was isolated after growth for 20 h (stationary phase) using the Qiagen Rneasy Mini kit according to the manufacturer’s instructions and after addition of 2 volumes of RNA protect reagent (Qiagen, N.V).
Reverse transcription and qRT-PCR
Expression of both the agrA and RNAIII genes was analyzed by quantitative reverse transcriptase PCR (qRT–PCR) as surrogate biomarkers of global agr operon activation. Expression levels of the housekeeping gyrB gene were used to normalize differences in mRNA quantification. In brief, 1 μg of total RNA was transcribed into cDNA using a Transcriptor First Strand cDNA Synthesis Kit (Roche Applied Science). Quantitative real time PCR was carried out on a LightCycler® 480 with LightCycler® 480 FastStart DNA Master Plus SYBR Green I (both Roche Applied Science). Previously described primers were used to amplify the RNAIII, agrA and gyrB transcripts [28, 33]. The experiments were performed using three biological replicates, each tested in triplicate. For each reaction, the ratio of the target gene (RNAIII or agrA) to gyrB transcripts was calculated based on the difference in cycle thresholds (2Ct_gyrB-Ct_target_gene) for all strains to obtain RNAIII and agrA relative gene expression levels in stationary phase in the presence and absence of the subMIC of oxacillin [33, 34]. The results were expressed as the n-fold difference in levels of transcription of the 2Ct_gyrBoxa-Ct_agrAoxa and 2Ct_gyrBoxa-Ct_RNAIIIoxa relative to 2Ct_gyrBcontrol_Ct_agrAcontrol and 2Ct_gyrBcontrol-Ct_RNAIIIcontrol, respectively.
Genotyping analysis of agr locus by whole genome sequencing
The DNA sequences of agr locus were obtained after that assembling of reads obtained from whole genome sequencing of all strains included in this study before of exposure to oxacillin subMICs. The sequencing was performed on the Illumina MiSeq platform (150 bp paired-end reads) using TruSeq DNA PCR free kits (Illumina, CA, USA). These agr nucleotide sequences of the entire agr locus were compared with those of agr group reference strains -agr I: NCTC 8325, agr II: N315; agr III: MW2- by multiple alignments using the Geneious server (Geneious 10.0.6).
Statistical analysis
In order to yield more accurate and reliable summary statistics continuous variables were expressed as mean and standard deviation (SD) from the expression data obtained of three biological replicates each tested in triplicate. These values were analyzed by Student’s t or Wilcoxon test, as appropriate. Significance was defined as p < 0.05. Data were stored and analyzed using SPSS software version 15.0 (SPSS, Chicago, IL, USA).
Nucleotide sequence accession number
The nucleotide sequences were deposited in GenBank (NCBI) under the accession numbers: MKYX00000000, MKYY00000000, MKYZ00000000, MKZA00000000, MKZB00000000, MKZC00000000, MKZD00000000 MKZE00000000, MKZF00000000, MKZG00000000, MKZH00000000, MKZI00000000, MKZJ00000000 and MKZK00000000.
Results
Expression of agr locus in the presence of oxacillin subMICs in stationary phase and its relationship with the CC, agr type and methicillin resistance
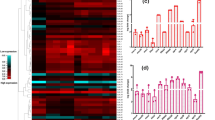
The exposure to subMICs of oxacillin in stationary phase resulted in a trend to higher RNAIII transcript levels than in absence of the antimicrobial agent for 11/14 strains being statistically significant for 5 strains of them (p < 0.05) (Fig. 1). Only SA_123 (CC45) (p: 0.735), SA_107 (CC30) (p: 0.028) and SA_170006 (CC5) (p: 0.043) exhibited reduced RNAIII expression (Table 2). We observed a similar pattern of relative gene expression for agrA, with increased levels of agrA mRNA in the presence of oxacillin subMICs for all strains, being statistically significant for 5 of them (p < 0.05), except SA_80001 (CC30) (p: 0.028), SA_107 (CC30) (p: 0.345) and SA_103 (CC5) (p: 0.465) which showed lower levels of expression (Fig. 1, Table 2, Additional file 2: Table S1). In addition, we observed that in four strains (SA_123, SA_103, SA_170006 and SA_80001), the change in the expression of RNAIII and agrA did not move in the same direction. However, these differences in the expression values for both agrA and RNAIII, in the presence and absence of oxacillin subMICs, were not statistically significant.

Effect of subMICs of oxacillin over RNAIII (a) and agrA (b) expression levels under oxacillin subMICs exposure in stationary growth phase. The expression levels of both agrA and RNAIII genes were determined by qRT–PCR experiments (three different experiments). The results are expressed as the n-fold variation of RNAIII (A) and agrA (B). An asterisk indicates a significant difference (P < 0.05)
In order to investigate the association between agr locus expression and methicillin resistance profile we studied the agr locus expression in the strains classified as MSSA and MRSA and we found higher agrA and RNAIII levels of gene expression for both groups in the presence of subMICs of oxacillin than in absence of this antibiotic, although this trend was not statistically significant (Table 3). When we analyzed by CC, we detected an increase in relative gene expression for RNAIII and agrA in the presence of oxacillin for both CC45 (p = 0.005 and p = 0.000, respectively) and CC15 (p = 0.005 and p = 0.001, respectively). Nevertheless, CC30 showed a decrease in gene expression for both RNAIII and agrA in the presence of subMICs of oxacillin, but the difference was not statistically significant.
With regard to agr type, agrII strains showed higher expression levels under oxacillin exposure for both RNAIII and agrA (Table 3).
Genotypic analysis of agr locus
After observing differences in the agr expression among the strains, we intended to know whether these strains harbored any mutations that might potentially be responsible for the agr functionality. In general, sequencing of the agr locus revealed similar mutation profiles among strains belonging to the same CC (Table 3). Strains from CC45-agrI showed up to 9 non-synonymous mutations in the agrB region and from 2 to 3 non-synonymous mutations in agrC, including missense mutations in SA_123 (417 Glu > Lys) and SA_170015 (103, Ile > Thr). Strains belonging to CC8-agrI exhibited mutations exclusively in the agrC and agrA genes. In the coding region of agrC, SA_190006 and SA_70002, displayed the same three non-synonymous mutations (42 Val > Gly; 262 Asp>Ala; 258 Phe > Ile). None of these mutations were present in SA_180015, although a frameshift deletion was detected (a409-1 bp, 138Lys/del). All CC5-agrII strains except SA_180009 displayed mutations: a single non-synonymous mutation (96, Ser > Leu) in SA_103, a frameshift one base pair deletion at position 313 in SA_80004, and a non-synonymous mutation at position 121 (121, Ile > Thr) in SA_170006. The two strains from CC15-agrII did not show any differences between them, with both exhibiting the same missense substitution at position 136 in agrA (136, Lys > Arg). No differences were found between the two CC30-agrIII strains, which showed the same genotypic profile with missense mutations in the coding region of agrB (49 Ile > Leu and 115 Ile > Leu) and agrC (55 Gly > Arg) (Table 4).
Relationship between mutation profile and expression of agr system
After analyzing both agrA and agrC genetic profile, we aimed to investigate the potential effect of these mutations on the expression of agr in stationary phase. Overall there were strain-to-strain differences in the relative levels of expression (Fig. 2), which were largely independent of CC, with only strains in CC5 all exhibiting similar levels of expression. Notably, the strains in CC5 also had the overall lowest levels of RNAIII and agrA expression (p = 0.024 and p = 0.016, respectively).

Relative expression of RNAIII (a) and agrA (b) in stationary growth phase. The expression levels of both agrA and RNAIII genes were determined by qRT–PCR experiments. The values are the means ± standard deviation (three different experiments)
Although it is difficult to determine the influence of mutations on the agr locus expression because of the variability found among strains, we studied whether the presence of the different mutations could have some impact on agr expression. In general, although some mutations detected did not seem to directly affect the expression of the agr system, the majority of them were associated with changes on the expression levels of RNAIII and/or agrA (Table 5). Thus, the mutation 42 Val > Gly found in the agrC in strains of CC45 and SA_190006 and SA_70002 of CC8 was associated with high expression levels for both RNAIII and agrA (specifically 5- and 6.9 fold, respectively). Likewise, mutations 262 Asp>Ala and 258 Phe > Ile detected in SA_190006 and SA_70002 (CC8) showed a 7.3-fold increase in RNAIII expression levels and a 5.8-fold increase in agrA expression levels (p < 0.05). In contrast, SA_180015 (CC8, ST8) and SA_80004 (CC5) both exhibited a deletion in agrC exhibited 93.5- and 148-fold decrease in RNAIII expression levels, and 9.2- and 15.1-fold decrease in agrA expression values for both strains, respectively (p < 0.001). The other CC5 strains also had low expression values for both RNAIII and agrA and, although we did not find any deletions, there were non-synonymous mutations in agrC in SA_103 (96Ser > Leu) and SA_170006 (121I > T) that were associated with 15.9- and 40.3-fold decrease in RNAIII expression and 4.3 and 6.7 in agrA expression, respectively (p < 0.001). However, SA_180009 of CC5 that also exhibited low expression did not have any mutations throughout the entire agr operon. We observed small and nonsignificant changes in the expression of RNAIII by the presence of mutation detected in agrA (136 Lys > Arg) in strains of CC15 (p = 0.478). Nevertheless, this mutation was linked to low expression values for agrA gene with a 2.3-fold decrease (p < 0.001). Finally, for CC30 strains, the mutation detected in agrC (55Gly > Arg) seems to be responsible of the low means values of expression obtained for both RNAIII and agrA (3.2- and 1.80-fold decrease, respectively) (p < 0.05).
Discussion
In the current study, we found that sub-lethal concentrations of oxacillin have an impact on the levels of agr gene expression in a selection of clinical MSSA and MRSA strains belonging to the main clonal complexes that were associated with invasive disease in Spanish hospitals, namely CC5, CC30, CC15, CC45 and CC8 [2,3,4]. These differences between the presence and absence of oxacillin subMICs resulted in increases or decreases in the levels of gene expression that were strain-dependent. In addition, we observed that the strains generally shared different mutations in agr locus according to the CC and that these mutations had an impact on the expression values of agr.
A recent review has shown that sub-lethal concentrations of different antibiotics such as beta-lactams are able to modulate the expression of virulence genes [23]. Our study shows a strain-dependent effect of oxacillin subMICs on the agr operon expression measured as transcriptional changes in the RNAIII and /or agrA mRNA in the clinical strains included in this study. We observed an increase relative to the control condition without oxacillin in 11/14 S. aureus clinical strains being statically significant for 5 of them. Additionally, strains with low expression levels under exposure to oxacillin subMICs were also detected. Because of the agr operon plays an important role in S. aureus virulence the exposure to subMICs of oxacillin could have significant consequences on the pathogenesis of infection. The initiation of transcription of RNAIII strictly relies upon the expression of agrA, in a RNAII-dependent (agrBDCA) manner [8, 9, 11]. Our data support these observations since the transcription of agrA was observed in all strains with RNAIII expression. However, in the presence of oxacillin subMICs, most strains showed the same direction of change in expression values for both genes, except for 4 strains that showed divergent expression values in RNAIII and agrA, although not statistically significant. This unexpected result could be due to other factors affecting on agrA expression. It is important to take into account the wide network of global (sarA, sarU, sarX, codY) and other virulence regulators (mgrA, sigB, saeSR) that have a direct or indirect effect on the agr [35, 36], and specifically on the P2 and P3 promoters to which agrA binds [37], or on agrC-agrA interaction [38]. When we stratified the expression results by CC, we observed higher expression values of agr locus in presence of oxacillin subMICs than in absence of this antimicrobial agent for both CC15 and CC45. However, the relatively small number of strains affords a substantial chance for statistical bias and does not allow performing proper multivariate analyses. On the other hand, methicillin resistance has been associated with a decrease in RNAIII expression under exposure to suboptimal concentrations of different antibiotics [28, 34, 39]. Our results do not seem to support this hypothesis, and only one MRSA strain (SA_170006) showed a reduction in the expression values relative to the control. However, taking into account that from the 4 MRSA (mecA positive) strains, only one is defined as an MRSA according to CLSI breakpoints (SA_170006) [40]. This subtle difference could be impacting on the correlation MSSA/MRSA with RNAIII/agrA expression. Indeed, previous studies [28, 34] suggested that the expression of mecA and either its potential to subtly affect peptidoglycan structure or its interaction with other cell wall–associated proteins prevent the auto-inducing peptide (AIP) from being detected. These results in an unresponsive agr system that could be directly attributed to induced levels of mecA expression because of RNAIII expression was repressed as mecA expression increased.
Although the role of agr system for pathogenesis in animal model is well established [41, 42] and agr-defective mutants have shown an attenuated virulence and agr blocking agents exhibit anti-infective properties in experimental S. aureus infections [43], several studies have reported that agr defective mutants are frequently recovered from patients with bacteremia, where the mutants are associated with persistent infection and poor outcome [9, 19,20,21]. According to previous reports we observed that 13/14 strains harbored both synonymous and non-synonymous mutations throughout the agr operon, especially in agrC and agrA [9, 17, 18]. Moreover, we have observed that while the genotypic analysis revealed similar mutations patterns accordingly to CC, the agr expression was clearly strain-dependent. Some of these mutations were associated to changes in agr gene expression in strains with and without a specific mutation. Deletions found in SA_180015 (CC8) and SA_80004 (CC5) in agrC were related with low expression levels and might be responsible for reduced gene expression in these strains. Both these deletions have previously been detected in strains with dysfunctional agr [9, 17, 18]. Strains of CC5 exhibited the most consistent agr gene expression although this group had the most divergent agr sequences within any CC, all mutations detected in this CC were associated with decreases expression values. Finally, our observations showed that CC30 exhibited low levels of agr gene expression with the presence of a previously identified mutation in agrC [18, 44]. We recognize that the attempt to correlate RNAIII / agrA expression with mutations within the agr operon without complementation experiments is problematic, since in most cases the sample size for any given mutation was one, making it difficult to draw strong correlations between mutations and agr expression. Even so, these results, collectively, indicate that mutations in agr have an impact on agr expression and could be correlated with levels of agr gene expression for strains belonging to the same CC. Nevertheless, although different studies have shown that mutations in agr-defective strains do not persist in natural populations and that inactivating mutations have a relatively short life, other limitation of this study is that we do not have the results from sequencing of the agr operon after exposure to antibiotic, and therefore, we do not know the influence that this selective pressure could exert on mutations in the agr locus. Furthermore, it is possible that mutations detected in the agr system may have an effect not only on the expression on the agr locus itself, but also on other virulence genes regulated by agr. However, although these results could be confirmed by complementation experiments with the wild type gene, we cannot ignore the potential effect that other regulators may also have on agr operon expression independently of the mutations detected [24, 29]. Our findings offer a valuable starting point for further studies that analyze gene expression changes induced by oxacillin subMICs from a whole transcriptomic approach would help to provide a more accurate overview of genes and regulators that compose the virulome framework in S. aureus and that may be affected under this condition.
Conclusions
We conclude that exposure to oxacillin subMICs have a stimulating or depressing effect on expression of the agr system for majority of the strains of S. aureus in our collection. In addition, the mutations we detected in the agr operon follow a common pattern among strains belonging to the same CC and could be responsible of changes in the agr expression. Consequently, the virulence mechanisms involved in the pathogenesis of infection could be affected. However, inpatient data supporting staphylococcal virulence modulation by antibiotics are rather frail, thus relevant clinical trials are necessary to extrapolate this finding to patient outcomes.
Abbreviations
- agr :
-
Accessory gene regulator
- CC:
-
Clonal complexes
- MIC:
-
Minimum inhibitory concentration
- MRSA:
-
methicillin resistant S. aureus
- MSSA:
-
Methicillin-susceptible S. aureus
- OD:
-
Optical density
- SD:
-
Standard deviation
- subMICs:
-
Sub-inhibitory concentrations
- TSB:
-
Tryptic soy broth
References
Fowler VG, Miro JM, Hoen B, Cabell CH, Abrutyn E, Rubinstein E, et al. Staphylococcus aureus endocarditis. JAMA. 2005;293:3012–21.
Bouchiat C, Moreau K, Devillard S, Rasigade JP, Mosnier A, Geissmann T, et al. Staphylococcus aureus infective endocarditis versus bacteremia strains: subtle genetic differences at stake. Infect Genet Evol. 2015;36:524–30.
Fowler VG, Nelson CL, McIntyre LM, Kreiswirth BN, Monk A, Archer GL, et al. Potential associations between hematogenous complications and bacterial genotype in Staphylococcus aureus infection. J Infect Dis. 2007;196:738–47.
Nienaber JJC, Sharma Kuinkel BK, Clarke-Pearson M, Lamlertthon S, Park L, Rude TH, et al. Methicillin-susceptible staphylococcus aureus endocarditis isolates are associated with clonal complex 30 genotype and a distinct repertoire of enterotoxins and adhesins. J Infect Dis. 2011;204:704–13.
Le KY, Otto M. Quorum-sensing regulation in staphylococci-an overview. Front Microbiol. 2015;6:1–8.
Cheung GYC, Joo HS, Chatterjee SS, Otto M. Phenol-soluble modulins - critical determinants of staphylococcal virulence. FEMS Microbiol Rev. 2014;38:698–719.
Wang R, Braughton KR, Kretschmer D, Bach T-HL, Queck SY, Li M, et al. Identification of novel cytolytic peptides as key virulence determinants for community-associated MRSA. Nat Med. 2007;13:1510–4.
Novick RP, Geisinger E. Quorum sensing in staphylococci. Annu Rev Genet. 2008;42:541–64.
Traber KE, Lee E, Benson S, Corrigan R, Cantera M, Shopsin B, et al. Agr function in clinical Staphylococcus aureus isolates. Microbiology. 2008;154:2265–74.
Painter KL, Krishna A, Wigneshweraraj S, Edwards AM. What role does the quorum-sensing accessory gene regulator system play during Staphylococcus aureus bacteremia? Trends Microbiol. 2014;22:676–85.
Novick RP, Projan SJ, Kornblum J, Ross HF, Ji G, Kreiswirth B, et al. The agr P2 operon: an autocatalytic sensory transduction system in Staphylococcus aureus. Mol Gen Genet. 1995;248:446–58.
Queck SY, Jameson-Lee M, Villaruz AE, Bach THL, Khan BA, Sturdevant DE, et al. RNAIII-independent target gene control by the agr quorum-sensing system: insight into the evolution of virulence regulation in Staphylococcus aureus. Mol Cell. 2008;32:150–8.
Novick RP, Ross HF, Projan SJ, Kornblum J, Kreiswirth B, Moghazeh S. Synthesis of staphylococcal virulence factors is controlled by a regulatory RNA molecule. EMBO J. 1993;12:3967–75.
Janzon L, Lfdahl S, Arvidson S. Identification and nucleotide sequence of the delta-lysin gene, hld, adjacent to the accessory gene regulator (agr) of Staphylococcus aureus. Mol Gen Genet. 1989;219:480–5.
Cheung GYC, Kretschmer D, Duong AC, Yeh AJ, Ho TV, Chen Y, et al. Production of an attenuated phenol-soluble Modulin variant unique to the MRSA clonal complex 30 increases severity of bloodstream infection. PLoS Pathog. 2014;10:e1004298.
Traber K, Novick R. A slipped-mispairing mutation in AgrA of laboratory strains and clinical isolates results in delayed activation of agr and failure to translate δ- and α-haemolysins. Mol Microbiol. 2006;59:1519–30.
Shopsin B, Eaton C, Wasserman GA, Mathema B, RXP A, Agolory S, et al. Mutations in agr do not persist in natural populations of methicillin-resistant Staphylococcus aureus. J Infect Dis. 2010;202:1593–9.
Smyth DS, Kafer JM, Wasserman GA, Velickovic L, Mathema B, Holzman RS, et al. Nasal carriage as a source of agr-defective staphylococcus aureus bacteremia. J Infect Dis. 2012;206:1168–77.
Seidl K, Chen L, Bayer AS, Hady WA, Kreiswirth BN, Xiong YQ. Relationship of agr expression and function with virulence and vancomycin treatment outcomes in experimental endocarditis due to methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. 2011;55:5631–9.
Fowler VG Jr, Sakoulas G, McIntyre LM, Meka VG, Arbeit RD, Cabell CH, et al. Persistent bacteremia due to methicillin-resistant Staphylococcus aureus infection is associated with agr dysfunction and low-level in vitro resistance to thrombin-induced platelet Microbicidal protein. J Infect Dis. 2004;190:1140–9.
Schweizer ML, Furuno JP, Sakoulas G, Johnson JK, Harris AD, Shardell MD, et al. Increased mortality with accessory gene regulator (agr) dysfunction in Staphylococcus aureus among Bacteremic patients. Antimicrob Agents Chemother. 2011;55:1082–7.
Cars O. Pharmacokinetics of antibiotics in tissues and tissue fluids: a review. Scand J Infect Dis Suppl. 1990;74:23–33.
Hodille E, Rose W, Diep BA, Goutelle S, Lina G, Dumitrescu O. The role of antibiotics in modulating virulence in Staphylococcus aureus. Clin Microbiol Rev. 2017;30:887–917.
Otto MP, Martin E, Badiou C, Lebrun S, Bes M, Vandenesch F, et al. Effects of subinhibitory concentrations of antibiotics on virulence factor expression by community-acquired methicillin-resistant Staphylococcus aureus. J Antimicrob Chemother. 2013;68:1524–32.
Dumitrescu O, Choudhury P, Boisset S, Badiou C, Bes M, Benito Y, et al. β-lactams interfering with PBP1 induce panton-valentine leukocidin expression by triggering sarA and rot global regulators of Staphylococcus aureus. Antimicrob Agents Chemother. 2011;55(7):3261–71.
Stevens DL, Ma Y, Salmi DB, McIndoo E, Wallace RJ, Bryant AE. Impact of antibiotics on expression of virulence-associated exotoxin genes in methicillin-sensitive and methicillin-resistant Staphylococcus aureus. J Infect Dis. 2007;195:202–11.
Joo HS, Chan JL, Cheung GYC, Otto M. Subinhibitory concentrations of protein synthesis-inhibiting antibiotics promote increased expression of the agr virulence regulator and production of phenol-soluble modulin cytolysins in community-associated methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. 2010;54:4942–4.
Rudkin JK, Laabei M, Edwards AM, Joo HS, Otto M, Lennon KL, et al. Oxacillin alters the toxin expression profile of community-associated methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. 2014;58:1100–7.
Subrt N, Mesak LR, Davies J. Modulation of virulence gene expression by cell wall active antibiotics in Staphylococcus aureus. J Antimicrob Chemother. 2011;66:979–84.
Rasigade J, Moulay A, Lhoste Y, Tristan A, Bes M, Vandenesch F, et al. Impact of sub-inhibitory antibiotics on fibronectin-mediated host cell adhesion and invasion by Staphylococcus aureus. BMC Microbiol. 2011;11:263–71.
Fernández-Hidalgo N, Ribera A, Larrosa MN, Viedma E, Origüen J, de Alarcón A, et al. Impact of Staphylococcus aureus phenotype and genotype on the clinical characteristics and outcome of infective endocarditis. A multicenter, longitudinal, prospective, observational study. Clin Microbiol Infect. 2017. https://doi.org/10.1016/j.cmi.2017.12.002.
San-Juan R, Viedma E, Chaves F, Lalueza A, Fortún J, Loza E, et al. High MICs for vancomycin and Daptomycin and complicated catheter-related bloodstream infections with methicillin-sensitive Staphylococcus aureus. Emerg Infect Dis. 2016;22:1057–66.
Abdelhady W, Chen L, Bayer AS, Seidl K, Yeaman MR, Kreiswirth BN, et al. Early agr activation correlates with vancomycin treatment failure in multi-clonotype MRSA endovascular infections. J Antimicrob Chemother. 2014;70:1443–52.
Rudkin JK, Edwards AM, Bowden MG, Brown EL, Pozzi C, Waters EM, et al. Methicillin resistance reduces the virulence of healthcare-associated methicillin-resistant staphylococcus aureus by interfering with the agr quorum sensing system. J Infect Dis. 2012;205:798–806.
Bronesky D, Wu Z, Marzi S, Walter P, Geissmann T, Moreau K, et al. Staphylococcus aureus RNAIII and its regulon link quorum sensing, stress responses, metabolic adaptation, and regulation of virulence gene expression. Annu Rev Microbiol. 2016;70:299–316.
Priest NK, Rudkin JK, Feil EJ, van den Elsen JMH, Cheung A, Peacock SJ, et al. From genotype to phenotype: can systems biology be used to predict Staphylococcus aureus virulence? Nat Rev Microbiol. 2012;10:791–7.
Rajasree K, Fasim A, Gopal B. Conformational features of the Staphylococcus aureus AgrA-promoter interactions rationalize quorum-sensing triggered gene expression. Biochem Biophys Rep. 2016;6:124–34.
Srivastava SK, Rajasree K, Fasim A, Arakere G, Gopal B. Influence of the AgrC-AgrA complex on the response time of Staphylococcus aureus quorum sensing. J Bacteriol. 2014;196:2876–88.
Waters EM, Rudkin JK, Coughlan S, Clair GC, Adkins JN, Gore S, et al. Redeploying β-lactam antibiotics as a novel Antivirulence strategy for the treatment of methicillin-resistant Staphylococcus aureus infections. J Infect Dis. 2017;215:80-7
CLSI. Performance standards for antimicrobial susceptibility testing. In: CLSI supplement M100. 27th ed. Wayne: Clinical and Laboratory Standards Institute; 2017.
Abdelnour A, Arvidson S, Bremell T, Rydén C, Tarkowski A. The accessory gene regulator (agr) controls Staphylococcus aureus virulence in a murine arthritis model. Infect Immun. 1993;61:3879–85.
Novick RP. Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol Microbiol. 2003;48:1429–49.
Wright JS, Jin R, Novick RP. Transient interference with staphylococcal quorum sensing blocks abscess formation. Proc Natl Acad Sci U S A. 2005;102:1691–6.
Sharma-Kuinkel BK, Mongodin EF, Myers JR, Vore KL, Canfield GS, Fraser CM, et al. Potential influence of Staphylococcus aureus clonal complex CC30 genotype and transcriptome on Hematogeneous infections. Open Forum Infect Dis. 2015;2(3):ofv093. https://doi.org/10.1093/ofid/ofv093.
Funding
This work was supported by the Ministerio de Economía y Competitividad of Spain, Instituto de Salud Carlos III the and co-financed by the European Regional Development Fund (ERDF) project “A way to achieve Europe” through the Spanish Network for Research in Infectious Diseases (REIPI) (RD16/0016/0002 and RD16/0016/0003) and the grants PI12/01205; PI15/02013; PI12/0179 and PI15/02125 (Health Research Fund; Department of Health; Spain).
E. Viedma was supported by Rio Hortega Program (Instituto de Salud Carlos III – CM15/00183).
The funding sources had no role in the study design, data collection and analysis, neither preparation of the manuscript.
Availability of data and materials
The datasets generated and/or analysed during the current study (nucleotide sequences) are available in the GenBank (NCBI): https://www.ncbi.nlm.nih.gov/genbank/wgs/. The accession numbers are: MKYX00000000, MKYY00000000, MKYZ00000000, MKZA00000000, MKZB00000000, MKZC00000000, MKZD00000000 MKZE00000000, MKZF00000000, MKZG00000000, MKZH00000000, MKZI00000000, MKZJ00000000 and MKZK00000000.
Author information
Authors and Affiliations
Contributions
EV and FC conceived and designed the experiments. EV, NL, NFH, JG, JV collected the isolates. Funding was obtained by FC and BA. Experiments were performed by EV and IMG. The data were analyzed by EV and DPM. EV prepared the manuscript draft. All authors agreed to be accountable for all aspects of the work. FC and DPM contributed in giving final approval of the version to be published. All authors reviewed and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
Figure S1. Growth curves for S. aureus strains included in the study in the presence of subMIC of oxacillin. Growth was monitored by determining optical density at 600 nm (O.D600). (JPEG 5949 kb)
Additional file 2:
Table S1. Expression levels changes in RNAIII and agrA locus under the exposure to oxacillin subMICs in the stationary growth phase. aOXA: Oxacillin. bPOS: fold change positive in the expression level under exposure to oxacillin subMIC. NEG: fold change negative in the expression level under exposure to oxacillin subMICs. cBold numbers indicate statistical significant results (P < 0.05). (DOCX 19 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

{kind=link}
Cite this article
Viedma, E., Pérez-Montarelo, D., Villa, J. et al. Sub-inhibitory concentrations of oxacillin modify the expression of agr locus in Staphylococcus aureus clinical strains belonging to different clonal complexes. BMC Infect Dis 18, 177 (2018). https://doi.org/10.1186/s12879-018-3088-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12879-018-3088-7