
Abstract
Background
S.aureus is a predominant pathogen that causes infection in critically ill patients, but little information exists regarding the characterization of S. aureus from different sources in burn patients in southeastern China.
Methods
We enrolled 125 patients with S. aureus infection in burns center between Jan 2014 and Dec 2015. S. aureus isolates were characterized by antimicrobial susceptibility test, toxin gene detection, and molecular typing with multilocus sequence type, staphylococcal protein A (spa) type, and staphylococcal cassette chromosome mec (SCCmec) type.
Results
Sixty-eight MRSA were isolated from SSTI and 31 from non-SSTI patients, respectively. Overall, the drug-resistant ability of S. aureus isolated from SSTI was higher than that from non-SSTI groups. SCCmecIII-CC239-t030 was the most common clone (38 from SSTIs, and 8 from non-SSTIs). Seg was the most common enterotoxin gene (21 from SSTIs and 33 from non-SSTIs). Isolates from SSTIs was more likely to carry seb (P = 0.04), while those from non-SSTIs tended to carry sea and seg (P = 0.002 and 0.01, respectively). Although isolates carried four hemolysin genes, there was no significant difference between them (P > 0.05).
Conclusion
SCCmecIII-CC239-t030 was the most common clone in Jiangxi burns center, China. The molecular characterization of S. aureus was quite different between SSTI and non-SSTI groups.
Similar content being viewed by others


Background
Millions of people have burns every year in the world. Only in China, approximately 5000–10,000 peoples sustain burn injuries per 1 million, and 10% of them require medical intervention. Jiangxi province locates in southeastern region in China, and having numerous fireworks factories. Consequently, there are many thermal burns patients in local burn units, and millions of dollars having been consumed every year.
Skin is the first barrier of defense against microbial invasion, and it becomes more prone to infection once it gets burns. Many pathogens are responsible for burn wound infections, including Staphylococcus spp., Enterococcus spp., Pseudomonas spp., Acinetobacter spp., and fungi [1]. However, Staphylococcus aureus (S.aureus) remains a leading cause of infections in burn centers [2]. The emergence and transmission of methicillin-resistant S. aureus (MRSA) in burn centers results in some poor outcomes such as prolonged hospitalization, bacteremia or sepsis, even death, which require further prevention and treatment efforts [3].
The main task for successful clinical treatment of microorganism infection depends on the knowledge of the characteristics of these infecting strains and their antibiotic susceptibility profiles. As a result, identification of the antibiotic susceptibility patterns of different strains in the burns center provides more appropriate treatments and decreased expenses. In addition to antibiotic resistance, another important factor that leads to treatment failure in burns patients is the production of different virulence factors. For example, these Panton-Valentine Leukocidin (PVL)-producing strains of S. aureus are associated with increased lethality compared to the PVL-negative strains [4]. Skin and soft tissue infections (SSTIs) still have been taken as the primary source in burns patients [5, 6] and antibacterial ointment is usually taken as the mainstay of treatment, different from non-SSTIs patients who treated with injectable antibiotics in China. It is necessary to know different characteristics between SSTI and non-SSTI groups for empirical therapy at the early infectious stage.
Molecular typing of S. aureus is of significant value as a tool to control infection [7]. At present, there are several molecular typing methods, including multilocus sequence typing (MLST), staphylococcal protein A (spa) typing, and staphylococcal cassette chromosome mec (SCCmec) typing. In general, a combination of typing methods could provide more detailed genetic linkage among these strains.
A high rate of isolated S. aureus corresponds to a high infectious level in burn centers [1,2,3]. However, there are few studies on the characterization of high isolated S. aureus from patients with burns in Jiangxi province, China. Many factors have action on effective treatment and control of S. aureus or MRSA infections. As a result, it is extremely important to know the different pattern of antimicrobial resistance and the prevalent clone of S. aureus or MRSA for local optimal clinical control.
Methods
Bacterial isolates
This study included all clinical isolates submitted for culture and sensitivity test at the burn center of the First Affiliated Hospital of Nanchang University in China, from January 1, 2014s to December 31, 2015s. S. aureus isolates were identified by using the VITEK-2 compact (BioMérieux, France) GP colorimetric identification card. Resistance to oxacillin or cefoxitin was confirmed by VITEK-2 compact AST-GP67 (BioMérieux, France), with resistance defined as an oxacillin minimum inhibitory concentration (MIC) ≥ 4 μg/mL and cefoxitin MIC ≥8 μg/mL [8]. In the case of multiple S. aureus isolates from the same patient, the first detected resistant specimen was included. In total, 125 S. aureus isolates were characterized, including 75 from SSTIs and 50 from non-SSTIs. The isolated strains stored at − 80 °C were thawed and sub-cultured on BHI medium prior to DNA extraction.
Antimicrobial susceptibility test (AST)
In addition to oxacillin and cefoxitin, other antimicrobials listed for routine reporting of S. aureus were included for AST by using VITEK-2 compact AST-GP67 (BioMérieux, France) test card for Gram-positive susceptibility. Antibiotics used for susceptibility test included benzylpenicillin, gentamicin, ciprofloxacin, levofloxacin, moxifloxacin, erythromycin, clindamycin, quinupristin/dalfopristin, linezolid, vancomycin, tetracycline, tigecycline, nitrofurantoin, rifampicin, and trimethoprim/sulfamethoxazole. The MIC interpretive standard or breakpoint values were set following the guidelines of Clinical and Laboratory Standard Institute M100-S23. S. aureus ATCC25923 was used as a quality control for MIC detection.
Detection of toxin genes
Genomic DNA was extracted using a DNA extraction kit (Sangon Biotech, Shanghai) with lysostaphin, according to the manufacturer’s instructions. Several clinically significant toxin genes were detected by PCR [9, 10], including lukS/F-PV (encoding Panton-Valentine leukocidin), tst (encoding toxic shock syndrome toxin 1), sea-see and seg-sej (encoding staphylococcal enterotoxins SEA-SEE and SEG-SEJ), and HLa-HLg (encoding staphylococcal hemolysin).
Detection of molecular types
All S. aureus isolates were investigated by multilocus sequence typing (MLST), and the products of seven housekeeping gene fragments were sequenced (Sangon Biotech, Shanghai) and compared with allele profiles from S. aureus database (www.mlst.net/). Spa and SCCmec typing of MRSA were performed as previously described [2] .
Statistical analysis
The chi-square or Fisher’s exact test was used as appropriate using the software package SAS 8.2 (SAS Institute Inc., Cary, NC, USA). A two-sided P value of < 0.05 was considered statistically significant.
Results
Clinical data
The median age of burns patients was 26 years and 7 months (range: 3 months - 79 years), as the sex distribution of male/female was 69/56 (55.2%/44.8%). All enrolled patients were from the burns center. The infectious source for burns patients included SSTI (75/125, 60.0%), respiratory tract (21/125, 16.8%), blood (14/125, 11.2%), and others (15/125, 12.0%).
Antimicrobial susceptibility test
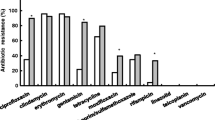
Ninety-nine isolates (68 isolated from SSTIs and 31 from non-SSTIs) were confirmed as MRSA and 26 isolates (7 isolated from SSTIs and 19 from non-SSTIs) confirmed as methicillin-susceptible S.aureus (MSSA), respectively. All 125 isolates were susceptible to vancomycin, linezolid, daptomycin, and quinupristin/dalfopristin. 15 isolates (3 from SSTIs and 12 from non-SSTIs) were susceptible to penicillin. The antimicrobial susceptibilities of S. aureus isolated from SSTIs were almost equal to those from non-SSTIs. These findings were summarized in Table 1.
Virulence factors
As shown in Table 2, seg (54/125, 43.2%) was the most frequently detected enterotoxin gene, of which 21 isolated from SSTIs and 33 isolates were from non-SSTIs. 19 isolates showed presence of lukS/F-PV, including 15 SSTIs and 4 non-SSTIs. No seh-positive isolates were detected. Staphylococcal HLa and HLg were the most frequently detected hemolysin genes, occurring in 118 isolates. S. aureus isolates from SSTIs were more likely to carry sea and seg (P = 0.002 and 0.01, respectively), whereas non-SSTI patients were more likely to carry seb (P = 0.04).
Molecular epidemiological characteristics
Among 125 S. aureus isolates, 21 sequence types (STs) and 27 spa types were acquired (Table 3). We detected 4 SCCmec type I, 12 SCCmec type II, 61 SCCmec type III, 20 SCCmec IV, and 2 SCCmec type V. As shown in Fig. 1, MSSA isolates expressed greater diversity in molecular characteristics than MRSA; 43 clones were present in these isolates, including 10 shared clones.

Distribution of major clones among 125 S.aureus isolates
Twenty sequence types (STs) from 13 clonal complexes (CCs) were identified among all 125 S. aureus isolates. The most common ST was ST239 (63/125, 50.4%), followed by ST5 (14/125, 11.2%), ST59 (7/125, 5.6%), and ST30 (7/125, 5.6%). ST239 was detected in 44 isolates from SSTIs and 19 isolates from non-SSTIs, and ST59 was found in 6 isolates from SSTIs and 1 isolate from non-SSTI. The distribution of S. aureus in ST239 and ST5 was significantly different between SSTIs and non-SSTIs (P = 0.057 or 0.006); however, there was no difference in the distribution of ST59 and ST30 between isolates from SSTI and non-SSTI groups (P = 0.174, 0.111). ST239 was the most common ST among lukS/F-PV-positive isolates, including 11 isolates (10 MRSA and 1 MSSA) from SSTIs and 8 isolates (6 MRSA and 2 MSSA) from non-SSTIs. The other STs in lukS/F-PV-positive isolates were ST88 (2 MRSA), ST25 (1 MRSA), and ST398 (1 MRSA).
The different 26 spa type was identified in this study. t30 (49/125, 39.2%) was the most common spa type, followed by t037 (16/125, 12.8%) and t062 (14/125, 11.2%). t30 was detected in 39 isolates from SSTIs and in 10 isolates from non-SSTIs, and t037 was detected in 9 isolates from SSTIs and 7 isolates from non-SSTIs. The former displayed a significant difference between isolates from SSTIs or non-SSTIs (P = 0.014), while the latter did not (P = 0.774).
Discussion
As a clinically important pathogen, S. aureus can cause many kinds of infections, such as SSTI, sepsis, pneumonia, and abscesses. As shown in the results section, there were 75 SSTIs, 14 blood or other bodily fluids, and 21 respiratory infection samples from S. aureus infected patients. Additionally, 68 isolates (15 community-acquired MRSA (CA-MRSA) and 53 hospital-acquired MRSA (HA-MRSA)) were obtained from SSTIs and 31 isolates (6 CA-MRSA and 25 HA-MRSA) from non-SSTIs. There was no significant difference between SSTI and non-SSTI groups in the occurrence of MRSA (P = 0.179). However, as shown in Table 1, S.aureus had higher resistance to erythromycin in SSTIs than that from non-SSTIs (P = 0.02), probably due to a high level of MRSA infection, which commonly integrates some important resistance genes [11]. Since its emergence in 1995 in Brazil, prevalence of ST239 has led to a high occurrence of hospital-acquired infection globally [12], as witnessed by S. aureus clone outbreaks in China, Africa, Australia, and America [13,14,15]. In this study, ST239 was found to be the most prevalent clone in the burn center (50.4%, 63/125), which is consistent to reports that ST239 is the most common clone in mainland China [16]. ST239 is usually associated with SSTIs disease [17], as a result, it was in expectation that patients with SSTIs had a higher proportion of infection caused by the ST239 clone than patients with non-SSTIs (P = 0.048). ST239 is mainly associated with staphylococcal cassette chromosome mec type III (ST239-SCCmecIII) [18]. As shown in Table 3, this study demonstrated evidence that more than half of the isolates from SSTIs were ST239-SCCmecIII-t030 or ST239-SCCmecIII-t037, and this number was higher than those isolates from non-SSTIs. ST239-SCCmecIII-t037 is closely related with ST239-SCCmecIII-t030, as the former is considered an ancestral ST239 spa type and has been gradually replaced by ST239-SCCmecIII-t030 in many places [19, 20]. ST5, found characterized in a total of 12 MRSA and 2 MSSA isolates, was the second most common ST in our study. ST5 has also been found to be among the most common genotype isolated from children with cystic fibrosis (CF) or nasal carriage [21, 22].
Comprehensively considered high antibiotic resistance and high virulence, some isolated S. aureus were a dangerous pathogen in this study. The harmfulness of S. aureus is partially determined by a series of virulence factors, which vary significantly according to different strains. As a result, to acquire valuable epidemiologic control in an endemic area, it is necessary to know the distribution of some important virulence factors. Staphylococcal enterotoxins (SEs) include a series of virulent factors associated with a severe disease, which usually causes food poisoning. SEs may suppress the motility of the human polymorphonuclear neutrophils through the inhibition of exoprotein expression, and allow S. aureus to invade and damage tissues. Although previous studies have confirmed that sea and seb are the most abundant toxin genes in clinical S. aureus isolates from patients in China [23], seg was the predominant SE gene detected in current study. There was significant difference observed between isolates from SSTIs and non-SSTIs (P = 0.009), which might represent regional characteristics. The study confirms that isolates from SSTIs predominantly carry seb (P = 0.042), which has been proved to be a hallmark of CA-MRSA [24]. There are four hemolysin genes in S. aureus, with the roles of which usually lead to the release of bacteriolytic enzyme that damages the surrounding cells by inhibiting blood cell function. HLa and HLb are the main toxins inducing pathological injury. Consistent with the report in Wenzhou, China [25], HLa, HLb, HLd, and HLg were all been detected in this study. These findings indicated that a high level of HLa, HLb, HLd, and HLg genes could be considered characteristic of MRSA strains in this region. There was no difference between isolates from SSTIs and non-SSTIs in these genes (P = 0.567–0.871, respectively). LukS/F-PV encodes Panton-Valentine leukocidin, an important virulent factor which usually damages neutrophils and is associated with SSTIs [26]. There is no connection between PVL-positive isolates among S. aureus isolated from SSTIs and those from non-SSTIs (P = 0.112), mostly because these strains originate from SSTIs.
Conclusion
Our study indicated that the infectious rate of MRSA was high in burns center and SSTIs had higher resistant levels than non-SSTIs. Additionally, SSTIs have different virulent factors profiles compared with non-SSTIs. The most prevalent clone was ST239-SCCmecIII- t030 in the burn center.
Abbreviations
- CLSI:
-
Clinical and Laboratory Standards Institute
- MRSA:
-
Methicillin-resistant S.aureus
- MSSA:
-
Methicillin-susceptibility S.aureus
- SCCmec:
-
Staphylococcal cassette chromosome mec
- SSTI:
-
Skin and soft tissue infection
References
Norbury W, Herndon DN, Tanksley J, Jeschke MG, Finnerty CC. Infection in burns. Surg Infect. 2016;17(2):250–5.
Chen X, Yang HH, Huangfu YC, Wang WK, Liu Y, Ni YX, Han LZ. Molecular epidemiologisc analysis of Staphylococcus aureus isolated from four burn centers. Burns. 2012;38(5):738–42.
Issler-Fisher AC, Mckew G, Fisher OM, Harish V, Gottlieb T, Maitz PK. Risk factors for, and the effect of MRSA colonization on the clinical outcomes of severely burnt patients. Burns. 2015;41(6):1212–20.
Holmes A, Ganner M, McGuane S, Pitt TL, Cookson BD, Kearns AM. Staphylococcus aureus isolates carrying Panton-Valentine leucocidin genes in. England and Wales: frequency, characterization, and association with clinical disease. J Clin Microbiol. 2005;43(5):2384–90.
Chen K, Huang Y, Song Q, Song Q, Chen X, Zeng L. Drug-resistance dynamics of Staphylococcus Aureus between 2008 and 2014 at a tertiary teaching hospital, Jiangxi Province, China. BMC Infect Dis. 2017;17(1):97.
Cen H, Wu Z, Wang F, Han C. Pathogen distribution and drug resistance in a burn ward: a three-year retrospective analysis of a single center in China. Int J Clin Exp Med. 2015;8(10):19188–99.
Xie X, Bao Y, Ouyang N, Dai X, Pan K, Chen B, et al. Molecular epidemiology and characteristic of virulence gene of community-acquired and hospital-acquired methicillin-resistant Staphylococcus aureus isolates in sun Yat-sen memorial hospital, Guangzhou, southern China. BMC Infect Dis. 2016;16(339)
CLSI. Performance standards for antimicrobial susceptibility test: 24th informational supplement M100-S24. Clinical and laboratory standard institute. PA, USA: Wayne; 2014.
Gu FF, Han LZ, Chen X, Wang YC, Shen H, Wang JQ, et al. Molecular characterization of Staphylococcus Aureus from surgical site infections in orthopedic patients in an orthopedic trauma clinical medical center in shanghai. Surg Infect(Larchmt). 2015;16(1):97–104.
Argudín MA, Tenhagen BA, Fetsch A, Sachsenröder J, Käsbohrer A, Schroeter A, et al. Virulence and resistance determinants of german Staphylococcus Aureus ST398 isolates from nonhuman sources. Appl Environ Microbiol. 2011;77(9):3052–60.
Teodoro CR, Mattos CS, Cavalcante FS, Pereira EM, dos Santos KR. Characterization of MLS(b) resistance among Staphylococcus aureus and staphylococcus epidemidis isolates carrying different SCCmec types. Microbiol Immunol. 2012;56(9):647–50.
Teixeira LA, Resende CA, Ormonde LR, Rosenbaum R, Figueiredo AM, de Lencastre H, et al. Geographic spread of epidemic multiresistant Staphylococcus aureus clone in Brazil. J Clin Microbiol. 1995;33(9):2400–4.
Abdulgader SM, Shittu AO, Nicol MP, Kaba M. Molecular epidemiology of methicillin-resistant Staphylococcus aureus in Africa: a systematic review. Front Microbiol. 2015;6(348)
Baines SL, Holt KE, Schultz MB, Seemann T, Howden BO, Jensen SO, et al. Convergent adaptation in the dominant global hospital clone ST239 of methicillin-resistant Staphylococcus aureus. MBio, 2015; 6(2): e00080.
Chen X, Sun K, Dong D, Luo Q, Peng Y, Chen F. Antimicrobial resistance and molecular characteristics of nasal Staphylococcal aurues isolates from newly admitted inpatients. Ann Lab Med. 2016;36(3):250–4.
Ning X, Sun M, Qiao Y, Dong F, Song W, Yao K, et al. Characterization of pediatric hospital-associated infection caused by methicillin-resistant Staphylococcus aureus in mainland China. Infect Dis (lond). 2015;47(6):410–7.
Changchien CH, Chen SW, Chen YY, Chu C. Antibiotic susceptibility and genomic variations in Staphylococcus aureus associated with skin and soft tissue infection (SSTI) disease groups. BMC Infect Dis. 2016;16(276)
Liu Y, Wang N, Du N, Shen H, Niu J, Ye H, et al. Molecular evidence for spread of two major methicillin-resistant Staphylococcal aureus clones with a unique geographic distribution in Chinese hospital. Antimicrob Agents Chemother. 2009;53(2):512–8.
Chen H, Liu Y, Jiang X, Chen M, Wang H. Rapid change of methicillin-resistant Staphylococcus aurues clones in a Chinese tertiary care hospital over a 15-year period. Antimicrob Agents Chemother. 2010;54(5):1842–7.
Goudarzi M, Bahramian M, Satarzadeh Tabrizi M, Udo EE, Fiqueiredo AM, et al. Genetic diversity of methicillin resistant Staphylococcus Aureus strains isolated from burn patients in Iran: ST239-SCCmec/t037 emerges as the major clone. Micro Pathog. 2017;105:1–7.
Vieira MA, Minamisava R, Pessoa-Júnior V, Lamaro-Cardoso J, Ternes YM, Andre MC, et al. Methicillin-resistant Staphylococcus aureus nasal carriage in neonates and children attending a pediatric clinics in Brazil. Braz J Infect Dis. 2014;18(1):42–7.
Liu Y, Zhang J, Zhong D, Ji L, Yang J, Philips J, et al. Characterization of Staphylococcus aureus isolates from pediatric patients with cystic fibrosis. World J Microbiol Biotechnol, 2016; 32(10): 162.
He W, Chen H, Zhao C, Zhang F, Li H, Wang Q, et al. Population structure and characterization of Staphylococcus aureus from bacteraemia at multiple hospitals in China: association between antimicrobial resistance, toxin genes and genotypes. Int J Antimicrob Agents. 2013;42(3):211–9.
Aguilar JL, Varshney AK, Wang X, Stanford L, Scharff M, Fries BC. Detection and measurement of staphylococcal enterotoxin-like K(SEI-K) secretion by Staphylococcus aureus clinical isolates. J Clin Microbiol. 2014;52(7):2536–43.
Yu F, Liu Y, Lv J, Lu C, Ding Y, Li D, et al. Antimicrobial susceptibility, virulence determinant carriage and molecular characteristics of Staphylococcus aureus isolates associated with skin and soft tissue infections. Braz J Infect Dis. 2015;9(6):614–22.
Shallcross LJ, Fragaszy E, Johnson AM, Hayward AC. The role of the Panton-valentine leucocidin toxin in staphylococcal disease: a systematic review and meta-analysis. Lancet Infect Dis. 2013;13(1):43–54.
Acknowledgements
We thank all study patients and staff in the regional burn center.
Funding
This work was partially supported by the National Natural Science Foundation of China (81660555).
Availability of data and materials
The datasets were generated during and/or done analysis during the current study are included in this published article.
Author information
Authors and Affiliations
Contributions
WZ conceived and designed the experiments. KSC, SRL, PQL, QYS, DL and TL performed the experiments. LBZ did the data analysis. KSC wrote the initial paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was approved by the Ethics Committee of the First Affiliated Hospital of Nanchang University (Approved number: 2,015,018). We confirmed that all informed consent were obtained from all participants, and all information was anonymized, and no individual information can be identified.
Consent for publication
No applicable.
Competing interests
All contributing authors declare no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Chen, K., Lin, S., Li, P. et al. Characterization of Staphylococcus aureus isolated from patients with burns in a regional burn center, Southeastern China. BMC Infect Dis 18, 51 (2018). https://doi.org/10.1186/s12879-018-2955-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12879-018-2955-6