
Abstract
Background
The characterization and research around the gut microbiome in older people emphasize microbial populations change considerably by losing the diversity of species. Then, this review aims to determine if there is any effect on the gut microbiota of adults older than 65 that starts an exercise intervention or improves physical activity level. Also, this review describes the changes in composition, diversity, and function of the gut microbiota of older subjects that had improved their physical activity level.
Methods
The type of studies included in this review were studies describing human gut microbiota responses to any exercise stimulus; cross-sectional studies focused on comparing gut microbiota in older adults with different physical activity levels—from athletes to inactive individuals; studies containing older people (women and men), and studies written in English. This review's primary outcomes of interest were gut microbiota abundance and diversity.
Results
Twelve cross-sectional studies and three randomized controlled trials were examined. Independently of the type of study, diversity metrics from Alpha and Beta diversity remained without changes in almost all the studies. Likewise, cross-sectional studies do not reflect significant changes in gut microbiota diversity; no significant differences were detected among diverse groups in the relative abundances of the major phyla or alpha diversity measures. Otherwise, relative abundance analysis showed a significant change in older adults who conducted an exercise program for five weeks or more at the genus level.
Conclusions
Here, we did not identify significant shifts in diversity metrics; only one study reported a significant difference in Alpha diversity from overweight people with higher physical activity levels. The abundance of some bacteria is higher in aged people, after an exercise program, or in comparison with control groups, especially at the genus and species levels. There needs to be more information related to function and metabolic pathways that can be crucial to understand the effect of exercise and physical activity in older adults.
Trial registration
PROSPERO ID: CRD42022331551.
Similar content being viewed by others

Background
According to the World Health Organization (WHO), the population aged 60 years will double by 2050, and it is projected that people older than 80 will triplicate. These increasing numbers could reach over two billion older adults in the following decades, becoming a significant health issue worldwide to ensure wellness [1].
The aging process is characterized by a progressive loss of physiological integrity, leading to impaired function and increased vulnerability to death [2]. This natural condition affects most living organisms because of the decline of functionality as aging progress, conducted by cellular damage [3]. This deterioration has been widely studied in humans because it is the primary risk factor for significant pathologies [4]. Lopez-Otín and colleagues 2013 enumerated nine candidate hallmarks that represent common aging denominators and contribute to determining the aging phenotype [2]. However, recent studies of aging have planned new hallmarks compromising inflammation and microbiome disturbance, among others [5]. This new perspective could better explain health outcomes related to aging diseases and therapeutic studies to achieve a high-quality lifestyle for older people.
Current advances in sequencing technologies and bioinformatics pipelines have identified notable changes in the gut microbiota/microbiome through the lifespan and its substantial effects on human health [6]. The microbiome refers to the combined genetic material of all the microorganisms (bacteria, fungi, protozoa, megafauna, and viruses) living in a particular environment; this term is explicitly used to denote the genetic and functional diversity of the microorganisms community and its relationship with the host [7, 8].
In brief, the diversity and abundance of taxa that make up the gut microbiota (refers to composition) are highly susceptible to change. This is because of external and internal factors that are inherent to the human being, such as birth mode [9], presence or absence of diseases [10], geographical location [11], and diet [12], among others. Previous research shows that the intestinal microbiota in healthy individuals is stable, especially when there is an absence of clinical manipulation (for example, indiscriminate use of antibiotics) and healthy lifestyle habits, such as an adequate diet and moderate to vigorous physical activity [13]. An adequate balance of bacteria in the digestive tract ensures the microbiota works in a symbiotic environment with the host, however, changes in diversity could lead to a reduction in the abundance of beneficial bacteria and an increase in the prevalence of potentially pathogenic microorganisms, also called dysbiosis [14,15,16,17].
Diet is one of the most relevant environmental factors in the investigation of the intestinal microbiome since it modulates the population of microorganisms considerably [18], factors related to diet and nutrition status are key to modulating the composition of microorganisms that inhabit throughout the digestive system [19].
The relationship between the consumption of microbiota-accessible carbohydrates (MACs) and the production of butyrate, as well as the abundance of bacteria that produce this short-chain fatty acid, has been explored in human studies [20]. Significant reductions in the consumption of this macronutrient lead to a drastic decrease in Bifidobacterium spp., Roseburia spp., and Eubacterium rectale. Other microbiota members like Clostridium spp. are important for colon cells since they release butyrate as a final product of fermentation. However, the consumption of various starches and fibers can define the type of bacteria that abounds or impacts the intestine [21]. Also, the breaking of large chains of amino acids results in the generation of metabolites such as hydrogen, methane, carbon dioxide, some SCFAs, and branched-chain amino acids (BCAAs). These metabolites resulting from the fermentation of amino acids fulfill a wide range of biological functions for the host; however, the abundance of some of these compounds may be related to inflammation processes or chronic diseases, since large amounts can be detrimental to the intestinal environment [22].
Although it is not clear the underlying mechanisms that drive changes in the gastrointestinal microbiota under exercise conditions, a few studies involving omics sciences provide possible pathways [23,24,25]. Scheinman et al. identified in a cohort of athletes that the genus Veillonella increased considerably after running a marathon. Subsequent analysis of the V. atypica strain led the authors to conclude that this microorganism promotes an improvement in race time because of its conversion metabolism of exercise-induced lactate into propionate, thus identifying a natural enzymatic process encoded in the microbiome that enhances athletic performance through the Cori cycle [24]. One of the most relevant results is how intestinal colonization of Veillonella increases the Cori cycle by providing an alternative method of lactate processing whereby systemic lactate is converted into SCFAs that re-enter the circulation. SCFAs are absorbed in the sigmoid and rectal region of the colon and enter the circulation through the pelvic plexus, bypassing the liver and draining through the vena cava to reach the systemic circulation directly [24]. Microbiome-derived SCFAs then directly and acutely enhance performance, suggesting that the microbiome might access lactate generated during periods of sustained exercise and convert it into these athletic performance-enhancing SCFAs.
From infancy to old age, the gut microbiome follows some patterns related to rapid change, becoming increasingly unique to individuals as they grow [26]. The characterization and research around the gut microbiome in older adults emphasize microbial populations change considerably by losing the diversity of species [27]; indeed, disturbances and diseases have been linked to these shifts [28, 29].
Recently, three independent cohorts comprising over 9000 individuals aged 18–87 characterized gut microbial patterns associated with age. They performed diversity analysis from multiple samples, paying particular attention to older adults. The results showed amplicon sequence variance (ASV) levels had a unique gut microbiome signature independent of sex or body max index and more related to age [26]. Otherwise, individuals over 80 exhibit continued microbial drift depending on health status. Wilmanski et al., identified microbiome patterns of healthy aging, such as depletion of core genera, primarily Bacteroides [26], and different microbial metabolic outputs in the blood, such as lower LDL cholesterol levels, higher levels of vitamin D and beneficial blood metabolites produced by gut microbes. These results are consistent with recent findings showing that host metabolism is crucial to understand the crosstalk between gut communities and the therapeutic alternatives [29, 30]. Despite the diet (a central shifter of the gut community [31, 32]), physical activity status is now considered a relevant factor in the study of the gut microbiome [33].
Physical activity (PA) is any movement produced by skeletal muscles that requires energy expenditure. The WHO includes leisure time, transport to and from places, and workdays as PA [34, 35]. The amount of activity can be quantified between low and vigorous intensity. Some types include walking, cycling, sports, and recreational activities [35], known to prevent and manage chronic non-transmissible diseases (stroke, diabetes, several cancers), many of which appear with aging [36].
PA and exercise training are well-known modifiable factors in aging, either for preventive medicine or chronic disease management. The protective effect and physiological response to exercise training have been extensively described [37, 38]: enhance the antioxidant response, promote activation of anabolic and mitochondrial biogenesis pathways in skeletal muscle [39], decrease inflammatory profile [40], improve insulin sensitivity, myokine profile and endothelial function [41, 42]. These changes confer multiple health outcomes, such as reducing symptoms of anxiety and depression [43], preventing falls and related injuries [41], improving all-cause mortality, an incident of type 2 Diabetes (T2D), specific cancers, or hypertension, and bone and muscular health. Physical exercise is associated with healthy aging, multisystemic benefits provided to this population are condensed in a multidimensional beneficial system; increased muscle synthesis, improved respiratory function, decreased blood pressure levels, and increased neurogenesis, as well as increased bone density muscle mass and loss of body fat percentage [44,45,46].
Several investigations have repeatedly shown that exposure to regular physical activity confers multiple positive effects on the aging process. The benefits of structured aerobic exercise programs are linked to better learning and cognitive performance on executive function and attentional control in aging [47, 48]. Bouts of physical activity also have a potential therapeutic capacity in conditions related to older adults, such as dementia [49]. Likewise, sufficient results from human and animal trials show the downregulation of pro-inflammatory cytokines and compounds by cardiovascular exercise [47, 50,51,52]. However, the effect of PA and exercise training on the composition and function of the gut microbiota in older people is not clear, considering the relevant role of the gut commensals for health outcomes and the modifications that confer augmenting PA [53,54,55,56,57].
Cross-sectional [54, 58, 59] and longitudinal [60,61,62] studies have sought to establish differences in the composition of the human gut microbiota related to physical activity level (PAL); however, the results are highly variable and sometimes contradictory. Only a few results suggest a significant difference in α and β diversity indicators between subjects with high and low PAL [54, 63]; meanwhile, other results show no change in the composition of gut microbiota related to exercise regimen [60, 64]. Modification of single bacteria taxa has been related to exercise stimulus, especially the increased abundance of Lactobacillus, Bifidobacterium, and Akkermansia [65]. Deeper analysis, specifically metabolome and metagenomic assays, shows significant changes in volatile compounds such as SCFAs [23] and unique members of the microbiome like Veillonella [24].
Studies seeking a link between physical activity and the gut microbiota include diverse age groups, such as older people [66,67,68,69,70] young adults [71, 72], adolescents [73], and mostly middle-aged women and men [54, 60, 61, 64, 74, 75]; likewise, diverse frequency, intensity, and type of exercise interventions can be found in these studies [24, 60, 76]. The growing evidence of the modulator effect of physical activity on the gut microbiota makes it relevant to conduct different systematic reviews where the type of population, type of studies, and type of exercise intervention are described.
Therefore, this systematic review aims to identify with the current and evidence whether starting an exercise program or improving PA level brings any notable change in the gut microbiota of adults older than 65 and whether these modifications are reflected in other physiological systems. This systematic review describes the changes in composition, diversity, and function of the gut microbiota of older adults that have improved their physical activity levels.
Methods
Criteria for considering studies for this review
Types of studies
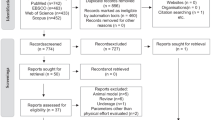
Since the gut microbiota research field in physical activity and exercise is growing, past reviews have showed that randomized control studies are few [53, 77]. For that reason, we consider involving: (a) studies describing human gut microbiota responses to any exercise stimulus (b) cross-sectional studies focused on comparing gut microbiota among older adults with different physical activity levels—from athletes to inactive individuals; (c) studies containing older adults women and men (+ 65 years old); (d) studies written in English. We excluded studies containing probiotic or prebiotic consumption and studies focused on diabetes and cancer. Reviews, comments, letters, interviews, and book chapters were also excluded. PRISMA Flow Diagram (Fig. 1) shows the screening process for this systematic review [78].

Preferred Reporting Items for Systematic Reviews [78]
Types of participants
Populations studied in this review were women and men in older adults, which means over 65 years old. Since it is challenging to reach the elderly with no disease or medical condition, we defined our population as aged functional subjects with no physical limitation or physical disability. Studies involving people aged 65 years and older with only two medical conditions related to older adults or healthy were included.
Types of interventions
The focus of this review is to determine if starting any exercise intervention could significantly change the gut microbiota; for that purpose, we have established the following eligibility criteria for types of intervention a) randomized controlled trials designed to improve any of the muscular strength, endurance, or flexibility components of fitness in the population named before and b) non-randomized controlled trials designed to improve physical activity level through lifestyle interventions, cross-sectional studies will also be included.
Types of outcome measures
The primary outcomes of interest are those related to the diversity and abundance of the gut microbiota. Secondary outcomes will focus on measures or approaches to the function of the gut microbiome. Also, quantification of physical activity level (E.g., median daily step counts) and outcomes related to maximum oxygen consumption and muscular strength will be considered.
The outcomes of interest for this review are:
-
-Gut Microbiota Abundance: one term frequently used in gut microbiota research is absolute abundance, which refers to the "unobservable actual abundance of a taxon in a unit volume of an ecosystem, such as the gut" [79]. It is essential to highlight that absolute and relative abundance are entirely different terms, according to Lin & Peddada. Changes in the absolute abundance of a single taxon can alter the relative abundance of all taxa [80].
These parameters are determined by the data got in the sequencing process; the next-generation sequencing (NGS) of the 16S rRNA helps describe microbial compositions in a niche. After a quality process, the 16S amplicon sequences can be clustered into Operational Taxonomic Units (OTUs) and Sequence Variants (SVs). In brief, observed counts of OTUs or SVs represent observed abundances of taxa in the sample [79,80,81].
-
-Gut Microbiota Diversity: Gut microbiota diversity refers to the number of different species present in a sample, niche, or ecosystem [82]. This review will be focused on stool samples provided by older adults involved in the studies that accomplished the criteria for inclusion. The microbial community in this niche has mainly been characterized in the past years [18]. The bacterial diversity defined by the numerical composition can be calculated with different indexes to determine the changes in the number of species [83]; Alpha diversity refers to the observed richness (number of taxa) and the relative abundances of those taxa (also known as evenness) within a sample. Meanwhile, Beta-diversity is defined as the variability in the microbial community composition among samples [84, 85].
Search methods for identification of studies
The search strategy is summarized in Table 1. The search terms "Elderly AND Gut Microbiota OR Gut Microbiome AND Physical Activity" were used in the bibliographic databases MEDLINE/Ovid, NIH/PubMed, and Academic Search Complete. This electronic search was done between May 14 and June 15, 2022, and other resources were not identified.
Quality assessment
Methodological quality and risk of bias for each study were assessed using the Risk Of Bias In Non-randomized Studies—of Interventions tool (ROBINS-I) [86] and the revised tool to assess the risk of bias in randomized trials (RoB 2) [87, 88].
Once a target trial specificity to the study was designed and confounding domains were listed, the risk of bias was explicitly assessed for the comparisons of interest to this review. The overall risk of biased judgment can be found in Supplementary Table 1 and Supplementary Table 2.
Results
Description of studies
After the electronic screening and evaluation of the pre-selected studies, we finally included fifteen studies in this review (Fig. 1). The type of study is significant cross-sectional, followed by controlled trials (randomized and non-randomized) and follow-up cohorts that were also included [89]. Table 2 collects relevant information from studies, such as medical conditions, age, and the number of participants who concluded the interventions and/or observations.
Results of the search
Likewise, cross-sectional studies did not reflect significant changes in gut microbiota diversity. No significant differences were detected among diverse groups in the relative abundances of the major phyla or alpha diversity measures (Chao 1, Simpson, Shannon; Kruskal–Wallis H test) [70, 89,90,91,92,93,94,95,96].
Otherwise, relative abundance analysis showed a significant change at the genus level in older adults who conducted an exercise program for five weeks or more. The relative abundance of Clostridioides difficile was significantly reduced (P = 0.03) [97], Clostridium subcluster XIVa shows a reduction in women who perform endurance exercise for twelve weeks meanwhile the genus Bacteroides shows a significantly increased [98].
When comparing the relative abundance of control and exercise groups, as shown in Table 3, authors inform significant differences in Bacteroides and Subdoligranulum [89] and a significant increase of Phascolarctobacterium and Mitsuokella in the exercise group [99]. Differential abundance analysis between two intervention groups conducted at the genus level showed that Haemophilus, Butyricicoccus, Eubacterium hallii, and Ruminiclostridium were reduced. In contrast, Coprobacter, and uncultured bacterium (from Rhodospirillales order) increased in the intervention group compared with the control group (all FDR P < 0.1) [28].
Similarly, the results described by Magzal et al. [96] in a cross-sectional study including 39 older adults suffering insomnia and classified into the groups high and low PAL shows that Bifidobacterium, Clostridium sensu stricto 1, Catenibacterium, Peptococcus, Holdemanella, and Butyricicoccus are among the genera present in more active individuals. Less active people had a higher relative abundance of the genera Barnesiella, Blautia, Lachnoclostridium, Christensenellaceae R-7 group, and UCG-005 [96].
Few studies report significant presence or abundance at the species level, Fielding, and collaborators showed that Faecalibacterium prausnitzii, Barnesiella intestinihominis, Bacteroides caccae, and Clostridium citroniae were higher in older adults with high fitness profile; meanwhile, a reduction in Eubacterium biforme, Desulfovibrio D168, and Escherichia coli was observed when compared to the Low Fitness group in a cross-sectional study where 29 older women and men (70 years) performed a short physical battery [92].
Various studies aimed to establish a correlation between important indicators of physical activity status such as maximal oxygen consumption or VO2 peak (ml/kg/min) [90, 97, 100], based on the results of cardiorespiratory fitness, older adults were divided into two or three functional groups, where those with higher values oscillate between 22.17 ± 0.51 [90], 23.2 ± 5.8 [100], and 27.3 ± 4.6 (ml/kg/min) [97]. Older people with less VO2 peak values are shown as statistically significantly lower. Correlation analysis exhibit that gut dysbiosis is associated with the reduced exercise capacity of elderly patients with hypertension [90].
Similar findings are observed in a randomized controlled trial [100], where 17 aged adults were assigned to exercise (HIT + RT) or a control group, eventually, after 12 weeks program measurement of VO2max (mL·kg − 1·min − 1) was performed, posterior analysis showed that the Shannon's index was positively correlated with VO2max changes; results suggest an association between microbiota richness and cardiorespiratory fitness improvements [100]. Other correlation analyses between the baseline relative abundance of specific microbiota families and the changes in body composition and cardio metabolic parameters showed that Bifidobacteriaceae abundance was positively correlated with fat mass and negatively with muscle mass. Equally, Paraprevotellaceae and Prevotellaceae were negatively correlated with fat mass and positively with muscle mass [100].
Table 3 shows a trend in the method employed to determine gut microbiota composition using the 16S rDNA amplicon based NGS. Only one selected study indicated Next Generation Sequencing (NGS) [70]. Meanwhile, another reported analyzing available data from The American Gut Project (AGP) [94, 101]. Table 3 summarizes the results related to the diversity and composition of the gut microbiota. Only significant differences between groups are described.
A deeper analysis of functional predictions shows that some metagenomic functions were significantly different between exercise and control periods (P < 0.05; FDR < 0.3). Based on the KEGG database, functions related to genetic information processing and nucleotide metabolism were overrepresented after a 5-week endurance exercise program in older Japanese men [97]. A similar analysis shows some crucial differences in 26 metagenomic functions when comparing high-fitness (HF) and low-fitness (LF) aged people. The authors emphasize that the expression of glutathione peroxidase (K00432; GPx) was higher, whereas the remaining 25 functions were lower in HF when compared with LF. GPx was the most highly expressed function (2 to 20-fold increased) compared to all other significant KEGG IDs. [92].
The physical activity frequency is also related to the relative abundance of microbial pathways. Zhu and collaborators suggest that regular exercise significantly modulated microbial function in older people because of the functional analysis performed in samples recovered from the American Gut Project [28]. In synthesis, the relative abundances of 18 pathways were significantly higher. In comparison, the abundances of 5 of those pathways were significantly lower in the daily or regular exercise group (DRE) than in the never or rare exercise group (NRE). These pathways include vitamin-related pathways, nucleotide metabolism-related pathways, glucose metabolism, and amino acid metabolism [94]. Some studies have involved direct quantification methods, such as untargeted metabolomics; Results reported by Castro-Mejía et al., describe significantassociations ( >|0.2| r) for ten gut metabolites and five plasma metabolites with lifestyle co-variables, such as steps per day which correlated positively with mono and di-saccharides metabolism and negatively with amino acid and lipid metabolism. Also, they did not find an essential difference in the concentrations of Short-Chain Fatty Acids (SCFA) from the fecal metabolome according to the high o lower fitness phenotype [93]. In contrast, Magzal, and collaborators report higher concentrations of total SCFA in people with lower physical activity levels [96]; here, acetate was the most prevalent SCFA in both groups. Analysis of the difference in these volatile compounds revealed that the less active group had significantly higher concentrations of propionate, isobutyrate, and valerate compared with the more active group. The magnitude of the difference in concentration between the study groups was higher for propionate (η2 = 16). The less active group also had significantly higher concentrations of total fecal SCFAs, compared to the more active activity group, with a medium effect size (η2 = 08) [96].
Finally, we identified a high variability in the frequency of physical activity both in longitudinal and cross-sectional studies. In brief, randomized trials included exercise protocols between a) 5 weeks of endurance exercise program comprising three ergometer sessions per week [97]; b) Supervised resistance training sessions, twice weekly for six weeks [66]; c) 8-week exercise training randomized controlled trial comprised aerobic and resistance exercise [99]; d) 12-week comparative trials, between aerobic exercise training or trunk muscle training [98]; 12-week training program included high-intensity training and resistance training three times per week [100]. We consider this data is not enough to describe the effect of different exercise intensities and durations on the composition and function of the gut microbiota of older people.
Discussion
This review summarizes 15 studies involving physical activity, exercise, and gut microbiota changes. In brief, three randomized control trials and 11 cross-sectional trials were analyzed to determine whether performing an exercise program, or higher levels of PA, are consequent to changes in the diversity, abundance, and functional parameters of the gut microbiota of older adults. Similar to reports from other systematic reviews, there are no significant shifts in diversity metrics (Alpha and Beta). Here, only one study from recovery data reported a significant difference in Alpha diversity from overweight people with higher PAL. Contrary to similar findings reported by Barton et al. [63], the microbiota alpha diversity of elderly athletes defined by the Shannon and Simpson index and the Chao1 index did not differ from that of the controls [89].
The abundance of some bacteria is higher in aged people, after an exercise program, or in comparison with control groups, especially at the genus level (Table 3). Some of these bacteria are from the Lachnospira and Lachnospiraceae NK4A136 group, these microbiota members have been described as potentially beneficial [102], because they are producers of SCFA [28], and the synthesis of these organic acids is usually linked to important roles in maintaining colonic host health as an energy source, regulator of gene expression, and anti-inflammatory agents [103], which might be beneficial for the host.
Similar to other studies in non-older adults, some results included in this systematic review suggest that regular exercise significantly modulated microbial function in elderly individuals the data proportionate so far is limited and few studies have included extra analysis such as metabolomic assays or metagenomic approach, where microbial compounds and relative pathways related to physical activity could discover [94], in contrast, other studies including non-older adults have reported significant findings by using specific analysis techniques and combination of omics technologies [69, 104, 105].
Here we highlight the association between the relative abundance of gut microbiota and physical function [99] and a reduced exercise capacity that is negatively associated with the core gut microbiota [90, 91]. We also identified in this systematic review that similar to results presented in cross-sectional studies with young adults [58, 106], consumption of oxygen by older men and women is correlated with species richness and higher diversity of bacterial members of the gut microbiota [90, 97, 100], which reinforce the hypothesis that effect of PAL is more related to functional outcomes rather than compositional indicators (such as diversity or abundance) further investigation is required.
Associations between physical activity and gut microbiota have yet to be extensively studied in older adults. Existing publications focusing on young adults and athletes show consistent results related to the production of SCFAs [63]. Also, bacteria such as Akkermansia muciniphila and Faecalibacterium prausnitzi have been described in the past [107]. In addition, the health status of older adults involved in biological and gut microbiota studies might be a limiting factor since including people with insomnia [96], Arterial Hypertension, Dyslipidemia, Hyperglycemia, Prostatic Hyperplasia [97], Primary Hypertension [90], Osteoporotic Fractures [91], Overweight and Obesity [94], Cardio Vascular Disease, Type 2 Diabetes mellitus [95] is present in this systematic review. These medical conditions have been reported as modulators of gut microbiota composition [23, 108, 109]. However, data availability for older adults is limited, and comparing healthy and unhealthy subjects could be complicated to perform.
Although the response to exercise and augment of PAL has been extensively studied in diverse biologic systems, such as the mitochondria, the muscle, the liver, and the neurologic system, among others, it is still unclear whether these changes are related to the gut microbiota in older adults. Past reviews and animal studies have linked the possible physiological response to exercise with the community of microorganisms that inhabit the gut [53, 110, 111]. We did not find consistent results that may reflect the modifications of the gut microbiome in other physiological systems. Some bacteria taxa whose abundance changed are beneficial for aged people (Table 3), such is the case of the genus Oscillospira which is a promising candidate for the next generation -of probiotics because of its capacity to produce butyrate [112]. Faecalibacterium and Coprococcus have been correlated with host quality of life indicators in humans diagnosed with depression [113], and some species of the genus Bacteroides and Parabacteroides are more extraordinary producers of γ-aminobutyric acid (GABA) [114]. Similar findings are described for Faecalibacterium prausnitzi that, besides promoting the production of metabolites, have been related to the decrease of inflammatory markers in patients with Alzheimer's-type dementia [115]. Eubacterium hallii is also considered a SCFAs producer, especially propionate [116], thanks to metagenomics. It has been discovered that Subdoligranulum MGS (metagenomics species) was co-abundantly found with Akkermansia muciniphila [117], a promising biomarker for nutritional status [118].
Otherwise, the study provided by Fielding and collaborators [92] looks to describe a correlation between muscle function and gut microbiota through the colonization of mice with microbiota from highly functional older adults. Although results are inconclusive, authors stated that bacteria taxa at the family-level Prevotellaceae, genus level Barnesiella and Prevotella, and species-level Barnesiella intestine hominis might be involved in mechanisms related to the maintenance of muscle strength in older adults [92].
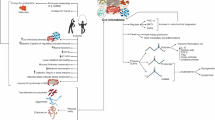
Deeper analysis performed in the studies included identifying metagenomic functions and metabolic pathways to describe some metabolic signatures related to vitamin, amino acid, and glucose functions. In contrast with other reports [33, 119, 120], we did not find an essential association between SCFA and physical activity in older adults. However, very few studies include metabolomic assays, and the data is limited (Fig. 2). This allows us to identify that including diverse sequencing methodologies and the analysis of related metabolites such as SCFAs and GABA in combination with metagenomic approximations could help to describe the role of physical activity on the gut microbiota of older adults in future studies.

Schematic representation of the data available from studies included in this systematic review. The amount of information available is mostly related to data from 16S rRNA sequencing and the identification of some bacteria associated to beneficial functions for the host; although very few studies used metagenomic approaches, some bacterial functions could be identified in future studies. Here, we identify only two studies describing SCFAs and results are inconclusive. Future directions could link the already known effect of exercise on brain and muscle function in older adults and the gut microbiome
Information on taxa and functions related to the benefits of performing PA has been relevant in the identification and isolation of probiotic candidates [121]. Also, including omics techniques, would give insights into the mechanisms underlying the effect of exercise on the gut microbiome of older adults and whether it differs from young people.
Conclusions
This review aimed to determine if there is any effect on the gut microbiota of adults older than 65 who start an exercise intervention or improve physical activity level. The studies identified do not address this systematic review's objectives. However, almost all the studies analyzed the diversity and abundance of the gut microbiota; there needs to be more information related to function and metabolic pathways that can be crucial to understand the effect of exercise and physical activity in older adults. It is essential to highlight the lack of randomized controlled trials in this field. Most of the studies included are observational, and interventions were mainly voluntary, based on physical exercise (aerobic or muscular) or to increase physical activity through lifestyle changes (increasing the number of steps). The lack of data related to gut microbiota analysis is a weakness that needs to be addressed in future studies.
Limitations of this review
Authors consider that some limitations of this review included publication bias because of one of the main criteria to report findings related to physical activity and gut microbiota of older adults, which was the statistical significance even though studies with results that do not show statistical significance may be clinically significant, and thus important to the findings of a systematic review. We also consider that the selection and inclusion of cross-sectional studies could be a potential limitation in this review. This is the first time that physical activity, microbiota, and older adults are compared in a systematic review.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Abbreviations
- OTUs:
-
Operational Taxonomic Units
- WHO:
-
World Health Organization
- ASV:
-
Amplicon Sequence Variance
- T2D:
-
Type 2 Diabetes
- SVs:
-
Sequence Variants
- PRISMA:
-
Preferred Reporting Items for Systematic Reviews
- AGP:
-
American Gut Project
- FGAS:
-
Frändin–Grimby Activity Scale
- PAL:
-
Physical Activity Level
- NGS:
-
Next-generation sequencing
- PA:
-
Physical Activity
- HF:
-
High Fitness
- LF:
-
Low fitness
- HIIT:
-
Hight Interval Intensity Training
- RT:
-
Resistance Training
- SCFA:
-
Short-Chain Fatty Acids
- GABA:
-
γ-Aminobutyric acid
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- DRE:
-
Regular Exercise Group
- NRE:
-
Never or Rare Exercise group
References
Partridge L, Deelen J, Slagboom PE. Facing up to the global challenges of ageing. Nature. 2018;561(7721):45–56.
López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;153(6):1194–217.
Gems D, Partridge L. Genetics of longevity in model organisms: debates and paradigm shifts. Annu Rev Physiol. 2013;75(1):621–44.
O’Toole PW, Shiels PG. The role of the microbiota in sedentary lifestyle disorders and ageing: lessons from the animal kingdom. J Intern Med. 2020;287(3):271–82.
Schmauck-Medina T, Molière A, Lautrup S, Zhang J, Chlopicki S, Madsen HB, et al. New hallmarks of ageing: A 2022 Copenhagen ageing meeting summary. Aging 2022;14. Available from: https://www.aging-us.com/article/204248/text.Cited 2022 Aug 28.
Miller ET, Svanbäck R, Bohannan BJM. Microbiomes as metacommunities: understanding host-associated microbes through metacommunity ecology. Trends Ecol Evol. 2018;33(12):926–35.
Asnicar F, Berry SE, Valdes AM, Nguyen LH, Piccinno G, Drew DA, et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. Nat Med. 2021;27(2):321–32.
Tan AH, Lim SY, Lang AE. The microbiome–gut–brain axis in Parkinson disease — from basic research to the clinic. Nat Rev Neurol. 2022;18(8):476–95.
Mitchell CM, Mazzoni C, Hogstrom L, Bryant A, Bergerat A, Cher A, et al. Delivery mode affects stability of early infant gut microbiota. Cell Rep Med. 2020;1(9): 100156.
Bayer G, Ganobis CM, Allen-Vercoe E, Philpott DJ. Defined gut microbial communities: promising tools to understand and combat disease. Microbes Infect. 2021;27: 104816.
Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez-Bello MG, Contreras M, et al. Human gut microbiome viewed across age and geography. Nature. 2012;486(7402):222–7.
Walker RL, Vlamakis H, Lee JWJ, Besse LA, Xanthakis V, Vasan RS, et al. Population study of the gut microbiome: associations with diet, lifestyle, and cardiometabolic disease. Genome Med. 2021;13(1):188.
Rinninella E, Raoul P, Cintoni M, Franceschi F, Miggiano GAD, Gasbarrini A, et al. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms. 2019;7(1):14.
Plovier H, Cani PD. Microbial impact on host metabolism: opportunities for novel treatments of nutritional disorders? Microbiol Spectr. 2017;5(3).
Li M, Wang M, Donovan SM. Early development of the gut microbiome and immune-mediated childhood disorders. Semin Reprod Med. 2014;32:74.
Bajinka O, Tan Y, Abdelhalim KA, Ozdemir G, Qiu X. Extrinsic factors influencing gut microbes, the immediate consequences and restoring eubiosis. AMB Express. 2020;10(1):130.
Liu W, Wang H, Wang Y, Li H, Ji L. Metabolic factors-triggered inflammatory response drives antidepressant effects of exercise in CUMS rats. Psychiatry Res. 2015;228(3):257–64.
Dieterich W, Schink M, Zopf Y. Microbiota in the Gastrointestinal Tract. Med Sci. 2018;6(4). Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6313343/.Cited 2020 Jul 30.
Maslowski KM. Metabolism at the centre of the host–microbe relationship. Clin Exp Immunol. 2019;197(2):193–204.
Fu X, Liu Z, Zhu C, Mou H, Kong Q. Nondigestible carbohydrates, butyrate, and butyrate-producing bacteria. Crit Rev Food Sci Nutr. 2019;59(sup1):S130–52.
González Hernández MA, Canfora EE, Jocken JWE, Blaak EE. The short-chain fatty acid acetate in body weight control and insulin sensitivity. Nutrients. 2019;11(8):1943.
Fan Y, Pedersen O. Gut microbiota in human metabolic health and disease. Nat Rev Microbiol. 2021;19(1):55–71.
Liu Y, Wang Y, Ni Y, Cheung CKY, Lam KSL, Wang Y, et al. Gut Microbiome fermentation determines the efficacy of exercise for diabetes prevention. Cell Metab. 2020;31(1):77-91.e5.
Scheiman J, Luber JM, Chavkin TA, MacDonald T, Tung A, Pham LD, et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat Med. 2019;25(7):1104–9.
Tian P, Bastiaanssen TFS, Song L, Jiang B, Jiang B, Zhang X, et al. Unraveling the microbial mechanisms underlying the psychobiotic potential of a bifidobacterium breve strain. Mol Nutr Food Res. 2021;65:e200074.
Wilmanski T, Diener C, Rappaport N, Patwardhan S, Wiedrick J, Lapidus J, et al. Gut microbiome pattern reflects healthy ageing and predicts survival in humans. Nat Metab. 2021;3(2):274–86.
Perez M, Ntemiri A, Tan H, Harris HMB, Roager HM, Ribière C, et al. A synthetic consortium of 100 gut commensals modulates the composition and function in a colon model of the microbiome of elderly subjects. Gut Microbes. 2021;13(1):1–19.
Muralidharan J, Moreno-Indias I, Bulló M, Lopez JV, Corella D, Castañer O, et al. Effect on gut microbiota of a 1-y lifestyle intervention with Mediterranean diet compared with energy-reduced Mediterranean diet and physical activity promotion: PREDIMED-Plus Study. Am J Clin Nutr. 2021;114(3):1148–58.
Hohman LS, Osborne LC. A gut-centric view of aging: Do intestinal epithelial cells contribute to age-associated microbiota changes, inflammaging, and immunosenescence? Aging Cell. 2022;21:e13700.
Lee JY, Tsolis RM, Bäumler AJ. The microbiome and gut homeostasis. Science. 2022;377(6601):eabp9960.
Murtaza N, OCuív P, Morrison M. Diet and the microbiome. Gastroenterol Clin North Am. 2017;46(1):49–60.
Houghton D, Hardy T, Stewart C, Errington L, Day CP, Trenell MI, et al. Systematic review assessing the effectiveness of dietary intervention on gut microbiota in adults with type 2 diabetes. Diabetologia. 2018;61(8):1700–11.
Aya V, Flórez A, Perez L, Ramírez JD. Association between physical activity and changes in intestinal microbiota composition: a systematic review. PLoS ONE. 2021;16(2): e0247039.
Guthold R, Stevens GA, Riley LM, Bull FC. Worldwide trends in insufficient physical activity from 2001 to 2016: a pooled analysis of 358 population-based surveys with 1·9 million participants. Lancet Glob Health. 2018;6(10):e1077–86.
World Health Organization. Physical activity. Available from: https://www.who.int/news-room/fact-sheets/detail/physical-activity.Cited 2022 Aug 29.
Lee IM, Shiroma EJ, Lobelo F, Puska P, Blair SN, Katzmarzyk PT. Effect of physical inactivity on major non-communicable diseases worldwide: an analysis of burden of disease and life expectancy. The Lancet. 2012;380(9838):219–29.
Lefferts WK, Davis MM, Valentine RJ. Exercise as an aging mimetic: a new perspective on the mechanisms behind exercise as preventive medicine against age-related chronic disease. Front Physiol. 2022;13. Available from: https://www.frontiersin.org/articles/https://doi.org/10.3389/fphys.2022.866792.Cited 2022 Aug 29.
Carapeto PV, Aguayo-Mazzucato C. Effects of exercise on cellular and tissue aging. Aging. 2021;13(10):14522–43.
Ringholm S, Gudiksen A, Halling JF, Qoqaj A, Rasmussen PM, Prats C, et al. Impact of aging and lifelong exercise training on mitochondrial function and network connectivity in human skeletal muscle. J Gerontol Ser A. 2022:glac164.
El Assar M, Álvarez-Bustos A, Sosa P, Angulo J, Rodríguez-Mañas L. Effect of physical activity/exercise on oxidative stress and inflammation in muscle and vascular aging. Int J Mol Sci. 2022;23(15):8713.
Angulo J, El Assar M, Álvarez-Bustos A, Rodríguez-Mañas L. Physical activity and exercise: strategies to manage frailty. Redox Biol. 2020;35: 101513.
Seals DR, Nagy EE, Moreau KL. Aerobic exercise training and vascular function with ageing in healthy men and women. J Physiol. 2019;597(19):4901–14.
Khodadad Kashi S, Mirzazadeh ZS, Saatchian V. A systematic review and meta-analysis of resistance training on quality of life, depression, muscle strength, and functional exercise capacity in older adults aged 60 years or more. Biol Res Nurs. 2022;13:10998004221120944.
Valenzuela PL, Castillo-García A, Morales JS, Izquierdo M, Serra-Rexach JA, Santos-Lozano A, et al. Physical exercise in the oldest old. Compr Physiol. 2019;9(4):1281–304.
Garatachea N, Pareja-Galeano H, Sanchis-Gomar F, Santos-Lozano A, Fiuza-Luces C, Morán M, et al. Exercise attenuates the major hallmarks of aging. Rejuvenation Res. 2015;18(1):57–89.
De Sousa LA, Lopes V, Horta J, Espregueira-Mendes J, Andrade R, Rebelo-Marques A. Therapeutics that can potentially replicate or augment the anti-aging effects of physical exercise. Int J Mol Sci. 2022;23(17):9957.
Chapman S, Aslan S, Spence J, DeFina L, Keebler M, Didehbani N, et al. Shorter term aerobic exercise improves brain, cognition, and cardiovascular fitness in aging. Front Aging Neurosci. 2013;5. Available from: https://www.frontiersin.org/articles/https://doi.org/10.3389/fnagi.2013.00075.Cited 2023 Mar 21.
Chekroud SR, Gueorguieva R, Zheutlin AB, Paulus M, Krumholz HM, Krystal JH, et al. Association between physical exercise and mental health in 1·2 million individuals in the USA between 2011 and 2015: a cross-sectional study. Lancet Psychiatry. 2018;5(9):739–46.
Barad Z, Augusto J, Kelly ÁM. Exercise-induced modulation of neuroinflammation in ageing. J Physiol;n/a(n/a). Available from: https://onlinelibrary.wiley.com/doi/abs/https://doi.org/10.1113/JP282894.Cited 2023 Mar 21.
Zhang Y, Cao RY, Jia X, Li Q, Qiao L, Yan G, et al. Treadmill exercise promotes neuroprotection against cerebral ischemia-reperfusion injury via downregulation of pro-inflammatory mediators. Neuropsychiatr Dis Treat. 2016;12:3161–73.
Zhang J, Guo Y, Wang Y, Song L, Zhang R, Du Y. Long-term treadmill exercise attenuates Aβ burdens and astrocyte activation in APP/PS1 mouse model of Alzheimer’s disease. Neurosci Lett. 2018;14(666):70–7.
Zhang SS, Zhu L, Peng Y, Zhang L, Chao FL, Jiang L, et al. Long-term running exercise improves cognitive function and promotes microglial glucose metabolism and morphological plasticity in the hippocampus of APP/PS1 mice. J Neuroinflammation. 2022;19(1):34.
Ticinesi A, Lauretani F, Tana C, Nouvenne A, Ridolo E, Meschi T. Exercise and immune system as modulators of intestinal microbiome: implications for the gut-muscle axis hypothesis. Exerc Immunol Rev. 2019;25:84–95.
Clarke SF, Murphy EF, O’Sullivan O, Lucey AJ, Humphreys M, Hogan A, et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut. 2014;63(12):1913–20.
Estaki M, Pither J, Baumeister P, Little JP, Gill SK, Ghosh S, et al. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome. 2016;4(1):42.
Petersen LM, Bautista EJ, Nguyen H, Hanson BM, Chen L, Lek SH, et al. Community characteristics of the gut microbiomes of competitive cyclists. Microbiome. 2017;5(1):98.
Jang LG, Choi G, Kim SW, Kim BY, Lee S, Park H. The combination of sport and sport-specific diet is associated with characteristics of gut microbiota: an observational study. J Int Soc Sports Nutr. 2019;16(1):21.
Durk RP, Castillo E, Márquez-Magaña L, Grosicki GJ, Grosicki GJ, Bolter ND, et al. Gut microbiota composition is related to cardiorespiratory fitness in healthy young adults. Int J Sport Nutr Exerc Metab. 2019;29:249.
O’Donovan C, Madigan SM, Garcia-Perez I, Rankin A, Sullivan OO, Cotter PD. Distinct microbiome composition and metabolome exists across subgroups of elite Irish athletes. J Sci Med Sport. 2020;23:63.
Allen JM, Mailing LJ, Niemiro GM, Moore R, Cook MD, White BA, et al. Exercise alters gut microbiota composition and function in lean and obese humans. Med Sci Sports Exerc. 2018;50(4):747–57.
Munukka E, Ahtiainen JP, Puigbò P, Jalkanen S, Pahkala K, Keskitalo A, et al. Six-week endurance exercise alters gut metagenome that is not reflected in systemic metabolism in over-weight women. Front Microbiol. 2018;9:2323.
Taniguchi H, Tanisawa K, Sun X, Sun X, Kubo T, Hoshino Y, et al. Effects of short-term endurance exercise on gut microbiota in elderly men. Physiol Rep. 2018;6:e13935.
Barton W, Penney NC, Cronin O, Garcia-Perez I, Molloy MG, Holmes E, et al. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut. 2018;67(4):625–33.
Estaki M, Pither J, Baumeister P, Little JP, Gill SK, Ghosh S, et al. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome. 2016;084(1):42.
Hughes RL. A Review of the Role of the Gut Microbiome in Personalized Sports Nutrition. Front Nutr. 2020;6. Available from: https://www.frontiersin.org/articles/https://doi.org/10.3389/fnut.2019.00191/full.Cited 2020 Feb 6.
Moore JH, Smith KS, Chen D, Lamb DA, Smith MA, Osburn SC, et al. Exploring the effects of six weeks of resistance training on the fecal microbiome of older adult males: secondary analysis of a peanut protein supplemented randomized controlled trial. Sports. 2022;10:65.
Morita E, Yokoyama H, Imai D, Takeda R, Ota A, Kawai E, et al. Aerobic exercise training with brisk walking increases intestinal bacteroides in healthy elderly women. Nutrients. 2019;11:868.
Zhu Q, Jiang S, Du G. Effects of exercise frequency on the gut microbiota in elderly individuals. MicrobiologyOpen. 2020;9(8):e1053-undefined.
Castro-Mejía JL, Khakimov B, Krych Ł, Bülow J, Bechshøft RL, Højfeldt G, et al. Physical fitness in community-dwelling older adults is linked to dietary intake, gut microbiota, and metabolomic signatures. Aging Cell. 2020;19(3):e13105-undefined.
Fart F, Rajan SK, Wall R, Rangel I, Ganda-Mall JP, Tingo L, et al. Differences in gut microbiome composition between senior orienteering athletes and community-dwelling older adults. Nutrients. 2020;12(9):2610.
Šket R, Debevec T, Kublik S, Schloter M, Schoeller A, Murovec B, et al. Intestinal metagenomes and metabolomes in healthy young males: inactivity and hypoxia generated negative physiological symptoms precede microbial dysbiosis. Front Physiol. 2018;9:198.
Gut Microbiota Composition Is Related to Cardiorespiratory Fitness in Healthy Young Adults. - PubMed - NCBI. Available from: https://www.ncbi.nlm.nih.gov/pubmed/29989465.Cited 2019 Nov 25.
Whisner CM, Maldonado J, Dente B, Krajmalnik-Brown R, Bruening M. Diet, physical activity and screen time but not body mass index are associated with the gut microbiome of a diverse cohort of college students living in university housing: a cross-sectional study. BMC Microbiol. 2018;1218(1):210.
Bressa C, Bailén-Andrino M, Bailen-Andrino M, Pérez-Santiago JD, González-Soltero R, Pérez M, et al. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLOS ONE. 2017;12:e0171352.
Hampton-Marcell JT, Eshoo TW, Cook MD, Gilbert JA, Horswill CA, Poretsky RS. Comparative analysis of gut microbiota following changes in training volume among swimmers. Int J Sports Med. 2020;41:292.
Kern T, Blond MB, Hansen TH, Rosenkilde MM, Rosenkilde M, Quist JS, et al. Structured exercise alters the gut microbiota in humans with overweight and obesity-A randomized controlled trial. Int J Obes. 2020;44:125.
Strasser B, Wolters M, Weyh C, Krüger K, Ticinesi A. The effects of lifestyle and diet on gut microbiota composition, inflammation and muscle performance in our aging society. Nutrients. 2021;13(6):2045.
Moher D, Liberati A, Tetzlaff J, Altman DG, Group TP. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. PLOS Med. 2009;6(7):e1000097.
Lin H, Peddada SD. Analysis of microbial compositions: a review of normalization and differential abundance analysis. Npj Biofilms Microbiomes. 2020;6(1):1–13.
Lin H, Peddada SD, Peddada SD. Analysis of compositions of microbiomes with bias correction. Nat Commun. 2020;11(1):3514.
Schloss PD. The effects of alignment quality, distance calculation method, sequence filtering, and region on the analysis of 16S rRNA gene-based studies. PLoS Comput Biol. 2010;6(7): e1000844.
Kim BR, Shin J, Guevarra R, Lee JH, Kim DW, Seol KH, et al. Deciphering diversity indices for a better understanding of microbial communities. J Microbiol Biotechnol. 2017;27(12):2089–93.
Anderson MJ, Crist TO, Chase JM, Vellend M, Inouye BD, Freestone AL, et al. Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist. Ecol Lett. 2011;14(1):19–28.
Walters KE, Martiny JBH. Alpha-, beta-, and gamma-diversity of bacteria varies across habitats. PLoS ONE. 2020;15(9): e0233872.
Anderson MJ, Ellingsen KE, McArdle BH. Multivariate dispersion as a measure of beta diversity. Ecol Lett. 2006;9(6):683–93.
Sterne JA, Hernán MA, Reeves BC, Savović J, Berkman ND, Viswanathan M, et al. ROBINS-I: a tool for assessing risk of bias in non-randomised studies of interventions. BMJ. 2016;12(355): i4919.
Higgins JPT, Altman DG, Gøtzsche PC, Jüni P, Moher D, Oxman AD, et al. The cochrane collaboration’s tool for assessing risk of bias in randomised trials. The BMJ. 2011;18(343): d5928.
Sterne JAC, Savović J, Page MJ, Elbers RG, Blencowe NS, Boutron I, et al. RoB 2: a revised tool for assessing risk of bias in randomised trials. BMJ. 2019;28(366): l4898.
Soltys K, Lendvorsky L, Hric I, Baranovicova E, Penesova A, Mikula I, et al. Strenuous physical training, physical fitness, body composition and bacteroides to prevotella ratio in the gut of elderly athletes. Front Physiol. 2021;1: 670989.
Yu Y, Mao G, Wang J, Zhu L, Lv X, Tong Q, et al. Gut dysbiosis is associated with the reduced exercise capacity of elderly patients with hypertension. Hypertens Res Off J Jpn Soc Hypertens. 2018;41(12):1036–44.
Langsetmo L, Johnson A, Demmer RT, Fino N, Orwoll ES, Ensrud KE, et al. The association between objectively measured physical activity and the gut microbiome among older community dwelling men. J Nutr Health Aging. 2019;23(6):538–46.
Fielding RA, Reeves AR, Jasuja R, Liu C, Barrett BB, Lustgarten MS. Muscle strength is increased in mice that are colonized with microbiota from high-functioning older adults. Exp Gerontol. 2019;127: 110722.
Castro-Mejía JL, Khakimov B, Krych Ł, Bülow J, Bechshøft RL, Højfeldt G, et al. Physical fitness in community-dwelling older adults is linked to dietary intake, gut microbiota, and metabolomic signatures. Aging Cell. 2020;19(3): e13105.
Zhu Q, Jiang S, Du G. Effects of exercise frequency on the gut microbiota in elderly individuals. MicrobiologyOpen. 2020;9(8): e1053.
Zhong X, Powell C, Phillips CM, Millar SR, Carson BP, Dowd KP, et al. The influence of different physical activity behaviours on the gut microbiota of older Irish adults. J Nutr Health Aging. 2021;25(7):854–61.
Magzal F, Shochat T, Haimov I, Tamir S, Asraf K, Tuchner-Arieli M, et al. Increased physical activity improves gut microbiota composition and reduces short-chain fatty acid concentrations in older adults with insomnia. Sci Rep. 2022;12(1):1–14.
Taniguchi H, Tanisawa K, Sun X, Kubo T, Hoshino Y, Hosokawa M, et al. Effects of short-term endurance exercise on gut microbiota in elderly men. Physiol Rep. 2018;6(23): e13935.
Morita E, Yokoyama H, Imai D, Takeda R, Ota A, Kawai E, et al. Aerobic exercise training with brisk walking increases intestinal bacteroides in healthy elderly women. Nutrients. 2019;11(4):868.
Zhong F, Wen X, Yang M, Lai HY, Momma H, Cheng L, et al. Effect of an 8-week exercise training on gut microbiota in physically inactive older women. Int J Sports Med. 2021;42(7):610–23.
Dupuit M, Rance M, Morel C, Bouillon P, Boscaro A, Martin V, et al. Effect of concurrent training on body composition and gut microbiota in postmenopausal women with overweight or obesity. Med Sci Sports Exerc. 2022;54(3):517–29.
McDonald D, Hyde E, Debelius JW, Morton JT, Gonzalez A, Ackermann G, et al. American gut: an open platform for citizen science microbiome research. mSystems. 2018;3(3):e00031.
Inoue T, Kobayashi Y, Mori N, Sakagawa M, Xiao JZ, Moritani T, et al. Effect of combined bifidobacteria supplementation and resistance training on cognitive function, body composition and bowel habits of healthy elderly subjects. Benef Microbes. 2018;9(6):843–53.
Mirzaei R, Dehkhodaie E, Bouzari B, Rahimi M, Gholestani A, Hosseini-Fard SR, et al. Dual role of microbiota-derived short-chain fatty acids on host and pathogen. Biomed Pharmacother. 2022;1(145): 112352.
Scheiman J, Luber JM, Chavkin TA, MacDonald T, Tung A, Pham LDD, et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat Med. 2019;25:1104.
Liu Y, Liu Y, Wang Y, Wang Y, Ni Y, Cheung CKY, et al. Gut microbiome fermentation determines the efficacy of exercise for diabetes prevention. Cell Metab. 2020;31:77.
Estaki M, Pither J, Baumeister P, Little JP, Gill SK, Ghosh S, et al. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome. 2016;4:42.
Frugé AD, Van der Pol W, Rogers LQ, Morrow CD, Tsuruta Y, Demark-Wahnefried W. Fecal akkermansia muciniphila is associated with body composition and microbiota diversity in overweight and obese women with breast cancer participating in a presurgical weight loss trial. J Acad Nutr Diet. 2020;120(4):650–9.
Kern T, Blond MB, Hansen TH, Rosenkilde M, Quist JS, Gram AS, et al. Structured exercise alters the gut microbiota in humans with overweight and obesity-A randomized controlled trial. Int J Obes 2005. 2020;44(1):125–35.
Brunt VE, Casso AG, Gioscia-Ryan RA, Sapinsley ZJ, Ziemba BP, Clayton ZS, et al. Gut microbiome-derived metabolite trimethylamine N-oxide induces aortic stiffening and increases systolic blood pressure with aging in mice and humans. Hypertens Dallas Tex 1979. 2021;78(2):499–511.
Ticinesi A, Mancabelli L, Tagliaferri S, Nouvenne A, Milani C, Del Rio D, et al. The gut-muscle axis in older subjects with low muscle mass and performance: a proof of concept study exploring fecal microbiota composition and function with shotgun metagenomics sequencing. Int J Mol Sci. 2020;21(23). Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7728056/.Cited 2020 Dec 28.
Ticinesi A, Tana C, Nouvenne A, Prati B, Lauretani F, Meschi T. Gut microbiota, cognitive frailty and dementia in older individuals: a systematic review. Clin Interv Aging. 2018;29(13):1497–511.
Yang J, Li Y, Wen Z, Liu W, Meng L, Huang H. Oscillospira - a candidate for the next-generation probiotics. Gut Microbes. 2021;13(1):1987783.
Valles-Colomer M, Falony G, Darzi Y, Tigchelaar EF, Wang J, Tito RY, et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat Microbiol. 2019;4(4):623–32.
Strandwitz P, Kim KH, Terekhova D, Liu JK, Sharma A, Levering J, et al. GABA-modulating bacteria of the human gut microbiota. Nat Microbiol. 2019;4(3):396–403.
Ueda A, Shinkai S, Shiroma H, Taniguchi Y, Tsuchida S, Kariya T, et al. Identification of faecalibacterium prausnitzii strains for gut microbiome-based intervention in alzheimer’s-type dementia. Cell Rep Med. 2021;2(9): 100398.
Engels C, Ruscheweyh HJ, Beerenwinkel N, Lacroix C, Schwab C. The Common Gut Microbe Eubacterium hallii also Contributes to Intestinal Propionate Formation. Front Microbiol. 2016;7. Available from: https://www.frontiersin.org/articles/https://doi.org/10.3389/fmicb.2016.00713/full#B12.Cited 2020 Oct 26.
Van Hul M, Le Roy T, Prifti E, Dao MC, Paquot A, Zucker JD, et al. From correlation to causality: the case of Subdoligranulum. Gut Microbes. 2020;12(1):1849998.
Zhang T, Li Q, Cheng L, Buch H, Zhang F. Akkermansia muciniphila is a promising probiotic. Microb Biotechnol. 2019;12(6):1109–25.
Clark A, Mach N. Exercise-induced stress behavior, gut-microbiota-brain axis and diet: a systematic review for athletes. J Int Soc Sports Nutr. 2016;13:43.
Ramos C, Gibson GR, Walton GE, Magistro D, Kinnear W, Hunter K. Systematic review of the effects of exercise and physical activity on the gut microbiome of older adults. Nutrients. 2022;14(3):674.
Giron M, Thomas M, Dardevet D, Chassard C, Savary-Auzeloux I. Gut microbes and muscle function: can probiotics make our muscles stronger? J Cachexia Sarcopenia Muscle. 2022;13:1460.
Acknowledgements
We thank Springer Nature for providing the waiver of the publication fees of this manuscript.
Funding
This work was funded by Dirección de Investigación e Innovación from Universidad del Rosario.
Author information
Authors and Affiliations
Contributions
VA and JDR: designed the study, conducted the analysis and drafted the manuscript. PJ and EM: extracted the information and validated the results. All authors approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplementary Table 1.
ROBINS-I risk of bias assessment summary: review authors' judgements about each methodological quality item for each non-randomized included study in this review.
Additional file 2: Supplementary Table 2.
A revised tool to assess risk of bias in randomized trials (RoB 2) summary: review authors' judgements about each methodological quality item for each randomized included study in this review.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Aya, V., Jimenez, P., Muñoz, E. et al. Effects of exercise and physical activity on gut microbiota composition and function in older adults: a systematic review. BMC Geriatr 23, 364 (2023). https://doi.org/10.1186/s12877-023-04066-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12877-023-04066-y