
Abstract
Davidia involucrata is a woody perennial and the only living species in the Genus Davidia. It is native to southern China where it holds cultural and scientific importance. However, D. involucrata is now an endangered species and its natural range includes low pH soils which are increasingly impacted by acid rain, nitrogen deposition and imbalanced nutrient cycling. The combination of these stresses also poses the additional risk of aluminum (Al) toxicity. Since the responses of D. involucrata to low pH and aluminum toxicity have not been investigated previously, a hydroponic experiment was conducted to examine the growth of one year old D. involucrata saplings after 50 d growth in a range of pH and Al conditions. Plant biomass, morphology, antioxidant enzyme activity, mineral concentrations and plant ecological strategy were compared at pH 5.8 and pH 4.0 without added Al (AlCl3) and in 0.1, 0.2 and 0.5 mM Al at pH 4.0. Our results showed that compared with pH 5.8, pH 4.0 (without added Al) not only inhibited root and shoot growth but also limited accumulation of nitrogen (N) and phosphorus (P) in leaves of D. involucrate. However, low Al concentrations (0.1 and 0.2 mM Al) at pH 4.0 partially restored the aboveground growth and leaf N concentrations, suggesting an alleviation of H+ toxicity by low Al concentrations. Compared with low Al concentrations, 0.5 mM Al treatment decreased plant growth and concentrations of N, P, and magnesium (Mg) in the leaves, which demonstrated the toxicity of high Al concentration. The results based on plant ecological strategy showed that D. involucrate decreased the competitiveness and favored its stress tolerance as pH changed from 5.8 to 4.0. Meanwhile, the competitiveness and stress tolerance of D. involucrata increased and decreased at low Al concentrations, respectively, and decreased and increased at high Al concentration, respectively. These trade-offs in ecological strategy were consistent with the responses of growth and antioxidant enzyme activity, reflecting a sensitive adaptation of D. involucrata to acid and Al stresses, which may aid in sustaining population dynamics. These findings are meaningful for understanding the population dynamics of D. involucrata in response to aluminum toxicity in acid soils.
Similar content being viewed by others


Introduction
Davidia involucrata Baill. (Family Davidiaceae) is a rare and endangered perennial tree species unique to China. This tree is a tertiary relict plant considered a “living fossil” and has been listed for priority state protection in the China Plant Red Data Book [1]. D. involucrata was once more widely distributed around the world but global climate changes in the Quaternary period decreased its distribution sharply. Currently, wild D. involucrata is largely restricted to the subtropical evergreen broad-leaved forests and the temperate deciduous broad-leaved forests of southwestern and south-central China [2, 3]. Reproduction and self-renewal are slow and difficult for this species and intensifying environmental stresses are decreasing its habitat further and increasing the risk of extinction [4]. Therefore, more study is required to understand the impact of specific environmental factors on fitness of D. involucrata to protect the remaining populations. Current research in this area mainly focuses on the impact of climate change factors such as temperature and water stress [1, 5, 6], while much less attention is being paid to soil factors.
As important limiting factors for plant growth and development, soil acidification and the accompanying increase of toxic aluminum (Al) are one of the important potential soil threats to the stability and sustainability of the wild population of D. involucrata. On the one hand, the distribution range of D. involucrata mainly comprises yellow and yellow-brown soils with pH values ranging from 4.0 to 6.5 [7, 8]. Furthermore, the soil acidification can be exacerbated by acid rain, nitrogen deposition, organic matter accumulation, and imbalanced soil nutrient cycling [9,10,11]. On the other hand, many soils in southwestern China also have high silicon and aluminum contents so that continued acidification can increase the concentration of Al in the soil solution, affect plant growth and threaten population stability [12]. Therefore, understanding the fitness status of D. involucrata under different levels of Al3+ treatment will help develop effective measures to protect the remaining D. involucrata populations. Unfortunately, until now, there has been no study clarifying how the fitness of D. involucrata will change under the threat of increased aluminum ions caused by soil acidification.
Generally, soluble Al3+ cations have negative effects on plant growth with the inhibition of root growth and changes in morphology being early symptoms of toxicity [13]. The resulting root systems are smaller and damaged and have a reduced capacity to take up water and nutrients [14, 15]. Previous studies have demonstrated that many woody perennials species in tropical and boreal forests are well adapted to acidic soils and can tolerate high concentrations of soluble Al [16]. In contrast to the high sensitivity exhibited of many crop species, these acid-soil adapted species are hardly affected by Al, and can even show a stimulation in growth by low to moderate concentrations of Al. This means the combination of low pH and Al can be either toxic or beneficial to plant growth depending on the species and conditions [9, 17, 18]. Thus, based on current information it is not possible to predict how low pH and Al affects the fitness of D. involucrata saplings.
The change in growth status is often regarded as an important reflection of the fate of plants in changing environments, but growth is actually a comprehensive reflection of trade-offs among three aspects of plant ecological strategy: resource acquisition, resistance to stress, and recovery after interference (such as regeneration and reproduction ability). These trade-offs provide an important insight into how the fitness of plant populations changes in changing environments. Because by deeply comparing the ecological strategies of plants in these three aspects, we can better understand when and how plants respond to environmental changes. Previous studies have attempted to explain these internal trade-offs in plants exposed to toxic Al often based on specific phenotypic, physiological or biochemical parameters [17, 19, 20]. However, due to the one-sided effect of individual parameters on physiological and metabolic processes, it is difficult to reveal the overall trade-offs of plant ecological strategies based solely on subjective indicators. Therefore, further quantitative and comprehensive understanding of the trade-offs of plant ecological strategies of D. involucrata under the changing Al concentrations may be more conducive to understanding of the fate of this rare and endangered plant under the threat of toxic Al in the future.
In order to understand the growth response and coping strategies of D. involucrata in relation to the increase of Al concentration under the acid condition, a hydroponic experiment with saplings of D. involucrata was conducted. Our objectives were to answer three questions: (1) Does low pH affect the growth of D. involucrata saplings? (2) Do different Al concentrations have different effects on the growth of D. involucrata saplings? (3) How do D. involucrata saplings alter their ecological strategies in response to different Al concentrations?
Materials and methods
Plant material, hydroponic culture and Al treatment
The one-year-old saplings of D. involucrata were obtained from a nursery (Yezhixin, Sichuan province) where the plants were grown from seed. After removal of the initial growth substrate, the roots were thoroughly rinsed with demineralized water. Then 40 saplings were randomly selected and transferred into 10 L tanks containing 8 L nutrient solution at pH of 5.8, with two saplings per tank, respectively. The nutrient solution consisted of (in µM) 625 KCl, 113 NH4Cl, 500 Ca (NO3)2, 125 MgSO4, 50 KH2PO4, 9 Fe-K2EDTA, 0.25 MnCl2, 0.02 CuSO4, 0.05 ZnSO4, 1.25 H3BO3, 0.005 Na2MoO4, and 0.00025 CoCl2 [21]. The nutrient solutions were changed weekly. The saplings were kept in this solution for two weeks to adapt to the hydroponic environment. After that, the 20 tanks were randomly divided into five groups with each group containing 4 tanks including 8 saplings. One group was randomly selected as control and treated with the above-mentioned nutrient solution at pH of 5.8. The other four groups were treated with the above-mentioned nutrient solution containing 0, 0.1, 0.2 and 0.5 mM AlCl3, respectively, at pH of 4.0. Al was added with vigorous stirring from a 1 M AlCl3 stock, freshly prepared on the day of use. The nutrient solutions were aerated vigorously by two air stones and aquarium pumps and renewed every three days.
These saplings of D. involucrata were cultivated in a naturally lit, and well-ventilated greenhouse with a day temperature range of 26–31 oC, a night temperature range of 20–24 oC, and a relative humidity range of 78–84%. Saplings were grown in these various treatments for 50 days.
Growth measurements
Before the experimental treatments, the fresh plants were weighed (FWbefore) and the leaf numbers (LNbefore) were counted before they were transferred into the tanks. At the end of the experiments, the total fresh weight (FWafter) and leaf numbers (LNafter) of each plant were measured again. As a relative measure, fresh weight increment (ΔFW) was defined as: ΔFW = FWafter - FWbefore; leaf number increment (ΔLN) was defined as: ΔLN = LNafter - LNbefore.
After that, two relatively young but fully expanded leaves from the upper part of the plant were selected for determining leaf functional traits and plant ecological strategy. Another fully opened leaf of the terminal leaflets of the uppermost part of the plant was chosen for measuring antioxidant enzymes. The remainder of the leaves was scanned using a Portable Laser Area Meter (LI-3000 C, Li-Cor Inc., Lincoln, NE) for leaf area. The remainder of the leaves and the shoot were dried for 48 h at 70 °C and weighed for leaf dry weight and shoot dry weight. The dried leaves were used for plant elemental measurements. The total leaf area (TLA) and total aboveground dry weight (DWab) were determined (including the leaves used for plant ecological strategy and antioxidant measurements).
The roots were scanned in a flatbed scanner (Epson Expression 10000XL) at a resolution of 400 dpi. The total root length, average root diameter and root volume were measured using WinRHIZO (Regent Instruments Inc., Quebec, QC, Canada). The new roots were separated and weighed after drying (RDWnew). Similarly, the remainder of the roots were also dried at 70 °C for 48 h and weighed (RDWre). The total root dry weight (DWroot) was determined as: DWroot = RDWnew + RDWre. The total individual dry weight (TDW) was determined as: TDW = DWab + DWroot. The dry weight increment was calculated by the relative water content of the plant. The root/aboveground biomass ratio was calculated as DWroot divided by DWab.
Plant ecological strategy assessments
The plant ecological strategies of D. involucrata individuals were evaluated using the Competitor-Stress tolerator-Ruderal (CSR) ‘trade-off theory’ to explore the potential coping strategies of D. involucrata in response to Al treatments [22, 23]. According to the capacity of the plant in resource acquisition, stress tolerance and recovery after interference, life history strategy of plant can be defined as a trade-off in three primary strategies: competitive (C-strategy: more investment in large leaf or root size to acquire resources), stress tolerant (S-strategy: invest more resources into defensive traits or metabolic processes to protect tissue from stress damages), and ruderal (R-strategy: more investment in regeneration and reproduction). This plant ecological strategy ‘trade-off theory’ provides us a holistic perspective regarding the response of D. involucrata under different Al concentrations, especially when such CSR strategy trade-offs were quantified based on plant functional traits as reported by Pierce et al. (2017) [24].
According to the model reported by Pierce et al. (2017) [24], the CSR strategy trade-offs of D. involucrata individuals from different treatments were evaluated based on three fundamental traits: specific leaf area (SLA), leaf dry matter content (LDMC) and individual leaf area (LA), with the use of the spreadsheet calculation tool ‘StrateFy’. The shifting of coping strategies of D. involucrata along Al3+ concentrations gradient was analyzed by comparing the C, S and R percentage values of plant individuals under different treatments, respectively.
The SLA, LDMC and LA were measured according to the standardized methodologies detailed by Harguindeguy et al. (2013) [25]. The two leaves for plant ecological strategy measurements of each individual were scanned to obtain LA (scanned leaf area divided by leaf number, cm2), and then were immediately sealed in the plastic bags, saturated with water and CO2 by spraying water and breathing. The plastic bags were stored in a dark and cool box for a minimum of 12 to 24 h to achieve complete leaf turgidity. After that, these leaves were dried with soft paper towel and weighed for leaf water-saturated fresh weights. Leaf dry weight was recorded after dried at 70 °C for 48 h. Subsequently, SLA (leaf area/leaf dry weight, cm2g− 1) and LDMC (100 × leaf dry weight/leaf water-saturated fresh weight, %) were calculated.
Antioxidant enzymes measurements
The leaf for antioxidant enzymes measurements of each individual was scanned and weighed to obtain leaf area and leaf fresh weight for total leaf area and total dry weight calculations. The collected leaves were homogenized with a mortar and pestle using liquid nitrogen. The total superoxide dismutase (SOD) activity was determined by measuring its ability to inhibit the photochemical reduction of nitroblue tetrazolium (NBT) [26, 27]. Peroxidase activity (POD) was measured according to the method of Chance and Maehly [28] with guaiacol as an electron donor, and the absorbance of the supernatant was determined at 470 nm. The activity was expressed in units of enzyme activity per minute per gram of tissue (min− 1 g− 1 FM). One enzyme unit was defined as the amount of enzyme causing an absorbance change of 0.1 min− 1 under standard conditions. The reaction of malondialdehyde (MDA) with thiobarbituric acid would produce a reddish-brown product under high temperature and acidic conditions, and determined by a spectrophotometer [20].
Plant elemental measurements
The total leaf N concentration was measured by the semi-micro Kjeldahl method. The K, Ca and Mg concentrations of leaves and Al concentration of roots were determined by atomic absorption spectrophotometer [19]. The leaf P concentration was measured using an automatic intermittent chemical analyzer (Cleverchem 200) after digestion with HNO3 in a microwave oven [27].
Statistical analyses
Data were analyzed using SPSS 19.0 software (SPSS Inc., Chicago, IL, USA). One-way ANOVAs were used to determine differences among treatments. For ANOVAs the data were tested for normality and equality of variations and, if necessary, natural log transformations were performed. Significant differences were determined at 95% significant level.
Results
Growth response
Without added Al, the fresh weight increment, dry weight increment, total leaf area, leaf numbers increment and dry weight of new roots of the saplings became obviously lower, and the root/aboveground biomass ratio became significantly greater when the pH values changed from 5.8 to 4.0 (Fig. 1).

The fresh weight increment (a), dry weight increment (b), total leaf area (c), leaf number increment (d), dry weight of new root (e) and root/aboveground biomass ratio (f) under 0.0, 0.0, 0.1, 0.2 and 0.5 mM Al treatments. The white bars are pH 5.8 and the gray bars are pH 4.0. The data show means ± SE (n = 8). Different letters in the same sub-figure denote significant differences at the P < 0.05 level
At pH 4.0, compared with 0 mM Al, the fresh weight increment, dry weight increment, total leaf area and leaf numbers increments were respectively 81% (P < 0.05), 44% (P = 0.14), 72% (P < 0.05) and 112% (P < 0.05) greater at 0.1 mM Al, and were respectively 98%, 62%, 101% and 143% greater at 0.2 mM Al (P < 0.05), but were not obviously changed at 0.5 mM Al (Fig. 1a-d). By comparison, the dry weight of new roots was not affected by 0.1 and 0.2 mM Al treatments, but showed a significant decrease at 0.5 mM Al (Fig. 1e). The root/aboveground biomass ratios at 0.1, 0.2 and 0.5 mM Al were 14% (P = 0.07), 20% (P < 0.05) and 27% (P < 0.05) lower than that at 0 mM Al, respectively (Fig. 1f).
Root morphology
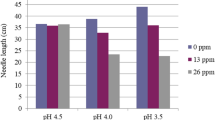
Without added Al, the root length, root volume, root surface area and root diameter of D. involucrata saplings were all significantly lower at pH 4.0 than these values at pH 5.8 (Fig. 2). For the Al treatments at pH 4.0, root length, root volume and root surface area showed small increases (~ 20%) at 0.2 mM Al and significant decreases (~ 27%) at 0.5 mM Al compared with control 0 mM Al (Fig. 2a-c).

The root length (a), root volume (b), root surface area (c), and root diameter (d) under 0.0, 0.0, 0.1, 0.2 and 0.5 mM Al treatments. The white bars are pH 5.8 and the gray bars are pH 4.0. The data show means ± SE (n = 8). Different letters in the same sub-figure denote significant differences at the P < 0.05 level
Concentrations of N, P, K, ca, mg in the leaves and Al in the roots
The leaf N and P concentrations of saplings at pH 4.0 (0 mM Al) treatment were significantly lower than these values at pH 5.8 treatment. At pH 4.0, along with the increase of Al concentrations from 0 mM, the leaf N concentrations significantly increased by 10% and 16% at 0.1 and 0.2 mM Al treatments and leaf P concentrations increased but not significantly (P = 0.061) at 0.1 and 0.2 mM Al treatments. However, at 0.5 mM Al, both N and P concentrations decreased to values similar to those at 0 mM Al (Fig. 3a, b).

The N (a), P (b), K (c), Ca (d) and Mg (e) concentrations in leaves and Al (f) in roots under 0.0, 0.0, 0.1, 0.2 and 0.5 mM Al treatments. The white bars are pH 5.8 and the gray bars are pH 4.0. The data show means ± SE (n = 4). Different letters in the same sub-figure denote significant differences at the P < 0.05 level
Leaf K and Ca concentrations were not affected by any Al treatment (Fig. 3c, d) while leaf Mg concentration showed a significant decrease at 0.5 mM Al compared with the other treatments (Fig. 3e). The Al concentration in the root tissues increased significantly as the external Al treatments increased in concentration (Fig. 3f).
Antioxidant enzyme activity in the leaves
The pH 4.0 (0 mM Al) treatment had no effect on POD and significantly increased SOD compared to levels measured at pH 5.8 (Fig. 4a, b). POD and MDA content of leaves showed no significant changes with any Al treatment compared with the pH 4.0 (0 mM Al) treatment (Fig. 4a, c). By comparison, SOD showed a 2.5-fold increase at 0.5 mM Al treatment compared to other Al treatments (Fig. 4b).

The peroxidase activity (POD) (a), superoxide dismutase activity (SOD) (b) and malondialdehyde content (MDA) (c) in leaves under 0.0, 0.0, 0.1, 0.2 and 0.5 mM Al treatments. The white bars are pH 5.8 and the gray bars are pH 4.0. The data show means ± SE (n = 5). Different letters in the same sub-figure denote significant differences at the P < 0.05 level
Leaf traits and plant ecological strategy
As shown in Fig. 5a and b, LA was significantly reduced and LDMC was significantly increased at pH 4.0 (0 mM Al) compared with the pH 5.8 treatment. When Al was added at pH 4.0, LA increased significantly at 0.1 and 0.2 mM Al by 42% and 49%, respectively, but decreased to a similar value as the 0 mM Al treatment at 0.5 mM Al. In contrast to the response of LA, compared with 0 mM Al treatment, LDMC displayed a decreasing trend at 0.1 and 0.2 mM Al and an increasing trend at 0.5 mM Al. Interestingly, compared to the 0.1 and 0.2 mM Al treatments, the 0.5 mM Al treatment significantly reduced LA and increased LDMC (Fig. 5a, b). Specific leaf area was not affected by any Al treatment (Fig. 5c).

The three functional traits: leaf area (a), leaf dry matter content (b), and specific leaf area (c), along with Grime’s CSR strategies: C-strategy (d), S-strategy (e), and R-strategy (f) of D. involucrata saplings under 0.0, 0.0, 0.1, 0.2 and 0.5 mM Al treatments. The white bars are pH 5.8 and the gray bars are pH 4.0. The data show means ± SE (n = 8). Different letters in the same sub-figure denote significant differences at the P < 0.05 level
According to the analysis using StrateFy, the D. involucrata saplings displayed a relatively weaker C-strategy and a significantly stronger S-strategy under pH 4.0 (0 mM Al) treatment compared to pH 5.8 treatment (Fig. 5d, e). At pH 4.0, compared with 0 mM Al treatment, the C-strategy increased by 11% (P < 0.05) and 7% (P = 0.058) in 0.1 and 0.2 mM Al while the S-strategy decreased by 73% (P < 0.05) and 41% (P = 0.068) at 0.1 and 0.2 mM Al treatments, respectively (Fig. 5d and e). By contrast, the 0.5 mM Al treatment significantly reduced C-strategy and greatly increased S-strategy, compared with 0.1 and 0.2 mM Al treatments (Fig. 5d, e). None of the pH or Al treatments had any effect on the R-strategy (Fig. 5f).
Discussion
Restricted growth of D. Involucrata under the acidic condition
Low pH has been reported to have various negative effects on morphology, physiology, and biomass accumulation in plants [29, 30]. In line with these previous reports, we found the dry weight of new roots and the increments of fresh weight, dry weight, leaf numbers and total leaf area of D. involucrata saplings were lower at pH 4.0 (0 mM Al) compared with pH 5.8.This suggested that the low soil pH condition, as well as the potential for further declines in soil pH value in the future, would pose a great threat to the growth of D. involucrata.
One of the reasons for this phenomenon of plant growth restriction caused by low pH may be that the increase of hydrogen ion concentration outside the roots inhibited the absorption of nutrients by the roots of D. involucrata, thus limiting the normal metabolism of the plants and the accumulation rate of organic matter. Our results showed that under the condition of pH 4.0, the leaf nitrogen and phosphorus contents were significantly lower than those under the pH 5.8 treatment. The study by Long et al. (2017) on Citrus sinensis and Citrus grandis also found that low pH not only significantly reduced the plant’s ability to absorb nutrients but also obviously inhibited the assimilation of carbon dioxide in the leaves [29]. According to previous studies at the plant cell level, the decrease in the amount of nutrient element absorption may be related to the inhibition of excess hydrogen ions on the function of plasma membrane H+-ATPase (PM H+-ATPase). Because PM H+-ATPase acts as a universal electrogenic H+ pump, and its function depends on the concentration gradient of H+ on both sides of the cell membrane. When the concentration of H+ in the environment is too high, this electrochemical proton gradient can be compromised, thereby limiting the transport of nutrients and other functions such as stomatal function [31].
The results also showed that the root/aboveground biomass ratio significantly increased, which indicated that aboveground plant properties responded more sensitively to low pH than root characteristics. This tendency of allocating more biomass underground implied a decrease in the efficiency of organic matter fixation through photosynthesis, per unit biomass, which also could be a significant factor contributing to the reduction in plant growth.
Concentration-dependent effects of aluminum ion
This study clearly showed that under the condition of pH 4, low concentration Al could significantly promote the aboveground growth of D. involucrata. For example, although low levels of Al (0.1 and 0.2 mM Al) had no effect on dry weight of new roots and root characteristics, the increments of fresh weight, dry weight, leaf numbers and total leaf area of D. involucrata were almost all significantly higher than those at 0 mM Al. This is similar to the promotion effect of low concentration aluminum treatment on the growth of Camellia japonica and maize as found by Liu et al. 2020 [32] and Wang et al. (2015) [33], respectively. However, inconsistent with our results, Rehmus et al. (2014) found that a low Al dose of 0.3 mM enhanced the root biomass of Tabebuia chrysantha tree seedlings, but did not improve shoot biomass [17]. This inconsistency may be attributed to the variations in the concentrations of nutrients in different organs. As suggested by Rehmus et al. (2014), the higher P concentrations in roots than those in the leaves may result in the root biomass increase at low concentration Al [17]. By comparison, our finding showed a higher N concentration in leaves of D. involucrata saplings at low Al concentrations (0.1 and 0.2 mM) compared with 0 mM Al treatment (pH 4.0). Increased N concentrations were likely beneficial to plant growth because N is necessary for protein synthesis. Therefore, as reported by Wang et al. (2015) who found a low Al concentration increased leaf protein content and thus promoted leaf growth [33], the increased leaf N concentrations may be one reason for the increased aboveground growth of D. involucrata under low pH.
This enhanced N concentration at low Al concentrations might be explained by alleviation of H+ toxicity. Previous studies showed that at low pH, low concentrations of Al could displace the highly toxic H+ ions from critical binding sites in the cell wall and on the plasma membrane [34]. They could also enhance PM H+-ATPase expression, thereby promoting H+ extrusion [35]. The enhanced PM H+-ATPase activity may regulate the function of nutrient transporters, such as ammonium transporters, to promote NH4+ uptake [36]. Because NH4+ uptake and assimilation are closely synchronized, the NH4+ assimilation would provide amino acids for protein synthesis and growth [36].
Our study also demonstrated that the growth of D. involucrata was inhibited by high Al concentration. For example, when the Al concentration was increased to 0.5 mM, the increments of fresh weight, dry weight, leaf numbers and total leaf area of the D. involucrata reduced to values similar to those of the 0 mM Al treatment. Moreover, consistent with the results found by Ryan et al. (2001) [37]and Hiranoet al. (2012) [38], the root growth and root traits of D. involucrata treated with 0.5 mM Al were worse than those treated with 0 mM Al. The inhibited absorption of nutrient elements may be one of the main reasons for the restricted growth of D. involucrata under high concentration aluminum treatment. Because we also found that under 0.5 mM Al treatment, leaf N and P concentrations decreased to the same level as under 0 mM Al treatment, and the leaf Mg concentration decreased to a significantly lower level than that under 0 mM Al treatment. The limited nutrient absorption capacity may be attributed to the reason that excess Al ions bind to the negatively charged phospholipid bilayers of the plasma membrane, which destabilizes the membrane potential and impedes the H+-ATPase’s ability to exclude protons, thereby affecting the transport of nutrients [39,40,41].
This concentration-dependent effect of Al suggested that low Al concentration in an acid environment may reduce the negative effect of low pH on the growth of D. involucrata. For the protection of D. involucrata population under the trend of soil acidification, we should be vigilant about the potential negative impacts of soil with high Al concentration on its growth.
Different coping strategies of D. Involucrata in response to acid and aluminum environments
Whether plants can cope with adverse environments through ecological strategy adjustments is the key to maintain their fitness as much as possible in stressful environments [42,43,44]. Our finding showed that D. involucrata saplings had a significantly greater S-strategy at pH 4.0 compared to pH 5.8, but had a relatively lower C-strategy, which is in line with the decreased biomass and increased SOD activity. These results indicated that under the acid condition, D. involucrata saplings adopted a trade-off strategy of reducing growth and enhancing resistance, thereby improving their survival ability.
Compared with 0 mM Al treatment (pH 4.0), C-strategy of D. involucrata saplings increased and S-strategy decreased at low Al concentrations (0.1 and 0.2 mM), but decreased and increased, respectively at 0.5 mM Al concentration. These changes were consistent not only with biomass increase at low Al concentrations (0.1 and 0.2 mM) and biomass decrease at 0.5 mM Al concentration, but also with the response of resistance physiological indicators. For instance, we found activation of antioxidants (POD and SOD) of D. involucrata saplings did not change at low Al concentrations, but SOD activity increased at 0.5 mM Al treatment. The trade-off between growth and resistance was also in line with the biochemical responses observed in previous studies. Specifically, under high Al stress, plants not only increased energy expenditure to enhance activation of antioxidants, but they also devoted more energy to exude organic acids or phenolic compounds [45]. These would alleviate Al toxicity within the plant or reduce Al concentration in the rhizosphere [14, 46, 47].
The strategic adjustments observed in D. involucrata suggested that, in response to acid and aluminum stress, the plant could modify its energy and resource allocation between growth and resistance. This adaptation provided an additional explanation for changes in growth patterns under varying Al conditions and might aid in sustaining population continuity. We believe that further research on the transformation of ecological strategies, as well as the Al concentration threshold for effective defense strategies, taking into account metabolic and gene expression changes, will benefit further understanding and predicting the population dynamics of D. involucrata under the worsening crisis of acid and aluminum stress.
Conclusions
The present study demonstrated that low pH (pH 4.0) inhibited the growth of D. involucrata saplings, and while low Al concentrations could alleviate the negative effects of low pH, as the Al concentration increased, the inhibitory effects on the growth were once again enhanced. Compared with pH 5.8, D. involucrata saplings adopted a trade-off strategy of reducing their competitiveness and increasing its resistance at pH 4.0. At pH 4.0, C-strategy and S-strategy were greater and lower respectively under low Al concentrations than those under 0 mM Al treatment, but as the Al concentration increased to 0.5 mM, these two strategies significantly decreased and increased, respectively. Our results illustrated that (1) the acid condition alone limited the growth of D. involucrate saplings, and the influence of Al depended on the concentration; (2) under the different aluminum treatments, D. involucrata can flexibly adjust its trade-off between competitive and stress tolerance strategies, which may better maintain its fitness and population continuation. Further studies on the growth changes and ecological strategies trade-off within a larger range of Al concentrations, along with research on related metabolic and gene expression processes, will benefit attempting to further explain and predict the dynamics of D. involucrata population under the Al stress.
Data availability
All data generated or analysed during this study are included in this published article.
References
Liu Q, Vetukuri RR, Xu W, Xu X. Transcriptomic responses of dove tree (Davidia Involucrata Baill.) To heat stress at the seedling stage. Forests. 2019;10. https://doi.org/10.3390/f10080656
Tang CQ, Dong Y-F, Herrando-Moraira S, Matsui T, Ohashi H, He L-Y, Nakao K, Tanaka N, Tomita M, Li X-S, et al. Potential effects of climate change on geographic distribution of the Tertiary relict tree species Davidia involucrata in China. Sci Rep. 2017;7:43822. https://doi.org/10.1038/srep43822
Long T, Tang J, Pilfold N, Zhao X, Dong T. Predicting range shifts of Davidia Involucrata Ball. Under future climate change. Ecol Evol. 2021;11:12779–89. https://doi.org/10.1002/ece3.8023
Wang Y, Wang Z, Xing H, Li J, Sen S. Prediction of potential suitable distribution of Davidia Involucrata Baill in China based on MaxEnt. Chin J Ecol. 2019;38:1230–7. https://doi.org/10.13292/j.1000-4890.201904.024
Liu Q, Feng Z, Xu W, Vetukuri RR, Xu X. Exogenous melatonin-stimulated transcriptomic alterations of Davidia involucrata seedlings under drought stress. Trees-Struct Funct. 2021;35:1025–38. https://doi.org/10.1007/s00468-021-02099-7
Liu Q, Xu L, Li Y, Xu W, Vetukuri RR, Xu X. Overexpression of an autophagy-related gene DiATG3 from Davidia involucrata improves plant thermotolerance by enhancing the accumulation of polyamines and regulating genes in calcium and MAPK signaling pathways. Environ Exp Bot. 2023;208. https://doi.org/10.1016/j.envexpbot.2023.105235
Li C, Peng X. Characteristics of soil nutrients in Davidia involucrata communities at different altitudes in Ya’an. J Sichuan Sci Technol. 2019;40:31–6. https://doi.org/10.16779/j.cnki.1003-5508.2019.03.006
Guo JH, Liu XJ, Zhang Y, Shen J, Han W, Zhang W, Christie P, Goulding K, Vitousek P, Zhang F. Significant acidification in major Chinese croplands. Science. 2010;327:1008–10. https://doi.org/10.1126/science.1182570
Bojorquez-Quintal E, Escalante-Magana C, Echevarria-Machado I, Martinez-Estevez M. Aluminum, a friend or foe of higher plants in acid soils. Frout Plant Sci. 2017;8:1767. https://doi.org/10.3389/fpls.2017.01767
Sade H, Meriga B, Surapu V, Gadi J, Sunita M, Suravajhala P, Kavi Kishor P. Toxicity and tolerance of aluminum in plants: tailoring plants to suit to acid soils. Biometals. 2016;29:187–210.
Ofoe R, Thomas RH, Asiedu SK, Wang-Pruski G, Fofana B, Abbey L. Aluminum in plant: benefits, toxicity and tolerance mechanisms. Frout Plant Sci. 2023;13:1085998. https://doi.org/10.3389/fpls.2022.1085998
Johnson AH, Siccama TG. Acid deposition and forest decline. Environ Sci Technol. 1983;17:A294–305. https://doi.org/10.1021/es00113a001. A.
Ryan PR, Ditomaso JM, Kochian LV. Aluminium toxicity in roots: an investigation of spatial sensitivity and the role of the root cap. J Exp Bot. 1993;44:437–46. https://doi.org/10.1093/jxb/44.2.437
Kochian LV, Piñeros MA, Liu J, Magalhaes JV. Plant adaptation to acid soils: the molecular basis for crop aluminum resistance. Annu Rev Plant Biol. 2015;66:571–98. https://doi.org/10.1146/annurev-arplant-043014-114822
Ma JF, Ryan PR. Understanding how plants cope with acid soils. Funct Plant Biol. 2010;37:iii–vi. https://doi.org/10.1071/FPv37n4_FO
Brunner I, Sperisen C. Aluminum exclusion and aluminum tolerance in woody plants. Frout Plant Sci. 2013;4:172. https://doi.org/10.3389/fpls.2013.00172
Rehmus A, Bigalke M, Valarezo C, Castillo JM, Wilcke W. Aluminum toxicity to tropical montane forest tree seedlings in southern Ecuador: response of biomass and plant morphology to elevated Al concentrations. Plant Soil. 2014;382:301–15. https://doi.org/10.1007/s11104-014-2110-0
Rehmus A, Bigalke M, Valarezo C, Castillo JM, Wilcke W. Aluminum toxicity to tropical montane forest tree seedlings in southern Ecuador: response of nutrient status to elevated Al concentrations. Plant Soil. 2015;388:87–97. https://doi.org/10.1007/s11104-014-2276-5
Naik D, Smith E, Cumming JR. Rhizosphere carbon deposition, oxidative stress and nutritional changes in two poplar species exposed to aluminum. Tree Physiol. 2009;29:423–36. https://doi.org/10.1093/treephys/tpn035
Guo P, Qi Y-P, Cai Y-T, Yang T-Y, Yang L-T, Huang Z-R, Chen L-S. Aluminum effects on photosynthesis, reactive oxygen species and methylglyoxal detoxification in two Citrus species differing in aluminum tolerance. Tree Physiol. 2018;38:1548–65. https://doi.org/10.1093/treephys/tpy035
Nguyen NT, Nakabayashi K, Thompson J, Fujita K. Role of exudation of organic acids and phosphate in aluminum tolerance of four tropical woody species. Tree Physiol. 2003;23:1041–50. https://doi.org/10.1093/treephys/23.15.1041
Grime JP. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. AmNat. 1977;111:1169–94. https://doi.org/10.1086/283244
Hodgson JG, Wilson PJ, Hunt R, Grime JP, Thompson K. Allocating C-S‐R plant functional types: a soft approach to a hard problem. Oikos. 1999;85:282–94. https://doi.org/10.2307/3546494
Pierce S, Negreiros D, Cerabolini BE, Kattge J, Díaz S, Kleyer M, Shipley B, Wright SJ, Soudzilovskaia NA, Onipchenko VG. A global method for calculating plant CSR ecological strategies applied across biomes world-wide. Funct Ecol. 2017;31:444–57. https://doi.org/10.1111/1365-2435.12722
Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte MS, Cornwell WK, Craine JM, Gurvich DE, et al. New handbook for standardised measurement of plant functional traits worldwide. Aust J Bot. 2013;61:167–234. https://doi.org/10.1071/bt12225
Beauchamp C, Fridovich I. Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Anal Biochem. 1971;44:276–87. https://doi.org/10.1016/0003-2697(71)90370-8
Zhang C, Chen M, Liu G, Huang G, Wang Y, Yang S, Xu X. Enhanced UV-B radiation aggravates negative effects more in females than in males of Morus alba saplings under drought stress. Environ Exp Bot. 2020;169:103903. https://doi.org/10.1016/j.envexpbot.2019.103903
Chance B, Maehly A. Assay of catalases and peroxidases. Methods Enzymol. 1955;2:764–75.
Long A, Zhang J, Yang L-T, Ye X, Lai N-W, Tan L-L, Lin D, Chen L-S. Effects of low pH on photosynthesis, related physiological parameters, and nutrient profiles of citrus. Frout Plant Sci. 2017;8:185. https://doi.org/10.3389/fpls.2017.00185
Zhang C-P, Meng P, Li J-Z, Wan X-C. Interactive effects of soil acidification and phosphorus deficiency on photosynthetic characteristics and growth in Juglans regia seedlings. Chin J Plant Ecol. 2014;38:1345–55. https://doi.org/10.3724/SP.J.1258.2014.00129
Li J, Guo Y, Yang Y. The molecular mechanism of plasma membrane H+-ATPases in plant responses to abiotic stress. J Genet Genomics. 2022;49:715–25. https://doi.org/10.1016/j.jgg.2022.05.007
Liu Y, Tao J, Cao J, Zeng Y, Li X, Ma J, Huang Z, Jiang M, Sun L. The beneficial effects of aluminum on the plant growth in Camellia japonica. J Soil Sci Plant Nutr. 2020;20:1799–809. https://doi.org/10.1007/s42729-020-00251-9
Wang L, Fan X-W, Pan J-L, Huang Z-B, Li Y-Z. Physiological characterization of maize tolerance to low dose of aluminum, highlighted by promoted leaf growth. Planta. 2015;242:1391–403. https://doi.org/10.1111/j.1365-2621.2003.tb14109.x
Kinraide TB. Aluminum enhancement of plant growth in acid rooting media. A case of reciprocal alleviation of toxicity by two toxic cations. Physiol Plant. 1993;88:619–25. https://doi.org/10.1111/j.1399-3054.1993.tb01380.x
Li Y, Zeng H, Xu F, Yan F, Xu W. H+-ATPases in plant growth and stress responses. Annu Rev Plant Biol. 2022;73:495–521. https://doi.org/10.1146/annurev-arplant-102820-114551
Zhang M, Wang Y, Chen X, Xu F, Ding M, Ye W, Kawai Y, Toda Y, Hayashi Y, Suzuki T. Plasma membrane H+-ATPase overexpression increases rice yield via simultaneous enhancement of nutrient uptake and photosynthesis. Nat Commun. 2021;12:735. https://doi.org/10.1038/s41467-021-20964-4
Ryan PR, Delhaize E, Jones DL. Function and mechanism of organic anion exudation from plant roots. Annu Rev Plant Physiol Plant Mol Biol. 2001;52:527–60. https://doi.org/10.1146/annurev.arplant.52.1.527
Hirano Y, Frey B, Brunner I. Contrasting reactions of roots of two coniferous tree species to aluminum stress. Environ Exp Bot. 2012;77:12–8. https://doi.org/10.1016/j.envexpbot.2011.10.007
Zhang J, Wei J, Li D, Kong X, Rengel Z, Chen L, Yang Y, Cui X, Chen Q. The role of the plasma membrane H+-ATPase in plant responses to aluminum toxicity. Frout Plant Sci. 2017;8:1757. https://doi.org/10.3389/fpls.2017.01757
Ahn SJ, Sivaguru M, Chung GC, Rengel Z, Matsumoto H. Aluminium-induced growth inhibition is associated with impaired efflux and influx of H+ across the plasma membrane in root apices of squash (Cucurbita pepo). J Exp Bot. 2002;53:1959–66. https://doi.org/10.1093/jxb/erf049
Gupta N, Gaurav SS, Kumar A. Molecular basis of aluminium toxicity in plants: a review. Am J Plant Sci. 2013;4:21–37. https://doi.org/10.4236/ajps.2013.412A3004
Morales M, Munné-Bosch S. Oxidative stress: a master regulator of plant trade-offs? Trends Plant Sci. 2016;21:996–9. https://doi.org/10.1016/j.tplants.2016.09.002
Wang J, Zhang C, Yang H, Mou C, Mo L, Luo P. Plant community ecological strategy assembly response to yak grazing in an alpine meadow on the eastern tibetan Plateau. Land Degrad Dev. 2018;29:2920–31. https://doi.org/10.1002/ldr.3050
He Z, Webster S, He SY. Growth–defense trade-offs in plants. Curr Biol. 2022;32:R634–9. https://doi.org/10.1016/j.cub.2022.04.070
Rahman SU, Han JC, Ahmad M, Ashraf MN, Khaliq MA, Yousaf M, Wang YC, Yasin G, Nawaz MF, Khan KA, et al. Aluminum phytotoxicity in acidic environments: a comprehensive review of plant tolerance and adaptation strategies. Ecotox Environ Safe. 2024;269:15. https://doi.org/10.1016/j.ecoenv.2023.115791
Cuenca G, Herrera R, Medina E. Aluminium tolerance in trees of a tropical cloud forest. Plant Soil. 1990;125:169–75.
Jones D, Ryan P. Aluminum toxicity. Encyclopedia of applied plant sciences: plant physiology and development. Elsevier; 2016. pp. 211–8.
Acknowledgements
Not applicable.
Funding
This work was supported by the Sichuan Science and Technology Program (2018SZDZX0036) and the Fundamental Research Funds of China West Normal University (18Q037).
Author information
Authors and Affiliations
Contributions
C. Z. designed the experiment, conceptualization and wrote the original draft. J. W., J. G., H. Y. and X. D. implemented the experiment. C. Z. and J. W. revised the manuscript.
Corresponding author
Ethics declarations
Ethics approval, guidelines and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, J., Guo, J., Yang, H. et al. Low levels of Al stimulate the aboveground growth of Davidia involucrata saplings. BMC Plant Biol 24, 465 (2024). https://doi.org/10.1186/s12870-024-05173-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-024-05173-7