
Abstract
Spike length (SL) is one of the most important agronomic traits affecting yield potential and stability in wheat. In this study, a major stable quantitative trait locus (QTL) for SL, i.e., qSl-2B, was detected in multiple environments in a recombinant inbred line (RIL) mapping population, KJ-RILs, derived from a cross between Kenong 9204 (KN9204) and Jing 411 (J411). The qSl-2B QTL was mapped to the 60.06–73.06 Mb region on chromosome 2B and could be identified in multiple mapping populations. An InDel molecular marker in the target region was developed based on a sequence analysis of the two parents. To further clarify the breeding use potential of qSl-2B, we analyzed its genetic effects and breeding selection effect using both the KJ-RIL population and a natural mapping population, which consisted of 316 breeding varieties/advanced lines. The results showed that the qSl-2B alleles from KN9204 showed inconsistent genetic effects on SL in the two mapping populations. Moreover, in the KJ-RILs population, the additive effects analysis of qSl-2B showed that additive effect was higher when both qSl-2D and qSl-5A harbor negative alleles under LN and HN. In China, a moderate selection utilization rate for qSl-2B was found in the Huanghuai winter wheat area and the selective utilization rate for qSl-2B continues to increase. The above findings provided a foundation for the genetic improvement of wheat SL in the future via molecular breeding strategies.
Key message
A major stable QTL for spike length was identified in wheat, and its genetic effect on yield related-traits as well as its potential use value in molecular breeding programs were characterized.
Similar content being viewed by others


Introduction
Wheat (Triticum aestivum L.) is a major food crop that is grown across the globe and increasing its yielding potential is important if world hunger levels are to be alleviated [1, 2]. Continuous population growth and the reduction in global arable land mean that it is particularly important to cultivate new wheat varieties with high and stable yield potentials [3].
Spike length (SL) is an important component of spike morphology in wheat and has an important influence on yield [2, 4]. Previous reports have indicated that SL was usually closely correlated with spike compactness (SCN), thousand kernel weight (TKW), and grain yield per plant (GYPP) [5,6,7,8,9]. In addition, Jantasuriyarat et al. [10] reported that SL was positively correlated with spikelet number per spike (SNPS) and that an allele from W7984 could have simultaneous genetic effects on both SCN and SL. Moghaddam et al. [11] reported that SL was positively correlated with shoot biomass, straw biomass per plant, and grain yield. Donmez et al. [12] reported that SL was positively correlated with harvest index, aboveground biomass, and grain yield after evaluating grain yield related traits. Therefore, identifying and verifying the genetic loci that control SL is crucial to the genetic improvement of wheat yields [3, 13].
Advances in molecular biology and quantitative genetics have led to the mapping of genes and numerous quantitative trait loci (QTLs) controlling SL on almost all the 21 chromosomes in wheat [13,14,15,16,17,18,19]. For example, the Q gene located on chromosome 5A allows the grain to be free-threshed and is closely related to SL, plant height (PH), and SCN [20, 21]. In addition, Zhang et al. [22] reported that TaAIRP2-1B was a regulator of SL in wheat; Ji et al. [23] mapped QSl.cib-5A on chromosome 5A and reported that it explained 7.88–26.6% of the phenotypic variation; Gao et al. [24] mapped QSL.caas-4AS and QSL.caas-4AL.1 on chromosome 4A and demonstrated that they individually explained 4.5–12.3% and 6.8–11.9% of the phenotypic variation, respectively; and Yu et al. [2] mapped 13 QTLs on chromosomes 1A, 1B, 3D, 4A, 5A, 5D, 6A, 7A, and 7B and showed that they individually explained 3.3–25% of the phenotypic variation. A QTL cluster associated with SL was also mapped onto chromosome 5A by Zhai et al. [25]. The abovementioned QTLs for SL have led to the breeding of wheat varieties with high and stable yields via molecular breeding technology. However, few studies have analyzed the genetic and breeding selection effects of the genes/QTLs related to SL.
Here, a major and stable quantitative trait locus (QTL) for SL, qSl-2B, was detected in multiple environments in a Kenong 9204 × Jing 411 derived recombinant inbred line (KJ-RIL) and was mapped to the 60.06–73.06 Mb region on chromosome 2B (based on the genome assembly for Kenong 9204, WheatOmics, http://202.194.139.32/). To further clarify its breeding potential, we analyzed the genetic and breeding selection effects of qSl-2B using the KJ-RIL population and a natural mapping population consisting of 316 breeding varieties/advanced lines. In addition, an InDel molecular marker, 2B-ID63, that is closely linked to qSl-2B was developed and the results showed that it improved wheat development based on molecular marker-assisted selection. Our results provided a foundation for the genetic improvement of wheat SL-related traits in the future and provided more molecular markers for molecular breeding programs that aimed to improve the yield potential of wheat.
Materials and methods
Plant materials and field trials
A total of 188 recombinant inbred line (RIL) populations, referred to “KJ-RILs”, derived from a cross between Kenong 9204 (KN9204) and Jing 411 (J411), were used in this study. Among them, KJ129 was not used in this study because a large number of the SNP markers were not present in the genotype. KN9204, J411, and the KJ-RILs were provided by the Center for Agricultural Resources Research, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences. The parents and the KJ-RIL populations were grown in ten different environments for phenotype evaluation. The low nitrogen (LN) environments were located in the following places: 2011–2012 in Shijiazhuang (E1-LN), 2012–2013 in Shijiazhuang (E3-LN), 2012–2013 in Beijing (E5-LN), 2012–2013 in Xinxiang (E7-LN), and 2013–2014 in Shijiazhuang (E9-LN). The high nitrogen (HN) environments were located in the following places: 2011–2012 in Shijiazhuang (E2-HN), 2012–2013 in Shijiazhuang (E4-HN), 2012–2013 in Beijing (E6-HN), 2012–2013 in Xinxiang (E8-HN), and 2013–2014 in Shijiazhuang (E10-HN). Detailed information regarding soil nitrogen content, experimental design, etc., can be found in our previous reports [4, 26,27,28,29,30]. The natural mapping population consisted of 316 breeding varieties/advanced lines and was planted in four environments for phenotype evaluation: 2020–2021 in Shijiazhuang, 2020–2021 in Weifang, 2020–2021 in Pulagu, and 2020–2021 in Yantai, China. A randomized block design with two replicates was used in each of the four environments. A three-row plot with rows that were 1.5 m long and 0.25 m apart was used, and 30 seeds were planted in each row. All experimental fields were routinely managed according to local standards.
Data collation and analysis
Based on the methods described in Cui et al. [16, 31], nine traits, SL, PH, grain length (GL), GYPP, spike number per plant (SNPP), TKW, SCN, kernel number per spike (KNPS), and SNPS, were measured in the natural mapping population and the KJ-RILs. QGAStation2.0 [32] was used to analyze the best linear unbiased estimators (BLUEs) for SL under LN (E1, E3, E5, E7, and E9) and HN (E2, E4, E6, E8, and E10) conditions and two datasets, BLUE-LN and BLUE-HN, were obtained. Both datasets were used in the QTL mapping analysis. Similarly, QGAStation 2.0 was used to analyze the BLUEs for yield-related traits in the natural mapping population grown in the four environments. A broad-sense heritability (H2) analysis was also performed using QGAStation 2.0. A one-way ANOVA and a phenotypic correlation analysis of SL with other traits were also performed using SPSS17.0 (https://spss-64bits.en.softonic.com/). The KJ-RIL and the natural mapping populations were used to perform an association analysis of the genotype and phenotypic data to estimate the genetic effect of qSl-2B on yield-related traits. FineBI 5.0 (https://www.finebi.com/product/fbi5) was used to analyze the selection effect of qSl-2B on the breeding process across the different regions and years.
Genotype acquisition and QTL detection
The DNA from the 187 KJ-RILs and the 316 accessions in the natural mapping population was extracted using the CTAB method according to Stacey et al. [33] with minor modification. The genotypes of the KJ-RIL and the natural mapping populations were screened using the 660 K and 55 K SNP arrays, respectively. The physical position of each SNP in the KN9204 genome assembly (unpublished data) was obtained by a local BLAST analysis (ftp://ftp.ncbi.nlm.nih.gov/blast/execuTABLEs/release/). A high density linkage map containing 119566 loci has been previously released by Cui et al. [34] and was based on a linkage analysis of the KJ-RIL population. In the present study, the physical positions rather than the genetic positions of the markers were used in the QTL mapping analysis.
Quantitative trait loci detection was performed using the IciMapping V4.2 BIP module format (http://www.isbreeding.net) with a logarithm of odds (LOD) threshold based on 1,000 permutations, 1.0 Mb as the step size, a stepwise regression of P = 0.001, and an I-error of 0.05.
Development of the InDel molecular markers
Based on a 10×genome resequencing of KN9204 and J411, a 51 bp insertion and deletion (InDel) polymorphic DNA sequence was identified at KN2B: 63085329 of the KN9204 genome. This was then used for polymorphic molecular marker development. PrimerServer in WheatOmics1.0 (http://202.194.139.32/) was used to design the polymorphic InDel PCR primers and the primer sequences are listed in Table S1. The amplification of InDel molecular markers in the KJ-RIL population and the linkage analysis indicated that this marker could be used as a diagnostic marker to identify qSl-2B in molecular breeding programs.
Results
Phenotypic analysis of spike length in the KJ-RILs
The phenotypic analysis of SL in the KJ-RIL mapping populations in the different environments are shown in Table 1. The SL mean in the KJ-RILs ranged from 6.69 cm to 9.55 cm and the absolute values of kurtosis and skewness were both less than 1, indicating that the phenotypic data for SL in the 187 KJ-RILs were approximately normally distributed. The phenotypic variability and the broad-sense heritability of SL in the KJ-RILs across the 10 environments were 8.17–10.22% and 0.52–0.92, respectively. Therefore, genetic factors were the main cause of phenotypic variation and SL was a typical quantitative trait that can be used in a QTL mapping analysis.
The correlation analyses between SL and other yield-related traits under LN and HN conditions are shown in Table 2. The results showed that SL was significantly positively correlated with KNPS, SNPS, PH, and GL under both LN and HN conditions, but significantly negatively correlated with SCN. There was no significant correlation between SL and GYPP, SNPP, or TKW.
QTL detection of qSl-2B
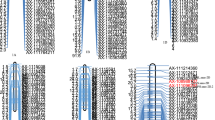
The QTL analysis showed that qSl-2B was consistently detected in the E4, E5, E6, E9, E10 as well as in the LN-BLUE and HN-BLUE datasets (Table 3; Fig. 1). It also explained 0.50–7.18% of the SL phenotypic variation in the KJ-RILs, with LOD values of 3.72–6.45. The favored alleles from KN9204 that augmenting SL, with the additive effect value ranging from 0.12 to 0.21. The physical information showed that qSl-2B was mapped within approximately 13 Mb at KN2B:60.06–73.06 Mb. Significantly, in five of the seven datasets shown in Table 3, qSl-2B was mapped to within 0.48 Mb (64.93–65.41 Mb) and was between the AX-109436459 and AX-111560829 SNPs. Furthermore, qSl-2B could be detected in three datasets representing the HN condition in Table 3 and the three datasets representing the LN condition, indicating that qSl-2B did not respond to nitrogen stress.

LOD value distributions for qSl-2B in the 10 environmental datasets and the 2 BLUE datasets. The abscissa shows the physical location of the SNP marker used for QTL positioning analysis in the reference genome of KN9204 and the ordinate is the LOD value corresponding to the different environments. The green segment is the estimated confidence region for qSl-2B and the markers in red were used in the subsequent genetic effect analysis
InDel molecular markers
Based on the genome DNA sequence information for KN9204 and J411, a total of 308 InDels (larger than 5 bp) and 357 SNPs were identified within 5.46 Mb (59.95–65.41 Mb) on chromosome 2B. A close linkage PCR-based InDel molecular marker, 2B-ID63 (63,085,329), for qSl-2B was developed. It could clearly and stably amplify the 421 bp and 472 bp target fragments from KN9204 and J411, respectively, and these PCR product sizes were consistent with the DNA re-sequencing results for the two parental lines (Fig. 2). Therefore, 2B-ID63 could be used to further analyze the genetic effects of the spike length and yield traits for wheat and provide molecular markers for the genetic improvement of yield potential.

Electrophoresis results for the PCR products amplified by 2B-ID63 in the 187 KJ-RILs. M: DNA marker. K: KN9204. J: J411. Numbers 1–48 are the amplified products in the KJ-RILs
Genetic effect of qSl-2B on yield-related traits in the KJ-RIL mapping population
The LOD peak position showed that AX-110741879 (KN2B: 59.95 Mb), AX-109425224 (KN2B: 60.19 Mb), AX-109436459 (KN2B: 64.93 Mb), AX-111560829 (KN2B: 65.41 Mb), AX-110566860 (KN2B: 72.03 Mb), AX-109316004 (KN2B: 73.42 Mb) and 2B-ID63 (KN2B: 63.09 Mb) was the close linkage markers for qSl-2B. Therefore, these markers were used to classify the 187 KJ-RILs into two groups. Since partial data for yield-related traits were missing in the E9 and E10 environments, the phenotypic data from the first 8 environments combined with genotypes were used to analyze for the genetic effect of qSl-2B on yield-related traits. The genotype identical to KN9204 was defined as Hap-KN9204 and the genotype identical to J411 was defined as Hap-J411. The results showed that the excellent alleles from KN9204 could significantly increase SL in four of the eight environment datasets (Fig. 3), with an average increase of 5.31%, but could only significantly increase SNPP in one environment, with an average increase rate of 10.25%. However, it significantly decreased SCN in seven of the eight environments with an average decrease of 7.19%, significantly decreased SNPS in one of the eight environments with an average decrease rate of 2.59%. The Hap-KN9204 allele had no significant effect on either GYPP, GL, TKW, PH or KNPS.

Genetic effects of qSl-2B on yield related traits in 8 of the 10 environments based on the KJ-RILs. SL: spike length, KNPS: kernel number per spike, SNPS: spikelet number per spike, PH: plant height, SCN: spike compactness, GL: grain length, GYPP: grain yield per plant, SNPP: spike number per plant, and TKW: thousand kernel weight. *, Significant difference at P < 0.05; **, highly significant difference at P < 0.01
Pyramiding effect analysis of qSl-2B under different genetic backgrounds
In addition to qSl-2B, we also identified two stable additive QTLs for SL in the KJ population, namely qSl-2D and qSl-5A [30]. The positive alleles of the increased SL of qSl-2D and qSl-5A were from J411. The pyramiding effect analysis for these two QTLs and qSl-2B was performed. Based on the genotype of tightly linked markers of these three QTLs, the KJ-RILs mapping population was divided into 8 groups. The results showed that as the number of positive alleles increases under LN and HN, the length of the SL also tends to be increase longer, which confirms the significant additive effect of all three QTLs (Fig. 4a). Compared with the phenotype of BAA, the pyramiding of the two favorable QTL alleles (BBB) increased SL by 14.54% and 15.18% under LN and HN, respectively.
In order to further illustrate additive effect of qSl-2B, we divided the KJ-RILs population into four groups based on genotype of tightly linked markers of qSl-2B, qSl-2D and qSl-5A. The results showed that the additive effect of qSl-2B ranged from 0.47 to -0.05 (Fig. 4b-e). Of these, the additive effect of qSl-2B for SL was significant in the backgrounds of AA (− −) and AB (− +) (Fig. 4b-c). The additive effect was higher when both qSl-2D and qSl-5A harbor negative alleles under LN and HN. In the two backgrounds of BA (+ −) and BB (+ +), qSl-2B had no significant effect on SL (Fig. 4d-e). When both qSl-2D and qSl-5A harbor positive alleles, the additive effect was negative. These results suggest that the expression of qSl-2B might be repressed in these backgrounds to some extent.

Pyramiding effect analysis of qSl-2B in different genetic backgrounds under LN and HN environments. (a) In the figure on the left, the Arabic numerals above each bar indicate the serial ID of the lines ranked according to SL value, from high to low SL. The Arabic numerals below each bar indicate number of positive alleles that increase SL. Arabic numerals at the bottom of each bar chart indicate the number of lines in each group. The figure on the right shows the origins of the positive and negative alleles for the three QTLs and the meaning of the code symbols. (b-e) The additive effects of qSl-2B in different genetic backgrounds under both HN and LN conditions. And the table at the bottom of in each bar chart shows the number of lines and the average SL (± SD) for each group. *, Significant difference at P < 0.05; **, highly significant difference at P < 0.01
Genetic and breeding selection effects of qSl-2B based on the natural mapping population
Three SNP molecular markers, i.e., AX-110426649 (TT/CC, KN2B: 60.21 Mb), AX-110383086 (TT/CC, KN2B: 65.64 Mb) and AX-108838208 (TT/CC, KN2B: 71.04 Mb), which were adjacent to the LOD peak position for qSl-2B, were used to genotype the 316 accessions in the natural mapping population (Table S2). The haplotype of Hap-CC-CC-TT, which was identical to KN9204, was defined as the excellent haplotype, Hap-TT-TT-CC, which was identical to J411, was defined as a non-excellent haplotype, and Hap-CC-TT-TT, Hap-CC-TT-CC, Hap-TT-CC-TT, Hap-TT-CC-CC, Hap-CC-CC-CC, and Hap-TT-TT-TT were defined as the recombinants. Heterozygous type is a line with a heterozygous genotype at any one to three of the three loci. Among the 316 accessions, the recombinants made-up the largest proportion, accounting for 34.81% of the total samples; the excellent haplotype and non-excellent haplotype accounting for 22.78% and 26.27%, respectively; and heterozygous type and missing data accounted for 5.70% and 10.44%, respectively (Table 4). These results indicated that qSl-2B was not fully utilized in traditional wheat breeding programs.
The genetic effects of qSl-2B on yield related traits were analyzed using the 316 accessions in the natural mapping population. The results showed that the excellent haplotype of Hap-CC-CC-TT could significantly increase TKW and SCN with average increase rates of 7.71% and 4.78%, respectively (Fig. 5). However, the Hap-CC-CC-TT haplotype significantly decreased SL, SNPS, SNPP, PH, and KNPS, with average decreases of 8.26%, 2.33%, 7.55%, 10.40%, and 5.40%%, respectively. However, its effects on GYPP and GL were not significant. Some of the results were inconsistent with those for the KJ-RIL population and may have been influenced by genetic background and environmental factors.

Genetic effects of qSl-2B on yield-related traits based on the 316 accessions in the natural mapping population. SL: spike length, KNPS: kernel number per spike, SNPS: spikelet number per spike, PH: plant height, SCN: spike compactness, GL: grain length, GYPP: grain yield per plant, SNPP: spike number per plant, TKW: thousand kernel weight. *, Significant difference at P < 0.05; **, highly significant difference at P < 0.01; ns: no significant difference
The breeding selection effects of qSl-2B at the spatio-temporal level were also analyzed based on geographical distribution using the natural mapping population (Fig. 6a). After discarding accessions with no detailed information, the 279 selected breeding varieties/advanced lines were divided into nine subgroups, which were those from Beijing, Hebei Province, Henan Province, Qinghai Province, Shandong Province, Shaanxi Province, Sichuan Province, Jiangsu Province, and countries/areas outside China. The results showed that the number of varieties with Hap-CC-CC-TT was highest in Jiangsu Province and Hebei Province, accounting for 50% and 43.75% of the total samples, respectively. In Shandong Province, Beijing and Henan Province, approximately 39.13%, 35.71% and 21.88% of the total samples, respectively, contained the excellent haplotype, but its utilization rates in Sichuan Province, Qinghai Province, and Shaanxi Province were lower, being 6.25%, 4.55%, and 0%, respectively. However, the utilization rates of the excellent haplotype were relatively low at 13.43%, but the non-excellent haplotype (Hap-TT-TT-CC) rate was 37.31% in the countries/areas that were outside China. The above results indicated that the utilization of qSl-2B considerably differed among regions/areas.
In terms of time span, the selection effect of qSl-2B was analyzed using 177 time-traceable varieties in the natural mapping population (Fig. 6b). In the 1990s, the non-excellent haplotype of Hap-TT-TT-CC was dominant and was present in up to 34.48% of the total samples, while the excellent haplotype of Hap-CC-CC-TT only accounted for 13.79% of the total samples. In the 2000s, the excellent haplotype proportion was 24.44%, while that of the non-excellent haplotype was 12.22% and those of the recombinants and the heterozygous type were 37.78% and 8.89%, respectively. In the 2010s, the excellent haplotype proportion was 32.76%, while that of the non-excellent haplotype was 27.59% and those of the recombinants and heterozygous type were 17.24% and 5.17%, respectively. These results indicated that wheat breeders have paid increasing attention to qSl-2B and applied its positive alleles to wheat breeding programs since the 1990s.

Analysis of the selection effect for qSl-2B haplotypes from the spatial (a) and time span (b) perspectives in breeding programs based on variety/advanced lines. Excellent Hap: Hap-CC-CC-TT; non-excellent Hap: Hap-TT-TT-CC; recombinants: Hap-CC-TT-TT, Hap-CC-TT-CC, Hap-TT-CC-TT, Hap-TT-CC-CC, Hap-CC-CC-CC, and Hap-TT-TT-TT; Heterozygous type is a line with a heterozygous genotype at any one to three of the three loci. Miss is a line where the haplotype data was missing. The corresponding radians of each haplotype in the chart were generated automatically by the software and have no practical significance
Discussion
Comparison of qSl-2B with that in previous studies
Spike length is a trait controlled by multiple genes and has important effects on wheat yield [35, 36]. Therefore, the identification and characterization of the QTL for SL is important when attempting to genetically improve yield potential based on a molecular breeding strategy. In this study, the SL of the KJ-RILs was evaluated in multiple environments under both LN and HN conditions. Among the five planting locations, except for the E7 and E8 in Xinxiang, the SL of the KJ-RILs was higher under HN than that under LN conditions. These findings indicated that SL was negatively affected by low nitrogen stress (Table 1). This result is consistent with previous reports, which suggested that nitrogen is an important nutrient for crop growth and development, including wheat [37,38,39].
Based on the genotypic data for the KJ-RILs, qSl-2B was mapped to the 60.06–73.06 Mb interval on chromosome 2B (based on the KN9204 genome assembly), and it was mapped to the KN2B: 64.93–65.41 Mb region in five of the 10 datasets, corresponding to the CS2B: 55.52–55.99 Mb section of the Chinese spring reference genome. To date, numerous QTLs for SL have been documented (http://wheatqtldb.net/yield_new.php) (Table S3) [9, 40,41,42,43,44,45,46,47,48,49,50,51]. Of these, Arif et al. [51] identified six QTLs controlling SL at 30.5 Mb, 144.65 Mb, 248.15 Mb, 546.61 Mb, 772.79 Mb, and 775.17 Mb on chromosome 2B, respectively. Under different soil moisture conditions, QSl.cob-2B.1 was stably detected in three of the four environments in a DH mapping population by El-Feki et al. [48]. This QTL, which was localized within the CS2B: 47.53–88.83 Mb section, coincided with the physical position of qSl-2B in this study. Based on a genome-wide association analysis using a wheat 90 K iSelect SNP genotyping assay, Sun et al. [47] identified 129 significant SNPs that were associated with SL, among which BS00022060_51 was located at CS2B: 52.7 Mb and therefore overlapped with qSl-2B. They also reported that it accounted for 4.4–21.0% of the phenotypic variation in this natural mapping population. The above results implied that qSl-2B could be identified in multiple mapping populations, which suggested that it may have important breeding applications.
According to meta-QTL analyses undertaken by recent studies [52,53,54], many other yield-related QTLs also coincide with the physical position of qSl-2B (Tables S3, S4, S5 and Fig. 7). A total of seven MQTLs on chromosome 2B were reported by Saini et al. [52] (Fig. 7a); of which MQTL2B.7 was located at CS2B: 42.28–59.18 Mb and harbored QTLs for TKW, SL, PH, GYPP and other yield-related traits. MQTL-2B-3 harbored QTL clusters for TKW, tiller number, KNPS, GYPP, and other yield-related traits that covered the CS2B: 50.71–80.12 Mb genome region (Fig. 7b) [53]. Zhang et al. [54] reported a QTL cluster called 2B.1 at CS2B: 44.8–106.00 Mb, which was associated with spikelet, GYPP, grain weight per tiller (GW), KNPS, and biomass (Fig. 7c). In addition, Zhang et al. [54] also found that the Ppd-B1 gene (gene bank number DQ885765) is nearby this QTL cluster (2B.1) at 56.2 Mb, in which the copy number variation affects yield-related traits such as GYPP, TKW, KNPS, biomass, spikelet, PH, and keycard. The above results indicated that gene clusters or genes with pleiotropic effects on yield-related traits might existed near qSl-2B and that more attention should be paid to the key chromosomal segments in molecular breeding programs designed to improve yield potential.

Meta-QTL and QTL clusters for spike-related traits on chromosome 2B reported in recent years. The figures a and b represent MQTL reported by Saini et al. [52] and Yang et al. [53], respectively. Figure c represents the QTL clusters reported by Zhang et al. [54]. The red segment on the three chromosomes is qSl-2B in the figure
Genetic effect analysis of qSl-2B effects on yield related traits
Spike length is an important yield-related trait and is closely related to TKW, KNPS, GYPP, etc. Qu et al. [55] reported that SL was significantly positively correlated with TKW; Liu et al. [56] found that there were significant positive correlations among SL, SNPS, and KNPS; Guo et al. [57] reported that SL was significantly positively correlated with sterile spikelet number (SSN) and KNPS; and Li et al. [58] showed that SL and PH were significantly positively correlated with each other. In this study, SL was significantly positively correlated with SNPS, KNPS, GL, and PH and significantly negatively correlated with SCN (Table 2). These results were generally consistent with previous studies.
We also clarified the genetic effects of qSl-2B on other yield traits using the KJ-RIL and the natural mapping populations. In the natural mapping population, the genetic effect of Hap-KN9204 on SL was opposite to that of the KJ-RILs with alleles from KN9204 increasing SL in the KJ-RILs, but decreasing SL in the natural mapping population (Figs. 3 and 5). Besides, Hap-KN9204 had a positive effect on SNPP and negative effect on SCN, which is also completely reversed in natural mapping populations. The large confidence interval for the primary QTL mapping results and the high recombination frequency in the target region might account for this inconsistency. Fine mapping and map-based cloning of qSl-2B would verify this suggestion. In addition, QTL mapping based on a bi-parental mapping population could only identify the alleles that produce improvements compared to the two given parental lines, whereas in the natural mapping population, all or at least most of the possible allelic variation due to the target gene could be identified, which meant that the best alleles for a given gene could be identified. This might also account for the different results for the genetic effect of qSl-2B in the two different mapping populations. Future haplotype analyses of the target gene could be used to confirm this suggestion. In addition, Hap-KN9204 had positive effect on TKW, and negative effect on PH and KNPS only in natural mapping populations. However, Hap-KN9204 had a negative effect on SNPS in both the KJ-RIL and the natural mapping populations. The reason for this result may be that gene clusters and a unique gene underlying qSl-2B could exist in the target region.
The qSl-2B selection utilization rate and future application potential
Traditional breeding technologies are mainly based on empirical selection for a specific phenotype, which is a slow process and the selection efficiency is generally low. Therefore, traditional breeding technologies cannot meet the demands associated with increasing wheat consumption through the genetic improvement of wheat yield potential. The 5G breeding approach will bring much-needed disruptive changes to crop improvement [59]. The key genes and close linkage molecular markers are very important when using the 5G breeding approach. To evaluate the potential application value of qSl-2B in future molecular breeding programs, we analyzed the selective utilization rate for qSl-2B in different regions and over different time periods. The results showed that the excellent Hap-CC-CC-TT haplotype from KN9204 was selected for use in some regions, but there were large differences among regions. In the regions outside China, the selective utilization rate for Hap-CC-CC-TT was low, indicating that breeders in countries outside China should pay more attention to this QTL. In China, Hap-CC-CC-TT selection was higher in Jiangsu, Shandong, and Hebei provinces, and Beijing particularly in the Huanghuai winter wheat area; but was lower in Henan Province, Beijing. Shanxi, Qinghai, and Sichuan. Different regions have different climates, soils, and other growth conditions and the focus associated with breeding for the genetic improvement of yield potential may vary. These variations might account for the large differences in the qSl-2B selection and utilization rates.
The selection and utilization of the excellent Hap-CC-CC-TT haplotype increased over time. This might have been due to the involvement of molecular breeding technology. Advances in molecular marker development have meant that QTLs associated with yield-related traits can be traced and used more efficiently. It is worth noting that the excellent Hap-CC-CC-TT haplotype selection effect is still relatively low and that qSl-2B still has great application potential in future breeding programs.
In conclusion, we remapped qSl-2B, a major QTL for SL and its genetic effects on yield-related traits were characterized using two different mapping populations. The breeding selection effect analysis indicated that qSl-2B had great application potential in future breeding programs. An InDel molecular marker in the target region was also developed and provided new molecular marker resources for wheat molecular breeding programs that aimed to genetically improve yield potential.
Data availability
All data supporting the results of this study are available within the paper and its supplementary data published online.
References
Ma Z, Zhao D, Zhang C, Zhang Z, Xue S, Lin F, Kong Z, Tian D, Luo Q. Molecular genetic analysis of five spike-related traits in wheat using RIL and immortalized F2 populations. Mol Genet Genomics. 2007;277:31–42.
Yu M, Mao SL, Chen GY, Pu ZE, Wei YM, Zheng YL. QTLs for uppermost internode and spike length in two wheat RIL populations and their affect upon plant height at an individual QTL level. Euphytica. 2014;200:95–108.
Li T, Deng G, Su Y, Yang Z, Tang Y, Wang J, Qiu X, Pu X, Li J, Liu Z, Zhang H, Liang J, Yang W, Yu M, Wei Y, Long H. Identification and validation of two major QTLs for spike compactness and length in bread wheat (Triticum aestivum L.) showing pleiotropic effects on yield-related traits. Theor Appl Genet. 2021;134:3625–41.
Cao M, Dong J, Wang H, Cai Y, Ma T, Zhou X, Xiao J, Li S, Chen L, Xu H, Zhao C, Wu Y, Sun H, Ji J, Cui F, Qin R. Identification of a major stable QTL for spikelet number in wheat (Triticum aestivum L.) and its genetic effects analysis on yield-related traits. Euphytica. 2002;218:96.
Schuler SF, Bacon RK, Gbur EE. Kernel and Spike Character Influence on Test Weight of Soft Red Winter Wheat. Crop Sci. 1994;34:1309–13.
Khaliq I, Irshad A, Ahsan M. Awns and flag Leaf Contribution towards Grain Yield in Spring Wheat (Triticum aestivum L). Cereal Res Commun. 2008;36:65–76.
Wu X, Cheng R, Xue S, Kong Z, Wan H, Li G, Huang Y, Jia H, Jia J, Zhang L, Ma Z. Precise mapping of a quantitative trait locus interval for spike length and grain weight in bread wheat (Triticum aestivum L). Mol Breed. 2014;33:129–38.
Yao H, Xie Q, Xue S, Luo J, Lu J, Kong Z, Wang Y, Zhai W, Lu N, Wei R, Yang Y, Han Y, Zhang Y, Jia H, Ma Z. HL2 on chromosome 7D of wheat (Triticum aestivum L.) regulates both head length and spikelet number. Theor Appl Genet. 2019;132:1789–97.
Liu H, Shi Z, Ma F, Xu Y, Han G, Zhang J, Liu D, An D. Identification and validation of plant height, spike length and spike compactness loci in common wheat (Triticum aestivum L). BMC Plant Biol. 2022;22:568.
Jantasuriyarat C, Vales MI, Watson CJ, Riera-Lizarazu O. Identification and mapping of genetic loci affecting the free-threshing habit and spike compactness in wheat (Triticum aestivum L). Theor Appl Genet. 2004;108:261–73.
Moghaddam M, Ehdaie B, Waines JG. Genetic variation and interrelationships of agronomic characters in landraces of bread wheat from southeastern Iran. Euphytica. 1997;95:361–9.
Donmez E, Sears RG, Shroyer JP, Paulsen GM. Genetic gain in yield attributes of Winter Wheat in the Great Plains. Crop Sci. 2001;239:114–23.
Xu X, Li X, Zhang D, Zhao J, Jiang X, Sun H, Ru Z. Identification and validation of QTLs for kernel number per spike and spike length in two founder genotypes of wheat. BMC Plant Biol. 2022;22:146.
Sourdille P, Tixier MH, Charmet G, Gay G, Cadalen T, Bernard S, Bernard M. Location of genes involved in ear compactness in wheat (Triticum aestivum) by means of molecular markers. Mol Breed. 2000;6:247–55.
Kumar N, Kulwal PL, Balyan HS, Gupta PK. QTL mapping for yield and yield contributing traits in two mapping populations of bread wheat. Mol Breed. 2007;19:163–77.
Cui F, Li J, Ding A, Zhao C, Wang L, Wang X, Li S, Bao Y, Li X, Feng D, Kong L, Wang H. Conditional QTL mapping for plant height with respect to the length of the spike and internode in two mapping populations of wheat. Theor Appl Genet. 2011;122:1517–36.
Cui F, Ding A, Li J, Zhao C, Wang L, Wang X, Qi X, Li X, Li G, Gao J, Wang H. QTL detection of seven spike-related traits and their genetic correlations in wheat using two related RIL populations. Euphytica. 2012;186:177–92.
Mladenov V, Dimitrijevic M, Petrović S, Boćanski J, Banjac B, Kondic-Spika A, Trkulja D. Genetic analysis of spike length in wheat. Genetika. 2019;51:167–78.
Kuang CH, Zhao XF, Yang K, Zhang ZP, Ding L, Pu ZE, Ma J, Jiang QT, Chen GY, Wang JR, Wei YM, Zheng YL, Li W. Mapping and characterization of major QTL for spike traits in common wheat. Physiol Mol Biol Plants. 2020;26:1295–307.
Simons KJ, Fellers JP, Trick HN, Zhang Z, Tai YS, Gill BS, Faris JD. Molecular characterization of the Major Wheat Domestication Gene Q. Genetics. 2006;172:547–55.
Xie Q, Li N, Yang Y, Lv Y, Yao H, Wei R, Sparkes DL, Ma Z. Pleiotropic effects of the wheat domestication gene Q on yield and grain morphology. Planta. 2018;247:1089–98.
Zhang J, Li C, Li L, Xi Y, Wang J, Mao X, Jing R. RING finger E3 ubiquitin ligase gene TaAIRP2-1B regulates spike length in wheat. J Exp Bot. 2023;13:erad226.
Ji G, Xu Z, Fan X, Zhou Q, Yu Q, Liu X, Liao S, Feng B, Wang T. Identification of a major and stable QTL on chromosome 5A confers spike length in wheat (Triticum aestivum L). Mol Breed. 2021;41:56.
Gao F, Wen W, Liu J, Rasheed A, Yin G, Xia X, Wu X, He Z. Genome-wide linkage mapping of QTL for Yield Components, Plant Height and Yield-related physiological traits in the Chinese Wheat Cross Zhou 8425B/Chinese Spring. Front Plant Sci. 2015;6:1099.
Zhai H, Feng Z, Li J, Liu X, Xiao S, Ni Z, Sun Q. QTL Analysis of Spike Morphological Traits and Plant Height in Winter Wheat (Triticum aestivum L.) using a high-density SNP and SSR-Based linkage map. Front Plant Sci. 2016;7:1617.
Cui F, Fan X, Zhao C, Chen M, Ji J, Zhang W, Li J. A novel genetic map of wheat: utility for mapping QTL for yield under different nitrogen treatments. BMC Genet. 2014;15:57.
Cui F, Fan X, Chen M, Zhang N, Zhao C, Zhang W, Han J, Ji J, Zhao X, Yang L, Zhao Z, Tong Y, Wang T, Li J. QTL detection for wheat kernel size and quality and the responses of these traits to low nitrogen stress. Theor Appl Genet. 2016;129(3):469–84.
Fan X, Zhang W, Zhang N, Chen M, Zheng S, Zhao C, Han j, Liu J, Zhang X, Song L, Ji J, Liu X, Ling H, Tong Y, Cui F, Wang T, Li J. Identification of QTL regions for seedling root traits and their effect on nitrogen use efficiency in wheat (Triticum aestivum L). Theor Appl Genet. 2018;131:2677–98.
Zhao C, Zhang N, Wu Y, Liu C, Fan X, Yan X, Xu H, Ji J, Cui F. QTL for spike-layer uniformity and their influence on yield-related traits in wheat. BMC Genet. 2019;20:23.
Fan X, Cui F, Ji J, Zhang W, Zhao X, Liu J, Meng D, Tong Y, Wang T, Li J. Dissection of Pleiotropic QTL Regions Controlling Wheat Spike Characteristics under Different Nitrogen Treatments Using Traditional and conditional QTL mapping. Front Plant Sci. 2019;10:187.
Cui F, Zhao C, Li J, Ding A, Li X, Bao Y, Li J, Ji J, Wang H. Kernel weight per spike: what contributes to it at the individual QTL level? Mol Breed. 2013;31:265–78.
Chen G, Zhu Z, Zhang F, Zhu J. Quantitative genetic analysis station for thegenetic analysis of complex traits. Chin Sci Bull. 2012;57:2721–6.
Stacey J, Isaac PG. Isolation of DNA from plants. Methods Mol Biol. 1994;28:9–15.
Cui F, Zhang N, Fan X, Zhang W, Zhao C, Yang L, Pan R, Chen M, Han J, Zhao X, Ji J, Tong Y, Zhang H, Jia J, Zhao G, Li J. Utilization of a Wheat660K SNP array-derived high-density genetic map for high-resolution mapping of a major QTL for kernel number. Sci Rep. 2017;7:3788.
Jin J, Liu D, Qi Y, Ma J, Zhen W. Major QTL for seven yield-related traits in common wheat (Triticum aestivum L). Front Genet. 2020;11:1012.
Li Y, Gao J, Zhang R, Song G, Zhang S, Li W, Li G. Identification of new QTL for yield-related traits in Chinese landrace and elite wheat varieties through a genome-wide linkage mapping. Euphytica. 2020;216:124.
Peng S, Buresh RJ, Huang J, Zhong X, Zou Y, Yang J, Wang G, Liu Y, Hu R, Tang Q, Cui K, Zhang F, Dobermann A. Improving nitrogen fertilization in rice by sitespecific N management. A review. Agron Sustain Dev. 2010;30:649–56.
Cormier F, Foulkes M, Hirel B, Gouache D, Moenne-Loccoz Y, Gouis JL. Breeding for increased nitrogen-use efficiency: a review for wheat (T. Aestivum L). Plant Breed. 2016;135:255–78.
Xing P, Zhang X, Li D, Wang H, Bao Y, Li X. Genome-wide association study identified novel genetic loci controlling internode lengths and plant height in common wheat under different nitrogen treatments. Euphytica. 2022;218:146.
Sourdille P, Cadalen T, Guyomarc’h H, Snape JW, Perretant MR, Charmet G, Boeuf C, Bernard S, Bernard M. An update of the courtot × Chinese spring intervarietal molecular marker linkage map for the QTL detection of agronomic traits in wheat. Theor Appl Genet. 2003;106:530–8.
Marza F, Bai GH, Carver BF, Zhou WC. Quantitative trait loci for yield and related traits in the wheat population Ning7840 × Clark. Theor Appl Genet. 2006;112:688–98.
Wu X, Chang X, Jing R. Genetic insight into yield-associated traits of wheat grown in multiple rain-fed environments. PLoS ONE. 2012;7:e31249.
Liu G, Jia L, Lu L, Qin D, Zhang J, Guan P, Ni Z, Yao Y, Sun Q, Peng H. Mapping QTLs of yield-related traits using RIL population derived from common wheat and tibetan semi-wild wheat. Theor Appl Genet. 2014;127:2415–32.
Xu Y, Wang R, Tong Y, Zhao H, Xie Q, Liu D, Zhang A, Li B, Xu H, An D. Mapping QTLs for yield and nitrogen-related traits in wheat: influence of nitrogen and phosphorus fertilization on QTL expression. Theor Appl Genet. 2014;127(1):59–72.
Narjesi V, Mardi M, Hervan EM, Azadi A, Naghavi MR, Ebrahimi M, Zali AA. Analysis of quantitative trait loci (QTL) for grain yield and agronomic traits in wheat (Triticum aestivum L.) under normal and salt-stress conditions. Plant Mol Biol Rep. 2015;33:2030–40.
Xu YF, Li SS, Li LH, Ma FF, Fu XY, Shi ZL, Xu HX, Ma PT, An DG. QTL mapping for yield and photosynthetic related traits under different water regimes in wheat. Mol Breed. 2017;37:34.
Sun C, Zhang F, Yan X, Zhang X, Dong Z, Cui D, Chen F. Genome-wide association study for 13 agronomic traits reveals distribution of superior alleles in bread wheat from the Yellow and Huai Valley of China. Plant Biotechnol J. 2017;15(8):953–69.
El-Feki WM, Byrne PF, Reid SD, Haley SD. Mapping quantitative trait loci for agronomic traits in Winter Wheat under different soil moisture levels. Agronomy. 2018;8:133.
Li F, Wen W, Liu J, Zhang Y, Cao S, He Z, Rasheed A, Jin H, Zhang C, Yan J, Zhang P, Wan Y, Xia X. Genetic architecture of grain yield in bread wheat based on genome-wide association studies. BMC Plant Biol. 2019;19:168.
Pretini N, Vanzetti LS, Terrile II, Donaire G, González FG. Mapping QTL for spike fertility and related traits in two doubled haploid wheat (Triticum aestivum L.) populations. BMC Plant Biol. 2021;21:353.
Arif MAR, Shokat S, Plieske J, Ganal M, Lohwasser U, Chesnokov YV, Kocherina NV, Kulwal P, Kumar N, McGuire PE, Sorrells ME, Qualset CO, Börner A. A SNP-based genetic dissection of versatile traits in bread wheat (Triticum aestivum L). Plant J. 2021;108:960–76.
Saini DK, Srivastava P, Pal N, Gupta PK. Meta-QTLs, Ortho-meta-QTLs and candidate genes for grain yield and associated traits in wheat (Triticum aestivum L). Theor Appl Genet. 2022;135(3):1049–81.
Yang Y, Amo A, Wei D, Chai Y, Zheng J, Qiao P, Cui C, Lu S, Chen L, Hu YG. Large-scale integration of meta-QTL and genome-wide association study discovers the genomic regions and candidate genes for yield and yield-related traits in bread wheat. Theor Appl Genet. 2021;134(9):3083–109.
Zhang J, She M, Yang R, Jiang Y, Qin Y, Zhai S, Balotf S, Zhao Y, Anwar M, Alhabbar Z, Juhász A, Chen J, Liu H, Liu Q, Zheng T, Yang F, Rong J, Chen K, Lu M, Islam S, Ma W. Yield-related QTL clusters and the potential candidate genes in two wheat DH populations. Int J Mol Sci. 2021;22:11934.
Qu X, Li C, Liu H, Liu J, Luo W, Xu Q, Tang H, Mu Y, Deng M, Pu Z, Ma J, Jiang Q, Chen G, Qi P, Jiang Y, Wei Y, Zheng Y, Lan X, Ma J. Quick mapping and characterization of a co-located kernel length and thousand-kernel weight-related QTL in wheat. Theor Appl Genet. 2022;135(8):2849–60.
Liu J, Xu Z, Fan X, Zhou Q, Cao J, Wang F, Ji G, Yang L, Feng B, Wang T. A Genome-Wide Association Study of Wheat Spike Related Traits in China. Front Plant Sci. 2018;9:1584.
Guo J, Shi W, Zhang Z, Cheng J, Sun D, Yu J, Li X, Guo P, Hao C. Association of yield-related traits in founder genotypes and derivatives of common wheat (Triticum aestivum L). BMC Plant Biol. 2018;18(1):38.
Li T, Li Q, Wang J, Yang Z, Tang Y, Su Y, Zhang J, Qiu X, Pu X, Pan Z, Zhang H, Liang J, Liu Z, Li J, Yan W, Yu M, Long H, Wei Y, Deng G. High-resolution detection of quantitative trait loci for seven important yield-related traits in wheat (Triticum aestivum L.) using a high-density SLAF-seq genetic map. BMC Genomic Data. 2022;23:37.
Varshney RK, Sinha P, Singh VK, Kumar A, Zhang Q, Bennetzen JL. 5Gs for crop genetic improvement. Curr Opin Plant Biol. 2020;56:190–6.
Acknowledgements
Not applicable.
Funding
This work was funded by Taishan scholar young expert (grant no. 20230119), the Major Basic Research Project of Natural Science Foundation of Shandong Province, China (grant no. ZR2019ZD16), the Key R & D Program of Shandong Province, China (Major Innovation Project) (grant no. 2022LZG002-2), the National Natural Science Foundation of China (grant nos. 32101726, 32072051 and 32272119), the Shandong Provincial Fund for Excellent Young Scholars (Grant No. ZR2022YQ19), the Natural Science Foundation of Shandong Province (ZR2022MC119), and the Yantai science and technology plan project (2023ZDCX023).
Author information
Authors and Affiliations
Contributions
FC, HD, CZ and XS analyzed the data and drafted the manuscript. HD, CW, YC, FW and XS performed the genotype identification and phenotype evaluation. YW, HS, RQ and CL helped design the study and revised the manuscript. All of the authors have read and approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Ethical approval and consent to participate
These experiments complied with the ethical standards in China.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ding, H., Wang, C., Cai, Y. et al. Characterization of a wheat stable QTL for spike length and its genetic effects on yield-related traits. BMC Plant Biol 24, 292 (2024). https://doi.org/10.1186/s12870-024-04963-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-024-04963-3