
Abstract
Background
Nutrients and heavy metals can be lost from soils via leaching, and arbuscular mycorrhizal fungi (AMF) can influence these events. Soil column experiments were carried out to examine whether leaching intensity and AMF can alter nutrient and Cd uptake in white clover plants and the extent of their losses through leaching.
Results
The presence of AMF significantly increased shoot and total biomass, as well as increased N, P, Cu and Zn uptake independent of water amount applied; while root P and Cu uptakes were promoted by AMF at any water amount treatments. Higher water amounts led to reductions in total N, K and Zn uptake for AMF-colonized plants in comparison to moderate water amount treatments. In the absence of AMF, white clover at low water amount treatment exhibited maximal root Cd uptake. At high water amount treatments, the presence of AMF significantly decreased leachate volumes and the amount of leached NH4+, K and Cd while AMF significantly increased the amounts of leached NO3−.
Conclusions
Overall we found that AMF-colonized white clover plants reduced NH4+, K and Cd loss from soils but increased the risk of NO3− loss under high intensity leaching conditions.
Similar content being viewed by others


Introduction
Nutrient loss from soils through leaching threatens global ecosystems by decreasing soil fertility and productivity and has a negative environmental impact [1, 2]. Plants play a major role in preventing leaching; thus, enhancement in efficiency of plant nutrient utilization is a measure to reduce the environmental impact of leaching [3, 4]. In addition, solubility and mobility of some toxic elements (e.g., Cd) are high, leading to migration of Cd in the polluted soils through rain [5, 6]. Thus, it was suggested that reduction of Cd migration in soil-plant systems is beneficial for environmental safety [7].
Soil biota was also reported to modulate the effects on nutrient loss from leaching [3]. Terrestrial plants have evolved mechanisms that optimize nutrient acquisition by expanding their nutrient interception areas, which is able to minimize nutrient loss after rainfall leaching [4]. Arbuscular mycorrhizal fungi (AMF) colonized plant root hairs expand hyphal networks into surrounding soils [8, 9]. AMF can change soil physical properties through forming macroaggregates that alter the surface traits [10, 11] and increase the capability of surface adsorption of soils [12]. Plants can also be directly benefited by acquiring P, K, Ca, Mg, Cu, Zn, NH4+ and NO3−via AMF [13,14,15,16,17,18]. Overall, AMF act to reduce mineral N and P losses that are linked to increases in efficiency of plant nutrient uptake and stabilization of plant-soil systems [19, 20].
AMF have been shown to promote plant growth and accumulate Cd in Cd-polluted soils by the following mechanisms: (1) increasing antioxidant enzyme activity [21]; (2) elevating plant photosynthesis [22]; (3) increasing mineral nutrient uptake of host plants particularly P [23] and (4) secreting glomalin-related soil proteins [24]. Rainfall also induces heavy metal leaching on the surface layer of contaminated soils [5]. Cd can be absorbed by AMF hyphae intertwined with soil surface particles [11]. Heavy rainfall also results in Cd leaching from the soil surface to bottom layers, while AMF have been demonstrated to reduce Cd leaching in maize-planted soils [7]. However, little is known about how rainfall intensity and AMF affect plant growth and the uptake as well as leaching of both nutrients and Cd at different growth stages.
In the current study, we examined the effects of AMF and leaching intensity on element uptakes, as well as dissolved nutrients and Cd ions leaching in soils planted with white clover (Trifolium repens L.). We hypothesized that (1) AMF would increase plant growth and nutrient uptake regardless of rainfall amount, and (2) AMF would reduce dissolved ion leaching especially under high intensity rainfall conditions. We designed soil column experiments to monitor the uptake and leaching dynamics in different mowing and leaching events.
Results
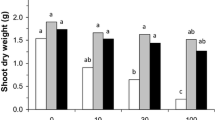
AMF-inoculated plants exhibited significantly higher shoot biomass than plants without inoculation at the mowing and harvest stage, regardless of water amount (Fig. 1). Similarly, N, P, Cu and Zn accumulated in the plant shoots followed a similar trend as plant shoot biomass (Fig. 2A, B, G and H). The root uptake of P and Cu was also improved by AMF (Fig. 2B and G). The shoot uptakes of K, Ca and Mg were increased by AMF only at the mowing stage but these element uptakes in shoots were not elevated by AMF under each leaching event conditions at harvest (Fig. 2C, D and E). The shoot Fe uptake at the mowing stage was only increased by AMF in the moderate water amount treatment (Fig. 2F).

Effects of arbuscular mycorrhizal fungi (AMF) and water amount on plant biomass. Different lower letters denote the significant differences in shoot and root biomass according to Duncan’s post hoc test. Shoot I and II designate plant materials collected at the mowing and harvest stages, respectively. Different upper letters denote the significant differences in total biomass according to Duncan’s post hoc test

Plant nutrient uptake in different arbuscular mycorrhizal fungi (AMF) and water amount treatments. Different lower letters denote the significant differences in shoot and root nutrient uptake according to Duncan’s post hoc test; Shoot I and II designate plant materials collected at the mowing and harvest stages, respectively. Different upper letters denote the significant differences in total nutrient uptake according to Duncan’s post hoc test. If no significant difference was found among treatments, no letter is indicated
Water treatment amount did not affect shoot biomass at either at mowing or harvest stage (Fig. 1). Moderate water amount significantly reduced root K, Mg, Zn and Cd uptakes in non-AMF plants, whereas only root K uptake was decreased by moderate water amount in AMF plants (Figs. 2 and 3). The total K and Zn uptakes for the AMF-colonized plants for the high water amount treatments were significantly lower than those of moderate water amount treatments (Fig. 2 C and H). L-AMF treatment exhibited maximum Cd uptake in the roots. Nevertheless, shoot Cd uptakes were increased by AMF at the mowing and harvest stages (Fig. 3).

Plant Cd uptakes in different arbuscular mycorrhizal fungi (AMF) and water amount treatments. Different lower letters denote the significant differences in shoot and root Cd uptakes according to Duncan’s post hoc test. Shoot I and II designate plant materials collected at the mowing and harvest stages, respectively. Different upper letters denote the significant differences in total Cd uptakes according to Duncan’s post hoc test
In the moderate and high water amount treatments, AMF resulted in a significant decrease of leachate volumes in the first leaching event. Although leachate volumes were not significantly different in the subsequent second, third and fourth leaching events for the AMF-colonized versus non-colonized plants, the sum of leachate volume of the four leaching events was significantly reduced in the presence of AMF when high water amounts were applied (Fig. 4A).

Leachate volume and amount of leached dissolved C and N in different arbuscular mycorrhizal fungi (AMF) and water amount treatments. Different lower letters denote the significant differences in each leaching event according to Duncan’s post hoc test. Different upper letters denote the significant differences in total leachate volume and amount of leached dissolved C and N according to Duncan’s post hoc test. If no significant difference was found among treatments, no letter is indicated
The amounts of NH4+ found in leachates for the three water amount treatments displayed different patterns. AMF had no influence on the amounts of leached NH4+ in the low water treatments for every leaching event. In contrast, AMF decreased the amounts of leached NH4+ only in the first leaching event when moderate water amount was applied. In both first and second leaching events, the amounts of leached NH4+ were reduced when high water amount was applied. Thus, for the moderate and high water treatments, the total amounts of leached NH4+ over the 4 leaching events of AMF inoculation treatments were less than these of non-AMF inoculation treatments (Fig. 4B).
The total amounts of leached NO3− differed from the trends for NH4+. In the low and moderate water treatments, the amounts of leached NO3− were not affected by AMF for all leaching events. Nevertheless, with high water amount inputs, the presence of AMF significantly increased the amount of leached NO3− in the first and fourth leaching events, as well as the total amount of leached NO3− over the 4 leaching events. In contrast, AMF significantly decreased the amount of leached NO3− in the third leaching event. The DOC in the leachates was only increased by water amount but not by AMF for all leaching events (Fig. 4D).
In this study, Fe, Cu and Zn concentrations in leachate were not detected by ICP-OES (data not shown). In the low water treatments, it was found that AMF increased the total amounts of leached P, Ca and Mg only in the third leaching event (Fig. 5A, C, D). In the moderate water treatments, the amounts of leached P, K, Ca and Mg were not significantly altered by AMF except for a reduction of P in the second leaching event (Fig. 5A-D). Compared with the treatments that were not colonized with AMF, the presence of AMF significantly decreased the amount of leached K in the third and fourth leaching events. Reduction of total amount of leached K and Cd in the high water treatment appeared in the AMF inoculation group (Fig. 5B).

Amount of leached element in different arbuscular mycorrhizal fungi (AMF) and water amount treatments. Different lower letters denote the significant differences in each leaching event according to Duncan’s post hoc test. Different upper letters denote the significant differences in total amount of leached element according to Duncan’s post hoc test. If no significant difference was found among treatments, no letter is indicated
It was found that soil DOC concentration in the M + AMF treatment was significantly higher than M-AMF treatment (Fig. 6C). The maximal soil DTPA-Cd concentration was found in the H-AMF treatment group (Fig. 6E). Soil pH was not significantly different for the groups with low and moderate water amount inputs. Compared with L-AMF treatment, pH values for H-AMF and H + AMF treatments decreased 0.1 and 0.36 units, respectively (Fig. 6D).

Soil chemical properties in different arbuscular mycorrhizal fungi (AMF) and water amount treatments. Different lower letters denote the significant differences according to Duncan’s post hoc test
Discussion
AMF has been reported to increase host plant biomass [22, 25, 26]. However, not every nutrient or mineral was equally promoted by the presence of AMF [16, 18, 27, 28]. Our results indicated that AMF increased shoot biomass, as well as N, P, Cu and Zn uptake at every growth stage, whereas shoot K, Ca and Mg uptakes were not increased by AMF at every growth stage when leaching intensity increased (Figs. 1 and 2). Therefore, we concluded that the nutrient uptake in response to AMF varied, dependent on the plant growth stage and the type of nutrients. In addition, it was speculated that the reduction of nutrient element uptake did not limit plant shoot growth regardless of water amounts applied. Increased shoot Cd uptake induced by increased biomass inAMF-inoculated plants has been previously reported [16, 18, 27, 28], which might explain the enhancement of Cd uptakes in AMF-inoculated plant shoot from our findings (Fig. 3).
A recent study found that soils containing plant roots had higher non-capillary porosity and saturated hydraulic conductivity than soils without plant roots, which facilitated preferential migration of soil solutions [29]. In this study, we did not observe increases in root biomass following AMF inoculation (Fig. 1), but significant reductions in leachate volumes occurred in the first leaching event when moderate and high water were applied (Fig. 4A). The reasons for this phenomenon may include (1) AMF-colonized plants could facilitate root growth at the early growth stages, (2) a higher water consumption in AMF-colonized plant shoots was accompanied by ahigher biomass, and (3) other unknown root morphological characteristics that were not yet identified in this experiment could also influence leaching.
These results were partly consistent with the hypothesis that AMF can reduce soil NH4+ loss, especially under conditions of increased rainfall intensity (Fig. 4B). Similar NH4+ losses that occurred via leaching have been reported in other studies [13, 20, 30]. AMF colonized plants can absorb N as amino acids [31], NO3− [26] or NH4+ [32]. Reduction in N losses via leaching has also been linked to the promotion of plant growth, N assimilation and NH4+ immobilization in soils involving AMF-root systems in pasture soils [13]. In addition, reduction of leachate volume also contributes to decreases of leached N [20, 33, 34]. In our study, increased plant N uptake and reduction of leachate volumes both contributed to the reductions of NH4+ loss through leaching (Figs. 2 A, 4A).
Previous studies have demonstrated that NH4+ is preferentially absorbed over NO3− in mycorrhizal plants [32, 35]. In particular, AMF exert their effects on N leaching for NH4+ but not for NO3− [13]. In contrast, neither AMF nor water volume influenced the amount of leached NH4+ whereas NO3− loss through leaching was less for AM-colonized plants as compared with non-colonized plants when water regime was regular [36]. In red clover mycorrhizal grassland microcosms, the amount of leached NO3− may also be associated with the N fixation by Rhizobia spp., resulting in N loss via leaching [4, 37]. The amounts of leached NO3− in our experiments were greater for AMF-colonized plants with high water treatments (Fig. 3 C). It was predicted that an enhancement of N fixation in the presence of AMF may influence the available N pool in soil and lead to variable levels of NO3− loss by leaching in our study. AMF can alter soil microbial communities including those involved in N-cycling processes, which could lead to N losses via denitrification as previously reported [4, 38, 39]. N leaching from plant-soil systems induced by rainfall events is influenced by numerous factors including soil characteristics, plant root systems, soil organisms and their interactions [1]. Therefore, the levels of N fixation, soil N-cycling microorganisms and nitrification processes that existed in our experimental plant-root systems may have influenced NO3− loss in different ways at different growth stages.
Plant roots and AMF hyphae both produce large amounts of exudates that are released into soils, including organic carbon derived from photosynthesis [10]. Root exudates such as the low-molecular weight malic, succinic and citric acids were enhanced by AMF-colonization of soils contaminated by heavy metals [40]. However, our results indicated that AMF did not influence the dynamics of DOC losses (Fig. 4D).
Generally, AMF promote P uptake in host plants [14] and reduce leached P in soils in normal rainfall events [13, 41]. However, we found that P uptake in plants was enhanced by AMF at each of water treatment groups (Fig. 2B), but the total P lost by leaching was unaltered by the presence of AMF regardless of water amount (Fig. 5A). Nevertheless, at low water amount treatments, the presence of AMF significantly increased the total amount of leached P, Ca and Mg in the third leaching event (Fig. 5A, C, D). The total amount of leached P, K, Ca and Mg increased with water amount increased (Fig. 5). All of these results indicated that nutrient leaching is dependent on plant growth stages, rainfall intensity, nutrient type, soil nutrient levels and instantaneous plant nutrient uptake status.
Plant root systems can directly take up nutrients and heavy metals, thereby lowering these ion concentrations in soils [42]. Maize roots could decrease concentration of Cd in the interflow while had no effect on leached Cd from the contaminated soils; whereas AMF could reduce concentration of Cd in the interflow and in the leachate [7]. The role of AMF played in Cd uptake could be explained by changes in soil macroaggregates and total glomalin-related soil protein (T-GRSP) content in Cd-contaminated soils [7, 43]. Additionally, soil macroaggregates possessing relatively higher mass Cd loading reduced Cd mobility and its export from contaminated soils [44]. However, the presence of AMF did not result in increase in total Cd uptake in white clover plants in our experiments (Fig. 3). Therefore, Cd uptake in plants may not be the primary reason for the reduction in the amount of leached Cd when high water treatment was applied. Although AMF reduced soil pH that might increase the risk of heavy metal mobilization, the soil available Cd was not affected by AMF in the high water treatment group (Fig. 6). We only measured the soil available Cd before plant harvest, thus, more information regarding the dynamics of soil available Cd is needed in the future to explain the reduction of Cd loss via leaching.
Conclusions
The increased shoot biomass, as well as uptake of N, P, Cu and Zn in AMF-colonized white clover plants partly support the hypothesis that AMF may increase plant growth and nutrient uptake regardless of rainfall amount. Reduced losses of soil NH4+, K and Cd induced by AMF occurred under high intensity rainfall conditions. Opposite to our assumption, AMF significantly reduced NH4+, K and Cd loss from soils but increased soil NO3− loss when high intensity leaching occurred. The changes of soil in different growth stages will help us to further clarify the mechanism of rainfall on plant element absorption and soil ion leaching in future studies.
Materials and methods
Substrate and microbial inoculum propagation
The location of soil collection was Baima Town of Nanjing, Jiangsu Province, China. The soils were mixed with sand (2:1) and then sterilized by at 120 °C in an autoclave. The chemical properties of mixtures used for this study were listed in Table S1. A mixture of AMF inocula containing Glomus aggregatum, G. etunicatum, G. intraradices, G. tortuosum and G. versiforme was used in this experiment. Cadmium chloride solution was added into soil and mixed well. Cd concentration of the mixtures was 3 mg Cd kg− 1. The mixture was maintained in the greenhouse for six months before use.
Experimental design
The experiments in this study included three water amount treatments (i.e., low (L), moderate (M) and high (H)) and two AMF treatments (i.e., plants lacking AMF (–AMF) and plants inoculated with (+ AMF)), resulting in 6 different treatments (i.e., L–AMF, L + AMF, M–AMF, M + AMF, H–AMF and H + AMF) and each treatment was conducted in replicates. The variety of white clover used in this experiment is haifa. White clover seeds were sown in pots (35 cm in height and 16 cm in diameter) filled with 6 kg of substrate soil on November 26, 2018. In the + AMF treatments, white clover was inoculated with 200 g of mycorrhizal fungi whereas 200 g sterilized inoculants used for AMF propagation were added in the –AMF treatments. To maintain microbial communities, each of all treatments received 50 mL soil microbial filtrate after screening through a nylon mesh (25 μm). Twenty-five days after sowing, the clover was thinned to 15 seedlings per pot. On April 7, 2019, the plants were cut to 10 cm, which was designated as the mowing event. Plants were harvested by hand approximately on May 13, 2019, and this was designated the harvest event. During the growth period, leaching events were carried out on March, 9, March 18, April 9 and April 19, 2019, respectively. In each leaching event, deionized water were applied in the low, moderate and high water amount treatments (i.e., 720, 1440 and 2160 mL), respectively. Drainage holes at the bottom of the pots were used to collect the leachate.
Measurements
Clover shoot biomass and elemental uptake were calculated as the sum of biomass or element uptake in the mowing and harvest events. The elemental uptake was calculated as concentrations in the plant tissues multiplied by the biomass. Total biomass was the sum of shoot and root biomass, and total element uptake was the sum of shoot and root element uptake. Total leachate volume and total amount of element loss was the sum of leachate volume or the amount of element loss from all four leaching events. Mycorrhizal colonization was determined as previously described [45].
Analytical measurements of P, K, Ca, Mg, Fe, Cu, Zn and Cd were conducted using inductively coupled plasma-optical emission spectrometry (ICP-OES). Clover samples were digested in a nitric:perchloric acid mixture (3:1 v/v) [27]. Total N in plant tissues was determined after digestion in H2SO4:H2O2 solution and its concentrations were determined by spectrophotometry as previously described [46]. Diethylenetriamine pentaacetic acid (DTPA) was used to extract Cd from soils as previously described [47]. NH4+ was extracted using 0.5 M K2SO4 and measured using the indophenol blue method. Dissolved organic carbon (DOC) levels in soils were quantified with a TOC-TN analyzer as previously described [48]. NH4+, DOC and elements in leachates were also determined as described above. NO3− was measured using a dual wavelength method as previously described [48]. Soil pH was determined using a pH meter in the supernatant of a soil:deionized water mixture (in 1:5 ratio). Available P in soils was measured by the molybdenum blue method as previously described [49].
Statistical analyses
Statistical analyses were conducted using SPSS version 13.0 (IBM, Chicago, Ill, USA). Duncan multiple comparisons were used to compare differences among different treatments.
Availability of data and materials
The data will be available on request by contacting corresponding author.
Abbreviations
- AMF:
-
arbuscular mycorrhizal fungi
- N:
-
nitrogen
- P:
-
phosphorus
- K:
-
potassium
- Ca:
-
calcium
- Mg:
-
magnesium
- Fe:
-
Ferrum
- Cu:
-
copper
- Zn:
-
Zinc
- Cd:
-
cadmium
- DOC:
-
dissolved organic carbon
- DTPA:
-
diethylenetriamine pentaacetic acid
References
Cameron KC, Di HJ, Moir JL. Nitrogen losses from the soil/plant system: a review. Ann Appl Biol. 2013;162:145–173.
Galloway JN, Aber JD, Erisman JW, Seitzinger SP, Howarth RW, Cowling EB, Cosby BJ. The nitrogen cascade. Bioscience. 2003;53:341–356.
Bender SF, van der Heijden MGA. Soil biota enhance agricultural sustainability by improving crop yield, nutrient uptake and reducing nitrogen leaching losses. J Appl Ecol. 2015;52:228–239.
Cavagnaro TR, Bender SF, Asghari HR, van der Heijden MGA. The role of arbuscular mycorrhizas in reducing soil nutrient loss. Trends Plant Sci. 2015;20:283–290.
Beesley L, Moreno-Jimenez E, Clemente R, Lepp N, Dickinson N. Mobility of arsenic, cadmium and zinc in a multi-element contaminated soil profile assessed by in-situ soil pore water sampling, column leaching and sequential extraction. Environ Pollut. 2010;158:155–160.
Zheng SA, Zhou W, Xue YH, Yin JF, Zheng XQ. Effect of simulated rain on leaching of cadmium from 22 typical soils and key factors identification. Environ Chem. 2013;37:3489–3496.
He Y-m, Yang R, Lei G, Li B, Jiang M, Yan K, Zu Y-q, Zhan F-d, Li Y. Arbuscular mycorrhizal fungi reduce cadmium leaching from polluted soils under simulated heavy rainfall. Environ Pollut. 2020;263:114406.
Simard SW, Beiler KJ, Bingham MA, Deslippe JR, Philip LJ, Teste FP. Mycorrhizal networks: Mechanisms, ecology and modelling. Fungal Biology Reviews. 2012;26:39–60.
Smith SE, Read DJ. Mycorrhizal Symbiosis. London: Academic Press; 2008.
Frey SD. Mycorrhizal fungi as mediators of soil organic matter dynamics. In: DJ, Futuyma, editors. Annual Review of Ecology, Evolution, and Systematics. Vol. 50. Palo Alto: Annual Reviews; 2019.
Janouskova M, Pavlikova D. Cadmium immobilization in the rhizosphere of arbuscular mycorrhizal plants by the fungal extraradical mycelium. Plant Soil. 2010;332:511–520.
Wang Q, Mei DG, Chen JY, Lin YS, Liu JC, Lu HL, Yan CL. Sequestration of heavy metal by glomalin-related soil protein: Implication for water quality improvement in mangrove wetlands. Water Res. 2019;148:142–152.
Bender SF, Conen F, Van der Heijden MGA. Mycorrhizal effects on nutrient cycling, nutrient leaching and N2O production in experimental grassland. Soil Biol Biochem. 2015;80:283–292.
Johansen A, Jakobsen I, Jensen ES. Hyphal transport by a vesicular-arbuscular mycorrhizal fungus of N applied to the soil as ammonium or nitrate. Biology Fertil Soils. 1993;16:66–70.
Lehmann A, Veresoglou SD, Leifheit EF, Rillig MC. Arbuscular mycorrhizal influence on zinc nutrition in crop plants – A meta-analysis. Soil Biol Biochem. 2014;69:123–131.
Liu M, Che Y, Wang L, Zhao Z, Zhang Y, Wei L, Xiao Y. Rice straw biochar and phosphorus inputs have more positive effects on the yield and nutrient uptake of Lolium multiflorum than arbuscular mycorrhizal fungi in acidic Cd-contaminated soils. Chemosphere. 2019;235:32–39.
Marschner H, Dell B. Nutrient-uptake in mycorrhizal symbiosis. Plant Soil. 1994;159:89–102.
Zhang F, Liu M, Li Y, Che Y, Xiao Y. Effects of arbuscular mycorrhizal fungi, biochar and cadmium on the yield and element uptake of Medicago sativa. Sci Total Environ. 2019;655:1150–1158.
Mardhiah U, Caruso T, Gurnell A, Rillig MC. Arbuscular mycorrhizal fungal hyphae reduce soil erosion by surface water flow in a greenhouse experiment. Appl Soil Ecol. 2016;99:137–140.
van der Heijden MGA. Mycorrhizal fungi reduce nutrient loss from model grassland ecosystems. Ecology. 2010;91:1163–1171.
Rozpadek P, Wezowicz K, Stojakowska A, Malarz J, Surowka E, Sobczyk L, Anielska A, Wazny R, Miszalski Z, Turnau K. Mycorrhizal fungi modulate phytochemical production and antioxidant activity of Cichorium intybus L. (Asteraceae) under metal toxicity. Chemosphere. 2014;112:217–224.
Gao TT, Liu XM, Shan L, Wu Q, Liu Y, Zhang ZJ, Ma FW, Li C. Dopamine and arbuscular mycorrhizal fungi act synergistically to promote apple growth under salt stress. Environ Exp Bot. 2020;178:11.
Liu M, Zhao Z, Chen L, Wang L, Ji L, Xiao Y. Influences of arbuscular mycorrhizae, phosphorus fertiliser and biochar on alfalfa growth, nutrient status and cadmium uptake. Ecotoxicol Environ Safety. 2020;196:110537.
Qiao YH, Crowley D, Wang K, Zhang HQ, Li HF. Effects of biochar and Arbuscular mycorrhizae on bioavailability of potentially toxic elements in an aged contaminated soil. Environ Pollut. 2015;206:636–643.
Cao J, Wang C, Ji D. Improvement of the soil nitrogen content and maize growth by earthworms and arbuscular mycorrhizal fungi in soils polluted by oxytetracycline. Sci Total Environ. 2016;571:926–934.
Cavagnaro TR, Barrios-Masias FH, Jackson LE. Arbuscular mycorrhizas and their role in plant growth, nitrogen interception and soil gas efflux in an organic production system. Plant Soil. 2012;353:181–194.
Liu M, Sun J, Li Y, Xiao Y. Nitrogen fertilizer enhances growth and nutrient uptake of Medicago sativa inoculated with Glomus tortuosum grown in Cd-contaminated acidic soil. Chemosphere. 2017;167:204–211.
Xiao Y, Liu M, Chen L, Ji L, Zhao Z, Wang L, Wei L, Zhang Y. Growth and elemental uptake of Trifolium repens in response to biochar addition, arbuscular mycorrhizal fungi and phosphorus fertilizer applications in low-Cd-polluted soils. Environ Pollut. 2020;260:113761.
Jiang XJ, Liu W, Chen C, Liu J, Yuan Z-Q, Jin B, Yu X. Effects of three morphometric features of roots on soil water flow behavior in three sites in China. Geoderma. 2018;320:161–171.
Asghari HR, Cavagnaro TR. Arbuscular mycorrhizas enhance plant interception of leached nutrients. Funct Plant Biol. 2011;38:219–226.
Whiteside MD, Treseder KK, Atsatt PR. The brighter side of soils: Quantum dots track organic nitrogen through fungi and plants. Ecology. 2009;90:100–108.
Tanaka Y, Yano K. Nitrogen delivery to maize via mycorrhizal hyphae depends on the form of N supplied. Plant Cell Environ. 2005;28:1247–1254.
Asghari HR, Chittleborough DJ, Smith FA, Smith SE. Influence of arbuscular mycorrhizal (AM) symbiosis on phosphorus leaching through soil cores. Plant Soil. 2005;275:181–193.
Corkidi L, Merhaut DJ, Allen EB, Downer J, Bohn J, Evans M. Effects of mycorrhizal colonization on nitrogen and phosphorus leaching from nursery containers. HortScience. 2011;46:1472–1479.
Govindarajulu M, Pfeffer PE, Jin HR, Abubaker J, Douds DD, Allen JW, Bucking H, Lammers PJ, Shachar-Hill Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature. 2005;435:819–823.
Bowles TM, Jackson LE, Cavagnaro TR. Mycorrhizal fungi enhance plant nutrient acquisition and modulate nitrogen loss with variable water regimes. Global Chang Biol. 2018;24:e171–82.
Wagg C, Bender SF, Widmer F, van der Heijden MGA. Soil biodiversity and soil community composition determine ecosystem multifunctionality. PNAS. 2014;111:5266–5270.
Bender SF, Plantenga F, Neftel A, Jocher M, Oberholzer H-R, Koehl L, Giles M, Daniell TJ, van der Heijden MGA. Symbiotic relationships between soil fungi and plants reduce N2O emissions from soil. Isme J. 2014;8:1336–1345.
Veresoglou SD, Chen B, Rillig MC. Arbuscular mycorrhiza and soil nitrogen cycling. Soil Biol Biochem. 2012;46:53–62.
Ren CG, Kong CC, Wang SX, Xie ZH. Enhanced phytoremediation of uranium-contaminated soils by arbuscular mycorrhiza and rhizobium. Chemosphere. 2019;217:773–779.
Martinez-Garcia LB, De Deyn GB, Pugnaire FI, Kothamasi D, van der Heijden MGA. Symbiotic soil fungi enhance ecosystem resilience to climate change. Global Change Biol. 2017;23:5228–5236.
Kim KR, Owens G, Kwon SI. Influence of Indian mustard (Brassica juncea) on rhizosphere soil solution chemistry in long-term contaminated soils: A rhizobox study. J Environ Sci. 2010;22:98–105.
Zhang ZF, Mallik A, Zhang JC, Huang YQ, Zhou LW. Effects of arbuscular mycorrhizal fungi on inoculated seedling growth and rhizosphere soil aggregates. Soil Tillage Res. 2019;194:7.
Li SS, Wang M, Zhao ZQ, Li XY, Han Y, Chen SB. Alleviation of cadmium phytotoxicity to wheat is associated with Cd re-distribution in soil aggregates as affected by amendments. RSC Adv. 2018;8:17426–17434.
Giovannetti M, Mosse B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980;84:489–500.
Yuen SH, Pollard AG. Determination of nitrogen in agricultural materials by the Nessler reagent. 2. Micro-determinations in plant tissues and in soil extracts. J Sci Food Agric. 2010;5:364–369.
Xiao Y, Wang L, Zhao Z, Che Y. Biochar shifts biomass and element allocation of legume-grass mixtures in Cd-contaminated soils. Environ Sci Pollut Res. 2020;27:10835–10845.
Xiao Y, Li Y, Che Y, Deng S, Liu M. Effects of biochar and nitrogen addition on nutrient and Cd uptake of Cichorium intybus grown in acidic soil. Inter J Phytoremed. 2018;20:398–404.
Olsen SR, Sommers LE. Phosphorus. In: Page AL, editor. Methods of soil analysis: part 2 chemical and microbiological properties. Madison: American Society of Agronomy, Soil Science Society of America; 1982. pp. 403–430.
Acknowledgements
Not applicable.
Funding
The work was supported by the National Natural Science Foundation of China (No. 31971745, 32171688), the China Agriculture Research System of MOF and MARA and Jiangsu Qinglan Project.
Author information
Authors and Affiliations
Contributions
YX designed, performed this experiment and wrote this paper, LC revised this paper. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Xiao, Y., Chen, L. Arbuscular mycorrhizal fungi reduce potassium, cadmium and ammonium losses but increases nitrate loss under high intensity leaching events. BMC Plant Biol 22, 365 (2022). https://doi.org/10.1186/s12870-022-03741-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-022-03741-3