
Abstract
Background
Plant defense against herbivores begins with perception. The earlier plant detects the harm, the greater plant will benefit in its arm race with the herbivore. Before feeding, the larvae of the rice pest Cnaphalocrocis medinalis, initially spin silk and fold up a leaf. Rice can detect and protect itself against C. medinalis feeding. However, whether rice could perceive C. medinalis leaf rolling behavior is currently unknown. Here, we evaluated the role of leaf rolling by C. medinalis and artificial leaf rolling in rice plant defense and its indirect effect on two important C. medinalis parasitoids (Itoplectis naranyae and Apanteles sp.) through a combination of volatile profiling, gene-transcriptional and phytohormonal profiling.
Results
Natural leaf rolling by C. medinalis resulted in an increased attraction of I. naranyae when compared to the undamaged plant after 12 h. Volatile analysis revealed that six out of a total 22 components significantly increased in the headspace of C. medinalis rolled plant when compared to undamaged plant. Principal component analysis of these components revealed similarities in the headspace of undamaged plant and artificially rolled plant while the headspace volatiles of C. medinalis rolled plant deferred significantly. Leaf rolling and feeding by C. medinalis up-regulated the plant transcriptome and a series of jasmonic acid (JA) and salicylic acid (SA) related genes. While feeding significantly increased JA level after 12 to 36 h, rolling significantly increased SA level after 2 to 12 h. Compared to artificial rolling, natural rolling significantly increased JA level after 36 h and SA level after 2 and 12 h.
Conclusions
Our findings suggest that natural leaf rolling by C. medinalis can be perceived by rice plant. The detection of this behavior may serve as an early warning signal in favor of the rice plant defenses against C. medinalis.
Similar content being viewed by others


Background
Throughout their life cycle, plants experience biotic and abiotic stresses, which can severely affect their growth and yield [1,2,3]. Globally, herbivore insects are among the most serious biotic problems and have close relationships with plants [4]. While insects have evolved ways to find hosts, plants are under selection pressure to evade detection or defend themselves when attacked [4,5,6].
To fend off insect herbivores, plants have evolved intricate and dynamic defense systems. Phytohormones play an important role in regulating plant defenses [7,8,9]. The phytohormones, jasmonic acid (JA) and salicylic acid (SA), and their derivatives play predominant roles in signal transduction of plant defenses against pathogens and herbivorous insects [10, 11]. Plant hormones can mediate downstream regulation of plant defenses, including direct defense mechanisms such as the production of defensive proteins and enzymes, and indirect defense mechanisms involving the production of volatile blends [12,13,14].
Plant volatiles are released naturally, and their emission can change in response to different stress factors [15, 16]. With plant–insect co-evolution, herbivore-induced plant volatiles (HIPVs) have become cost-saving weapons for plant defenses, and are useful in attracting natural enemies of herbivores [17,18,19]. Host recognition by natural enemies is specific, as they identify volatile blends from various plant species. However, natural enemies differ in their ability to distinguish HIPVs source depending on the distance of the natural enemy to the cues [20,21,22].
Conflicts in interactions of plants with herbivores begin with perception. Interestingly, plants can identify attacks from different insect species and mount several defenses [23, 24]. Plants perceive insect herbivores’ activities like oviposition, feeding, walking on the leaf surface, as well as chemical cues from insect oral secretion and frass [25,26,27]. Spinning is an important activity of silk-producing insects. The ability to produce silk has evolved in many groups of insects, and is used to accomplish a wide array of activities that enhance survival [28,29,30]. Several insects’ silk has been reported to attract parasitic wasps. For instance, Apanteles melanoscelus (Ratzeburg) (Hymenoptera: Braconidae) has a positive response to gypsy moth silk, and silk extracts of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) larvae also attract parasitic wasps [31, 32]. However, no research has shown whether rolling behaviors following spinning can mediate plant defenses.
In Asia, the rice leaf folder, Cnaphalocrocis medinalis Guenée (Lepidoptera: Pyralidae), is one of the most important insect pest of paddy rice [33]. Severe feeding by this pest often affects the growth of rice plant leading to yield loss [34, 35]. Various wasp parasitoids of C. medinalis commonly exist in rice fields, such as Itoplectis naranyae (Ashmead) (Hymenoptera: Ichneumonidae) and Apanteles sp. (Hymenoptera: Braconidae) [36, 37]. The characteristic behavior of C. medinalis larvae is to roll a leaf longitudinally, by spinning silk before feeding [38, 39]. There are reports indicating that the feeding behavior of C. medinalis can induce rice defenses, but there is no evidence to indicate whether leaf rolling alone prior to feeding by the pest, can be perceived by the plant [40, 41].
Early detection of herbivores is beneficial for plants to develop effective and sustainable defenses. It is therefore important to determine whether rice plant can detect the early threat of C. medinalis leaf rolling, by initiating defenses against the pest [42]. In this study, we investigated the dynamics of rice plant responses to C. medinalis leaf rolling, C. medinalis leaf rolling and feeding and artificial leaf rolling, and compared with undamaged plant. To achieve this goal, we integrated results from wasps’ behavioral assays, plant volatile profiles, plant transcriptional and phytohormonal profiles.
Results
Behavior of natural enemies
We hypothesized that leaf rolling behavior of C. medinalis larvae can trigger rice plant defenses. Therefore, we tested the preferences of two parasitic wasps (I. naranyae and Apanteles sp.) associated with C. medinalis larva. Itoplectis naranyae significantly preferred natural rolling treatment, when compared to undamaged plant after 12 h (x2 = 6.3684, df = 1, P = 0.0116) (Fig. 1a). However, both wasp species showed no significant preferences for naturally rolled plant after 24 h induction, when compared to undamaged plant (Fig. 1b). To explore whether sole rolling of plant leaf could be detected by natural enemies, we also tested the behavior choices of wasps between undamaged plant and artificially rolled plant. However, no differences in preferences for both wasp species between undamaged plant and artificially rolled plant were observed after 12 and 24 h (Fig. 1c and d).

Olfactometer responses of Itoplectis naranyae and Apanteles sp. to odors f from artificial rolling plant and natural rolling plant treatments after 12 h (a, c) and 24 h (b, d) versus undamaged plant. Each wasp was released one at a time in the main arm of the Y-tube and given 5 min to make a choice between the test or control odor. Means with asterisk are significantly different (P < 0.05)
Analysis of rice plant volatiles
To explore the reason for natural enemy behavior, we analyzed plant volatiles in undamaged plant, artificially rolled plant, and naturally rolled plant. In total, 22 volatile components were identified, including several terpenes, aldehydes, ketones, and esters (Table 1). Generally, these components occurred at lower amounts in the headspace of undamaged plant and artificially rolled plant, and at higher amounts in the headspace of naturally rolled plant. Of the 22 components identified, six were significantly higher after 12 and 24 h in the headspace of naturally rolled plant (P < 0.05). Principal Component Analysis also showed that the headspace volatiles of undamaged plant and artificially rolled plant were similar, but the headspace volatiles of naturally rolled plant differed significantly from both undamaged plant and artificially rolled plant (P < 0.05), especially after 12 h of damage (Fig. 2a and b).

Principal Component Analysis comparing volatile blends from undamaged plant, artificial rolling and natural rolling plant treatments. a after 12 h attack and (b) after 24 h attack. Grouping pattern of samples were with respect to the first two principal components and Hotelling’s ellipse at 95% confidence interval for the observations. Each point represents one sample
Global transcriptome changes in rice leaves in response to Cnaphalocrocis medinalis infestation
We used high-throughput sequencing to elucidate global transcript abundance in rice, and then compared the expression profiles of artificial rolling, natural rolling and rolling and feeding plant against undamaged plant. Compared to the undamaged plant, rolling and feeding treatment induced a drastic change in rice plant transcriptional levels while artificial rolling induced slight changes (Fig. 3). Compared to undamaged plant, 1073 genes were up-regulated and 870 genes were down-regulated in natural rolling plant while 2899 and 1807 genes were up-regulated and down-regulated in rolling and feeding plant, respectively (Table 2). The KEGG enrichment analyses showed that many defense- or stress-related pathways were activated, including plant hormone pathways (see Additional file 1: Figure S1). Moreover, the transcriptional responses of rice plant’s JA and SA pathways associated with the different treatments used in this study were identified using the GO tool (Fig. 4). Among these, genes such as lipoxygenase (LOX), hydroperoxide dehydratase (AOS), allene oxide cyclase (AOC), 12-oxophytodienoic acid reductase (OPR) and their homologs associated with the JA pathways were up-regulated, especially following the rolling and feeding treatment (Fig. 4a). For the SA pathway, TGA transcription factors (TGA), pathogenesis-related (PR) protein and regulatory protein NPR1 (NPR1) genes were up-regulated in natural rolling treatment while phenylalanine ammonia-lyase (PAL), TGA and NPR1 genes were up-regulated in rolling and feeding treatment when compared to the undamaged treatment (Fig. 4b).

Comparative analysis of changes in the rice leaf transcriptome in response to artificial rolling, natural rolling and rolling and feeding by Cnaphalocrocis medinalis after 12 h. Genes associated with a q ≤ 0.05 for at least one time point were used to construct the Venn diagram

Comparative analysis of gene transcriptomes associated with jasmonic acid and salicylic acid pathways. a gene transcriptional differences related to jasmonic acid pathway and (b) gene transcriptional differences related to salicylic acid pathway. Note: enoyl-CoA hydratase/3-hydroxyacyl-CoA dehydrogenase (MFP2); 12-oxophytodienoic acid reductase (OPR); lipoxygenase (LOX); hydroperoxide dehydratase (AOS); acetyl-CoA acyltransferase 1 (ACAA1); allene oxide cyclase (AOC); transcription factor MYC2 (MYC2); acyl-CoA oxidase (ACX); TGA transcription factors (TGA); phenylalanine ammonia-lyase (PAL); pathogenesis-related (PR) protein; regulatory protein NPR1 (NPR1)
Jasmonic acid and salicylic acid analyses
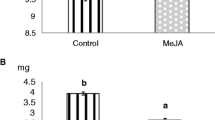
Based on transcript profile changes associated with hormone signaling, we measured JA and SA levels in natural rolling, rolling and feeding and artificial rolling treatments and compared to the undamaged plant by using LC/MS. Generally, similar and increasing trends in JA and SA biosynthesis levels were observed in both artificial rolling treatment and natural rolling treatment at all tested time points (Fig. 5). The accumulation of JA was greater in rolling and feeding treatment than other treatments from 12 h. Jasmonic acid level in natural rolling treatment showed no significant difference from 0 h to 24 h but was significantly higher when compared to its level in artificially rolled plant (P < 0.01, Fig. 5a). Salicylic acid level was significantly higher in natural rolling treatment after 2 h and 12 h than in other treatments (P < 0.01). However, rolling and feeding produced significantly higher levels of SA after 24 h and 36 h when compared to other treatments (P < 0.01, Fig. 5b).

Plant phytohormones produced after natural rolling and rolling and feeding by Cnaphalocrocis medinalis, and artificial rolling of rice leaf at different time points. a jasmonic acid biosynthesis and (b) salicylic acid biosynthesis. Mean ± SEM. Means followed by different letter indicates significant differences between the treatments at specific time points (P < 0.01)
Discussion
The relationship between plants and insects has received much attention in recent years, with most studies focusing on plant resistance to herbivorous insects [43, 44]. Plant–insect co-evolution is essential for species diversity and survival [5]. For plants to develop effective and sustainable defenses against aggressors, early detection of the aggressors is essential [45]. In our study, rice leaf rolling by C. medinalis resulted in increased plant JA and SA levels, and in the release of plant volatile blends which helped the plant to attract natural enemies of the herbivore (Fig. 6).

Rice plant can perceive the leaf rolling behavior of Cnaphalocrocis medinalis, release blends of volatiles (terpenes, aldehydes, ketones, esters) and recruit the natural enemy of the pest
Parasitoids often use different cues to locate their host [22]. In our study, we observed that I. naranyae significantly preferred naturally rolled plant over undamaged plant after 12 h of pest infestation. However, both wasp species showed no significant preferences for naturally rolled plant after 24 h induction when compared to undamaged plant. Similarly, both wasp species neither preferred undamaged nor artificially rolled plant after 12 and 24 h. There are no reports about the host cues employed by these two wasps in locating their target host. Thus, further studies should be carried out to unravel the cues involved in the wasps’ host location strategies.
Plant respond to stress in a series of steps after the stress is perceived [46, 47]. In general, wounding can induce direct and indirect plant defenses, such as changes in plant volatiles and attracting natural enemies of the attacker [48, 49]. Our results showed that natural rolling of rice leaf by C. medinalis significantly increased six volatile components while artificial rolling significantly increased only one volatile component after 12 h of induction. Principal component analysis showed that the headspace volatiles of natural rolling treated plant significantly differed, especially after 12 h of damage. We also found that the strong responses of wasp species to odors emanating from plants challenged by C. medinalis, was likely due to HIPVs, which modified their volatile organic compound (VOC) profiles when compared to undamaged plants.
Analysis of rice plant phytohormones showed significant differences in JA and SA levels in naturally rolled plant when compared to artificially rolled plant: for SA after 2 h to 24 h, and for JA after 36 h. It seems that leaf rolling behavior had a superimposed effect on plant immunity, when compared to artificial rolling. Therefore, our results suggest a correlation of plant defenses linked to C. medinalis leaf rolling behavior. The proteomics of Bombyx mori Linnaeus (Lepidoptera: Bombycidae), a silk-producing insect, revealed that many enzymes, protease inhibitors, and other unknown proteins are involved in plant defenses against its silk production. Among these, 40 protease inhibitors were identified as being involved in regulating or protecting the host plant from infestation [50]. However, the silk components from C. medinalis, which might have triggered plant responses, were not determined in the current study. Future studies should unravel these specific components and active ingredients involved in C. medinalis silk production and concomitant plant defenses. Additionally, treatments involving the application of silk only and application of silk and artificial rolling with silk maybe needed to assess whether similar defense responses of the plant will be recovered as reported in this study.
Our study also investigated plant defenses against feeding behaviors in order to explore differences in rice defense between rolling and feeding behaviors at the transcriptomic and metabolic level. Plant transcriptomic analysis showed an up-regulation of JA-related genes in the rolling and feeding treatment (Fig. 4a). Some SA-related genes were also up-regulated in the natural rolling and rolling and feeding treatments. Our result indicated that both natural rolling and rolling and feeding treatments could be perceived by rice plant, but the plant defense mechanisms differed in hormone levels. Jasmonic acid is famous for plant defense against insects’ attack and wounding. Clearly, feeding greatly increase the amount of JA when compared to JA levels in other treatments. Salicylic acid also plays an important role in early defense against herbivore insects. For instance, both Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae) and Pieris rapae (Linnaeus) (Lepidoptera: Pieridae) infestation raised SA level in cotton and arabidopsis, respectively [14, 51]. We recommend further studies to determine whether JA–SA synergism could occur at the level of gene expression in C. medinalis-challenged MH63 rice plant. Further studies are also needed to determine the mechanisms, the elicitors, and the coupled gas chromatography/electroantennogram detection active components involved in this tritrophic system.
Conclusions
Our study showed that similar to other tritrophic systems, HIPVs and the phytohormones JA and SA mediated rice-C. medinalis-I. naranyae tritrophic interactions [22, 52]. We provide the first evidence that rolling of rice leaves by C. medinalis induces JA and SA signaling pathways and HIPVs emission as plant defenses. In addition, our study showed differences in rice defenses among artificial rolling, natural rolling and rolling and feeding treatments. This could be due to the presence of different elicitors in rice’s ability to detect and defend itself against its aggressors. Our findings also provide a new beginning in exploring the effects of leaf rolling for silk-producing insect on associated plants and natural enemies.
Methods
Plants and insects
In this study, we used the rice (Oryza sativa L.) indica variety Minghui 63 (MH63), a restorer line with genetic stability. Single pre-germinated seeds of MH63 were sown in plastic pots (10 cm diameter × 8 cm in height), filled with a mixture of 0.5 g compound fertilizer (N:P:K = 14%:16%:15%, respectively) in a greenhouse at Huazhong Agricultural University, China. Plants were grown under natural light conditions at 28 ± 4 °C, 75 ± 5% relative humidity, and photoperiod ratios of 12/12 h, light/dark. The plants were watered daily and used at the tillering stage for both rearing and experimenting.
The C. medinalis colony was initiated from naturally occurring individuals, collected from rice fields at Xiaogan (113.91°E; 31.92°N), China. The insects were maintained in the laboratory on rice at 26 ± 2 °C, 75 ± 5% relative humidity, and photoperiod ratios of 16/8 h, light/dark. The fifth instar stage of the insect pest was used for all the experiments.
Adults of I. naranyae and Apanteles sp. were collected from paddy rice fields in Xiaogan, China and used in the Y-tube olfactomer experiment. Prior to the experiments, adults of each parasitoid species were fed on 10% honey solution until mating. After mating, parasitoids were transferred to the behavioral laboratory, at least 30 min prior to bioassay, to allow them to acclimatize to the experimental conditions.
Plant treatments
Flag leaf of rice plant was subjected to the following treatments: (i) undamaged (healthy rice plant); (ii) artificial rolling; (iii) natural rolling (leaf rolling without feeding by C. medinalis); and (iv) rolling and feeding (C. medinalis rolled and fed leaf). For artificial rolling treatment, we used a white thin thread made of inorganic materials to tie around the rice leaf to mimic C. medinalis silk spinning and leaf rolling behavior. For natural rolling treatment, insects were carefully observed and removed from the leaf immediately after rolling was completed, to prevent feeding on the leaves. Individual plants were induced by artificial rolling and natural rolling for 12 or 24 h before olfactometer assays and volatile analysis. Plants were induced by artificial rolling, natural rolling and rolling and feeding for 12 h before transcriptomic analysis and induced by artificial rolling, natural rolling and rolling and feeding for 2, 12, 24, or 36 h before phytohormonal analysis. Healthy undamaged plants acted as the control.
Behavioral response of parasitoid species to Cnaphalocrocis medinalis induced host plant volatiles
Responses of I. naranyae and Apanteles sp. females to different plant odor sources, in the absence of any visual cues, were tested using a Y-tube olfactometer (arm length: 18 cm and internal diameter: 1.5 cm). One arm of the tube was connected to the odor source and the other arm to the control treatment. A 60 W incandescent lamp bulb provided illumination. Charcoal-filtered clean air was passed through Teflon tubes into each arm of the olfactometer at a flow rate of 200 mL min− 1, and pulled out of the main arm of the olfactometer at the same rate, by a battery-powered portable vacuum pump (Sensen, Zhejiang, China).
Individual female parasitoid species were released one at a time in the main arm of the Y-tube. The pump system was turned on to test preferences for either the test or control odor, and each female was given 5 min to make a choice. The position of the test and the control arms of the Y-tube were changed after every three tested parasitoids to avoid positional bias. The connecting Teflon tubes and plant treatments were routinely replaced after six trials. A minimum number of 15 individual female parasitoid species acting as replicates were used for each pair of tests and control odor sources. The same environmental conditions, as described above, were maintained in the bioassay room.
Collection and analysis of plant volatiles
We used a closed-loop dynamic headspace volatile collection system to collect volatiles for 8 h, as described by Sun et al. [53]. Purified air was pushed into a glass jar containing each plant treatment at 500 mL min− 1 and drawn from the jar through vacuum containing traps, packed with Super Q adsorbent (200 mg each; Alltech Associates Inc. Deerfield, IL, USA). The air purification system consisted of (1) charcoal (activated carbon, 6 to 14 mesh, Fisher Scientific), (2) 5A molecular sieves (beads, 8 to 12 mesh, Sigma-Fluka), and (3) silica gel Rubin (drying agent free of metal salts, silica gel, Sigma-Fluka). The collection was replicated four times for each odor source.
After 8 h of volatile trapping using Super Q, each filter was eluted with 1 mL of N-hexane acetate, spiked with 10 μL of internal standard (0.1 mg/mL nonyl acetate, Sigma), and stored at − 40 °C until chemical analysis. Volatile extracts were analyzed by coupled gas chromatography/mass spectrometry (GC/MS) (QP-2010, Shimadzu, Shiga, Japan), equipped with an HP-5 MS fused-silica column (30 m × 0.25 mm × 0.25 μm) (Agilent Technologies, USA). Helium (1 mL min− 1) was used as the carrier gas, and an initial oven temperature of 40 °C was maintained for 1 min, ramped at 8 °C min− 1 to 300 °C, and held for 5 min. Compounds were identified using the quasi-molecular ions. In addition, structural assignments of several compounds were confirmed using authentic standards on the GC/MS under the same conditions employed for crude volatile analysis. Quantification was based on calibration curves (peak areas) generated from authentic standards of identified compounds. The peak area of each component was compared to the percent relative amount of internal standard peak area.
RNA isolation, library construction and sequencing
Total RNA extractions of plant leaves were performed using Trizol Reagent (Invitrogen Life Technologies). RNA purity and quantity were determined using a Nanodrop ND 1000 instrument (Nanodrop Technologies, Wilmington, DE, USA). Sequencing libraries were generated using the TruSeq RNA Sample Preparation Kit (Illumina, San Diego, CA, USA). Each treatment included three biological replicates.
Bioinformatics analysis
Gene expression patterns were clustered using Cutadapt (v1.15) software to filter the sequencing data and to generate high quality sequences (clean data) for further analysis. Genes were mapped to ‘terms’ in the Gene Ontology (GO) database and the number of differentially enriched genes were calculated for each term. DESeq (1.30.0) was used to analyze differential gene expression with screening conditions as follows: expression difference multiple |log2FoldChange| > 1 and significance of P < 0.05. Then, terms with significant enrichment of differentially enriched genes were calculated by hypergeometric distribution. GO enrichment analysis was used to obtain GO functional terms with significant enrichment of differentially expressed genes and revealed the possible functions. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis was used to conduct the enrichment analysis of differentially expressed genes. We counted the number of differentially expressed genes at different levels of KEGG pathway, and determined the metabolic pathways and signaling pathways that the differentially expressed genes mainly participated in. For each treatment, three biological replicates were used from individual plants.
Analysis of jasmonic acid and salicylic acid
Rice leaves (approximately 50 mg fresh weight) were collected, immediately frozen in liquid nitrogen, and stored at − 80 °C until analysis. Analysis was performed as described previously [55]. After grinding in liquid nitrogen, the 50 mg of each sample was extracted in 500 μL extraction solvent (2-propanol/H2O/concentrated HCl, v/v/v) in a 2:1:0.002 ratio and spiked with 10 μL internal standards (5 μg/mL Dihydrojasmonic acid and 5 μg/mL D4-Salicylic acid; Sigma). Samples were homogenized in a thermostatic mixer (MTC-100, Guangzhou, China) for 30 min at 100 rpm and at 4 °C. Then, 1 mL dichloromethane was added to the samples and homogenized under the same conditions. Samples were centrifuged at 13,000 g and at 4 °C for 5 min, and the bottom aqueous phase collected into clean 2 mL Agilent bottles. Samples were then dried completely under a hooded chamber using nitrogen evaporator and dissolved by adding 100 μl of methanol. A 0.22 μm organic filtration membrane was used to purify the samples and stored at − 20 °C until analysis.
Samples were analyzed using a liquid chromatography/mass spectrometry (LC/MS) system (Xevo G2-XS Qtof) and maintained in a thermostat-controlled chamber at 4 °C. Jasmonic acid and SA levels were calculated from the ratio of the endogenous hormone peak and the known internal standard. The peak area of each component was compared to the percent relative amount of internal standard peak area. For each treatment, four to six replicates were sampled.
Data analyses
Chi-squared tests were used to show differences in parasitoid species for the different categories of host plant odors. One-way ANOVA was used to show differences in phytohormone (JA and SA) levels and volatile components for the different treatment categories. Principal component analysis of plant volatiles was conducted and plotted using SIMCA software. High-throughput sequencing was used to elucidate global transcript abundance in rice plant, and compared the expression profiles of undamaged, artificial rolling, natural rolling and rolling and feeding plant treatments. All tests were performed at P < 0.05 statistical significance level and all data were analyzed in SPSS version 19.0 software.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Abbreviations
- GC/MS:
-
Gas chromatography/mass spectrometry
- HIPVs:
-
Herbivore-induced plant volatiles
- JA:
-
Jasmonic acid
- LC/MS:
-
Liquid chromatography/mass spectrometry
- MH63:
-
Minghui 63
- SA:
-
Salicylic acid
- VOC:
-
Volatile organic compounds
References
Sharma HC, Sujana G, Rao DM. Morphological and chemical components of resistance to pod borer, Helicoverpa armigera in wild relatives of pigeonpea. Arthropod-Plant Inte. 2009;3:151–61.
Ramegowda V, Senthil-Kumar M. The interactive effects of simultaneous biotic and abiotic stresses on plants: mechanistic understanding from drought and pathogen combination. J Plant Physiol. 2015;176:47–54.
Pandey P, Irulappan V, Bagavathiannan MV, Senthil-Kumar M. Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Front Plant Sci. 2017;8:537.
Bruce TJ. Interplay between insects and plants: dynamic and complex interactions that have coevolved over millions of years but act in milliseconds. J Exp Bot. 2014;66:455–65.
Agrawal AA. Current trends in the evolutionary ecology of plant defence. Funct Ecol. 2011;25:420–32.
Martinezmedina A, Flors V, Heil M, Mauchmani B, Cmj P, Pozo MJ, et al. Recognizing plant defense priming. Trends Plant Sci. 2016;21:818–22.
Navarro L, Bari R, Achard P, Lisón P, Nemri A, Harberd NP, et al. DELLAs control plant immune responses by modulating the balance of jasmonic acid and salicylic acid signaling. Curr Biol. 2008;18:650–5.
Bari R, Jones JDG. Role of plant hormones in plant defence responses. Plant Mol Biol. 2009;69:473–88.
Nishiyama R, Watanabe Y, Leyva-Gonzalez MA, Ha CV, Fujita Y, Tanaka M, et al. Arabidopsis AHP2, AHP3, and AHP5 histidine phosphotransfer proteins function as redundant negative regulators of drought stress response. P Natl Acad Sci USA. 2013;110:4840–5.
Robert-Seilaniantz A, Grant M, Jones JD. Hormone crosstalk in plant disease and defense: more than just jasmonate-salicylate antagonism. Annu Rev Phytopathol. 2011;49:317–43.
Das A, Lee SH, Hyun TK, Kim SW, Kim JY. Plant volatiles as method of communication. Plant Biotechnol Rep. 2013;7:9–26.
Erb M, Meldau S, Howe GA. Role of phytohormones in insect-specific plant reactions. Trends Plant Sci. 2012;17:250–9.
Xu S, Zhou W, Pottinger S, Baldwin IT. Herbivore associated elicitor-induced defences are highly specific among closely related nicotiana species. BMC Plant Biol. 2015;15:2.
Aljbory Z, Chen MS. Indirect plant defense against insect herbivores: a review. Insect Sci. 2018;25(1):2–23.
Delphia CM, Mescher MC, Moraes CMD. Induction of plant volatiles by herbivores with different feeding habits and the effects of induced defenses on host-plant selection by thrips. J Chem Ecol. 2007;33:997–1012.
Yuan JS, Himanen SJ, Holopainen JK, Chen F, Stewart CNJ. Smelling global climate change: mitigation of function for plant volatile organic compounds. Trends Ecol Evol. 2009;24:323–31.
Kessler A. Plant responses to insect herbivory: the emerging molecular analysis. Annu Rev Plant Biol. 2002;53:299–328.
Dicke M, Hilker M. Induced plant defenses: from molecular biology to evolutionary ecology. Basic Appl Ecol. 2003;4:3–14.
Hare JD. Ecological role of volatiles produced by plants in response to damage by herbivorous insects. Annu Rev Entomol. 2011;56:161–80.
Ponzio C, Cascone P, Cusumano A, Weldegergis BT, Gols R. Volatile-mediated foraging behaviour of three parasitoid species under conditions of dual insect herbivore attack. Anim Behav. 2016;111:197–206.
Puente M, Magori K, Kennedy GG, Gould F. Impact of herbivore-induced plant volatiles on parasitoid foraging success: a spatial simulation of the Cotesia rubecula, Pieris rapae, and Brassica oleracea system. J Chem Ecol. 2008;34:959–70.
Aartsma Y, Bianchi FJJA, Wopke VDW, Poelman EH, Dicke M. Herbivore-induced plant volatiles and tritrophic interactions across spatial scales. New Phytol. 2017;216:1054–63.
Diezel C, Dahl CCV, Gaquerel E, Baldwin IT. Different lepidopteran elicitors account for cross-talk in herbivory-induced phytohormone signaling. Plant Physiol. 2009;150:1576.
Acevedo FE, Rivera-Vega LJ, Chung SH, Ray S, Felton GW. Cues from chewing insects — the intersection of DAMPs, HAMPs, MAMPs and effectors. Curr Opin Plant Biol. 2015;26:80–6.
Reddy GV, Holopainen JK, Guerrero A. Olfactory responses of Plutella xylostella natural enemies to host pheromone, larval frass, and green leaf cabbage volatiles. J Chem Ecol. 2002;28:131–43.
Hilker M, Meiners T. Early herbivore alert: insect eggs induce plant defense. J Chem Ecol. 2006;32:1379–97.
Hilker M, Meiners T. How do plants “notice” attack by herbivorous arthropods? Biol Rev. 2010;85:267–80.
Wilson EO, Hölldobler B. Sex differences in cooperative silk-spinning by weaver ant larvae. P Natl Acad Sci USA. 1980;77:2343–7.
Berger A. Ballooning activity of chilo partellus larvae in relation to size of mother, egg batches, egg and larvae and age of mother. Entomol Exp Appl. 1989;50:125–32.
Diss AL, Kunkel JG, Montgomery ME, Leonard DE. Effects of maternal nutrition and egg provisioning on parameters of larval hatch, survival and dispersal in the gypsy moth, Lymantria dispar L. Oecologia. 1996;106:470–7.
Weseloh RM. Effects on behavior of Apanteles melanoscelus females caused by modifications in extraction, storage, and presentation of gypsy moth silk kairomone. J Chem Ecol. 1977;3:723–35.
Ha DM, Choi SH, Shim JK, Jung DO, Song KS, Nho SK, et al. Behavioral attraction of two parasitoids, Venturia canescens and Bracon hebetor, to silk extracts of a host Plodia interpunctella. J Asia Pac Entomol. 2006;9:287–92.
Inoue H, Kamiwada H, Fukamachi S. Seasonal changes in adult density and female mating status of the rice leaf roller Cnaphalocrocis medinalis guenee (lepidoptera: pyralidae) in paddy fields of southern Kyushu, Japan. Jpn J Appl Entomol Z. 2004;48:177–83.
Fraenkel G, Fallil F, Kumarasinghe K. The feeding behaviour of the rice leaf folder, Cnaphalocrocis medinalis. Entomol Exp Appl. 1981;29:147–61.
Nathan SS, Chung PG, Murugan K. Effect of botanical insecticides and bacterial toxins on the gut enzyme of the rice leaf folder Cnaphalocrocis medinalis. Phytoparasitica. 2004;32:433–43.
Rani WBB, Amutha R, Muthulakshmi S, Indira K, Mareeswari P. Diversity of rice leaf folders and their natural enemies. Res J Agric Biol Sci. 2007;3:394–7.
Gurr GM, Read DMY, Catindig JLA, Cheng J, Liu J, Lan LP, et al. Parasitoids of the rice leaffolder Cnaphalocrocis medinalis and prospects for enhancing biological control with nectar plants. Agr Forest Entomol. 2012;14:1–12.
Khan ZR, Rueda BP, Caballero P. Behavioral and physiological responses of rice leaffolder Cnaphalocrocis medinalis, to selected wild Rices. Entomol Exp Appl. 1989;52:7–13.
Fraenkel G, Fallil F. The spinning (stitching) behaviour of the rice leaf folder, Cnaphalocrocis medinalis. Entomol Exp Appl. 2011;29:138–46.
Saravanakumar D, Muthumeena K, Lavanya N, Suresh S, Rajendran L, Raguchander T, et al. Pseudomonas-induced defence molecules in rice plants against leaffolder (Cnaphalocrocis medinalis) pest. Pest Manag Sci. 2007;63:8.
Kalaivani K, Kalaiselvi MM, Senthil-Nathan S. Effect of methyl salicylate (MeSA) induced changes in rice plant (Oryza sativa) that affect growth and development of the rice leaffolder, Cnaphalocrocis medinalis. Physiol Mol Plant P. 2018;101:116–26.
Zebelo SA, Maffei ME. Role of early signalling events in plant-insect interactions. J Exp Bot. 2015;66:435–48.
Helms AM, De Moraes CM, Tröger A, Alborn HT, Francke W, Tooker JF, et al. Identification of an insect-produced olfactory cue that primes plant defenses. Nat Commun. 2017;8(1):337.
Shikano I, Rosa C, Tan CW, Felton GW. Tritrophic interactions: microbe-mediated plant effects on insect herbivores. Annu Rev Phytopathol. 2017;55:annurev-phyto-080516-035319.
Woldemariam MG, Ahern K, Jander G, Tzin V. A role for 9-lipoxygenases in maize defense against insect herbivory. Plant Signal Behav. 2018;13(1):00 Zhou W, Thomas Brockmöller, Ling Z, Omdahl A, Baldwin IT, Xu S: Evolution of herbivore-induced early defense signaling was shaped by genome-wide duplications in nicotiana. Elife 2016, 5:5.
Bittner N, Trauer-Kizilelma U, Hilker M. Early plant defence against insect attack: involvement of reactive oxygen species in plant responses to insect egg deposition. Planta. 2017;245:993–1007.
Reymond P, Weber H, Damond M, Farmer EE. Differential gene expression in response to mechanical wounding and insect feeding in Arabidopsis. Plant Cell. 2000;12:707–19.
Jacobo-Velázquez DA, González-Agüero M, Cisneros-Zevallos L. Cross-talk between signaling pathways: the link between plant secondary metabolite production and wounding stress response. Sci Rep-UK. 2015;5:8608.
Dong Z, Zhao P, Wang C, Zhang Y, Chen J, Wang X, et al. Comparative proteomics reveal diverse functions and dynamic changes of Bombyx mori silk proteins spun from different development stages. J Proteome Res. 2013;12:5213–22.
Bi JL, Murphy JB, Felton GW. Does salicylic acid act as a signal in cotton for induced resistance to Helicoverpa zea? J Chem Ecol. 1997;23:1805–18.
van Poecke RMP, Dicke M. Induced parasitoid attraction by Arabidopsis thaliana: involvement of the octadecanoid and the salicylic acid pathway. J Exp Bot. 2002;53:1793–9.
Sun X, Zhou W, Liu H, Zhang AJ, Ai CR, Zhou SS, et al. Transgenic Bt rice does not challenge host preference of the target pest of rice leaffolder, Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). PLoS One. 2013;8:e79032.
Pan X, Welti R, Wang X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography-mass spectrometry. Nat Protoc. 2010;5:986–92.
Acknowledgements
We appreciate the Testing and Analysis Center of Huazhong Agricultural University staff for technical assistance during laboratory experiments.
Funding
This research was financially supported by the National Key Research and Development Program (No. 2017YFE0113900) and the Special Technical Innovation of Hubei Province (2017ABA146). The funding body played no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Author information
Authors and Affiliations
Contributions
J-HS, ZS, HL and M-QW conceived and designed the experiments; J-HS, X-JH, CW, LL and CNF performed the experiments; J-HS, ZS, H-NJ and F-FL analyzed the data; J-HS, ZS, CNF and M-QW wrote the paper. All contributing authors agreed to the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All plant materials (including seeds) used in our study were permitted and obtained from the National Key Laboratory of Crop Genetic Improvement, Huazhong Agricultural University. Experimental research on plant, including collection of plant materials, complied with institutional, national, or international guidelines.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests。.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1: Figure S1.
Kyoto Encyclopedia of Genes and Genomes of rice plants. (a) undamaged plant vs natural rolling plant by Cnaphalocrocis medinalis and; (b) artificially rolled plant vs natural rolling plant by C. medinalis, (c) natural rolling plant vs rolling and feeding by C. medinalis.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Shi, JH., Sun, Z., Hu, XJ. et al. Rice defense responses are induced upon leaf rolling by an insect herbivore. BMC Plant Biol 19, 514 (2019). https://doi.org/10.1186/s12870-019-2116-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-019-2116-0