
Abstract
Histone deacetylase (HDAC) inhibitors affect cell homeostasis, gene expression, and cell cycle progression and promote cell terminal differentiation or apoptosis. However, the effect of HDAC inhibition on SH-SY5Y cells, which are neuroblastoma cells capable of differentiating into neurons under specific conditions, such as in the presence of retinoic acid (RA), is unknown. In this study, we hypothesized that HDAC inhibitors induced the neuronal differentiation of SH-SY5Y cells. To test this hypothesis, we used phase contrast microscopy, immunocytochemistry (ICC), qPCR, and western blotting analysis. MS-275 and valproic acid (VPA), two HDAC inhibitors, were selected to evaluate neuronal differentiation. It was confirmed that cells treated with MS-275 or VPA differentiated into mature neurons, which were distinguished by bipolar or multipolar morphologies with elongated branches. In addition, the mRNA expression of neuronal markers (Tuj1 and NEFH) and the oligodendrocyte marker (CNP) was significantly increased with MS-275 or VPA treatment compared to that with RA treatment. In addition, the protein expression of the other neuronal markers, Tuj1 and NeuN, was highly increased with HDAC inhibitor treatments compared to that with RA treatment. Furthermore, we confirmed that noncanonical Wnt signaling was upregulated by HDAC inhibitors via MAPK signaling and the Wnt/JNK pathway. Therefore, both MS-275 and VPA promoted the differentiation of SH-SY5Y cells into mature neurons via the Wnt signaling pathway.
Similar content being viewed by others

Introduction
Histone deacetylase (HDAC) inhibitors affect various cellular signals, such as cell proliferation, terminal differentiation, apoptosis, cell homeostasis, cell cycle arrest, and gene expression [1, 2]. When histone acetyltransferase (HAT) transfers the acetyl groups of acetyl-CoA to the lysine residues in the NH2 terminal tails of core histones, the chromosome is uncoiled into chromatin, making transcription easier [3]. When HDACs remove the acetyl group, the chromatin condenses back into a chromosome [1, 4]. This epigenetic mechanism activates cell proliferation and makes transcription difficult [1]. Conversely, HDAC inhibitors prevent histone lysine residues from being deacetylated. Because acetylation leads to uncoiled chromatin, it activates transcription and affects gene expression.
MS-275, which is an HDAC1, HDAC2, and HDAC3 inhibitor, is a candidate for treating autism and cancers such as advanced breast cancer and metastatic lung cancer [1, 5, 6]. Some studies suggest that MS-275 improves social and synaptic function associated with autism and alleviates postoperative cognitive dysfunction by reducing hippocampal neuroinflammation [6, 7]. MS-275 also induces the odontoblast differentiation of human dental pulp stem cells and the neurogenic differentiation of human adipose tissue-derived mesenchymal stem cells [1, 8]. Valproic acid (VPA) is a class I and class IIa HDAC inhibitor that has been clinically used as a therapeutic agent for the treatment of breast cancer, intracranial glioma, bipolar disorder, brain metastases, spinal muscular atrophy, and migraine [1, 9, 10]. VPA can act on the cardiovascular system and can be used to manage diabetes mellitus; it also has anti-inflammatory and neuroprotective effects [11]. VPA has been shown to alter estrogen receptor status and metastatic potential and modulates the tumor immune response of breast cancer cells [9]. Other studies suggest that VPA promotes the neuronal differentiation of adipose tissue-derived stem cells via the Wnt signaling pathway or the inducible nitric oxide synthase-soluble guanylyl cyclase signaling pathway [1, 12]. One study also showed that VPA enhances the neuronal differentiation of spiral ganglion neural stem cells with much longer neurite outgrowth via the Wnt/β-catenin signaling pathway [13].
The Wnt signaling pathway, which is associated with proliferation, apoptosis, differentiation, and the cell cycle, can be classified into the canonical and noncanonical pathways [14]. In the canonical pathway, when Wnt (Wnt1, Wnt2, or Wnt3α) is activated, it can bind with the frizzled (Fzd) receptor and stimulate the coreceptors, low-density lipoprotein receptor-related protein 5 (LRP5) or LRP6 and dishevelled (Dvl), to inhibit β-catenin proteolysis [15]. β-Catenin dissociates from the destruction complex, composed of Axin1, adenomatosis polyposis coli (APC), phosphorylated glycogen synthase kinase 3β (p-GSK3β), and casein kinase 1 (CK1), and translocates into the nucleus leading to gene transcription [15, 16]. The noncanonical pathway, which does not regulate β-catenin, has the following two downstream signals; the Wnt/Ca2+ pathway and the Wnt/JNK pathway. Wnt4, Wnt5α, or Wnt11, which are known to be noncanonical forms of Wnt, bind with Fzd and Dvl, which leads to phospholipase C (PLC) activation in the Wnt/Ca2+ pathway. Increased Ca2+ stimulates protein kinase C (PKC), calcineurin, and calcium/calmodulin-dependent protein kinase II (CaMKII), and nuclear factor of activated T-cell (NFAT) translocates into the nucleus leading to gene transcription [14]. Several studies have demonstrated that activated Dvl stimulates phosphorylated ERK and JNK and leads to gene transcription and neuronal differentiation via c-Jun in the Wnt/JNK pathway [1, 14].
Although some studies have assessed neuronal differentiation using HDAC inhibitors, the effects of neuronal differentiation on SH-SY5Y cells by HDAC inhibitors have not been well demonstrated [17,18,19]. Therefore, in this study, we confirmed the effect of HDAC inhibitors on the differentiation of SH-SY5Y cells into mature neurons via the Wnt signaling pathway.
Materials and methods
Cell lines and reagents
We used the human neuroblastoma cell line, SH-SY5Y (RRID: CVCL_0019l ATCC® CRL-2266), which can differentiate into mature neuronal cells under certain conditions. The cells were grown in Dulbecco's modified Eagle's medium (DMEM; WELGENE, Gyeongsan-si, Republic of Korea) supplemented with 10% fetal bovine serum (FBS; EMD Millipore Corp., Burlington, Massachusetts, USA), 1% penicillin–streptomycin (WELGENE), and 0.2% amphotericin B (Life Technologies Corporation, Carlsbad, USA) at 37℃ in a 5% CO2 incubator. Retinoic acid (RA; 10 μM), a known reagent for the neuronal differentiation of SH-SY5Y cells, was dissolved in dimethyl sulfoxide (DMSO; Sigma–Aldrich, Burlington, MA, USA) and used as a positive control in this study. We selected two HDAC inhibitors, MS-275 and VPA, and dissolved them in DMSO as described in a previous study [1]. The concentration of the HDAC inhibitors was determined according to several references based on the inhibition of HDAC, activation of HAT [20,21,22,23,24,25], and neuronal differentiation [1, 26,27,28]. Differences in shape were confirmed by taking images using a phase-contrast microscope; neuronal differentiation occurred most actively with HDAC inhibitor treatment. In addition, the cell viability was determined by MTT assay and then the concentration of the HDAC inhibitors was chosen.
Immunocytochemistry (ICC)
For ICC, the cells (5 × 104 cells/mL) were cultured on poly-L-lysine-coated aclar plastic coverslips as described previously [29]. Medium containing 1% FBS (EMD Millipore Corp.) was changed to medium containing the HDAC inhibitors. The cells were fixed with 4% paraformaldehyde (PFA; T&I Co., Chuncheon-si, Republic of Korea) for 15 min at room temperature. The cells were then blocked with 0.5% Triton X-100 (Sigma–Aldrich) for 20 min and 10% normal goat serum (NGS; Vector Laboratories, Inc., Burlingame, USA) for 30 min. Primary antibodies were added for 1.5 h, and the secondary antibodies were then added and kept in the dark for 1 h. For staining nuclei, 4′,6-diamidino-2-phenylindole (DAPI, 1 μg/mL, Life Technologies Corporation) was added for 30 min. The cells were then imaged with a Zeiss LSM510 confocal microscope (Carl Zeiss, Jena, Germany) after mounting [29]. The primary antibodies used were microtubule-associated protein 2 (MAP2, 1:200; Cell Signaling Technology, Danvers, USA) and neurofilament-H (NFH, 1:400; Cell Signaling Technology). Alexa 488-conjugated goat anti-rabbit (Invitrogen Co., Waltham, USA) and Alexa 594-conjugated goat anti-mouse (Invitrogen Co.) were used as the secondary antibodies. All experiments were repeated at least three times.
RNA isolation and cDNA synthesis
For qPCR, cells (5 × 105 cells/mL) treated with HDAC inhibitors were harvested using TRIzol (TaKaRa Co., Shimogyo-ku, Japan) as described previously [1]. Recombinant RNase Inhibitor (TaKaRa Co.), Go script Buffer Mix Oligo dT (Promega Co., Madison, USA), and Go script Enzyme Mix (Promega Co.) were added to synthesize cDNA according to the manufacturer’s instructions. A PCR instrument (Takara Co. ) was used for denaturing RNA and synthesizing cDNA. The synthesized cDNA was stored at 4 °C.
Quantitative PCR (qPCR)
qPCR analyses were performed with SYBR Green Premix Ex Taq (Takara Co. ) and LightCycler 480 II (Roche Holding AG., Basel, Swiss) at 60 °C, as recommended by the manufacturer’s instructions. All primers listed in Table 1 were purchased from Bioneer and CosmoGenetech (Seoul, Korea). Each sample was analyzed in three replicate reactions of 10 μL.
Western blotting analysis
For western blotting analysis, cells (5 × 105 cells/mL) were cultured and lysed in a lysis buffer (1 M Tris pH 7.5, 1 M NaCl, 0.5 M EDTA, 10% Nonidet P-40, 100% glycerol, 50 mg/mL leupeptin, 50 mg/mL aprotinin, 0.2 M PMSF, 0.1 M Na3VO4, 1 M NaF) as described previously [37]. The western blot membranes containing proteins were incubated with specific antibodies against neurofilament-heavy chain (NFH, 1:1000), Tuj1 (1:1000), synaptophysin (SYP, 1:1000), neuronal nuclei (NeuN, 1:2000), Wnt5α/β (1:1000), Fzd5 (1:1000), Dvl2 (1:1000), Dvl3 (1:1000), Axin1 (1:1000), PKC (1:1000) p-ERK (1:1000), ERK (1:1000), p-JNK (1:1000), JNK (1:1000), c-Jun (1:1000), and GAPDH (1:3000) [37, 38]. Horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG antibody and HRP-conjugated goat anti-mouse IgG antibody were used as the secondary antibodies. All primary and secondary antibodies were purchased from Cell Signaling Technology and Santa Cruz Biotechnology. The immunoblotted bands were measured by Immobilon Crescendo Western HRP substrate (EMD Millipore Corporation). ImageJ software was used for quantitative analysis of immunoblotted bands [37]. All experiments were repeated at least three times.
Statistical analysis
Statistical analyses were performed with GraphPad Prism® 5.0 software (GraphPad Software Inc., San Diego, CA, USA). Differences among groups were assessed using two-way ANOVA. Repeated measures were assessed by ANOVA with significance levels of *p < 0.05, **p < 0.01, and ***p < 0.001 compared to the control; and #p < 0.05, ##p < 0.01, and ###p < 0.001 compared to the RA-treated group. If mean values between populations showed statistically significant changes, they were analyzed by post–hoc tests (Bonferroni). ICC, qPCR, and western blotting data were analyzed using one–way ANOVA and post–hoc tests (Tukey’s).
Results
Morphological changes
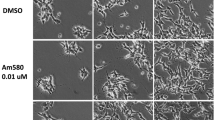
To assess whether the cell morphology was changed with HDAC inhibitor treatment, we imaged cells treated with HDAC inhibitors under a microscope for 1 week. After RA and HDAC inhibitor (MS-275 or VPA) treatment, the cells had bipolar or multipolar neurites with elongated branches compared to the control cells. These morphological changes suggest that the HDAC inhibitors induced the differentiation of SH-SY5Y cells into mature neurons (Fig. 1).

Neuron-like morphology after HDAC inhibitor treatment. SH-SY5Y cells were cultured with RA (10 μM), MS-275 (500 nM), or VPA (20 μM) and images were taken on the 1st and 7th days under a microscope. HDAC inhibitors (MS-275 and VPA) induced neuronal differentiation effectively compared to control and RA treatments. The scale bar represents 100 μm
We measured cell death in different concentrations following treatment with HDAC inhibitors, and the chosen concentrations did not have an effect on the death of the cells (Additional file 1: Fig. S1). These data confirmed that the cell viabilities were decreased with high concentrations of HDAC inhibitors [21, 25, 39]. In addition, to investigate the inhibition of histone deacetylation with HDAC inhibitor treatment, the protein expression of HDAC was determined using western blotting (Additional file 1: Figs. S2 and S3). We figured out that the concentration of HDAC inhibitors affected histone modification, as in previous reports [20,21,22,23,24,25].
Neuronal differentiation of SH-SY5Y cells with HDAC inhibitors
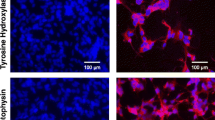
To determine the increase in neuronal markers, we analyzed SH-SY5Y cells treated with HDAC inhibitors using immunocytochemistry with primary antibodies against MAP2 and NFH. MAP2 is located in the dendrites of neurons and the soma, and NFH, a component of the cytoskeleton, is usually located in axons. Figure 2 shows the neurite outgrowth and morphology of mature neurons treated HDAC inhibitors compared to the control and RA groups. Neuronal marker-positive cells were increased with the following HDAC inhibitor treatments; 32.79 ± 3.313% in control, 50.18 ± 3.037% in RA, 68.48 ± 0.5366% in MS-275, and 66.74 ± 4.477%, in VPA (Fig. 2b).

Analysis of neuronal markers by fluorescent immunocytochemistry. a Fluorescent immunocytochemistry showing MAP2-positive cells (green) and NFH-positive cells (red) following HDAC inhibitor treatment compared to the control and RA treated cells. DAPI (blue) was used to stain the nuclei of cells. The scale bar represents 20 μm. b The number of positive cells that expressed MAP2 and NFH was measured. The ratio of positive cells to nuclei was calculated for each group (n = 4). All experiments were repeated at least three times. ***p < 0.001 compared to the control group, #p < 0.05 and ##p < 0.01 compared to the RA group
Neuronal marker genes were highly expressed with HDAC inhibitor treatments
In a previous study, we investigated the effect of HDAC inhibitors on stem cell differentiation into mature neurons, oligodendrocytes, or astrocytes [1]. Here, we determined that the mRNA expression of NEFH, a mature neuronal marker, and CNP, an oligodendrocyte marker, was significantly increased with VPA group compared to RA group (Fig. 3) using qPCR analyses. Interestingly, the expression of Tuj1 was increased with MS-275 treatment. These results suggest that MS-275 and VPA are effective inducers of neuronal differentiation and are, therefore, novel drugs for SH-SY5Y cell differentiation [40, 41].

qPCR assay. We analyzed the gene expression of MAP2, NEFL, NEFH, NEFM, CNP, Tuj1, and GFAP by qPCR. The gene expression of most neuronal markers was increased with HDAC inhibitor treatment compared to RA treatment. The mRNA expression was normalized to the RA-treated cells. GAPDH was used as a control. All experiments were repeated at least three times. ##p < 0.01 and ###p < 0.001 compared to the RA group
Regulation of neuronal differentiation via Wnt/JNK signaling
We studied the protein expression of neuronal markers, such as NFH, Tuj1, SYP, and NeuN, by western blotting analysis. The expression of NeuN, Tuj1, SYP, and NFH was increased with HDAC inhibitors and RA treatment compared to the control group. Interestingly, Tuj1 and SYP were significantly upregulated following treatment with HDAC inhibitors compared that with to RA treatment (Fig. 4a, b, and Additional file 1: Fig. S4).

Expression of neuronal markers and Wnt-related signals. The levels of neuronal markers (a, b), Wnt-related proteins (c and d), and MAPK signaling (e and f) were determined via western blotting analysis. GAPDH was used as a control. Original uncut western blotting images are shown in Additional file 1: Figs. S4, S5, S6. All experiments were repeated at least three times. *p < 0.05, **p < 0.01, and ***p < 0.001 compared to the control group
Previously, our lab and several others studied the Wnt signaling pathway, which is related to neuronal differentiation [1, 38, 42, 43]. Therefore, here we investigated whether the Wnt signaling pathway affects neuronal differentiation following HDAC inhibition. Interestingly, Wnt5α/β, which is related to noncanonical Wnt signaling, was highly increased with the HDAC inhibitors and RA treatment compared to the control group. Similarly, Fzd5, which is located around the cytoplasmic membrane and binds to canonical and noncanonical Wnt, was increased with the HDAC inhibitor and RA treatment compared to the control group. Dvl is connected with Fzd and regulates downstream signaling. Dvl2 was decreased while Dvl3 was increased following treatment with the HDAC inhibitors. Importantly, Axin1, which interacts with GSK3β and β-catenin, was decreased with VPA treatment compared to the control group (Fig. 4c, d, and Additional file 1: Fig. S5).
The noncanonical Wnt signaling pathway independent of β-catenin is classified as the Wnt/Ca2+ and Wnt/JNK pathways. To assess the activation of noncanonical Wnt signaling with HDAC inhibitor treatment, phosphorylation-specific antibodies were used as previously described [1]. In the Wnt/Ca2+ pathway, the expression of PKC was stimulated by HDAC inhibitor treatment. In the Wnt/JNK pathway, phosphorylated ERK was activated by HDAC inhibitor treatment. Interestingly, the expression of phosphorylated JNK was highly upregulated with RA treatment. The expression of c-Jun, the transcription factor of the noncanonical pathway, was increased with MS-275, VPA, and RA treatments compared to the control (Fig. 4e, f, and Additional file 1: Fig. S6). Taken together, these data show that HDAC inhibitors induced neuronal differentiation of SH-SY5Y cells by activating the noncanonical Wnt signaling pathway, specifically, the Wnt/JNK pathway (Fig. 5).

Noncanonical Wnt signaling for neuronal differentiation with HDAC inhibitors. HDAC inhibitors activate Wnt4, Wnt5α, Wnt 11, and the Fzd receptor. Fzd activates Dvl and stimulates the Wnt/Ca2+ or Wnt/JNK pathway. In the Wnt/Ca2+ pathway, activated Dvl promotes Ca2+, which stimulates PKC. In the Wnt/JNK pathway, activated Dvl promotes phosphorylated ERK and phosphorylated JNK. Finally, c-Jun, a transcription factor, binds to DNA and promotes neuronal differentiation. Here, we demonstrate that Wnt4/5α/11 can modulate Dvl/PKC signaling and the Dvl/p-ERK/p-JNK pathway for neuronal differentiation following HDAC inhibition. Orange color indicates the signaling observed in this study
Discussion
HDAC inhibitors are related to the induction of apoptosis and autophagy, cell cycle arrest, hormone signaling, immune effects, neuroprotection, and neuronal differentiation [1, 2, 44, 45]. Some HDAC inhibitors, such as romidepsin, vorinostat, or ricolinostat, are already approved or advanced as clinical candidates for cancer therapy [46]. MS-275, another HDAC inhibitor, decreases cell proliferation and induces the differentiation of human dental pulp stem cells into odontoblast-like cells [8] and human mesenchymal stem cells [1]. MS275 also has been known to differentiate U87MG glioblastoma multiforme cells into neural cells when it synergized with 8-CPT-cAMP [47]. Another HDAC inhibitor, VPA, has been known to effective inducer of differentiation. For example, it could stimulate neurite outgrowth and prolong cell survival via the ERK [45] and JNK [1] pathway. In addition, VPA could induce the differentiation of PC12 cells which are the neuroblastoma cell into mature neurons [48]. Other groups were reported that adipose tissue-derived stem cells could also differentiate into neuronal cells by treating VPA [12, 49]. Similarly, it has been reported that VPA has a potential as an agent of neurogenic differentiation via activating histone H3 core acetylation in embryonic stem cells [50] and osteogenic differentiation in tonsil-derived mesenchymal stem cells [51]. Animal studies have also shown that VPA treatment in the rat cerebral cortex induced differentiation via activation of the ERK-P21 Cip/WAFI pathway [45, 52]. In this study, we demonstrated that MS-275 and VPA were effective inducers of neuronal differentiation via the activated ERK/JNK pathway by Wnt signaling.
We investigated the effect of HDAC inhibitors (MS-275 and VPA) on neuronal differentiation by observing morphology via phase contrast and ICC. The expression of both MAP2 [53] and NFH [54, 55] was markedly increased with HDAC inhibitors (MS-275 or VPA) compared to the control and RA-treated cells. Previous studies have reported that HDAC inhibitors can regulate cell proliferation and cell apoptosis in numerous cancer cell lines [56, 57]. Similarly, cell proliferation was not increased with MS275 and VPA treatment. However, the neurites were elongated and the numbers increased (Figs. 1 and 2).
HAT is important for astrocyte differentiation, and HDAC is important for oligodendrocyte differentiation [1, 58]. We also obtained the similar results by qPCR analysis that acetylation via HDAC inhibitors promotes the differentiation of oligodendrocytes. In the western blotting analysis, we assessed whether the protein expression of NeuN, Tuj1, SYP, and NFH was increased with HDAC inhibitors. NeuN, which is a soluble nuclear protein related to terminal neuronal differentiation, was used to estimate neuronal cell loss in NDDs [55]. Tuj1 can be used to determine microtubule protofilament stability and was used to evaluate the expression of neuronal markers [59]. SYP, which is a membrane glycoprotein located in presynaptic vesicles, regulates the endocytosis of synaptic vesicles [55, 60]. NFH plays a role in axonal architecture and neurite outgrowth [55]. In this study, the expression of NeuN, Tuj1, SYP, and NFH was highly increased with HDAC inhibitors compared to that with the control. In addition, the levels of Tuj1 and SYP were enhanced with HDAC inhibitors compared to that following RA treatment. These data suggest that MS-275 and VPA could be used as effective inducers of neuronal differentiation (Figs. 3, 4 and Additional file 1: Fig. S4 ).
The Wnt signaling pathway is classified as either the canonical pathway, which is dependent on β-catenin, or the noncanonical pathway, which is independent of β-catenin [15, 16]. Noncanonical Wnt signaling is further classified as the Wnt/Ca2+ and Wnt/JNK pathways. In our results, Wnt4, Wnt5α, and Wnt11 were activated by the HDAC inhibitors and bound to the Fzd receptor to stimulate Dvl (Fig. 4 and Additional file 1: Figs. S5, S6). Stimulated Dvl activates PLC and PKC, and the subsequent signals increase Ca2+ via IP3-activated PKC, calcineurin, and CaMKII [14]. Activated calcineurin removes the phosphate group from NFAT, and NFAT translocate to the nucleus to activate transcription and differentiation in the Wnt/Ca2+ pathway [14]. In the Wnt/JNK pathway, activated Dvl stimulates phosphorylated ERK and JNK and activates c-Jun, a transcription factor, to increase transcription and cell differentiation. It has been reported that a single Wnt5α can promote neuronal differentiation via the Wnt5α/JNK pathway in human adipose tissue-derived stem cells [38]. Another study has reported that VPA can induce the differentiation of human pluripotent stem cells into spermatogonial stem-cell like cells by activating Wnt signaling pathway [61]. We obtained similar results in SH-SY5Y cells treated with HDAC inhibitors. The increased expression of Wnt5α, Fzd5, PKC, p-ERK, p-JNK, and c-Jun with HDAC inhibitors suggests that MS-275 and VPA activated the noncanonical Wnt signaling pathway to promote the neuronal differentiation of SH-SY5Y cells. In addition, MS-275 induces the PP2A B-type subunit PR130, which is a modulator of Wnt signaling, in various cell types [62,63,64]. The PP2A-PR130 complex modulates the Wnt signal transduction pathway by restricting the functions of the intracellular Wnt regulator naked cuticle [62]. The expression of PR130 might be increased with HDAC inhibitor treatment in NB cells. This assumption requires further experimental evidence.
Some animal studies have suggested that HDAC inhibitors can be used as potential future therapeutics for neurological diseases. For example, it has been known that VPA promotes hippocampal neurogenesis and cell proliferation via the Wnt/β-catenin signaling pathway in the transgenic mouse model of Alzheimer’s diseases [65], Parkinson’s disease [66], and amyotrophic lateral sclerosis [66, 67]. In addition, VPA has been reported to induce synaptogenesis, axonal regeneration, and synaptic plasticity in the mice model of traumatic brain injury [68]. Several studies have been reported that Wnt regulators have been used as potential drugs for cancer treatment [1, 16, 69]. It has also been reported that the Wnt signaling pathway could be a therapeutic target to induce bone growth and skeletal tissue regeneration [70]. So, we expect that the Wnt signaling pathway could also be used as a therapeutic target for neurological disorders such as Alzheimer’s disease. Our next goal is to find a novel drug targeting the Wnt signaling pathway for neurological disorders and to investigate neuronal differentiation by histone acetylation in other cell lines. Taken together, these results indicate that MS-275 and VPA promote transcription and neuronal differentiation in SH-SY5Y cells by acetylation of lysine residues mediated by the noncanonical Wnt signaling.
Availability of data and materials
The datasets generated or analyzed during this study are available in the article and Additional file.
Abbreviations
- HDAC:
-
Histone deacetylase
- NB:
-
Neuroblastoma
- RA:
-
Retinoic acid
- VPA:
-
Valproic acid
- ICC:
-
Immunocytochemistry
- qPCR:
-
Quantitative PCR
- MAP2:
-
Microtubule-associated protein 2
- NEFL:
-
Neurofilament light chain
- NEFM:
-
Neurofilament medium chain
- NEFH:
-
Neurofilament heavy chain
- Tuj1:
-
β-Tubulin
- CNP:
-
2',3'-Cyclic-nucleotide 3'-phosphodiesterase
- GFAP:
-
Glial fibrillary acidic protein
- GAPDH:
-
Glyceraldehyde 3-phosphate dehydrogenase
- NeuN:
-
Neuronal nuclear protein
- SYP:
-
Synaptophysin
- Fzd:
-
Frizzled
- Dvl:
-
Dishevelled
- PKC:
-
Protein kinase C
- ERK:
-
Extracellular signal-regulated kinase
- JNK:
-
C-Jun N-terminal kinase
- APC:
-
Adenomatosis polyposis coli
- p-GSK3β:
-
Phosphorylated glycogen synthase kinase 3β
- CK1:
-
Kinases casein kinase 1
- PLC:
-
Phospholipase C
- NFAT:
-
Nuclear factor of activated T-cell
- DMEM:
-
Dulbecco's modified Eagle's medium
- FBS:
-
Fetal bovine serum
- DMSO:
-
Dimethyl sulfoxide
- PFA:
-
Paraformaldehyde
- NGS:
-
Normal goat serum
- DAPI:
-
4′,6-Diamidino-2-phenylindole
References
Jang S, Jeong HS. Histone deacetylase inhibition-mediated neuronal differentiation via the Wnt signaling pathway in human adipose tissue-derived mesenchymal stem cells. Neurosci Lett. 2018;668:24–30.
Liu Z, et al. HDAC inhibitor LMK235 promotes the odontoblast differentiation of dental pulp cells. Mol Med Rep. 2018;17(1):1445–52.
Miksiunas R, et al. Cardiomyogenic differentiation potential of human dilated myocardium-derived mesenchymal stem/stromal cells: the impact of HDAC inhibitor SAHA and biomimetic matrices. Int J Mol Sci. 2021;22(23):12702.
Shanmugam G, Rakshit S, Sarkar K. HDAC inhibitors: targets for tumor therapy, immune modulation and lung diseases. Transl Oncol. 2022;16: 101312.
Bradner JE, et al. Chemical genetic strategy identifies histone deacetylase 1 (HDAC1) and HDAC2 as therapeutic targets in sickle cell disease. Proc Natl Acad Sci U S A. 2010;107(28):12617–22.
Ma K, et al. Histone deacetylase inhibitor MS-275 restores social and synaptic function in a Shank3-deficient mouse model of autism. Neuropsychopharmacology. 2018;43(8):1779–88.
Wu Y, et al. Histone deacetylase inhibitor MS-275 alleviates postoperative cognitive dysfunction in rats by inhibiting hippocampal neuroinflammation. Neuroscience. 2019;417:70–80.
Lee EC, et al. The histone deacetylase inhibitor (MS-275) promotes differentiation of human dental pulp stem cells into odontoblast-like cells independent of the MAPK signaling system. Int J Mol Sci. 2020;21(16):5771.
Heers H, et al. Valproic acid as an adjunctive therapeutic agent for the treatment of breast cancer. Eur J Pharmacol. 2018;835:61–74.
Park SA, et al. Combination treatment with VPA and MSCs-TRAIL could increase anti-tumor effects against intracranial glioma. Oncol Rep. 2021;45(3):869–78.
Singh D, et al. Hidden pharmacological activities of valproic acid:a new insight. Biomed Pharmacother. 2021;142: 112021.
Okubo T, et al. Valproic acid promotes mature neuronal differentiation of adipose tissue-derived stem cells through iNOS-NO-sGC signaling pathway. Nitric Oxide. 2019;93:1–5.
Moon BS, Lu W, Park HJ. Valproic acid promotes the neuronal differentiation of spiral ganglion neural stem cells with robust axonal growth. Biochem Biophys Res Commun. 2018;503(4):2728–35.
Chae WJ, Bothwell ALM. Canonical and non-canonical Wnt signaling in immune cells. Trends Immunol. 2018;39(10):830–47.
Bugter JM, Fenderico N, Maurice MM. Mutations and mechanisms of WNT pathway tumour suppressors in cancer. Nat Rev Cancer. 2021;21(1):5–21.
Fu WB, Wang WE, Zeng CY. Wnt signaling pathways in myocardial infarction and the therapeutic effects of Wnt pathway inhibitors. Acta Pharmacol Sin. 2019;40(1):9–12.
Frumm SM, et al. Selective HDAC1/HDAC2 inhibitors induce neuroblastoma differentiation. Chem Biol. 2013;20(5):713–25.
Kiweler N, et al. The histone deacetylases HDAC1 and HDAC2 are required for the growth and survival of renal carcinoma cells. Arch Toxicol. 2018;92(7):2227–43.
Kiweler N, et al. Histone deacetylase inhibitors dysregulate DNA repair proteins and antagonize metastasis-associated processes. J Cancer Res Clin Oncol. 2020;146(2):343–56.
Camphausen K, et al. Enhancement of xenograft tumor radiosensitivity by the histone deacetylase inhibitor MS-275 and correlation with histone hyperacetylation. Clin Cancer Res. 2004;10(18 Pt 1):6066–71.
Franci G, et al. The class I-specific HDAC inhibitor MS-275 modulates the differentiation potential of mouse embryonic stem cells. Biol Open. 2013;2(10):1070–7.
Gojo I, et al. Phase 1 and pharmacologic study of MS-275, a histone deacetylase inhibitor, in adults with refractory and relapsed acute leukemias. Blood. 2007;109(7):2781–90.
Gottlicher M, et al. Valproic acid defines a novel class of HDAC inhibitors inducing differentiation of transformed cells. EMBO J. 2001;20(24):6969–78.
Lee S, et al. Histone deacetylase inhibitors decrease proliferation potential and multilineage differentiation capability of human mesenchymal stem cells. Cell Prolif. 2009;42(6):711–20.
Rosato RR, Almenara JA, Grant S. The histone deacetylase inhibitor MS-275 promotes differentiation or apoptosis in human leukemia cells through a process regulated by generation of reactive oxygen species and induction of p21CIP1/WAF1 1. Cancer Res. 2003;63(13):3637–45.
Gurvich N, et al. Histone deacetylase is a target of valproic acid-mediated cellular differentiation. Cancer Res. 2004;64(3):1079–86.
Jang S, Hwang J, Jeong HS. The role of histone acetylation in mesenchymal stem cell differentiation. Chonnam Med J. 2022;58(1):6–12.
Jang S, Jeong HS. Data for the effect of histone deacetylase inhibitors on voltage- and ligand-gated ion channel gene expression in neurogenic induced-human adipose tissue-derived mesenchymal stem cells. Data Brief. 2018;17:1314–9.
Jang S, et al. Functional neural differentiation of human adipose tissue-derived stem cells using bFGF and forskolin. BMC Cell Biol. 2010;11:25.
Mages B, et al. The cytoskeletal elements MAP2 and NF-L show substantial alterations in different stroke models while elevated serum levels highlight especially MAP2 as a sensitive biomarker in stroke patients. Mol Neurobiol. 2021;58(8):4051–69.
Loonstra FC, et al. Neuroaxonal and glial markers in patients of the same age with multiple sclerosis. Neurol Neuroimmunol Neuroinflamm. 2023;10(2): e200078.
Pipis M, et al. Charcot-Marie-Tooth disease type 2CC due to NEFH variants causes a progressive, non-length-dependent, motor-predominant phenotype. J Neurol Neurosurg Psychiatry. 2022;93(1):48–56.
Luck K, et al. A reference map of the human binary protein interactome. Nature. 2020;580(7803):402–8.
Haenig C, et al. Interactome mapping provides a network of neurodegenerative disease proteins and uncovers widespread protein aggregation in affected brains. Cell Rep. 2020;32(7): 108050.
Crabtree DV, et al. Tubulins in the primate retina: evidence that xanthophylls may be endogenous ligands for the paclitaxel-binding site. Bioorg Med Chem. 2001;9(8):1967–76.
Baptista I, et al. TKTL1 knockdown impairs hypoxia-induced glucose-6-phosphate dehydrogenase and glyceraldehyde-3-phosphate dehydrogenase overexpression. Int J Mol Sci. 2022;23(7):3574.
Jang S, et al. Neuroprotective effects of (-)-epigallocatechin-3-gallate against quinolinic acid-induced excitotoxicity via PI3K pathway and NO inhibition. Brain Res. 2010;1313:25–33.
Jang S, Park JS, Jeong HS. Neural differentiation of human adipose tissue-derived stem cells involves activation of the Wnt5a/JNK signalling. Stem Cells Int. 2015;2015: 178618.
Cho HH, et al. Induction of osteogenic differentiation of human mesenchymal stem cells by histone deacetylase inhibitors. J Cell Biochem. 2005;96(3):533–42.
Brenig K, et al. The proteomic landscape of cysteine oxidation that underpins retinoic acid-induced neuronal differentiation. J Proteome Res. 2020;19(5):1923–40.
Zhao H, et al. Combined use of bFGF/EGF and all-trans-retinoic acid cooperatively promotes neuronal differentiation and neurite outgrowth in neural stem cells. Neurosci Lett. 2019;690:61–8.
Yang H, et al. Folic acid promotes proliferation and differentiation of porcine pancreatic stem cells into insulin-secreting cells through canonical Wnt and ERK signaling pathway. J Steroid Biochem Mol Biol. 2021;205: 105772.
Abuna RPF, et al. Frizzled 6 disruption suppresses osteoblast differentiation induced by nanotopography through the canonical Wnt signaling pathway. J Cell Physiol. 2020;235(11):8293–303.
Jenke R, et al. Anticancer therapy with HDAC inhibitors: mechanism-based combination strategies and future perspectives. Cancers. 2021;13(4):634.
Shukla S, Tekwani BL. Histone deacetylases inhibitors in neurodegenerative diseases. Neuroprotect Neuronal Different Front Pharmacol. 2020;11:537.
Ho TCS, Chan AHY, Ganesan A. Thirty years of HDAC inhibitors: 2020 insight and hindsight. J Med Chem. 2020;63(21):12460–84.
Liu X, et al. Differential regulation of H3K9/H3K14 acetylation by small molecules drives neuron-fate-induction of glioma cell. Cell Death Dis. 2023;14(2):142.
Kelkawi AHA, et al. Differentiation of PC12 cell line into neuron by Valproic acid encapsulated in the stabilized core-shell liposome-chitosan nano carriers. Int J Biol Macromol. 2022;210:252–60.
Satoh A, et al. Valproic acid promotes differentiation of adipose tissue-derived stem cells to neuronal cells selectively expressing functional N-type voltage-gated Ca(2+) channels. Biochem Biophys Res Commun. 2022;589:55–62.
Baumann C, et al. Changes in chromatin accessibility landscape and histone H3 core acetylation during valproic acid-induced differentiation of embryonic stem cells. Epigenet Chromatin. 2021;14(1):58.
Yu Y, et al. Valproic acid-induced CCN1 promotes osteogenic differentiation by increasing CCN1 protein stability through HDAC1 inhibition in tonsil-derived mesenchymal stem cells. Cells. 2022;11(3):534.
Jung GA, et al. Valproic acid induces differentiation and inhibition of proliferation in neural progenitor cells via the beta-catenin-Ras-ERK-p21Cip/WAF1 pathway. BMC Cell Biol. 2008;9:66.
Dehmelt L, Halpain S. The MAP2/Tau family of microtubule-associated proteins. Genome Biol. 2005;6(1):204.
De Schaepdryver M, et al. Serum neurofilament heavy chains as early marker of motor neuron degeneration. Ann Clin Transl Neurol. 2019;6(10):1971–9.
Ramalingam M, Jang S, Jeong HS. Therapeutic effects of conditioned medium of neural differentiated human bone marrow-derived stem cells on rotenone-induced alpha-synuclein aggregation and apoptosis. Stem Cells Int. 2021;2021:6658271.
Bertrand P. Inside HDAC with HDAC inhibitors. Eur J Med Chem. 2010;45(6):2095–116.
Chen IC, Sethy B, Liou JP. Recent update of HDAC inhibitors in lymphoma. Front Cell Dev Biol. 2020;8: 576391.
Marin-Husstege M, et al. Histone deacetylase activity is necessary for oligodendrocyte lineage progression. J Neurosci. 2002;22(23):10333–45.
Ti SC, Alushin GM, Kapoor TM. Human beta-tubulin isotypes can regulate microtubule protofilament number and stability. Dev Cell. 2018;47(2):175–90.
Daly C, et al. Synaptophysin regulates clathrin-independent endocytosis of synaptic vesicles. Proc Natl Acad Sci U S A. 2000;97(11):6120–5.
Wang X, et al. Valproic acid promotes the in vitro differentiation of human pluripotent stem cells into spermatogonial stem cell-like cells. Stem Cell Res Ther. 2021;12(1):553.
Creyghton MP, et al. PR130 is a modulator of the Wnt-signaling cascade that counters repression of the antagonist Naked cuticle. Proc Natl Acad Sci U S A. 2006;103(14):5397–402.
Dzulko M, et al. The PP2A subunit PR130 is a key regulator of cell development and oncogenic transformation. Biochim Biophys Acta Rev Cancer. 2020;1874(2): 188453.
Goder A, et al. HDAC1 and HDAC2 integrate checkpoint kinase phosphorylation and cell fate through the phosphatase-2A subunit PR130. Nat Commun. 2018;9(1):764.
Zeng Q, et al. Valproic acid stimulates hippocampal neurogenesis via activating the Wnt/beta-catenin signaling pathway in the APP/PS1/Nestin-GFP triple transgenic mouse model of Alzheimer’s disease. Front Aging Neurosci. 2019;11:62.
Gyawali A, et al. Monocarboxylate transporter functions and neuroprotective effects of valproic acid in experimental models of amyotrophic lateral sclerosis. J Biomed Sci. 2022;29(1):2.
Kim T, et al. HDAC inhibition by valproic acid induces neuroprotection and improvement of PD-like behaviors in LRRK2 R1441G transgenic mice. Exp Neurobiol. 2019;28(4):504–15.
Liu S, et al. Combined cell grafting and VPA administration facilitates neural repair through axonal regeneration and synaptogenesis in traumatic brain injury. Acta Biochim Biophys Sin. 2022;54(9):1289–300.
Cheng X, et al. Therapeutic potential of targeting the Wnt/beta-catenin signaling pathway in colorectal cancer. Biomed Pharmacother. 2019;110:473–81.
Houschyar KS, et al. Wnt pathway in bone repair and regeneration—what do we know so far. Front Cell Dev Biol. 2018;6:170.
Acknowledgements
I would like to thank Professor Junho Lee (Department of Biotechnology, Chonnam National University) for my first experiences in the laboratory and encouraging my research as a research scientist.
Funding
This research was supported by grants from the National Research Foundation of Korea (Grant Numbers NRF-2021R1I1A3060435 and NRF-2020R1F1A1076616); a Grant from the Chonnam National University Hospital Biomedical Research Institute (BCRI22051); Chonnam National University (Grant Number: 2022-0036) and a Grant from the Jeollanam-do Science and Technology R&D Project (Development of Stem Cell-Derived New Drug), funded by Jeollanam-do, Korea.
Author information
Authors and Affiliations
Contributions
JC designing and conducting experiments, collection of results, drawing the diagram, statistical analyses, writing and revising the manuscript; JH, designing experiments and teaching the methods; MR, teaching the methods and statistical analyses; HSJ; conceptualization of the idea, teaching the methods, funding acquisition, and statistical analyses; SJ, conceptualization of the idea, teaching the methods, statistical analyses, funding acquisition, and revising the manuscript. . All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors claim that there are no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1
. Additional figures.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Choi, J., Hwang, J., Ramalingam, M. et al. Effects of HDAC inhibitors on neuroblastoma SH-SY5Y cell differentiation into mature neurons via the Wnt signaling pathway. BMC Neurosci 24, 28 (2023). https://doi.org/10.1186/s12868-023-00798-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12868-023-00798-0