
Abstract
Objectives
Colibacillosis is a frequent enteric disease in the pig industry that causes significant economic losses. The objective of this study was to investigate the molecular characteristics of fluoroquinolone (FQ)-resistant E. coli isolates from suckling piglets with colibacillosis.
Results
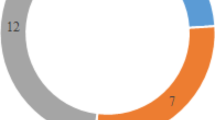
A total of 43 FQ-resistant E. coli isolates were tested in this study and all isolates showed multi-drug resistance (MDR) and mutations in quinolone resistance determining regions (gyrA or parC). Especially, FQ-resistant E. coli isolates with double mutations in both gyrA and parC were shown a high FQs minimum inhibitory concentration (≥ 64 mg/L for ciprofloxacin, ≥ 128 mg/L for enrofloxacin, and ≥ 256 mg/L for norfloxacin). Among 43 FQ-resistant E. coli isolates, 12 (27.9%) were showed plasmid-mediated quinolone resistance (PMQR) positive E. coli. Prevalence of PMQR gene, aac(6’)-Ib-cr, qnrS, and qepA, were identified in 7, 3, and 2 E. coli isolates, respectively. We identified the following in PMQR-positive E. coli isolates: the tetracycline resistance genes tetD (12 isolates, 100.0%), tetE (12 isolates, 100.0%), tetA (11 isolates, 91.7%), and tetB (1 isolate, 8.3%); β-lactamases–encoding blaCMY-2 (10 isolates, 83.3%), blaTEM-1 (7 isolates, 58.3%), blaOXA-1 (7 isolates, 58.3%), blaSHV-1 (3 isolates, 16.7%), and blaAAC-2 (1 isolate, 8.3%); and the chloramphenicol resistance genes (10 isolates, 83.3%); the sulfonamide resistance genes sul1 (9 isolates, 75.0%) and sul2 (10 isolates, 83.3%); the aminoglycoside modifying enzyme gene aac(3)-II (2 isolates, 16.7%). The F4 (7 isolates, 58.3%), LT:STb:EAST1 (5 isolates, 41.7%), and paa (3 isolates, 25.0%) were most common fimbrial antigen, combinations of toxin genes, and non-fimbrial adhesins genes, respectively. All PMQR-positive E. coli carried class I integrons but only 4 isolates carried the gene cassette. The most prevalent plasmid replicon was FIB (9 isolates, 75.0%), followed by FIC, HI1, and N (7 isolates, 58.3%), respectively.
Conclusions
Because FQ-resistant E. coli can serve as a reservoir of FQ resistant genetic determinants that can be transferred to pathogenic bacteria in humans or pigs, this represents a public health hazard.
Similar content being viewed by others


Background
Colibacillosis caused by Escherichia coli (E. coli) in pigs is the most frequent enteric disease and an important cause of death in suckling piglets [1]. This disease may result in significant economic losses in the pig industry because of mortality, decreased weight gain, and costs for treatments, vaccinations and feed supplements [2]. The use of antimicrobial drugs such as β-lactams, aminoglycosides, and fluoroquinolones (FQs) has remained the primary option for controlling colibacillosis.
FQs are broad-spectrum antimicrobials agents and have been used for the treatment of various infections caused by E. coli or other gram-negative bacteria. The World Health Organization (WHO) has classified FQs as ‘‘critically important antimicrobials’’ because of their clinical importance in both human and animal medicine [3]. Because the importance of FQs in humans and animals is increasing, FQ-resistant bacteria are a major concern in the treatment of infectious diseases. Notably, FQ-resistant E. coli have emerged in Korea after enrofloxacin was licensed to be used for veterinary purposes [4].
Multiple mechanisms are involved in resistance to FQ in Enterobacteriaceae. The major mechanisms of resistance to FQ involve mutations of chromosomal genes encoding DNA gyrase and/or topoisomerase IV [5, 6]. In addition, three different plasmid-mediated quinolone resistance (PMQR) determinants have been described: qnr-mediated inhibition of quinolone binding to DNA [7, 8], qepA encoded efflux pump [9], and the aac(6’)-Ib-cr mediated FQ acetylation [10, 11]. The PMQR genes confer only low-level resistance to FQ; however, they can be spread horizontally among enterobacteria and facilitate the selection of resistant mutants following exposure to FQ [12]. Although studies from several countries have documented the prevalence and characteristics of FQ-resistance in healthy pigs [13,14,15], there is still limited information regarding the molecular characteristics of FQ-resistant and PMQR-positive E. coli isolated from suckling piglets with diarrhea. Therefore, the purpose of this study was to investigate the molecular characteristics of FQ-resistant E. coli isolates from suckling piglets with colibacillosis.
Materials and methods
Bacterial isolates
E. coli isolates collected from each colibacillosis clinical case in suckling piglets from 2007–2018 were tested. The farms consisted of 42 different pig herds (50 to 100 sows per each herd). The aseptically collected intestinal contents and feces were inoculated on MacConkey agar(BD Biosciences, Sparks, MD) containing 4 mg/mL of ciprofloxacin (CIP; Sigma-Aldrich, St.Louis, MO). Subsequently, suspected E. coli colonies were identified by VITEK II system (bioMéreiux, France). Thus, a total of 43 CIP-resistant E. coli were tested in this study.
Antimicrobial susceptibility testing
The disk diffusion method was performed according to the Clinical and Laboratory Standards Institute (CLSI) guidelines [16]. The 19 antimicrobial disks (BD Biosciences) used in this study were amikacin (30 μg), amoxicillin-clavulanate (20/10 μg), ampicillin (10 μg), aztreonam (30 μg), cefazolin (30 μg), cephalothin (30 μg), cefoxitin (30 μg), cefepime (30 μg), chloramphenicol (30 μg), colistin (10 μg), doxycycline (30 μg), gentamicin (10 μg), kanamycin (30 μg), nalidixic acid (30 μg), neomycin (30 μg), penicillin (10 U), streptomycin (10 μg), tetracycline (30 μg), and trimethoprim-sulfamethoxazole (1.25/23.75 μg). The minimum inhibitory concentration (MIC) for CIP, enrofloxacin (ENR), and norfloxacin (NOR) was determined by standard agar dilution methods using the Mueller–Hinton agar (BD Biosciences) method according to the guidelines of the CLSI [16]. The breakpoints of CIP, NOR, and ENR were also determined according to the guidelines of the CLSI [16]. E. coli ATCC 25,922 was included as a quality control. Multidrug-resistance (MDR) was defined as acquired resistance to at least one agent in three or more antimicrobial classes [17].
O-Serogroup typing
O-serogroup typing was performed using rabbit antisera purchased from SSI (Serum Staten Institute, Denmark) with the slide agglutination technique of the Animal and Plant Quarantine Agency (Gimcheon, Korea). A standard strain was obtained from Dr. J.M. Fairbrother (OIE E. coli Reference Laboratory, Quebec, Canada).
Identification of mutations in quinolone-resistance-determining regions and detection of plasmid-mediated quinolone-resistance genes
PCR was carried out to amplify quinolone resistance determining regions (QRDRs) of the target genes (gyrA, and parC) to identify mutations in 43 FQ-resistant E. coli isolates using primers and conditions described previously [18, 19]. The PCR products were purified using GFX PCR DNA and the gel band purification kit (Amersham Bioscience, Freiburg, Germany), and sequenced by an automatic sequencer (Cosmogenetech, Seoul, Korea). The sequences were aligned with those in the GenBank nucleotide database using the Basic Local Alignment Search Tool (BLAST) program available through the National Center for Biotechnology Information website (http://www.ncbi.nlm.nih.gov/BLAST). PMQR genes (qnrA, qnrB, qnrC, qnrD, qnrS, aac(6’)-Ib-cr, and qepA) were detected by PCR amplification and sequencing analysis, as described in Table S1.
PCR amplification and sequencing of antimicrobial resistance and virulence genes from PMQR-positive E. coli
For detection of antimicrobial resistance genes and virulence genes, PCR was performed using DNA extracted from PMQR-positive E. coli isolates. Primers used in the study are listed in Table S1. All PMQR-positive E. coli isolates were tested for resistance genes related to aminoglycosides (aac (3)-II, and ant(2’’)-I), β-lactam antimicrobials (blaCTX-M families, blaTEM, blaSHV, blaOXA, and pAmpC), chloramphenicols (cmlA and catA1), sulfonamides (sul1 and sul2), and tetracyclines (tetA, tetB, tetC, tetD, tetE and tetG). The virulence factor genes associated with the toxins (LT, STa, STb, Stx2e, and EAST-1), fimbriae (F4, F5, F6, F18, and F41), and non-fimbrial adhesins (AIDA-1, paa, eae) were also confirmed by PCR as previously described [20].
Plasmid replicon typing and detection of integrons and gene cassettes
For plasmid replicon typing and detection of integrons and gene cassettes, PCR was performed using DNA extracted from PMQR-positive E. coli isolates. The primers used in this study targeted 18 different replicons [21] and class 1 and 2 integrons (Table S1.). Gene cassettes were tested for integron-positive isolates. The PCR products of the gene cassettes were sequenced as described above.
Conjugation assay
To determine the transferability of PMQR and other genes, conjugation assays were performed using the broth mating method, with E. coli J53 used as the recipient as previously described [22]. Transconjugants were selected on MacConkey agar (BD Biosciences) plates containing sodium azide (100 µg/ml; Sigma-Aldrich) and ampicillin or tetracycline (100 μg/ml; Sigma-Aldrich). Transferability was confirmed by antimicrobial susceptibility tests and PCR for molecular analysis as described above.
Results
Antimicrobial resistance
MDR patterns of FQ-resistant E. coli isolates from colibacillosis are shown in Table 1. Among the 43 FQ-resistant E. coli, all isolates showed MDR against 6 to 11 classes of antimicrobial agents. The rates of resistance to the various antimicrobial classes were as follows: aminoglycosides (43/43, 100.0%), penicillins (43/43, 100.0%), quinolones (43/43, 100.0%), tetracyclines (42/43, 97.7%), phenicols (39/43, 90.7%), β-lactam/β-lactamase inhibitor combinations (32/43, 74.4%), folate pathway inhibitors (32/43, 74.4%), cephems (31/43, 72.1%), monobactams (7/43, 16.3%), and polypeptides (6/43, 14.0%). The rate of resistance to 9 antimicrobial classes was the highest at 39.5%, and one (2.3%) FQ-resistant E. coli isolate was identified resistance to 11 antimicrobial classes.
Molecular characteristics of FQ-resistant E. coli
The molecular characteristics of 43 FQ-resistant E. coli isolates are shown in Table 2. Among the isolates, 36 isolates were classified into 17 O-serogroups, and 6 isolates were ungrouped. The most common serogroup was O149 (12 isolates, 27.9%). The gyrA amino acid substitutions were S83L (43 isolates), D87N (22 isolates), D87G (5 isolates), and D87E (2 isolates), and the parC mutations were S80I (23 isolates), S80R (9 isolates), E84A (5 isolates), S80K (3 isolates), S80N (1 isolates), S80W (1 isolates), E84G (1 isolates), A56C (1 isolates), and S57Q (1 isolates). The MIC ranges for CIP, ENR, and NOR were 4–256 mg/mL, 8–512 mg/ml, and 8–512 mg/mL, respectively, and isolates with double mutations in gyrA were relatively higher than those of other isolates with single mutations in gyrA. In particular, FQ-resistant E. coli isolates with a high level of MICs range (≥ 64 mg/L for CIP, ≥ 128 mg/L for ENR, and ≥ 256 mg/L for NOR) were shown to carry double mutations in gyrA in combination with double mutations in parC. PMQR genes were detected in 12 (27.9%) of the 43 FQ-resistant E. coli isolates. The aac(6’)-Ib-cr, qnrS, and qepA genes were identified in seven, three, and two FQ-resistant E. coli isolates, respectively. Among 12 PMQR-positive E. coli isolates, one isolate that showed the highest MICs for CIP (256 mg/mL), ENR (512 mg/mL), and NOR (512 mg/mL), also carried the PMQR gene aac(6’)-Ib-cr.
Characterization of PMQR-positive E. coli
The prevalence of antimicrobial resistance genes is shown in Table 3. All PMQR-positive E. coli isolates carried the following β-lactamase encoding genes: blaCMY-2 (10 isolates, 83.3%), blaTEM-1 (7 isolates, 58.3%), blaOXA-1 (7 isolates, 58.3%), blaSHV-1 (3 isolates, 16.7%), and blaAAC-2 (1 isolate, 8.3%). Tetracycline-resistance genes were detected in all PMQR-positive E. coli isolates as follows: tetD (12 isolates, 100.0%), tetE (12 isolates, 100.0%), tetA (11 isolates, 91.7%), and tetB (1 isolate, 8.3%). Two types of aminoglycoside-modifying enzyme genes were examined, but aac(3)-II was found only in 2 (16.7%) PMQR-positive E. coli isolates. The sul1 and sul2 sulfonamide-resistance genes were detected in 9 isolates (75.0%) and 10 isolates (83.3%), respectively. The cmlA chloramphenicol-resistance gene was found in 10 isolates (83.3%). Distributions of the virotypes are shown in Table 3. Seven (58.3%) and two (16.7%) of the isolates contained fimbrial antigen F4 and F41, respectively. The most prevalent combinations of toxin genes were LT:STb:EAST1 (5 isolates, 41.7%), followed by EAST1 (3 isolates, 25.0%), LT:STa:STb (1 isolates, 8.3%), STa:STb:EAST1 (1 isolates, 8.3%), STa:STb (1 isolates, 8.3%), Sta (1 isolates, 8.3%). The paa and AIDA non-fimbrial adhesins genes were detected in 3 isolates (25.0%) and 2 isolates (16.7%), respectively. Among 12 PMQR-positive FQ-resistant E. coli isolates, all isolates were found to have class 1 integrons. Class 1 integrons contained four types of gene cassette arrangements, aadA1-dfrA1 (2 isolates), aadA1-aadA2- aadB (1 isolate), and aadA1-aadA2- aadB-cmlA6 (1 isolate). Eight isolates did not carry any of the gene cassettes. A total of 10 plasmid replicon types were identified in all 12 PMQR-positive E. coli isolates. The most common plasmid replicon was FIB (9 isolates, 75.0%), followed by FIC, HI1, and N (7 isolates, 58.3%), respectively. Transferability was only identified in 7 isolates among 12 PMQR-positive FQ-resistant E. coli isolates.
Discussion
Suckling piglets are vulnerable to colibacillosis for many reasons such as changes in environmental conditions, a decline in maternal antibody titers, and various stresses. Antimicrobials are used in intensive pig production systems to control infectious diseases. In particular, FQs are highly effective antimicrobial class with many advantages including high oral absorption, large volume of distribution, and broad-spectrum antimicrobial activity [23]. However, increasing use of these agents has led to rising rates of resistance to FQs in E. coli worldwide [24]; thus, the probability of treatment failure may be increased [25]. In this study, all FQ-resistant E. coli isolates showed were MDR (resistance to more than six antimicrobial agents) with high levels of resistance to several antimicrobials: aminoglycosides (100.0%), penicillins (100.0%), tetracyclines (97.7%), and phenicols (90.7%). In particular, five isolates showed resistance to more than 10 classes. These results were consistent with previous studies showing co-association of resistance to other classes of antimicrobials and high MDR rates among FQ-resistant E. coli [26, 27]. This occurs because PMQR genes and other antimicrobial-resistance genes are linked together on the plasmid [26].
The O-serogroup is considered one of the major virulence factors of E. coli and variety of O-serogroups has been associated with diarrhea [28,29,30]. In the present study, 17 O-serogroups were detected and O149 was most common. The O149 serogroup has been determined to be the dominant serogroup in cases of diarrhea in many countries including China and Canada [31, 32]. In addition, in this study, all isolates (100%) had gyrA mutations, and 29 isolates (67.4%) were double amino acid substitutions (S83L plus substitution in aspartic acid 87). These results were consistent with previous studies showing that DNA gyrase is the primary target of FQ in gram-negative bacteria, and gyrA mutations are dominant mutations in E. coli [33]. Moreover, 37 (86.0%) FQ-resistant E. coli isolates had mutations at codon 80 in parC in the QRDRs, and the most common type of amino-acid substitution was S80I in parC. Previous studies reported that the substitution S80I was most frequently observed among substitutions in the QRDR of parC [34, 35]. These results are consistent with those observed in E. coli isolates from humans [36, 37]. Also, the MICs ranges of isolates with double mutations in gyrA were relatively higher than those of other isolates with single mutations in gyrA. Notably, FQ-resistant E. coli isolates carrying double mutations in gyrA in combination with double mutations in parC were identified at high levels of MICs (≥ 64 mg/L for CIP, ≥ 128 mg/L for ENR, and ≥ 256 mg/L for NOR). These results are consistent with those of recent studies showing that the total number of point mutations in QRDR was positively correlated with the increased MIC.
In this study, 12 FQ-resistant E. coli isolates carried three types of PMQR genes, aac(6’)-Ib-cr (7 isolates), qnrS (3 isolates), and qepA (2 isolates). These PMQR variants have been previously detected in E. coli from livestock, including in healthy animals and retail meats in the Czech Republic [38], China [39], and the United States [40], as well as from swine in Korea [14]. Also, to acquire FQ resistance, bacteria usually require at least double mutations in QRDRs [41]. However, in this study, two isolates (4 mg/L for CIP, 8 mg/L for ENR, and 8 mg/L for NOR) had only a single mutation in gyrA and harbored qnrS in its plasmid. This result showed that PMQR genes play a role in FQ resistance [42].
The rise of antimicrobial resistance is thought to be closely associated with the widespread transfer of resistance genes between bacterial species. In this study, all 12 PMQR-positive E. coli isolates carried a variety of antimicrobial resistance genes such as blaCMY, blaTEM, blaOXA, blaSHV, blaAAC, tetA, tetB, tetD, tetE, aac(3)-II, sul1, sul2, and cmlA and harbored mobile elements such as integrons and gene cassettes at the same time. The bla genes hydrolyze the characteristic β-lactam ring and confer resistance to most β-lactam antimicrobials, including cephalosporins [43]. Previous studies reported that the PMQR genes in bla positive-E. coli were detected at high levels [44]. The presence of the PMQR genes may be significantly associated with the β-lactamase gene, perhaps due to common carriage on a plasmid in Enterobacteriaceae [45]. Also, all 12 PMQR-positive E. coli isolates harbored class 1 integrons and four isolates also contained gene cassettes aadA or dfrA or both genes. These genes are frequently detected in class 1 integrons isolated from animals and humans in Korea [46]. Therefore, integrons in PMQR-positive E. coli isolates from suckling piglets may have acquired the genetic determinants of drug resistance, which could become a concern.
Plasmids are small DNA molecules that are distinct from chromosomes and can provide beneficial effects to bacteria such as antibiotic resistance through horizontal gene transfer [21]. In this study, 10 plasmid replicon types were identified in all 12 PMQR-positive E. coli isolates. The most common plasmid replicon was IncF plasmids including FIB, and FIC. IncF plasmids were associated with the important role in the spread of virulence and resistance to important classes of antimicrobials including quinolones, β-lactams, TEs, sulfonamides, chloramphenicol, and aminoglycosides among Enterobacteriaceae [47].
The detection of E. coli virulence factors is important for diagnosing and establishing preventative measures for colibacillosis [48]. In this study, toxin genes LT, STb, and Sta were detected in nine (75.0%), nine (75.0%), and one (8.3%) PMQR-positive E. coli isolates, respectively. These LT, STa, and STb genes damage the vessel and cause edema and a high mortality in pigs [49]. Also, the most prevalent fimbriae antigen was F4 (7 isolates, 58.3%). F4 fimbriae have been frequently detected in piglets in several countries such as Japan, Europe, and the United States [50,51,52]. Previous studies reported that paa is known to have a high association with F4 [50, 53]. In this study, 6 (50.0%) PMQR-positive E. coli isolates were identified as having the paa gene and coexisting with the F4 gene. Although the specific role of the paa gene in the development of pathogenic E. coli has not yet been clearly defined, various virotypes may also appear as a result of horizontal gene transferability of the paa gene [54].
Conclusions
This study investigated the molecular characteristics of FQ-resistant E. coli isolated from suckling piglets with colibacillosis. All FQ-resistant E. coli isolates showed an MDR phenotype, and the most prevalent of the mutations were double point mutations in gyrA and a single mutation in parC. Also, FQ-resistant E. coli isolates with PMQR genes carried various antimicrobial genes and harbored mobile elements and plasmid replicons. Antimicrobial resistance may become a serious problem because many drugs are probably ineffective for the treatment of colibacillosis and resistance elements can be horizontally transferred on pig farms. Also, this represents a public health hazard because FQ-resistant E. coli can serve as a reservoir of FQ resistant genetic determinants that can be transferred to pathogenic bacteria in humans or pigs. These data support the critical need for comprehensive surveillance of antimicrobial resistance on pig farms.
Availability of data and materials
Quinolone resistance determining regions sequences of the E. coli isolates have been deposited in DDBJ database (http://getentry.ddbj.nig.ac.jp/) under the accession numbers LC716475 (SSC-1), LC716476 (SSC-2), LC716477 (SSC-3), LC716478 (SSC-4), LC716479 (SSC-7), LC716480 (SSC-8), LC716481 (SSC-10), LC716482 (SSC-11), LC716483 (SSC-12), LC716484 (SSC-13), LC716485 (SSC-14), LC716486 (SSC-15), LC716487 (SSC-16), LC716488 (SSC-17), LC716489 (SSC-19), LC716490 (SSC-20), LC716491 (SSC-21), LC716492 (SSC-22), LC716493 (SSC-23), LC716494 (SSC-26), LC716495 (SSC-27), LC716496 (SSC-28), LC716497 (SSC-29), LC716498 (SSC-30), LC716499 (SSC-31), LC716500 (SSC-32), LC716501 (SSC-33), LC716502 (SSC-34), LC716503 (SSC-35), LC716504 (SSC-36), LC716505 (SSC-37), LC716506 (SSC-38), LC716507 (SSC-39), LC716508 (SSC-40), LC716509 (SSC-41), LC716510 (SSC-42), LC716511 (SSC-43), LC716512 (SSC-44), LC716513 (SSC-45), LC716514 (SSC-46), LC716515 (SSC-47), LC716516 (SSC-48), and LC716517 (SSC-49).
Abbreviations
- E. coli :
-
Escherichia coli
- FQ:
-
Fluoroquinolone
- CIP:
-
Ciprofloxacin
- PMQR:
-
Plasmid-mediated quinolone resistance
- MIC:
-
Minimum inhibitory concentration
- ENR:
-
Enrofloxacin
- NOR:
-
Norfloxacin
- MDR:
-
Multidrug-resistance
- QRDRs:
-
Quinolone resistance determining regions
References
Luppi A. Swine enteric colibacillosis: diagnosis, therapy and antimicrobial resistance. Porcine Health Manag. 2017;3:16.
Fairbrother JM, Gyles CL. Diseases of Swine Chapter: Colibacillosis. 9th ed. In: Straw BE, Zimmerman JJ, D’ Allaire S, Taylor DJ, editors. Oxford: Wiley Blackwell; 2006. p. 387–95. https://bmcmicrobiol.biomedcentral.com/articles/10.1186/s12866-022-02604-z.
WHO. Critically Important Antimicrobials for Human Medicine – 5th Rev. Geneva: World Health Organization; 2017. Available online: http://apps.who.int/iris/bitstream/handle/10665/255027/9789241512220-eng.pdf?sequence=1.
Hong J, Kim JM, Jung WK, Kim SH, Bae W, Koo HC, et al. Prevalence and antibiotic resistance of campylobacter spp. isolated from chicken meat, pork, and beef in Korea, from 2001 to 2006. J Food Prot. 2007;70:860–6.
Yoshida H, Bogaki M, Nakamura M, Nakamura S. Quinolone resistance determining region in the DNA gyrase gyrA gene of Escherichia coli. Antimicrob Agents Chemother. 1990;34:1271–2.
Heisig P. Genetic evidence for a role of parC mutations in development of high-level fluoroquinolone resistance in Escherichia coli. Antimicrob Agents Chemother. 1996;40:879–85.
Tran JH, Jacoby GA. Mechanism of plasmid mediated quinolone resistance. PNAS. 2002;99:5638–42.
Poirel L, Rodriguez-Martinez JM, Mammeri H, Liard A, Nordmann P. Origin of plasmid-mediated quinolone resistance determinant QnrA. Antimicrob Agents Chemother. 2005;49:3523–5.
Yamane K, Wachino JI, Suzuki S, Kimura K, Shibata N, Kato H, et al. New plasmid-mediated fluoroquinolone efflux pump, QepA, found in an Escherichia coli clinical isolate. Antimicrob Agents Chemother. 2007;51:3354–60.
Park CH, Robicsek A, Jacoby GA, Sahm D, Hooper DC. Prevalence in the United States of aac (6’)-Ib-cr encoding a ciprofloxacin-modifying enzyme. Antimicrob Agents Chemother. 2006;50:3953–5.
Robicsek A, Strahilevitz J, Jacoby GA, Macielag M, Abbanat D, Park CH, et al. Fluoroquinolone-modifying enzyme: a new adaptation of a common aminoglycoside acetyltransferase. Nat Med. 2006;12:83–8.
Robicsek A, Jacoby GA, Hooper DC. The worldwide emergence of plasmid-mediated quinolone resistance. Lancet Infect Dis. 2006;6:629–40.
Kuo H, Chou C, Tu C, Gong S, Han C, Liao J, et al. Characterization of plasmid-mediated quinolone resistance by the qnrS gene in Escherichia coli isolated from healthy chickens and pigs. Vet Med. 2009;54:473–82.
Hu YS, Shin S, Park YH, Park KT. Prevalence and Mechanism of Fluoroquinolone Resistance in Escherichia coli Isolated from Swine Feces in Korea. J Food Prot. 2017;80:1145–51.
Cheng P, Yang Y, Li F, Li X, Liu H, Fazilani SA, et al. The prevalence and mechanism of fluoroquinolone resistance in Escherichia coli isolated from swine farms in China. BMC Vet Res. 2020;16:258.
CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals (VET01-S3). 3rd ed. Clinical and Laboratory Standards Institute, Wayne, PA. 2015.
Magiorakos AP, Srinivasan A, Carey RB, Carmeli Y, Falagas ME, Giske CG, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18:268–81.
Dutta S, Kawamura Y, Ezaki T, Nair GB, Iida K, Yoshida S. Alteration in the GyrA subunit of DNA gyrase and the ParC subunit of topoisomerase IV in quinolone-resistant Shigella dysenteriae serotype 1 clinical isolates from Kolkata India. Antimicrob Agents Chemother. 2005;49:1660–1.
Bai H, Du JF, Hu M, Qi J, Cai YN, Niu WW, et al. Analysis of mechanisms of resistance and tolerance of Escherichia coli to enrofloxacin. Ann Microbiol. 2012;62:293–8.
Do KH, Byun JW, Lee WK. Prevalence of O-serogroups, virulence genes, and F18 antigenic variants in Escherichia coli isolated from weaned piglets with diarrhea in Korea during 2008–2016. J Vet Sci. 2019;20:43–50.
Johnson TJ, Wannemuehler YM, Johnson SJ, Logue CM, White DG, Doetkott C, et al. Plasmid replicon typing of commensal and pathogenic Escherichia coli isolates. Appl Environ Microbiol. 2007;73:1976–83.
Tamang MD, Nam HM, Jang GC, Kim SR, Chae MH, Jung SC, et al. Molecular characterization of extended-spectrum-b-lactamase-producing and plasmid-mediated AmpC b-lactamase-producing Escherichia coli isolated from stray dogs in South Korea. Antimicrob Agents Chemother. 2012;56:2705–12.
Patel K, Goldman JL. Safety Concerns Surrounding Quinolone Use in Children. J Clin Pharmacol. 2016;56:1060–75.
Threlfall EJ, Cheasty T, Graham A, Rowe B. High-level resistance to ciprofloxacin in Escherichia coli. Lancet. 1997;349:403.
Li Y, Chen L, Wu X, Huo S. Molecular characterization of multidrug-resistant avian pathogenic Escherichia coli isolated from septicemic broilers. Poult Sci. 2015;94:601–11.
Mitra S, Mukherjee S, Naha S, Chattopadhyay P, Dutta S, Basu S. Evaluation of co-transfer of plasmid-mediated fluoroquinolone resistance genes and blaNDM gene in Enterobacteriaceae causing neonatal septicaemia. Antimicrob Resist Infect Control. 2019;8:46.
Seo KW, Lee YJ. Prevalence and Characterization of Plasmid-Mediated Quinolone Resistance Determinants qnr and aac(6’)-Ib-cr in Ciprofloxacin-Resistant Escherichia coli Isolates from Commercial Layer in Korea. J Microbiol Biotechnol. 2020;30:1180–3.
Gyles CL. Fimbriae of Escherichia coli. In: Gyles CL, editor. Escherichia coli in Domestic Animals and Humans. Wallingford.: CAB International; 1994. p. 399–436.
Blanco M, Lazo L, Blanco JE, Dahbi G, Mora A, López C, et al. Serotypes, virulence genes, and PFGE patterns of enteropathogenic Escherichia coli isolated from Cuban pigs with diarrhea. Int Microbiol. 2006;9:53–60.
Sato JPH, Takeuti KL, Andrade MR, Koerich PKV, Tagliari V, Bernardi ML, et al. Virulence profiles of enterotoxigenic Escherichia coli isolated from piglets with post-weaning diarrhea and classification according to fecal consistency. Pesq Vet Bras. 2016;36:253–7.
Noamani BN, Fairbrother JM, Gyles CL. Virulence genes of O149 enterotoxigenic Escherichia coli from outbreaks of postweaning diarrhea in pigs. Vet Microbiol. 2003;97:87–101.
Chen X, Gao S, Jiao X, Liu XF. Prevalence of serogroups and virulence factors of Escherichia coli strains isolated from pigs with postweaning diarrhoea in eastern China. Vet Microbiol. 2004;103:13–20.
Bolon MK. The newer fluoroquinolones. Med Clin N Am. 2011;95:793–817.
Uchida Y, Mochimaru T, Morokuma Y, Kiyosuke M, Fujise M, Eto F, et al. Geographic distribution of fluoroquinolone-resistant Escherichia coli strains in Asia. Int J Antimicrob Agents. 2010;35:387–91.
Yang J, Luo Y, Li J, Ma Y, Hu C, Jin S, et al. Characterization of clinical Escherichia coli isolates from China containing transferable quinolone resistance determinants. J Antimicrob Chemother. 2010;65:453–9.
Vila J, Ruiz J, Marco F, Barcelo A, Goñi P, Giralt E, et al. Association between double mutation in gyrA gene of ciprofloxacin-resistant clinical isolates of Escherichia coli and MICs. Antimicrob Agents Chemother. 1994;38:2477–9.
Domínguez E, Zarazaga M, Sáenz Y, Briñas L, Torres C. Mechanisms of antibiotic resistance in Escherichia coli isolates obtained from healthy children in Spain. Microb Drug Resist. 2002;8:321–7.
Röderova M, Halova D, Papousek I, Dolejska M, Masarikova M, Hanulik V, et al. Characteristics of Quinolone Resistance in Escherichia coli Isolates from Humans, Animals, and the Environment in the Czech Republic. Front Microbiol. 2017;7:2147.
Yu T, Jiang X, Fu K, Liu B, Xu D, Ji S, et al. Detection of extended-Spectrum beta-lactamase and plasmid-mediated quinolone resistance determinants in Escherichia coli isolates from retail meat in China. J Food Sci. 2015;80:M1039–43.
Pereira RV, Foditsch C, Siler JD, Dulièpre SC, Altier C, Garzon A, et al. Genotypic antimicrobial resistance characterization of E. coli from dairy calves at high risk of respiratory disease administered enrofloxacin or tulathromycin. Sci Rep. 2020;10:19327.
Ruiz J. Mechanisms of resistance to quinolones: target alterations, decreased accumulation and DNA gyrase protection. J Antimicrob Chemother. 2003;51:1109–17.
Jacoby GA. Mechanisms of resistance to quinolones. Clin Infect Dis. 2005;41:S120–6.
Paterson DL, Bonomo RA. Extended-spectrum beta-lactamases: a clinical update. Clin Microbiol Rev. 2005;18:657–86.
Seo KW, Lee YJ. Detection of plasmid-mediated quinolone resistance genes in b-lactamase-producing Escherichia coli isolates from layer hens. Poult Sci. 2019;98:1480–7.
Xue G, Li J, Feng Y, Xu W, Li S, Yan C, et al. High prevalence of plasmid-mediated quinolone resistance determinants in Escherichia coli and Klebsiella pneumoniae isolates from pediatric patients in China. Microb Drug Resist. 2017;23:107–14.
Kang HY, Jeong YS, Oh JY, Tae SH, Choi CH, Moon DC, et al. Characterization of antimicrobial resistance and class 1 integrons found in Escherichia coli isolates from humans and animals in Korea. J Antimicrob Chemother. 2005;55:639–44.
Yang QE, Sun J, Li L, Deng H, Liu BT, Fang LX, et al. IncF plasmid diversity in multi-drug resistant Escherichia coli strains from animals in China. Front Microbiol. 2015;6:964.
Fairbrother JM, Nadeau E, Gyles CL. Escherichia coli in postweaning diarrhea in pigs: an update on bacterial types, pathogenesis, and prevention strategies. Anim Health Res Rev. 2005;6:17–39.
Nguyen UV, Coddens A, Melkebeek V, Devriendt B, Goetstouwers T, Poucke MV, et al. High susceptibility prevalence for F4+ and F18+ Escherichia coli in Flemish pigs. Vet Microbiol. 2017;202:52–7.
Zhang W, Zhao M, Ruesch L, Omot A, Francis D. Prevalence of virulence genes in Escherichia coli strains recently isolated from young pigs with diarrhea in the US. Vet Microbiol. 2007;123:145–52.
Kusumoto M, Hikoda Y, Fujii Y, Murata M, Miyoshi H, Ogura Y, et al. Emergence of a multidrug-resistant Shiga toxin-producing enterotoxigenic Escherichia coli lineage in diseased swine in Japan. J Clin Microbiol. 2016;54:1074–81.
Luppi A, Gibellini M, Gin T, Vangroenweghe F, Vandenbroucke V, Bauerfeind R, et al. Prevalence of virulence factors in enterotoxigenic Escherichia coli isolated from pigs with post-weaning diarrhoea in Europe. Porcine Health Manag. 2016;2:20.
Byun JW, Jung BY, Kim HY, Fairbrother JM, Lee MH, Lee WK. O-serogroups, virulence genes of pathogenic Escherichia coli and pulsed-field gel electrophoresis (PFGE) patterns of O149 isolates from diarrhoeic piglets in Korea. Vet Med (Praha). 2013;58:468–76.
Leclerc S, Boerlin P, Gyles C, Dubreuil JD, Mourez M, Fairbrother JM, et al. Paa, originally identified in attaching and effacing escherichia coli, is also associated with enterotoxigenic E. coli. Res Microbiol 2007;158:97–104. https://pubmed.ncbi.nlm.nih.gov/17125971/.
Acknowledgements
Not applicable.
Funding
This work was supported by “Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through Agriculture, Food and Rural Affairs Convergence Technologies Program for Educating Creative Global Leader, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (grant number: 320005–4)”.
This paper was supported by research funds for newly appointed professors of Jeonbuk National University in 2021.
Author information
Authors and Affiliations
Contributions
KWS and WKL, designed the study. KWS and KHD conducted the experiments. KWS and KHD performed the data arrangement. KWS wrote and edited the manuscript. KHD carried out the statistical analysis. KWS, KHD and WKL assumed overall responsibility for the accuracy and integrity of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
There was no need to approve of ethics because all isolates in this study were already isolated from feces or dead carcasses in Korean swine farm and NCCP.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Figure S1-1. LC716475 (SSC-1) blast alignment. Figure S1-2. LC716476 (SSC-2) blast alignment. Figure S1-3. LC716477 (SSC-3) blast alignment. Figure S1-4. LC716478 (SSC-4) blast alignment. Figure S1-5. LC716479 (SSC-7) blast alignment. Figure S1-6. LC716480 (SSC-8) blast alignment. Figure S1-7. LC716481 (SSC-10) blast alignment. Figure S1-8. LC716482 (SSC-11) blast alignment. Figure S1-9. LC716483 (SSC-12) blast alignment. Figure S1-10. LC716484 (SSC-13) blast alignment. Figure S1-11. LC716485 (SSC-14) blast alignment. Figure S1-12. LC716486 (SSC-15) blast alignment. Figure S1-13. LC716487 (SSC-16) blast alignment. Figure S1-14. LC716488 (SSC-17) blast alignment. Figure S1-15. LC716489 (SSC-19) blast alignment. Figure S1-16. LC716490 (SSC-20) blast alignment. Figure S1-17. LC716491 (SSC-21) blast alignment. Figure S1-18. LC716492 (SSC-22) blast alignment. Figure S1-19. LC716493 (SSC-23) blast alignment. Figure S1-20. LC716494 (SSC-26) blast alignment. Figure S1-21. LC716495 (SSC-27) blast alignment. Figure S1-22. LC716496 (SSC-28) blast alignment. Figure S1-23. LC716497 (SSC-29) blast alignment. Figure S1-24. LC716498 (SSC-30) blast alignment. Figure S1-25. LC716499 (SSC-31) blast alignment. Figure S1-26. LC716500 (SSC-32) blast alignment. Figure S1-27. LC716501 (SSC-33) blast alignment. Figure S1-28. LC716502 (SSC-34) blast alignment. Figure S1-29. LC716503 (SSC-35) blast alignment. Figure S1-30. LC716504 (SSC-36) blast alignment. Figure S1-31. LC716505 (SSC-37) blast alignment. Figure S1-32. LC716506 (SSC-38) blast alignment. Figure S1-33. LC716507 (SSC-39) blast alignment. Figure S1-34. LC716508 (SSC-40) blast alignment. Figure S1-35. LC716509 (SSC-41) blast alignment. Figure S1-36. LC716510 (SSC-42) blast alignment. Figure S1-37. LC716511 (SSC-43) blast alignment. Figure S1-38. LC716512 (SSC-44) blast alignment. Figure S1-39. LC716513 (SSC-45) blast alignment. Figure S1-40. LC716514 (SSC-46) blast alignment. Figure S1-41. LC716515 (SSC-47) blast alignment. Figure S1-42. LC716516 (SSC-48) blast alignment. Figure S1-43. LC716517 (SSC-49) blast alignment.
Additional file 2:
Table S1. Primers used for PCR and DNA sequencing.
Additional file 3:
Table S2. History of samples and isolates separated from each sample.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Seo, K., Do, KH. & Lee, WK. Molecular characteristics of fluoroquinolone-resistant Escherichia coli isolated from suckling piglets with colibacillosis. BMC Microbiol 22, 216 (2022). https://doi.org/10.1186/s12866-022-02632-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12866-022-02632-9