
Abstract
Background
Citrobacter strains are opportunistic pathogens often responsible for serious enteric as well as extra-intestinal diseases, and therefore the O-antigenic scheme, still in use in diagnostic identification, should be set for proper serotyping. The structures of more than 30 different Citrobacter O-antigens (O-polysaccharide chains of the lipopolysaccharides) of 43 Citrobacter O-serogroups have been elucidated so far. However, relationships between strains in several heterogeneous serogroups still need to be clarified by immunochemical studies. These include complex serogroups O3 and O8, represented by 20 and 7 strains, respectively, which are the subject of the present work. Earlier, the O-polysaccharide structures have been determined for Citrobacter O3 strain Be35/57 (PCM 1508) and Citrobacter O8 strain Be64/57 (PCM 1536).
Results
Serological studies (immunoblotting) carried out on Citrobacter lipopolysaccharides from different strains ascribed to serogroups O3 and O8 showed that each of these serogroups should be divided into non-cross-reacting subgroups. Based on the results of chemical analyses and 1H and 13C NMR spectroscopy the structure of Citrobacter O-antigens from strains PCM 1504 (O6) and PCM 1573 (O2) have been established. Chemical data combined with serological analyses showed that several Citrobacter strains should be reclassified into other serogroups.
Conclusions
Immunochemical studies carried out on Citrobacter LPS, described in this paper, showed the expediency of reclassification of: 1) strains PCM 1504 and PCM 1573 from serogroups O6 and O2 to serogroups O3 and O8, respectively, 2) strains PCM 1503 and PCM 1505 from serogroups O3 and O8 to new serogroups O3a and O8a, respectively.
Similar content being viewed by others

Background
Bacteria of the genus Citrobacter of the family Enterobacteriaceae are normal inhabitants of animal and human intestinal tract. Some Citrobacter strains are often associated with extraintestinal disorders, among which the most significant are neonatal meningitis and brain abscess [1, 2].
Serotyping and classification of these bacteria is important for diagnostic purposes. The genus Citrobacter is taxonomically most closely related to Salmonella and Escherichia coli. Currently, strains of the genus Citrobacter are classified into 11 genomospecies based on genetic studies [3]. Serological heterogeneity of Citrobacter strains is defined by the structure diversity of the O-antigen [4,5,6], which represents the O-specific polysaccharide chain (OPS) of the cell-surface lipopolysaccharide (LPS). Based on the LPS O-antigens, Citrobacter strains are divided into 43 O-serogroups [7] and 20 chemotypes [5].
Structural analysis of the OPS is crucial for unambiguous assignment of serotypes and their cross-reactivity. This is especially important in the case of Citrobacter where heterogeneity of strains within particular serogroup is observed and after re-classification some serotypes and O-antigens are overlapping in different species. Structures of the OPS and core domains of LPS are an efficient tool in classification of Gram-negative bacteria. Structural analysis of Citrobacter is an example for such approach enabling fine taxonomic classification of these bacteria. Elucidation of structures of over 30 different Citrobacter OPS improved the serological classification of strains and explained multiple cross-reactions between Citrobacter and other genera of the family Enterobacteriaceae, such as Hafnia, Escherichia, Klebsiella, and Salmonella [6, 8] (and refs cited herein). Although many Citrobacter OPS structures have been established, in several heterogeneous serogroups the O-antigens require further immunochemical studies.
The present work is devoted to serogroups O3 and O8, represented by 20 and 7 strains, respectively. The OPS structures have been determined for Citrobacter O3 strain Be35/57 (PCM 1508) [9] and Citrobacter O8 strain Be64/57 (PCM 1536) [10]. Serological studies of number of strains belonging to serogroups O3 and O8 indicated that each of these serogroups can be further divided into two non-cross-reacting subgroups. Moreover, it was shown that two strains from other serogroups, PCM 1504 (O6) and PCM 1573 (O2), should be reclassified into O3 and O8 serogroups, respectively. Two other strains, PCM 1503 and PCM 1505, should be reclassified from serogroups O3 and O8 into new serogroups. These studies prove that the existing O-antigenic scheme used for serotyping, classification and diagnostic purposes needs modifications to make it consistent with the recent taxonomic changes and serological and structural data on the Citrobacter O-antigens.
Methods
Bacterial strains, cultivation, isolation of LPS and OPS
Citrobacter strains listed in Table 1 were derived from the Polish Collection of Microorganisms (PCM) of the L. Hirszfeld Institute of Immunology and Experimental Therapy (Wrocław, Poland). Analysis of bacteria performed by the matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF) with standard procedure using Bruker Daltonics UltrafleXtreme spectrometer and Biotyper 3.1 software proved the classification of all strains to Citrobacter genus. Bacteria were cultivated in a Davis broth medium supplemented with casein hydrolysate and yeast extract (Difco), with aeration at 37 °C for 24 h, then harvested and freeze-dried.
The LPS were isolated from bacterial mass by the phenol-water procedure [11], recovered from water phase and purified as described [12]. The LPS (called LPS I) from two strains (Be35/57 and Be64/57) were isolated by dialysis of the phenol-water extract without separation of layers and purified with cold aq 50% trichloroacetic acid to precipitate proteins and nucleic acid [6]. The yield of the LPS was 1.5–2.5% of dry bacterial mass.
LPS was heated with 1% acetic acid at 100 °C for 1–2 h and the carbohydrate-containing supernatant was fractionated by gel filtration chromatography on a Sephadex G-50 Fine column (100 × 2.0 cm) in 0.05 M pyridinium acetate buffer pH 5.4 to obtain a high-molecular mass O-polysaccharide.
Preparation of sera and immunochemical analysis
Rabbit sera against whole cells of Citrobacter strains of serotype O3 (PCM 1497 and PCM 1508) and O8 (PCM 1533, 1536, 1572, 2539) were prepared as described earlier [13]. Sera against cells of strains PCM 1503, 1505 and PCM 1531 were from previous studies [8, 14, 15]. The animal studies were conducted in strict accordance with the ethical guidelines established by the National Ethics Committee and approved by the First Local Ethics Commission at the Institute of Immunology and Experimental Therapy, Polish Academy of Sciences (LKE 53/2009).
SDS-PAGE of LPS was performed by the method of Laemmli [16]. The gels were stained with the silver reagent [17] or immunoblotted according to [18]. Briefly, after separation in SDS-PAGE, the LPS was transblotted from the gel onto a PVDF membrane (Immobilon P, Millipore). The air-dried membrane was incubated with anti-Citrobacter serum diluted in TBS-T (20 mM Tris-HCl, 50 mM NaCl, 0.05% Tween-20, pH 7) containing 1% BSA, washed with TBS-T and incubated with goat anti-rabbit IgG conjugated with alkaline phosphatase diluted in TBS-T. The immunoblot was visualised with the staining reagent (nitro-blue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate in 0.05 M Tris/HCl pH 9.5 containing 5 mM MgCl2).
Passive hemagglutination assay was performed as described previously [13]. The LPS were at first heated (1 mg/ml PBS, 100 °C, 2 h) or alkali treated (0.25 M NaOH, 56 °C, 1 h) [19]. The sheep erythrocytes (0.2 ml of packed cells) were coated with a suspension of 1 mg LPS/ml PBS at 37 °C for 1 h. The hemagglutination assay was performed with 1% erythrocytes and ten-fold dilutions of serum, all in PBS at 37 °C for 2 h. Results were expressed as the reciprocal titres of the serum dilutions.
Sugar and methylation analysis
A sample of the OPS was hydrolyzed with 2 M TFA (120 °C, 2 h), monosaccharides were converted conventionally into the alditol acetates and analyzed by GLC-MS on a Hewlett-Packard 5971A instrument equipped with an HP-1 capillary glass column (12 m × 0.2 mm) using a temperature program of 150 °C (3 min) to 270 °C at 8 °C min−1. Methylation of the OPS was performed by the method of Gunnarsson [20]. The partially methylated monosaccharides were derived by hydrolysis of the methylated polysaccharide as in sugar analysis or with 10 M HCl (80 °C, 30 min), converted into the alditol acetates and analyzed by GLC-MS as above.
NMR spectroscopy
Polysaccharide samples were freeze-dried twice from a 99.9% D2O solution and dissolved in 99.95% D2O. 1H- and 13C–NMR spectra were recorded at 53 °C on a Bruker DRX-500 spectrometer (Germany) and chemical shifts are reported with internal acetone (δH 2.225, δС 31.45) as reference for calibration. The NMR spectra were recorded and data processed using standard Bruker software.
Results and discussion
Serological studies
The strains ascribed by several authors [4, 21, 22] to serogroups O3 and O8 are listed in Table 1. The names of species are used according to Miki et al. [21]. As the serological identity of several strains was ambiguous we have characterized them immunochemically. The passive hemagglutination test of Citrobacter O8 LPS with rabbit antisera against the whole bacteria revealed high degree of identity of strains PCM 1536 and PCM 1572, whereas the other LPS studied (PCM 2539) was different (Table 2). LPS of strain PCM 1533 showing low cross reactivity with LPS of PCM 1536 and PCM 1572 was considered as an R form, like LPS of strains PCM 1534 and PCM 1535 but their R character was not proved experimentally.
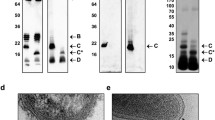
Within the entire serogroup O8 the OPS structure has been determined only for the Citrobacter Be64/57 (PCM 1536) [10], thus this strain is considered as a reference for the entire serogroup O8. In the immunoblotting assay (Fig. 1a, middle panel), the anti-1536 serum reacted with the homologous LPS as well as with LPS of strain PCM 1573, which has been originally ascribed to serogroup O2 [23]. This finding shows the expediency of transfer of Citrobacter PCM 1573 and PCM 1496 into serogroup O8 (Fig. 1a, Table 1). Data on the structure of the OPS of strain PCM 1573 presented below confirmed this conclusion.

SDS-PAGE and immunoblotting experiments for identification of Citrobacter strains in serogroups O8 (panel a) and O3 (panel b). Panel a) Silver-stained SDS-PAGE (left) and immunoblotting with anti-C. braakii-PCM 1536 (middle) and anti-C. youngae PCM 1505 (right) sera of LPS from C. youngae PCM 1505 (lane 1), C. youngae PCM 1573 (lane 2), and C. braakii PCM 1536 (lane 3 – LPS I [6], lane 4 – LPS). Panel b) Silver-stained SDS-PAGE (left) and immunoblotting with anti-C. braakii PCM 1536 (middle) and anti-C. youngae PCM 2539 (right) sera of LPS from C. youngae PCM 1504 (lane 1), C. youngae PCM 1505 (lane 2), C. braakii PCM 1536 (line 3), and C. youngae PCM 2539 (lane 4)
Lipopolysaccharides from strains PCM 1504 and PCM 1505, putative members of serogroup O8 or O6 (Table 1), were not recognized either by anti-1536 (O8) (Fig. 1a and b, middle panels) or anti-1531 (O6) [15] sera. The lack of reactivity of the anti-1505 serum with LPS of strain PCM 1573 (O8) confirms that strain PCM 1505 should not be classified within the serogroup O8 (Fig. 1a, right panel). Moreover, the structural studies revealed the occurrence of D-galactofuranose in the OPS of strain PCM 1505 [14], a component that is not characteristic for serogroup O8. Therefore, Citrobacter PCM 1505 should be assigned to a new serogroup.
The other Citrobacter strain, namely PCM 2539, has been considered as a member of serogroup O8 (Table 1). The serum anti-2539 reacted with the homologous LPS (Table 2) as well as with LPS of strain PCM 1504 (Fig. 1b, right panel). In turn, the LPS of strain PCM 1504 was recognized by anti-1508 and anti-1497 sera (anti O3 sera; Fig. 2). These data allowed to classify both PCM 2539 and PCM 1504 strains as the members of serogroup O3.

SDS-PAGE and immunoblotting experiments for determination the serotype of Citrobacter PCM 1504. Silver-stained SDS-PAGE (a) and immunoblotting with anti-C. youngae PCM 1508 (b) and anti-C. youngae PCM 1497 (c) sera of LPS from C. youngae PCM 1504 (lane 1), C. youngae O3 PCM 1508 (lane 2 – LPS I [6]), C. youngae O3 PCM 1509 (lane 3)
The serogroup O3 also appeared to be complex. The OPS structure has been determined for Citrobacter Be35/57 (PCM 1508) [9] and is considered as a reference structure for the serogroup O3. SDS-PAGE and immunoblotting studies (Fig. 3) showed that the strains of serogroup O3 display diverse pattern and can be divided into several groups. The strains PCM 1497, 1498, 1499, 1511, 1512, 1514, 1516, 1519, 1521, and PCM 1522 were recognized by anti-1508 (O3) serum. Anti-1497 serum reacted additionally with LPS of strains PCM 1503 and PCM 1518 (Fig. 3c). However, anti-1503 serum recognized only LPS of strains PCM 1503 and PCM 1518, indicating that they are serologically different. LPS of the strains PCM 1508, 1509, 1510, 1513, 1517, and PCM 1520 contained low amount of OPS or were already in the R form. In addition, the anti-1503 (not shown) or anti-1508 sera did not recognize the strain PCM 1515 (Fig. 3). The other immunochemical studies on the OPS of Citrobacter PCM 1503 [6, 8] and PCM 1505 [14] suggest that these two strains should be reclassified into new serogroups (Table 3). The structural and serological studies presented below indicated that serogroup O3 should be extended by adding strain PCM 1504, which has been ascribed previously to serogroup O6 or O8. In turn, strain PCM 1573 that has been classified previously to serogroup O2, should be transferred to serogroup O8 (Table 1).

Cross-reactivity of LPS from Citrobacter strains classified in O3 serogroup. Silver-stained SDS-PAGE (a) and immunoblotting with anti-C. youngae PCM 1508 (b) and anti-C. youngae PCM 1497 (c) sera of the indicated Citrobacter LPS
Structural analysis of the O-polysaccharides from Citrobacter PCM 1573 and PCM 1504 strains
In order to confirm the above serological results, the structural studies were performed on the OPS of strains PCM 1509, 1573, and PCM 1504. The corresponding LPS of Citrobacter PCM 1509, 1573 and PCM 1504 strains were recovered in yields 4.6%, 0.4%, and 1.65% of a dry bacterial mass, respectively. In SDS-PAGE, the LPS 1504 and LPS 1573 preparations showed a ladder-like pattern characteristic for LPS of S-type. LPS 1509 preparation in SDS-PAGE experiment has lost its smooth (S) character, therefore it has not been subjected to structural analysis. The mild acid hydrolysis of the LPS followed by fractionation of the carbohydrate material (38%, 59%, and 57% of the LPS weight) on Sephadex G-50 afforded the main fraction P1 (OPS), intermediate fraction P2, core oligosaccharide fraction P3 and Kdo-containing fraction P4. The yields of the OPS and core fractions were 35% and 27.8% for PCM 1504, 53.6% and 22.7% for PCM 1573, and 6.2% and 57.7% for PCM 1509, respectively, of the total material eluted from the column.
The sugar analysis of the OPS of strain PCM 1573 revealed the presence of two components, namely rhamnose and xylose in molar ratio 3.0:0.8 (hydrolysis with 2 M TFA). Methylation analysis showed the presence of terminal xylofuranose, 2-substituted rhamnopyranose, 3-substituted rhamnopyranose and 2,3-disubstituted rhamnopyranose in molar ratios 0.5:0.9:1.0:1.0 (2 M TFA). The 1H NMR and 13C NMR spectra of the OPS-1573 (Fig. 4a, b respectively) were identical to those of OPS from strains PCM 1536 [10] and PCM 1572 (data not shown), representing serogroup O8. Therefore, NMR data confirm the results of the chemical analysis and the expediency of classifying strain PCM 1573 into serogroup O8.

NMR spectra of the isolated OPS. Shown are the 1H NMR spectra (a, c) and 13C NMR spectra (b, d) of the OPS of Citrobacter PCM 1573 (a, b) and PCM 1504 (c, d). Expanded regions of the 1H NMR spectra are shown in the insets
Regarding the strain PCM 1504, sugar analysis of the OPS (hydrolysis with 2 M TFA) indicated the presence of rhamnose and mannose in molar ratio 1.9:1.0. Methylation analysis of the OPS revealed the presence of 4-substituted rhamnopyranose, 3-substituted rhamnopyranose, and 4-substituted mannopyranose residues in molar ratios 0.9:0.8:1.0, respectively. Moreover, the 1H NMR and 13C NMR spectra of the OPS of PCM 1504 strain (Fig. 4c, d, respectively) were identical to the spectra of Citrobacter strain Be35/57 [10] belonging to O3 serogroup. Thus, these data exclude the strain PCM 1504 from the O8 serogroup. The performed serological analysis indicated that the original smooth type LPS of strain PCM 1509 was identical to that of strain PCM 1508 (LPS I, O3) (Fig. 2).
Conclusions
In this paper we report that each of the complex serogroups O3 and O8 can be divided into two non-cross-reacting serogroups. The strains originally ascribed to the serogroups O2 (PCM 1573), O6 (PCM 1504, 1505) and O7 (PCM 1503) that O-antigenic structures are shown in Table 3, should be re-classified into serogroups O3 (PCM 1504) and O8 (PCM 1573). The strains PCM 1503 [8] and PCM 1505 [14] should be classified within new serogroups, namely O3a and O8a, respectively.
Change history
17 August 2017
An erratum to this article has been published.
Abbreviations
- GLC-MS:
-
Gas-liquid chromatography-mass spectrometry
- LPS:
-
Lipopolysaccharide
- NMR:
-
Nuclear magnetic resonance
- OPS:
-
O-polysaccharide
- TFA:
-
Trifluoroacetic acid
References
Doran TI. The role of Citrobacter in clinical disease of children: review. Clin Infect Dis. 1999;28:384–94.
Badger JL, Stins MF, Kim KS. Citrobacter freundii invades and replicates in human brain microvascular endothelial cells. Infect Immun. 1999;67:4208–15.
Brenner D, Grimont P, Steigerwalt A, Fanning G, et al. Classification of citrobacteria by DNA hybridization: designation of Citrobacter farmerii sp.nov., Citrobacter youngae sp.nov., Citrobacter braakii sp.nov., Citrobacter werkmanii sp.nov., Citrobacter sedlakii sp.nov. and three unnamed Citrobacter genomosp. Int J Syst Bacteriol. 1993;43:645–58.
Sedlak J, Slajsova M. On the antigenic relationships of certain Citrobacter and Hafnia cultures. J Gen Microbiol. 1966;43:151–8.
Keleti J, Lüderitz O, Mlynarcik D, Sedlak J. Immunochemical studies on Citrobacter O-antigens (lipopolysaccharides). Eur J Biochem. 1971;20:237–44.
Knirel YA, Kocharova NA, Bystrova OV, Katzenellenbogen E, Gamian A. Structures and serology of the O-specific polysaccharides of bacteria of the genus Citrobacter. Arch Immunol Ther Exp. 2002;50:379–91.
Sedlak J, Slajsova M. Antigen structure and antigenic relationships of the species Citrobacter. Zbl Bakt. 1966;200:369–74.
Kocharova NA, Katzenellenbogen E, Zatonsky GV, Gamian A, et al. Structure of the O-polysaccharide of Citrobacter youngae PCM 1503. Carbohydr Res. 2010;345:2571–3.
Kocharova N, Borisova S, Zatonsky G, Shashkov A, et al. Structure of the O-specific polysaccharide of Citrobacter freundii O3a,3b,1c. Carbohydr Res. 1998;306:331–3.
Kocharova N, Knirel Y, Shashkov A, Kochetkov N, et al. Structure of the Citrobacter freundii O8a,8b O-specific polysaccharide containing D-xylofuranose. Carbohydr Res. 1994;263:327–31.
Westphal O, Jann K. Bacterial lipopolysaccharides: extraction with phenol-water and further applications of the procedure. Methods Carbohydr Chem. 1965;5:83–91.
Romanowska E. Sepharose gel filtration method of purification of lipopolysaccharides. Anal Biochem. 1970;33:383–9.
Gamian A, Romanowska E, Romanowska A. Immunochemical studies on sialic acid-containing lipopolysaccharides from enterobacterial species. FEMS Microbiol Immunol. 1992;89:323–8.
Katzenellenbogen E, Kocharova N, Górska-Frączek S, Gamian A, et al. Structural and serological studies on the O-antigen show that Citrobacter youngae PCM 1505 must be classified to a new Citrobacter O-serogroup. Carbohydr Res. 2012;360:52–5.
Katzenellenbogen E, Kocharova NA, Zatonsky GV, Witkowska D, et al. Structural and serological studies on a new 4-deoxy-D-arabino-hexose-containing O-specific polysaccharide from the lipopolysaccharide of Citrobacter braakii PCM 1531 (serogroup 06). Eur J Biochem. 2003;270:2732–8.
Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–5.
Tsai CM, Frasch CE. A sensitive silver stain for detecting lipopolysaccharides in polyacrylamide gels. Anal Biochem 1982; 119: 115–19.
Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci U S A. 1979;76:4350–4.
Ahamed NM, Radziejewska-Lebrecht J, Widemann C, Mayer H. Reactivity of isolated lipopolysaccharides of Enterobacterial R-mutants with complete or incomplete core structures with lectins. Zentralblatt fur Bakteriol. Mikrobiol und Hyg Ser A-Medical Microbiol Infect Dis Virol Parasitol. 1980;247:468–82.
Gunnarsson A. N- and O-alkylation of glycoconjugates and polysaccharides by solid base in dimethyl sulphoxide/alkyl iodide. Glycoconj J. 1987;4:239–45.
Miki K, Tamura K, Sakazaki R, Kosako Y. Re-speciation of the original reference strains of serovars in the Citrobacter freundii (Bethesda-Ballerup group) antigenic scheme of west and Edwards. Microbiol Immunol. 1996;40:915–21.
Lányi B. Biochemistry and serology of Citrobacter. Methods Microbiol. 1984;15:144–71.
Mieszała M, Lipiński T, Kocharova N, Zatonsky G, et al. The identity of the O-specific polysaccharide structure of Citrobacter strains from serogroups O2, O20 and O25 and immunochemical characterization of C. Youngae PCM 1507 (O2a, 1b) and related strains. FEMS Immunol Med Microbiol. 2003;36:71–6.
Acknowledgements
Authors thank Professor Alexander S. Shashkov for help with NMR spectroscopy.
Funding
The study was financially supported by Wroclaw Centre of Biotechnology from programme The Leading National Research Centre (KNOW) for years 2014–2018 and it covered the cost of publishing.
Availability of data and materials
All data generated or analysed during this study are included in this published article.
Author information
Authors and Affiliations
Contributions
EK, AG designed the study. EK, MS, AKK, SG, MM carried out the laboratory work. EK, MS, NAK, YAK, AKK, SG, MM, AG analyzed the data. EK, MS, AG, AKK wrote the manuscript. All authors read and approved the final version of manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional information
The original version of this article was revised to correct discrepancies in Table 3.
An erratum to this article is available at https://doi.org/10.1186/s12866-017-1082-7.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Katzenellenbogen, E., Staniszewska, M., Kocharova, N.A. et al. Re-classification within the serogroups O3 and O8 of Citrobacter strains. BMC Microbiol 17, 169 (2017). https://doi.org/10.1186/s12866-017-1078-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12866-017-1078-3