
Abstract
Rapeseed (Brassica napus L.), accounts for nearly 16% of vegetable oil, is the world’s second produced oilseed. However, pod shattering has caused significant yield loses in rapeseed production, particularly during mechanical harvesting. The GH28 genes can promote pod shattering by changing the structure of the pod cell wall in Arabidopsis. However, the role of the GH28 gene family in rapeseed was largely unknown. Therefore, a genome-wide comprehensive analysis was conducted to classify the role of GH28 gene family on rapeseed pod shattering. A total of 37 BnaGH28 genes in the rapeseed genome were identified. These BnaGH28s can be divided into five groups (Group A-E), based on phylogenetic and synteny analysis. Protein property, gene structure, conserved motif, cis-acting element, and gene expression profile of BnaGH28 genes in the same group were similar. Specially, the expression level of genes in group A-D was gradually decreased, but increased in group E with the development of silique. Among eleven higher expressed genes in group E, two BnaGH28 genes (BnaA07T0199500ZS and BnaC06T0206500ZS) were significantly regulated by IAA or GA treatment. And the significant effects of BnaA07T0199500ZS variation on pod shattering resistance were also demonstrated in present study. These results could open a new window for insight into the role of BnaGH28 genes on pod shattering resistance in rapeseed.
Similar content being viewed by others


Introduction
Rapeseed (Brassica napus L., AACC, 2n = 38) is the second largest oil crop, accounting for about 16% of the total global vegetable oil production in the world [1]. Rapeseed pods readily dehisce and disperse their seeds at maturity, resulting in yield losses of 15–50% under unfavorable weather or machine harvesting conditions [2]. Therefore, cultivating new cultivars with pod shattering resistance is the most effective approach to reducing the yield losses in rapeseed production [3].
During pod shattering, cell separation in the dehiscence zone (DZ) is thought to caused by pectin degradation [4]. Polygalacturonase (PG), a member of the Glycoside Hydrolase family 28 (GH28), can catalyze the hydrolysis of α-(1–4) galactosidic bond cleavage and D-glucuronic acid bond in pectin molecules, resulting in cell separation [5, 6]. Especially, RABIDOPSIS DEHISCENCE ZONE POLYGALACTURONASE 1 (ADPG1) and ADPG2 were involved in the cell separation during Arabidopsis silique dehiscence [7]. The role of PG in pod shattering was also been reported in pea and soybean [8, 9]. Besides, QUARTET2 (QRT2), another member of the GH28 family, was involved in anther dehiscence and floral organ abscission in Arabidopsis [10]. QRT3 with polygalacturonase activity plays a direct role in the degradation of pollen mother cell wall [11].
In Brassica plants, a large number of studies have been carried out to investigate the mechanism of pod shattering (resistance). So far, eight pod shattering resistance related genes, including SHATTERPROOF1/2 (SHP1/2) [12], FRUITFULL (FUL) [13], INDEHISCENT (IND) [14], ALCATRAZ (ALC) [15], NAC SECONDARY WALL THICKENING PROMOTING FACTOR 1/2 (NST1/2) [16], POLYGlACTOURANAZE (PG) [17], REPLUMLESS (RPL) [18] and SPATULA (SPT) [19] have been reported. However, the effects of pectin and PG on rapeseed pod shattering were largely unknown.
Thus, investigating the role of GH28 on pod shattering in B. napus is of great significance in understanding the pod shattering mechanism. In present study, 37 BnaGH28 genes were identified in the B. napus genome by a homology sequence blast. The protein physicochemical properties, conserved motif, gene structure, cis-acting element, and tissue expression profile analysis were also conducted. Furthermore, Two BnaGH28 genes (BnaA07T0199500ZS and BnaC06T0206500ZS) significantly regulated by IAA or GA treatment were be found in present study. And the significant effects of BnaA07T0199500ZS variation on pod shattering resistance were also demonstrated. These findings could provide key information for developing pod shattering resistant cultivars by genetic approach in B. napus.
Materials and methods
Identification of BnaGH28 genes in the B. napus genome
For the Identification of BnaGH28 genes in the B. napus genome, a blastP analysis was performed in the Brassica napus pan-genome Information Resource (BnPIR) database [20] (http://cbi.hzau.edu.cn/bnapus/index.php) by using ten AtGH28 protein reference sequences from UniProt database [21](https://www.uniprot.org/). And then, BnaGH28s were screened according to the conserved protein structure by using the Simple Modular Architecture Research Tool (SMART) [22] (http://smart.embl.de/smart/set_mode.cgi?NORMAL=1/). For chromosome location analysis, the B. napus genome file, chromosome annotation information, and location information of BnaGH28 genes were downloaded from the BnPIR database. The Gene Location Visualization subroutine of the TBtools software was used to map the chromosome location of BnaGH28 genes.
Evolutionary relationship of the GH28 family
The full-length protein sequences of 10 AtGH28s, and 37 BnaGH28s were obtained from the UniProt database and BnPIR database, respectively. Protein sequences were aligned using the ClustalW program. The neighbor-joining (NJ) phylogenetic tree was constructed by the MEGA software. Bootstrap analysis was conducted with 1000 replications. Then the evolutionary tree was visualized by the online website iTOL [23] ( https://itol.embl.de/ ).
Analysis of collinearity replication relationship of BnaGH28 genes
To analyze the replication events involved in BnaGH28 genes between or within species, the genome and annotation files of A. thaliana and B. napus were obtained from the Ensemble Plants database (https://plants.ensembl.org/index.html) and BnPIR database, respectively. The OneStepMCScanX program of TBtools was used to analyze the collinearity between or within species of BnaGH28 genes.
Physicochemical properties of BnaGH28 proteins
The physicochemical properties including the number of amino acids (AA), molecular weight (MW), isoelectric point (pI), instability index (II), and grand average of hydropathicity (GRAVY) of BnaGH28 proteins were analyzed by the online software Expasy [24] (https://web.expasy.org/protparam/). Subcellular localization (SL) of BnaGH28 proteins was predicted by the online software Wolf PSORT [25] ( https://wolfpsort.hgc.jp/ ).
Conserved motifs and gene structure of the GH28 family
The online analysis software MEME [26] ( https://meme-suite.org/meme/ ) was used to analyze the conserved motifs of all BnaGH28 protein sequences, with the following parameters: the optimum width of motif, 6–50; the number of repetitions, any; the number of motifs, 20; the number of motif occurrences on each sequence is not limited. Then visualization was conducted by the TBtools software.
The rapeseed genome and annotation files were downloaded from the BnPIR database. The BnaGH28 gene structure was visualized by the subroutine Gene Structure View function of TBtools software.
Tissue expression profiles of BnaGH28 genes
In order to investigate the tissue expression profiles of BnaGH28 genes, the gene expression data in the root, stem, leaf, bud, filament, petal, pollen, sepal, cotyledon, seed, and silique of B. napus was obtained from the BnTIR database ( http://yanglab.hzau.edu.cn ).
Cis-acting elements and gene expression respond to phytohormone
For cis-acting elements analysis, the upstream 2000 bp promoter sequence of BnaGH28 genes was extracted from the B. napus genome by the TBtools software. Cis-acting elements in BnaGH28 promoter regions were predicted by the online software Plant CARE [27] (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/).
Primers were designed using qPrimerDB, and the specific primer sequence is shown in Table S1. In our preliminary experiment, 0–45 mg/L gibberellic acid 3 (GA3) or 0-500 mg/L auxin (IAA) were sprayed in rapeseed. Both 15 mg/L GA3 and 500 mg/L IAA could significantly increase pod shattering resistance in rapeseed (Fig. S1). For investigating BnaGH28 gene expression response to phytohormone, 15 mg/L GA3 or 500 mg/L IAA was sprayed in the silique fifteen days after flowering. Pods were collected for gene expression analysis after 7 days.
Variation analysis of BnaGH28
To analyze the effects of BnaGH28 gene variation on pod shattering resistance, the variation of two candidate genes and pod shattering resistance were investigated in a rapeseed micro-core collection. The sequence variation and haplotype information were extracted from our previous study (Table S2) [28]. During an evaluation of pod shattering resistance, a stem strength tester (YYD-1 A, Zhejiang Top Cloud-agri Technology Co., Ltd) was employed in the present study. Ten uniform fresh harvest siliques per accession were used to evaluate pod shattering resistance. The maximum pressure value of the middle 3 cm interval was recorded as the pod shattering force.
Results
Identification of BnaGH28 genes in the B. napus genome
To identify the GH28 family members in B. napus, ten AtGH28 protein sequences were used as query sequences for BLASTP analysis. A total of 37 BnaGH28 genes were identified in the B. napus genome based on transmembrane structure and conversed domain structures (Table S3). Chromosomal location results have shown that 37 BnaGH28 genes were unevenly distributed on 13 chromosomes. There were five BnaGH28 genes located on the chromosome A05, C03, and C04. There was only one BnaGH28 on chromosomes A01, A06, C01, and C08 (Fig. 1). It was interesting that the distribution of BnaGH28 genes on the A or C sub-genome of B. napus was similar. There were 18 and 19 GH28 genes located in the A and C sub-genome, respectively.

Distribution of BnaGH28 genes on Brassica napus chromosomes
Evolutionary relationship of GH28 family
To determine the evolutionary relationships of the GH28 family, an unrooted phylogenetic tree was constructed between 66 GH28s (37 from B. napus, 10 from A. thaliana, 8 from B. rapa, and 11 from B. oleracea) by MEGA software with NJ method. Results indicated that the 66 GH28s were grouped into five groups (Group A, B, C, D, and E) (Fig. 2). Group A contained 6 GH28 members (2 AtGH28s, 4 BnaGH28s), Group B contained 12 GH28 members (1 AtGH28, 2 BnaGH28s, 4 BraGH28s, 5 BolGH28s), Group C contained 10 GH28 members (2 AtGH28s, 8 BnaGH28s), Group D contained 9 GH28 members (1 AtGH28, 7 BnaGH28s, 1 BolGH28), and Group E contained 29 GH28 members (4 AtGH28s, 16 BnaGH28s, 4 BraGH28s, 5 BolGH28s).

Phylogenetic tree of BnaGH28, BraGH28, BolGH28 and AtGH28 proteins
In order to study the replication events of the BnaGH28 gene family in the B. napus genome, the interspecies collinearity of A. thaliana and B. napus and intraspecies collinearity were investigated by the TBtools software (Fig. 3). A large number of orthologous GH28 genes between B. napus and A. thaliana were identified in the present study (Fig. 3b). Within B. napus genome, the GH28 gene has been amplified to a certain extent (Fig. 3a). These indicating that GH28 gene replications might play a very important role in the development of B. napus.

Synteny analysis of GH28 among Arabidopsis thaliana and Brassica napus | (a) Intraspecies collinearity analysis; (b) Interspecies collinearity analysis. The coarse red line in the circle is the collinearity gene of GH28 genes among species, and the fine gray line indicates all the collinearity genes contained among species
Physicochemical properties of BnaGH28 proteins
In order to clarify the basic characteristics of BnaGH28 proteins, the AA, MW, pI, II, GRAVY, and SL of 37 BnaGH28 proteins were analyzed (Table S4). The length of BnaGH28 proteins ranged from 144 to 546 AA, with an average length of 411 AA. Their MW was ranged from 15501.33 (BnaA09T0529500ZS) to 58893.73 Da (BnaC06T0206500ZS). The pI was ranged from 4.68 (BnaA06T0042800ZS) to 9.33 (BnaC03T0372400ZS). And the GRAVY was ranged from − 0.537 (BnaA09T0529500ZS) to 0.255 (BnaA03T0214100ZS). By comparing the physicochemical properties between or within groups, it was found that the physicochemical properties of BnaGH28 were similar within groups, but largely different between different groups. The average pI was greater than 7, BnaGH28 proteins in groups A, D, and E are mostly alkaline. However, proteins in groups B and C are largely acidic with the average theoretical pI < 7. The stable proteins were mainly distributed in group C. Notably, BnaGH28 proteins in groups A and D are mostly hydrophobic; while BnaGH28 proteins in groups B, C, and E are hydrophilic (Table S4).
The SL results of 37 BnaGH28 proteins have shown that there were 17 BnaGH28 proteins located in the nucleus. There were 8, 6, 4, and 2 BnaGH28 proteins located in the extracell, vacuole, chloroplast, and cytoplasm, respectively (Table S4). It was also found that the subcellular localization of BnaGH28 was mostly consistent with its corresponding homologous in A. thaliana. However, BnaC03T0652200ZS and BnaC03T0372400ZS, homologous to an extracellular matrix located protein (AtPGLR1 and AtPGLR2), were located on the nucleus and vacuole in present study.
Conserved motifs and gene structure of BnaGH28
Twenty conserved motifs of 37 BnaGH28 were obtained by using the online software MEME. We found that almost all of the BnaGH28 proteins contained motif 5 (94.59%), indicating that motif 5 is relatively conserved in the GH28 family. However, only 4 BnaGH28 proteins contained motif 14 (Fig. 4a). Three catalytic reaction related amino acid sites (CGPGHGIS, SPNTDGI, and GDDC) and ion interaction related amino acid sites (RIK) were identified in motif 1 and motif 2 (Fig. S2). And almost all 37 BnaGH28 contained these four conserved functional amino acid sites.

Architecture of conserved motifs and gene structures of the GH28 family in Brassica napus | (a) Conserved motifs of BnaGH28. Boxes with different colors represented different conserved motifs; (b) Gene structure of BnaGH28 genes. Exons and introns are presented as filled green round-corner rectangles and thin single lines, respectively
Results of the distribution of conserved motifs in different groups have shown that proteins in the same group have similar motif distribution. It was found that motif 17 and motif 19 mainly existed in the members of group D. BnaGH28 proteins containing motif 14, motif 15 and motif 16 were mainly in group E (Fig. 4a). These indicate that the proteins in different subgroups may have large functional differentiation.
Gene structure analysis was performed to gain a deeper understanding of the GH28 gene expansion in B. napus. By aligning CDS and genomic sequences, we found that the structure and intron number were quite different among BnaGH28 genes. It was shown that BnaC03T0652200ZS is the shortest one, with only 551 bp. And BnaC04T0298700ZS is the longest BnaGH28, with 21,199 bp. Most genes have 4 introns in group D, except for the BnaA03T0214000ZS and BnaA03T0214100ZS genes. The number of introns in BnaA09T0529500ZS was very different from other branches which have 6–9 introns in the group E (Fig. 4b).
Tissue expression profiles of BnaGH28 genes
To explore tissue-specific expression profiles of BnaGH28 genes, the expression data of 9 tissues, including buds (4 mm), sepals, pollen, petals, filaments, cotyledons, roots, seeds (50 d after flowering) and pods (50 d after flowering), were obtained from the BnTIR database. Results have shown that only some BnaGH28 genes in group E were highly expressed in pods (Fig. 5a). To further explore the BnaGH28 gene expression characters during rapeseed silique development, the expression level of genes in silique at different development stages (2–60 days after flowering) were compared. Results indicated that the expression level of almost all BnaGH28 genes in groups A, B, C, and D was decreased with the silique development. However, eleven BnaGH28 genes in group E were highly expressed at the silique maturity stage (40–60 days after flowering) (Fig. 5b). These indicated that 11 BnaGH28 genes might play a role in pod development of B. napus.

Expression profiles of BnaGH28 genes | (a) Tissue expression profiles of BnaGH28 genes; (b) Expression patterns of BnaGH28 genes in developing siliques
Cis-elements and expression respond to phytohormones of BnaGH28
To better understand the transcriptional regulation and potential function of the BnaGH28 genes, cis-elements in the promoter sequences were predicted by the Plant Care database. Results have demonstrated that a large number of multiple stress- or hormone-related cis-elements have been identified in the promoter region of BnaGH28 genes (Fig. 6a). Among these, almost all BnaGH28 genes contained hormone-related cis-acting elements (auxin response elements (TGA elements), abscisic acid responsiveness elements (ABREs), gibberellin response elements (TATC-box), salicylic acid responsiveness (TCA-element), and MeJA-responsiveness (TGACG-motif and TGTCA-motif)). There 43.75% of genes in group E have IAA response elements. Moreover, all genes in groups C or E have the ABA response elements. These indicated that the BnGH28 expression level might be regulated by hormone signaling.

The cis-acting elements prediction of BnaGH28 and their expression level respond to phytohormones | (a) The cis-acting elements distribution of BnaGH28; (b) Expression level respond to phytohormones of BnaA07T0199500ZS; (c) Expression level respond to phytohormones of BnaC06T0206500ZS. GA3 and IAA represent siliques treated by GA3 or IAA for 7 days, respectively
For investigating the expression level of BnaGH28 response to phytohormones, siliques treated by GA or IAA for 7 days were employed in the present study. Only two genes’ expression levels can be detected among these 11-pod development related candidate genes (Fig. 6b-c). Results indicated that the expression levels of BnaA07T0199500ZS and BnaC06T0206500ZS were significantly downregulated by IAA and GA treatment. Thus, GA and IAA treatments might influence pod shattering resistance by regulating BnaGH28 gene expression in B napus.
Effects of BnaGH28 variation on pod shattering resistant
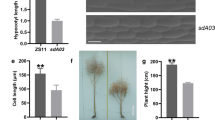
In order to clarify the effects of BnaGH28 gene variation on pod shattering resistance, the variation of 2 pod shattering resistance related candidate genes was investigated in a rapeseed micro-core collection. Results have shown that there were 7 variation sites in BnaA07T0199500ZS (Fig. 7a). However, no available variation (minor allele frequency > 0.05) was detected of BnaC06T0206500ZS in the present population. By comparing the pod shattering force between different variations of BnaA07T0199500ZS, we found that accessions carrying haplotypes 1 (Hap1) exhibited pod shattering resistance than Hap2 (Fig. 7b). These indicated BnaA07T0199500ZS might play vital role in pod shattering resistance in B. napus.

Effects of BnaGH28 gene variation on pod shattering resistance | (a) Variation sites of BnaA07T0199500ZS; (b) Effects of BnaA07T0199500ZS variation on rapeseed pod shatter-resistance
Discussions
Pod shattering, controlled by multiple processes, is a complex and important trait in rapeseed production [29]. The GH28 family including PG has been reported in pod shattering resistance in A. thaliana [30] and grain legumes [31]. In the current study, 37 BnaGH28 genes were identified via genome-wide analysis. This provides a chance to clarify the role of GH28 genes on rapeseed pod shattering resistance.
Members of the same group have a similar protein character, conserved motifs, gene structure, tissue expression profile, and cis-acting elements. These indicated that BnaGH28s might play an important role in pod shattering like its orthologous. AtADPG1/2 were involved in the cell separation during Arabidopsis silique dehiscence; AtQRT2 was involved in anther dehiscence in Arabidopsis; AtQRT3 plays a direct role in the degradation of pollen mother cell wall [32]. However, there were also some homologous genes with different motifs and gene structures. This indicates that these genes in different subgroups may have large functional differentiation [33].
As previous study reported that promoters can regulate plant growth, development, and physiological metabolism by controlling the gene expression at the right time, place, and level [34, 35]. By analyzing the cis-elements in the BnaGH28 family, we found that almost all BnaGH28 genes contained hormone-related cis-acting elements in promoter region. This indicated phytohormone could regulate pod shattering resistance by affecting BnaGH28 gene expression.
Previous report has demonstrated that IAA can influence pod shatter-resistance in Arabidopsis [36]. Relatively higher IAA content in cells can effectively inhibit the activity of endo-1,4-beta-glucanase and PG [37, 38]. Studies on the relationship between exogenous 2-methyl-4-chlorophenoxyacetic acid (4-CPA, an IAA analog) and pod shattering resistance have been conducted [39]. It was also be reported that the pod shattering resistance related genes IND and ALC have been influenced by GA and IAA action [14, 15, 40]. Thus, IAA or GA might play a critical role in pod shattering resistance by regulating the expression level of BnaA07T0199500ZS and BnaC06T0206500ZS in group E. Even the ABA response elements were identified in promoter region of genes in groups E. But, the effects of ABA on rapeseed pod shattering were few reported.
Furthermore, the gene allelic variations have affected pod shattering resistance in A. thaliana and B. napus [41,42,43,44]. In this study, Accessions carrying Hap1 of BnaA07T0199500ZS exhibited higher pod shattering resistance than that of Hap2. This indicates that the pod shattering resistance can be improved by integrating the excellent haplotypes [45]. All these findings provide critical information for investigating the molecular mechanism of GH28 regulating pod shattering resistance in B. napus. And the pod shattering resistant HAP 1 of BnaA07T0199500ZS provide genetic resources for breeding of pod shattering resistant cultivars.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Woodfield HK, Sturtevant D, Borisjuk L, Munz E, Guschina IA, Chapman KD, Harwood JL. Spatial and temporal mapping of key lipid species in Brassica napus Seeds. Plant Physiol. 1998;173(4):1998.
Zhai Y, Cai S, Hu L, Yang Y, Amoo O, Fan C, Zhou Y. CRISPR/Cas9-mediated genome editing reveals differences in the contribution of INDEHISCENT homologues to pod shatter resistance in Brassica napus L. Theor Appl Genet. 2019;132(7):2111–23.
Jia L, Jun W, Hui W, Wenxiang W, Rijin Z, Desheng M, Hongtao C, Juan Y, Harsh R, Qiong H. Multigenic control of pod shattering resistance in Chinese rapeseed germplasm revealed by genome-wide association and linkage analyses. Front Plant Sci. 2016;7:1058.
Liu QH, Stephen J. Physiological and molecular aspects of pod shattering resistance in crops. Czech J Genet Plant Breed 2019, 55(3).
Ting GE, Xue H, Rang-Jin X, University S. Recent advances in polygalacturonase gene in fruit tree species. Plant Physiol J. 2019;55:1075–88.
Yuan C, Zhang S, Hu R, Wei D, Tang Q, Wang Y, Tian S, Niu Y, Wang Z. Comparative transcriptome analysis provides insight into the molecular mechanisms of anther dehiscence in eggplant (Solanum melongena L). Genomics. 2021;113(2):497–506.
Sander L, Child R, Ulvskov, Albrechtsen M, Borkhardt B. Analysis of a dehiscence zone endo-polygalacturonase in oilseed rape (Brassica napus) and Arabidopsis thaliana: evidence for roles in cell separation in dehiscence and abscission zones, and in stylar tissues during pollen tube growth. Plant Mol Biol. 2001;46:469–79.
Dong R, Dong D, Luo D, Zhou Q, Chai X, Zhang J, Xie W, Liu W, Dong Y, Wang Y, et al. Transcriptome analyses reveal candidate pod shattering-Associated genes involved in the pod ventral sutures of common vetch (Vicia sativa L). Front Plant Sci. 2017;8:649.
Wang F, Sun X, Shi X, Zhai H, Tian C, Kong F, Liu B, Yuan X. A Global Analysis of the polygalacturonase gene family in soybean (Glycine max). PLoS ONE. 2016;11(9):e0163012.
Preuss D, Rhee S, Davis R. Tetrad Analysis Possible in Arabidopsis with mutation of the QUARTET (QRT) genes. Sci (New York NY). 1994;264(5164):1458.
Rhee SY, Osborne E, Poindexter PD, Somerville CR. Microspore separation in the quartet 3 mutants of Arabidopsis is impaired by a defect in a developmentally regulated polygalacturonase required for pollen mother cell wall degradation. Am Soc Plant Biologists. 2003;133(3):1170–80.
Jia L, Rijin Z, Wenxiang W, Hui W, Yu Q, Rosy R, Desheng M, Harsh R, Qiong H. A copia-like retrotransposon insertion in the upstream region of the SHATTERPROOF1 gene, BnSHP1.A9, is associated with quantitative variation in pod shattering resistance in oilseed rape. J Exp Bot. 2020;71(18):5402–13.
Peng PF, Li YC, Mei DS, Colasanti J, Fu L, Liu J, Chen YF, Hu Q. Expression divergence of FRUITFULL homeologs enhanced pod shatter resistance in Brassica napus. Genet Mol Res. 2015;14(1):871–85.
Angowski U, Goi O, Quille P, Stephenson P, Carmody N, Feeney E, Barton D, Stergaard L, O’Connell S. A plant biostimulant from the seaweed Ascophyllum nodosum (Sealicit) reduces pod shatter and yield loss in oilseed rape through modulation of IND expression. Sci Rep. 2019;9(1):16644.
Hua S, Shamsi IH, Guo Y, Pak H, Chen M, Shi C, Meng H, Jiang L. Sequence, expression divergence, and complementation of homologous ALCATRAZ loci in Brassica napus. Planta. 2009;230(3):493–503.
Mitsuda N. Masaru Ohme-Takagi: NAC transcription factors NST1 and NST3 regulate pod shattering in a partially redundant manner by promoting secondary wall formation after the establishment of tissue identity. Plant J. 2008;56(5):768–78.
Petersen M, Sander L, Child R, Van Onckelen H, Ulvskov P, Borkhardt B. Isolation and characterisation of a pod dehiscence zone-specific polygalacturonase from Brassica napus. Plant Mol Biol. 1996;31:517–27.
Chu W, Liu J, Cheng H, Li C, Fu L, Wang W, Wang H, Hao M, Mei D, Liu K, et al. A lignified-layer bridge controlled by a single recessive gene is associated with high pod-shatter resistance in Brassica napus L. Crop J. 2022;10(3):638–46.
Raman H, Raman R, Sharma N, Cui X, McVittie B, Qiu Y, Zhang Y, Hu Q, Liu S, Gororo N. Novel quantitative trait loci from an interspecific Brassica rapa derivative improve pod shatter resistance in Brassica napus. Front Plant Sci. 2023;14:1233996.
Song JM, Liu DX, Xie WZ, Yang Z, Guo L, Liu K, Yang QY, Chen LL. BnPIR: Brassica napus pan-genome information resource for 1689 accessions. Plant Biotechnol J. 2021;19(3):412.
Claire OD. Rolf, Apweiler: a guide to UniProt for protein scientists. Methods Mol Biol. 2010;694:25.
Schultz J, Milpetz F, Bork P, Ponting CP. SMART, a simple modular architecture research tool: identification of signaling domains. Proc Natl Acad Sci. 1998;95(11):5857–64.
Letunic I, Bork P. Interactive tree of life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021;49(W1):W293–6.
Gasteiger E, Gattiker A, Hoogland C, Ivanyi I, Appel RD, Bairoch AM. ExPASy: the proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003;31(13):3784–8.
Paul H, Keun-Joon P, Takeshi O, Naoya F, Hajime H, Adams-Collier CJ, Kenta N. WoLF PSORT: protein localization predictor. Nucleic Acids Res. 2007;35(Web Server issue):585–7.
Bailey TL, Boden M, Buske FA, Frith M, Grant CE, Clementi L, Ren J, Li WW, Noble WS. MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res. 2009;37(suppl2):W202–8.
Lescot M, Déhais P, Thijs G, Marchal K, Moreau Y, Van de Peer Y, Rouzé P, Rombauts S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002;30(1):325–7.
Hu J, Chen B, Zhao J, Zhang F, Xie T, Xu K, Gao G, Yan G, Li H, Li L, et al. Genomic selection and genetic architecture of agronomic traits during modern rapeseed breeding. Nat Genet. 2022;54(5):694–704.
Kuai J, Sun Y, Liu T, Zhang P, Zhou M, Wu J, Zhou G. Physiological mechanisms behind differences in pod shattering resistance in rapeseed (Brassica napus L.) varieties. PLoS ONE. 2016;11(6):e0157341.
Jaradat MR, Ruegger M, Bowling A, Butler H, Cutler AJ. A comprehensive transcriptome analysis of silique development and dehiscence in Arabidopsis and Brassica integrating genotypic, interspecies and developmental comparisons. Biotechnol Agric Food Chain. 2014;5(4):302–20.
Parker TA, Sassoum L, Paul G. Pod shattering in grain legumes: emerging genetic and environment-related patterns. Plant Cell. 2021;33(2):179–99.
Ogawa M, Kay P, Wilson S, Swain SM. ARABIDOPSIS DEHISCENCE ZONE POLYGALACTURONASE1 (ADPG1), ADPG2, and QUARTET2 are polygalacturonases required for cell separation during reproductive development in Arabidopsis. The Plant Cell 2009, 21(1): 216–233.
Zhang F, Chen T, Liu N, Hou X, Wang L, Cai Q, Li R, Qian X, Xu H, Zhu Z, et al. Genome-wide characterization of SDR gene family and its potential role in seed dormancy of Brassica napus L. BMC Plant Biol. 2024;24(1):21.
Li Y, Huang H, Shi Y, Huang S, Liu T, Xiao C, Tian X, Zhao P, Dai X, Huang T. Genome-wide identification, evolution and expression analyses of GA2ox Gene Family in Brassica napus L. Phyton-International J Experimental Bot. 2023;92(3):815–35.
Lyu M, Iftikhar J, Guo R, Wu B, Cao J. Patterns of expansion and expression divergence of the polygalacturonase Gene Family in Brassica oleracea. Int J Mol Sci. 2020;21(16):5706.
Sorefan K, Girin T, Liljegren SJ, Ljung K, Robles P, Galvan-Ampudia CS, Offringa R, Friml J, Yanofsky MF, Ostergaard L. A regulated auxin minimum is required for seed dispersal in Arabidopsis. Nature. 2009;459(7246):583–6.
Johnson-Flanagan AM, Spencer MS. Ethylene production during development of mustard (Brassica juncea) and canola (Brassica napus) seed. Plant Physiol. 1994;106(2):601–6.
Dal Degan F, Child R, Svendsen I, Ulvskov P. The cleavable N-terminal domain of plant endopolygalacturonases from clade B may be involved in a regulated secretion mechanism. J Biol Chem. 2001;276(38):35297–304.
Child RD, Ulvskov P, Van Onckelen HA, Botterman J. Strategies for engineering shatter resistance into oilseed rape pods. In Proc. 10th International Rapeseed Congress, Canberra, Australia 1999.
Kay P, Groszmann M, Ross JJ, Parish RW, Swain SM. Modifications of a conserved regulatory network involving INDEHISCENT controls multiple aspects of reproductive tissue development in Arabidopsis. New Phytol. 2013;197(1):73–87.
Raman H, Raman R, Kilian A, Detering F, Carling J, Coombes N, Diffey S, Kadkol G, Edwards D, McCully M, et al. Genome-wide delineation of natural variation for pod shatter resistance in Brassica napus. PLoS ONE. 2014;9(7):e101673.
Liljegren SJ, Ditta GS, Eshed Y, Savidge B, Yanofsky MF. SHATTERPROOF MADS-box genes control seed dispersal in Arabidopsis. Nature. 2000;404(6779):766–70.
Jenkins ES, Paul W, Craze M, Whitelaw CA, Weigand A, Roberts JA. Dehiscence-related expression of an Arabidopsis thaliana gene encoding a polygalacturonase in transgenic plants of Brassica napus. Plant Cell Environ. 2010;22(2):159–67.
Kadkol GP, Halloran GM, MacMillan RH. Evaluation of Brassica genotypes for resistance to shatter. II. Variation in siliqua strength within and between accessions. Euphytica. 1985;34:915–24.
Seo JH, Dhungana SK, Kang BK, Baek IY, Sung JS, Ko JY, Jung CS, Kim KS, Jun TH. Development and validation of SNP and InDel markers for pod-shattering tolerance in soybean. Int J Mol Sci. 2022;23(4):2382.
Funding
This work was supported by the National Key Research and Development Program of China (2022YFD2301401-3), the Open Project of Key Laboratory of Biology and Genetic Improvement of Oil Crops, Ministry of Agriculture and Rural Affairs, P. R. China (KF2021003), the Natural Science Research project of Universities in Anhui Province of China (KJ2021A0198), Young Talents of Anhui Agricultural University (rc312102), and the Special Fund for Anhui Agriculture Research System. The funding bodies had no role in study design, data collection and analysis, interpretation of data, decision to publish, or writing the manuscript.
Author information
Authors and Affiliations
Contributions
K.J. Zhou and F.G Zhang conceived the study. F.G Zhang, N. Liu, T.H Chen and K.J. Zhou designed the experiments. N. Liu, X.Z. Qian and H. Xu identified GH28 members from the Brassica napus genome. N. Liu, T.H Chen, L. Wang, Q.A Cai, R. Li, performed the Bioinformatics analysis and data collection. N. Liu, S. Zhou, and X.Z. Hou carried out the variation analyses. F.G Zhang and N. Liu wrote the paper. Z.H. Zhu, W.Y. Zheng, and Y. Yu participated in the modification of the manuscript. All the authors have read and approved the publication of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, F., Liu, N., Chen, T. et al. Genome-wide identification of GH28 family and insight into its contributions to pod shattering resistance in Brassica napus L.. BMC Genomics 25, 492 (2024). https://doi.org/10.1186/s12864-024-10406-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-024-10406-y