
Abstract
Introduction
The Ovar-DRB1 gene, a crucial element of the Major Histocompatibility Complex (MHC) Class II region, initiates adaptive immunity by presenting antigens to T-cells. Genetic diversity in sheep, particularly in MHC Class II genes like Ovar-DRB1, directly influences the specturm of presented antigens impacting immune responses and disease susceptability. Understanding the allelic diversity of Ovar-DRB1 gene in Sudan Desert Sheep (SDS) is essential for uncovering the genetic basis of immune responses and disease resistance, given the the breeds significance in Sudan's unique environment.
Methods
Utilizing Targeted Next-Generation Sequencing (NGS) we explore allelic diversity in Ovar-DRB1 gene within SDS. Successfully ampliying and and sequencing the second exon of this gene in 288 SDS samples representing six breeds provided a comprehensive allelic profile, enabling a detalied examination of the gene's genetic makeup.
Results
We identifed forty-six alleles, including four previously unreported, enrichness the genetic diversity of SDS breeds. These alleles exhibiting non-uniform distribution, varying frequencies across breeds, indicating a breed-specific genetic landscape. Certain alleles, known and novel, show higher frequencies in specific populations, suggesting potential associations with adaptive immune responses. Identifying these alleles sets the stage for investigating their functional roles and implications for disease resistance. Genetic differentiation among SDS breeds, as indicated by FST values and clustering analyses, highlights a unique genetic makeup shaped by geographic and historical factors. These differentiation patterns among SDS breeds have broader implications for breed conservation and targeted breeding to enhance disease resistance in specific populations.
Conclusion
This study unveils Ovar-DRB1 gene allelic diversity in SDS breeds through targeted NGS and genetic analyses, revealing new alleles that underscore the breeds’ unique genetic profile. Insights into the genetic factors governing immune responses and disease resistance emerge, promising for optimization of breeding strategies for enhanced livestock health in Sudan’s unique environment.
Similar content being viewed by others

Introduction
Major Histocompatibility Complex (MHC) genes play a crucial role in immune responses and disease resistance in various species, including sheep. Among the MHC genes, Ovar-DRB1 has garnered significant attention due to its involvement in antigen presentation and recognition [1, 2].Understanding the genetic diversity and variation within the Ovar-DRB1 gene is essential for breeding programs and conservation efforts in sheep populations [3].
Sudan Desert Sheep (SDS) are a unique breed adapted to the harsh environmental conditions of the Sudanese desert. These sheep possess remarkable traits, such as heat tolerance, drought resistance, infectious diseases tolerance and adaptation to sparse grazing resources, making them well-suited for arid regions. However, the genetic characteristics and diversity of the Ovar-DRB1 gene in SDS remain largely unexplored [4, 5].
To fill this knowledge gap, we conducted a comprehensive study using targeted Next Generation Sequencing (NGS) technology to genotype the Ovar-DRB1 gene in SDS. Targeted NGS allows for cost-effective and efficient genotyping of specific genomic regions, making it a powerful tool for investigating genetic diversity in targeted genes.
In recent studies, the genetic diversity of native sheep breeds reared in Turkey and Algeria has been investigated [6, 7]. These studies aimed to explore the impact of polymorphisms in the MHC gene region, which has the potential to influence future animal breeding strategies. In line with these efforts, [8] conducted Sanger sequencing of the highly polymorphic Exon 2 of the ovine DRB1 gene. Their study focused on six indigenous Turkish sheep breeds and two crossbreeds, aiming to uncover the diversity within this gene. Additionally, [9] described nine novel full-length Ovar-DRB1 sequences and introduced an improved direct-sequencing method for analyzing the entire exon 2 region of the Ovar-DRB1 gene, incorporating previously unknown intronic sequences. These investigations enhance our understanding of MHC gene polymorphisms and their potential significance for the development of f forthcoming breeding initiatives in animal populations.
In this paper, we present the results of our genotyping analysis of the Ovar-DRB1 gene in Sudan Desert Sheep using targeted NGS. We collected DNA samples from a representative population of Sudan Desert Sheep and employed a targeted sequencing approach to specifically capture and sequence the Ovar-DRB1 gene regions of interest. Through this targeted NGS approach, we obtained high-quality DNA sequence data for the Ovar-DRB1 gene in Sudan Desert Sheep. Our analysis focused on identifying genetic variants, assessing allele frequencies, and exploring the population structure and diversity within the Sudan Desert Sheep population [10, 11].
By elucidating the genetic diversity and variation within the Ovar-DRB1 gene, our study provides valuable insights into the adaptive potential and immune responses of Sudan Desert Sheep. This information is crucial for implementing effective breeding strategies, improving disease resistance, and ensuring the long-term health and resilience of this unique breed. Furthermore, the results of our study contribute to the broader understanding of MHC genetics in sheep populations. The targeted NGS approach employed in this research can serve as a valuable tool for future studies investigating the MHC diversity and functional significance in other sheep breeds or related species [12].
In conclusion, our study demonstrates the utility of targeted NGS for genotyping the Ovar-DRB1 gene in Sudan Desert Sheep, shedding light on the genetic diversity and variation within this important MHC gene. The findings of this research have practical implications for breeding programs, conservation efforts, and the broader understanding of MHC genetics in sheep populations.
Materials and methods
Sample population collection and genomic DNA extraction
A total of 288 blood samples were collected from various Sudan desert sheep (SDS) breeds, including Abrag (AB; n = 37), Ashgar (AS; n = 44), Buze´e (B; n = 23), Hamari (H; n = 72), Kabashi (K, n = 45), and Watish (W; n = 38). The samples were obtained from regions located in Al Gadarif, Al Jazirah, Sennar, and North Kurdufan States as illustrated in the map in (Fig. 1), showcasing the geographical diversity of the sampled Sudan desert sheep breeds. Prior consent was obtained from a shepherd, and precautions were taken to avoid sampling closely related individuals. Genomic DNA extraction was performed using the DNeasy® Blood and Tissue Kit (Qiagen, Germany) according to the manufacturer's instructions. This extraction method ensures the isolation of high-quality DNA suitable for downstream analyses.

Geographic Distribution of Sudan Desert Sheep (SDG) Breeds and Blood Sample Collection Sites: This map displays the geographic locations where blood samples were collected from various Sudan Desert Sheep (SDG) breeds across four states. The scale is in kilometers
Ethical approval
The study protocol was reviewed and approved by the Faculty of Veterinary Medicine, University of Khartoum. The research was conducted in accordance with their guidelines for sampling domestic animals in Sudan, ensuring ethical compliance in the collection and use of the samples.
PCR amplification and sequencing
For typing Ovar-DRB1 breeds, we implemented a method based on targeted Next-Generation Sequencing. First, the second exón of this gene were amplified using the primers Illumina_330: TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGNNNNNNATTAGCCTCYCCCCAGGAGKC and Illumina_329: GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGNNNNNNCACCCCCGCGCTCACCTCGCCGC based on a previous study amplifying the region using conventional PCR [13]. The PCR reaction mixture contained 12.5 μl of 2X KAPA HiFi HotStart ReadyMix (Kapa Bio-Systems, Wilmington, MA, USA), 200 nM of each primer and 2.5 μl of the genomic DNA. The reaction was performed at 95 °C for 3 min, followed by 35 cycles of 95 °C for 30 s, 60 °C for 30 s, 72 °C for 30 s and a final extension at 72 °C for 5 min. PCR amplicons were visualized by electrophoresis on 1.5% agarose gel stained with Gel-RedTM (Biotium, Hayward, CA). Illumina sequencing libraries were prepared by purifying the amplicons using AMPure XP (Beckman Coulter Life Sciences, USA) and sequencing adapters and index sequences were added using the Nextera XT Index Kit (Illumina, CA, USA). The sequencing run was conducted with a MiSeq Reagent Kit v3 (600 cycles) on an Illumina MiSeq device according to the manufacturer’s instructions.
Sequence data analysis and Ovar-DRB1 allele genotyping
The quality of obtained Fastq files was evaluated using FastQC software [14] FastQ files were analyzed following the best practice recommended by GATK Germline short variant discovery (SNPs + Indels) (https://gatk.broadinstitute.org/hc/en-us/articles/360035535932-Germline-short-variant-discovery-SNPs-Indels-). Briefly, raw reads were mapped to the ARS-UI_Ramb V2.0 Assembly version of the ovine (Ovis aries) genome (https://www.ncbi.nlm.nih.gov/assembly/GCF_016772045.1/) using BWA-MEM aligner [15] to generate BAM and SAM files. Variants (SNPs and INDELs) per sample were called using HaplotypeCaller to generate GVCF files and consolidated using the joint-call cohort to generate the two VCF files, one for SNPs and other for INDELs. Then, variants were filtered, refined, and annotated to obtain the final VCF file. The SnpEff version 4.3.1 software (https://pcingola.github.io/SnpEff/) Variants and haplotypes were visualized using the IGV software (reference). Heterozygous sites were confirmed when both alleles have balanced numbers of reads. Obtained DNA sequences were compared with previously reported Ovar-DRB1 alleles in the IPD-MHC database to assign the genotypes. Sequences were submitted to the GenBank (https://www.ncbi.nlm.nih.gov/genbank/) under accession numbers from OR488718-OR488752, OR488756, OR488764, OR488770, and OR636395.
Statistical analyses
Genetic diversity at allele level
Allele frequencies and the number of alleles (na) were obtained by direct counting. The distribution of alleles across breeds was analyzed and visualized by a Venn plot created using the R package ‘VennDiagram’ (http://cran.r-project.org/; [16]. The observed (hO) and unbiased expected (hE) heterozygosity of the Ovar-DRB1 locus were estimated according to [17] using the Arlequin 3.5 software for population genetic analyses [18]. FIS statistics for each breed were calculated using the Exact Test proposed by [19] and implemented in Genepop 4.7 software [20] to evaluate deviation from Hardy–Weinberg equilibrium (HWE). The Ewens–Watterson–Slatkin Exact Test of neutrality was performed using the method described by [21] and implemented in the Arlequin 3.5 program.
Genetic structure of the Sudan Desert Sheep
Genetic structure and genetic differentiation among sheep breeds were assessed using Wright’s FST statistics, calculated using the variance-based method of [19]. This parameter was estimated using Arlequin 3.5 and Genepop 4.7 software. The fixation index (FST) values were represented graphically using the pairFstMatrix.r function implemented in the R statistical environment. To condense the genetic variation at the Ovar-DRB1 locus, allele frequencies were used to perform a Principal Component Analysis (PCA) according to [22] implemented in Past software [23]. Nei’s 1972 standard genetic distances [24] Ds and DA [25] were calculated from allele frequencies and were used to perform cluster analysis using the unweighted pair-group method with arithmetic mean (UPGMA;[26] and the neighbor-joining algorithm (NJ; [27]. Confidence intervals for the groupings were estimated by bootstrap resampling of the data using 1000 replicates. Population genetic distances and trees were computed and visualized using the POPTREE2 [28].
Genetic diversity at sequence level
Nucleotide diversity (π) and pairwise differences in nucleotide substitutions between alleles (NPD) within each sheep breed were calculated using Arlequin 3.5. The mean number of nonsynonymous (dN), and synonymous (dS) nucleotide substitutions per site calculated as an average over all sequence pairs were estimated within each group using the modified Nei-Gojobori model [29] and Jukes–Cantor’s formula implemented in the software MEGA X [30].
The possibility that certain codon sites are under diversifying selection within each native Sudan breed was investigated using the Bayesian method implemented using OmegaMap [31]. This method incorporates intragenic recombination and does not assume a known fixed genealogy, so that recombination does not inflate the false detection rate of positive sites. The Ovar-DRB1 allele tree was conducted by using the Maximum Likelihood method and Tamura-Nei model [32] implemented into the MEGA X software. Ovar-DQB1*01:01:01 allele was included as an outgroup.The bootstrap consensus trees were inferred from 1000 replicates. These trees were constructed based on the complete second exon DNA sequences or only the codons that codify the antigen-binding site (ABS) amino acid motifs.
Results
Targeted NGS genotyping method
In the present work, a targeted NGS method was implemented to genotype the Ovar-DRB1 gene. The second exon of this gene was amplified from 288 animals belonging to SDS, and the PCR products were pooled and sequencing in a single run. This analysis allowed us to obtain a full coverage of the second exon (270 bp) that includes the β1 domain with an average depth of 32.61 ± 18.29 in 259 animals, failing only 29 samples. Overlapping of forward and reverse reads allowed to determine the allele phases.
Distribution of Ovar-DRB1 alleles in selected native Sudan Desert sheep breeds
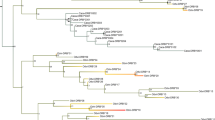
Targeted NGS genotyping allowed us to identify forty-six Ovar-DRB1 alleles (42 previously reported variants and four new alleles; Table 1 and Fig. 2) in the SDS breeds. The number of alleles (na) varied from 17 in B to 33 in H (Tables 1 and 2). The new Ovar-DRB1*Sudan1 was detected in four SDS breeds, whereas the remaining three new alleles were present in two heterozygous animals in Ashgar breed (Ovar-DRB1*Sudan2 and *Sudan3) and one animal in Hamari (Ovar-DRB1*Sudan4). Nucleotide and predicted amino acid sequences of the four new alleles are shown in Fig. 2 and compared with the Ovar-DRB1 DNA sequence of the ARS-UI_Ramb V2.0 Assembly. Figure 3 showed that the variants detected in Sudan desert sheep breeds were interspersed among the various clusters of the Ovar-DRB1 tree, including all the previously reported alleles and the four new alleles. A similar result was observed when the Ovar-DRB1 tree was inferred using codons corresponding to the ABS (Fig. S1).

Alignment of the nucleotide (a) and the predicted amino acid (b) sequences of the β1 domain encoded by four new Ovar-DRB1 alleles (accession numbers: OR488764 for Ovar-DRB1*SDS1, OR488756 for Ovar-DRB1*SDS2, OR488770 for Ovar-DRB1*SDS3, OR636395 for Ovar-DRB1*SDS24) derived from 259 Sudan Desert Sheep (37 animals of the Abrag, 44 of the Ashgar, 23 of the Buze´e, 72 of the Hamari, 45 of the Kabashi, and 38 of the Watish). Numbering refers to amino acid positions in the mature protein. Nucleotide and amino acid residues identical to those encoded by the Ovar-DRB1 sequence from the the ARS-UI_Ramb V2.0 Assembly version of the ovine (Ovis aries) genome are indicated by dots (https://www.ncbi.nlm.nih.gov/assembly/GCF_016772045.1/)

Neighbor-joining (NJ) tree constructed from the 270 bp nucleotide sequence that includes the β1 domain encoded by all reported Ovar-DRB1 alleles and the twenty-six new ones (From Ovar-DRB3*Sudan 1 to Ovar-DRB1*Sudan 26) detected in the Sudan desert sheep breeds. Numbers are bootstrap percentages that support each node. Bootstrapping was carried up with 1000 replicates to assess the reliability of individual branches. AB = Abrag, AS = Ashgar, B = Buze´e, H = Hamari, K = Kabashi, and W = Watish
A Venn diagram was constructed using data obtained in this study to analyze and illustrate the distribution of the forty-six Ovar-DRB1 alleles detected in the SDS sample analyzed. As shown in Fig. 4, this analysis revealed that out of the forty-six alleles identified in SDS, only ten were identified in all breeds, twelve variants were unique for one population, whereas the remaining were detected in from two to four breeds. In addition, variant Ovar-DRB1*17:02 only detected the Buze´e crossbreed population.

Venn plot of Ovar-DRB1 alleles shared by Sudan Desert Sheep breeds: Abrag, Ashgar, Hamari, Kabashi, and Watish breeds
As shown in Fig. S2, the SDS breeds have an even gene frequency distribution, with a high number of alleles with low frequency. Between five to eight alleles appeared with frequencies of > 5% within each population. These common alleles accounted for a cumulative gene frequencies that range from 54.20% in H to 80.43% in K; three (Ovar-DRB1*04:02, *06:03, and *10:01) of which were common in all SDS breeds, one (Ovar-DRB1*13:02) was detected in four out of the five populations, whereas thirty-four variants presented gene frequencies lower than 5% (Table 1, Fig. S3).
Nucleotide and amino acid diversity in the Ovar-DRB1 alleles found in Sudanese Desert Sheep breeds
Genetic diversity at the DNA and amino acid levels was evaluated using methods that.
compare the average amino acid and nucleotide substitutions for every pair of alleles within the breeds studied. Table 3 summarized the results obtained with these methods. The π values exceeded 0.055 (πrange = 0.055 – 0.074) and the mean number of pairwise difference values exceeded 16.73 (NPD range = 14.95 – 19.95) within Sudan Desert Sheep populations. Regarding amino acid diversity, the average dN and dS substitutions in these sheep breeds were calculated across the entire Ovar-DRB1 exon 2 and the ABS. The dN/dS ratio observed in SDS ranged from 0.026 to 0.043 for the entire second exon and from 0.212 to 0.236 when only ABS were considered.
Gene diversity, Hardy–Weinberg Equilibrium (HWE), and neutrality testing of BoLA-DRB3 variants found in Sudanese cattle breeds
As shown in Table 3, na ranged from 17 in B to 33 in H breeds, while he and ho were higher than 0.88 and 0.82, respectively, in all populations; which evidence of the high diversity values for SDS. It is widely accepted that the genetic diversity of MHC class II genes can be maintained by balancing or overdominance selection. For this reason, HWE and Slatkin’s exact neutrality test were carried out to evaluate this phenomenon in SDS populations. However, the HWE test showed that genotype frequencies did not significantly deviate from the theoretical proportion. Furthermore, Slatkin’s exact neutrality test has only evidence of signature of selection compatible with balancing selection in B despite the even gene frequency profile mentioned above (Table 2). The obtained p value in B (p = 0.014) was consistent with the theoretical proportion expected under balancing pressure toward several alleles with low gene frequencies. In addition, we estimated the selection index (ω) in each amino acid site to evaluate the presence of diversifying selection (ω > 1) along Ovar-DRB1 exon 2. These analyses showed high ω values in eleven sites in SDS, mainly located in the ABS (Fig. 5).

Estimated values of the selection index ω in each amino acid site along Ovar-DRB1 exon 2 in Sudan Desert Sheep breeds: Abrag, Ashgar, Buze´e, Hamari, Kabashi, and Watish. Arrows indicate the antigen-binding site (ABS)
Ovar-DRB1 genetic structure and levels of population differentiation in Sudan Desert Sheep
The average FST analysis showed a low but significant level of genetic differentiation for Ovar-DRB1 gene between SDS breeds (FST = 0.010, p < 0.01), and the pairwise FST values varied between 0 (AB—W) to 0.040 (AS—B) (Table S1 and Fig. 6). Significant differences were observed in fourteen out of the fifteen comparisons (p < 0.05). In addtion, Ovar-DRB1 allele frequencies from SDS were used to generate Nei’s DA and DS genetic distance matrices. Then, dendrograms were constructed from these distance matrices using UPGMA and NJ algorithms. All trees revealed congruent topologies, which were consistent with the historical and geographical location of the breeds, revealing two clusters which one included the easter breeds (AB, and W) and the other cluster comprised the western ones (AS, K and H), whereas B crossbreed located in intermediate position and AS diverge from the others breeds (Fig. 7a). In addition, Ovar-DRB1 allele frequencies were used to perform PCA analyses among SDS breeds. The first PC accounted for 51.00% of the data variability, and clearly exhibited a differentiation pattern between eastern (negative values) and western (positive values) breeds, while AS and B were located in opposite extreme positions of the plot (Fig. 7b). The first PC was primarily determined by the contribution of negative values of the Ovar-DRB1*03:03, *03:07, *04:02, *12:02, *13:03, *14:02, and *16:08 alleles, and the positive values of Ovar-DRB1*01:01, *03:11, *06:03, *08:03, *10:01, and *Sudan1. The second PC explained 24.36% of the total variation and contributed to differentiate western and eastern SDS breeds. Finally, the third PC accounted for 11.73% of the variance, discriminating the populations within each cluster. In conclusion, the PCA results agree with the overall clustering observed after NJ or UPGMA tree construction.

Graphic representation of calculated FST between population pairs using an R package pairFstMatrix.r. 1 = Abrag, 2 = Ashgar, 3 = Buze´e, 4 = Hamari, 5 = Kabashi, and 6 = Watish

(a) Neighbor-joining dendrogram constructed from a matrix of DA genetic distances. (b) Principal Component Analysis of allele frequencies from the Ovar-DRB1 gene in Sudan Desert Sheep breeds: Abrag, Ashgar, Buze´e, Hamari, Kabashi, and Watish
In addition, PCA using the gene frequencies of the amino acid motif of the five pockets (pocket 1, pocket 4, pocket 6, pocket 7, and pocket 9) involved in the antigen-binding function of the MHC complex were carried out. In general, the same previously described spatial distributions were obtained, being the exception the pocket 4 (Fig. S4a-e). In addition, the B crossbred population presents a divergence position than other breeds in the PCA plots based on ABS motifs frequencies. PC1 explained from 40.75 to 90.53 of the total variance, while PC2 accounted for 5.41 to 34.94. Figures S4a-e showed the amino acid motifs that mainly contribute to the breed location in these PCAs.
Discussion
The MHC Class II genes play a critical role in the immune response by presenting antigens to T-cells, thereby initiating the adaptive immune response [33]. This process is pivotal for effective pathogen recognition and the subsequent generation of immune responses tailored to combat a wide range of infectious agents [34]. Genetic diversity within MHC II genes, including Ovar-DRB1 in sheep, is particularly important as it directly influences the repertoire of antigens that can be presented to T-cells, enabling a more comprehensive immune response [2, 35, 36]. In sheep, MHC II gene variation has been associated with differences in immune responsiveness and disease susceptibility. Studies have demonstrated that allelic polymorphisms in MHC II genes are linked to variations in immune responses against specific pathogens, including bacteria, viruses, and parasites [37, 38]. The diversity of MHC II alleles contributes to the sheep's ability to recognize and respond to a wide array of pathogens, thereby enhancing disease resistance [39]. Therefore, the breed specific variants identified genetic variation within the Ovar-DRB1 gene significantly impacts immune system function in SDS. Allelic polymorphisms within Ovar-DRB1 affect antigen-MHC II binding, influencing the specificity and efficiency of immune responses [40, 41]. Specific Ovar-DRB1 alleles have been linked to differential resistance or susceptibility to diseases in sheep populations [42, 43], demonstrating the direct link between genetic diversity and immune response efficacy. The identification of breed-specific allelic diversity in the Ovar-DRB1 gene among SDS breeds holds significant implications for disease resilience and adaptation. MHC II genetic variations contribute to unique immune response profiles in different breeds [44]. Breed-specific MHC II alleles have been associated with enhanced disease resistance in various livestock species [45, 46]. The breed-specific variants identified in SDS breeds may confer differential resistance or susceptibility to regional diseases, shedding light on how these breeds have evolved to cope with specific challenges.
Comparative analysis of Ovar-DRB1 alleles in SDS with other sheep populations provides insights into genetic diversity and evolution. MHC II genes often exhibit population-specific allelic distributions due to ecological conditions and selection pressures [44]. Differences in prevalence and distribution of specific MHC II alleles have been documented in different geographic regions [47], suggesting local adaptation influences genetic diversity. Comparisons with other populations enhance understanding of allele distribution, genetic adaptation, and disease resistance strategies. The mean number of differences exceeded 16.73 (NPD range = 14.95 – 19.95) between all pairs of haplotypes in the sample serves as an estimate for the number of mutations that have occurred since the divergence of the haplotypes. This metric is instrumental in gauging genetic variation within populations. Elevated values signify increased genetic diversity, a factor that, in the context of DRB, may facilitate a response to a broad spectrum of pathogens. In comparison to other studies, the diversity observed in Ovar-DRB1 alleles aligns with that in other indigenous breeds [8, 48, 49] but surpasses that in commercially raised breeds within more controlled environments. This disparity can be attributed to natural selection, such as balancing selection in the case of DRB genes, recent gene migration between populations, and stochastic errors introduced by random genetic drift.
The diverse array of Ovar-DRB1 alleles in SDS breeds has implications for genetic conservation and breeding. Understanding immune-related gene diversity is pivotal for safeguarding livestock populations [50]. Breed-specific variants may hold adaptive advantages specific to local environments or diseases, making them valuable candidates for conservation efforts [51]. Breeders can strategically incorporate specific alleles associated with resistance to enhance disease resistance and livestock performance [52]. Harnessing genetic diversity within Ovar-DRB1 optimizes livestock health and productivity, contributing to sustainable and resilient production systems.
The exploration of Ovar-DRB1 diversity in SDS holds implications for enhancing livestock health and resilience. Genetic diversity within immune genes enhances the ability to combat diseases and adapt to changing environments [53]. Breed-specific alleles may offer advantages in adapting to environmental stressors, contributing to robustness [54]. The study's findings emphasize the importance of informed breeding and management practices, contributing to more sustainable and resilient livestock production systems. The study highlights the value of cross-disciplinary collaboration in understanding genetic diversity's impact on health outcomes. Geneticists, immunologists, and livestock managers contribute synergistic insights [55]. Collaboration informs breeding strategies and promotes livestock health, resilience, and sustainability.
While this study has provided valuable insights into the genetic diversity of the Ovar-DRB1 gene in SDS, there are certain limitations that warrant consideration. The sample size, although representative of multiple SDS breeds, may not fully encompass the broader genetic diversity present in these populations. Expanding the sample size to include additional individuals and breeds could provide a more comprehensive view of allelic variation [50]. Another limitation lies in the focus on the second exon of the Ovar-DRB1 gene (Ovar-DRB1.2). Future studies could explore the entirety of the gene or even extend the analysis to other immune-related genes to unravel a more complete picture of the genetic determinants shaping immune responses in SDS [56, 57]. Additionally, the present study has laid the groundwork for understanding allelic diversity, but functional investigations of these alleles are essential. Investigating the associations between specific alleles and immune responses against different pathogens could offer deeper insights into the practical implications of genetic variation [34]. The significance of the four new alleles reported here lies in the fact that functional MHC DRB genes constitute one of the most polymorphic loci in the mammalian genome. The polymorphisms within these genes play a central role in the immune response to pathogens, as the efficiency of binding to antigenic peptides depends on the amino acid motifs present in the pockets of DRB molecules, specifically in exon 2. According to the IPD-MHC database, 130 Ovar-DRB1 alleles have been reported, but their geographic and breed distribution is uneven. Some alleles are widely distributed across different regions and breeds, while others are detected solely in specific regions or closely related breed groups. Indigenous breeds serve as a crucial source of DRB genetic diversity. The geographic distribution of these alleles is a legacy of historical, stochastic (genetic drift), or selective (natural or artificial) factors. Conducting functional or association studies in Sudan Desert Sheep is essential to comprehend the contribution of these newly identified alleles.
Future directions for research could encompass longitudinal studies to monitor changes in allele frequencies over time and in response to evolving disease pressures. Integrating genomic data with phenotypic and environmental data would allow for a more comprehensive assessment of genotype–phenotype interactions and adaptive mechanisms within SDS populations.
Conclusion
The comprehensive exploration of allelic diversity within the Ovar-DRB1 gene in SDS has provided valuable insights into the genetic foundations of immune responses and disease resistance. Our findings underscore the intricate interplay between genetics and health outcomes in livestock populations. The identification of breed-specific variants and novel alleles within Ovar-DRB1 highlights the unique adaptive potential of different SDS breeds. These insights have direct implications for informed breeding strategies, disease management, and conservation efforts. By unraveling the complex relationships between genetic diversity and immune responses, this study contributes to the advancement of livestock health and resilience.
Future perspectives
As we move forward, several avenues for future research emerge from this study. Expanding the scope of genetic analysis beyond the Ovar-DRB1 gene to include other immune-related genes could provide a more comprehensive understanding of immune system dynamics in SDS populations. Longitudinal studies tracking allele frequencies over time and in response to changing disease pressures will shed light on the evolving genetic landscape and adaptive mechanisms. Integrating genomic data with phenotypic and environmental information will enable a holistic assessment of genotype–phenotype interactions. Collaborative efforts among geneticists, immunologists, livestock managers, and policy makers remain essential. Such interdisciplinary cooperation will facilitate the translation of genetic insights into practical strategies for enhancing livestock health, productivity, and resilience. Additionally, continued collaboration with international partners will enable cross-comparisons with other sheep populations, further enriching our understanding of genetic diversity and adaptation. In conclusion, this study serves as a stepping stone towards harnessing genetic diversity for the betterment of Sudan Desert Sheep populations. By addressing limitations, embracing cross-disciplinary collaboration, and embarking on future research endeavors, we can unlock the full potential of genetic diversity to ensure the health and prosperity of these valuable livestock breeds.
Availability of data and materials
The Illumina MiSeq sequences obtained were deposited to the DNA Data Bank of Japan (https://www.ncbi.nlm.nih.gov/sra/?term =) under accession number DRA017273. The processed sequences of the all detected alleles in the SDS were submitted to Genbank under accession numbers OR488718-OR488752, OR488756, OR488764, OR488770, and OR636395 (https://www.ncbi.nlm.nih.gov/Genbank/update.html).
References
Portanier E, Garel M, Devillard S, Maillard D, Poissant J, Galan M, et al. Both candidate gene and neutral genetic diversity correlate with parasite resistance in female Mediterranean mouflon. BMC Ecol. 2019;19:1–14.
Esmailnejad A, Ganjiani V, Hosseini-Nasab E, Nazifi S. Association of Ovar-DRB1 alleles with innate immune responses in sheep. Vet Med Sci. 2022;8(2):752–7.
Ballingall K, Herrmann-Hoesing L, Robinson J, Marsh S, Stear M. A single nomenclature and associated database for alleles at the major histocompatibility complex class II DRB1 locus of sheep. Tissue Antigens. 2011;77(6):546–53.
Gornas N, Weimann C, El Hussien A, Erhardt G. Genetic characterization of local Sudanese sheep breeds using DNA markers. Small Rumin Res. 2011;95(1):27–33.
Salim B, Alasmari S, Mohamed NS, Ahmed MKA, Nakao R, Hanotte O. Genetic variation and demographic history of Sudan desert sheep reveal two diversified lineages. BMC Genomics. 2023;24(1):118.
Özdil F, Ilhan F, Işık R. Genetic characterization of some Turkish sheep breeds based on the sequencing of the Ovar-DRB1 gene in the major histocompatibility complex (MHC) gene region. Arch Anim Breed. 2018;61(4):475–80.
Ameur AA, Yilmaz O, Nezih A, Cemal I, Gaouar SBS. Assessment of genetic diversity of Turkish and Algerian native sheep breeds. Acta Agric Slov. 2020;115(1):5–14.
Bay V, Keleş M, Aymaz R, Hatipoğlu E, Öner Y, Yaman Y. Documentation of extensive genetic diversity in the Ovar-DRB1 gene in native Turkish sheep. Anim Biotechnol. 2021;32(4):507–18.
Ballingall KT, Fardoe K, McKeever DJ. Genomic organisation and allelic diversity within coding and non-coding regions of the Ovar-DRB1 locus. Immunogenetics. 2008;60:95–103.
Van Tassell CP, Smith TP, Matukumalli LK, Taylor JF, Schnabel RD, Lawley CT, et al. SNP discovery and allele frequency estimation by deep sequencing of reduced representation libraries. Nat Methods. 2008;5(3):247–52.
Davey JW, Blaxter ML. RADSeq: next-generation population genetics. Brief Funct Genomics. 2010;9(5–6):416–23.
Mamanova L, Coffey AJ, Scott CE, Kozarewa I, Turner EH, Kumar A, et al. Target-enrichment strategies for next-generation sequencing. Nat Methods. 2010;7(2):111–8.
Ballingall KT, Tassi R. Sequence-based genotyping of the sheep MHC class II DRB1 locus. Immunogenetics. 2010;62:31–9.
Andrews S. FastQC: a quality control tool for high throughput sequence data. 2010. Available online at: http://www.bioinformatics.babraham.ac.uk/projects/fastqc.
Li H, Durbin R. Fast and accurate short read alignment with burrows-wheeler transform. Bioinformatics. 2009;25(14):1754–60.
Chen H, Boutros PC. VennDiagram: a package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinform. 2011;12(1):1–7.
Nei M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics. 1978;89(3):583–90.
Schneider S, Roessli D, Excoffier L. Arlequin, version 2.000: a software for population genetics data analysis. Genetics and Biometry Laboratory: University of Geneva, Geneva; 2000.
Weir BS, Cockerham CC. Estimating F-statistics for the analysis of population structure. Evolution. 1984;38:1358–70.
Rousset F. A complete re-implementation of the GENEPOP software for software for teaching and research. Mol Ecol Resour. 2008;8:103–6.
Slatkin M. A correction to the exact test based on the Ewens sampling distribution. Genet Res. 1996;68(3):259–60.
Cavalli-Sforza LL, Menozzi P, Piazza A. The history and geography of human genes. Princeton: Princeton Univ. Press; 1994.
Hammer Ø, Harper DA. Past: paleontological statistics software package for educaton and data anlysis. Palaeontol Electron. 2001;4(1):1.
Nei M. Genetic distance between populations. Am Nat. 1972;106(949):283–92.
Nei M, Tajima F, Tateno Y. Accuracy of estimated phylogenetic trees from molecular data: II Gene Frequency data. J Mol Evol. 1983;19:153–70.
Sneath PHA, Sokal RR. Numerical taxonomy. In: Kennedy D, Park RB, editors. The principles of numerical classification. San Fransisco: Freemann; 1973. p. 573.
Trees RP. The neighbor-joining method: a new method for. Mol Biol Evol. 1987;4(4):406–25.
Takezaki N, Nei M, Tamura K. POPTREE2: Software for constructing population trees from allele frequency data and computing other population statistics with Windows interface. Mol Biol Evol. 2010;27(4):747–52.
Nei M, Gojobori T. Simple methods for estimating the numbers of synonymous and nonsynonymous nucleotide substitutions. Mol Biol Evol. 1986;3(5):418–26.
Mega X. Molecular evolutionary genetics analysis across computing platforms; S Kumar, G Stecher, M Li, C Knyaz K Tamura. Mol Biol Evol. 2018;35(1):1547–9.
Wilson DJ, McVean G. Estimating diversifying selection and functional constraint in the presence of recombination. Genetics. 2006;172(3):1411–25.
Tamura K, Nei M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol. 1993;10(3):512–26.
Aida Y, Takeshima SN, Baldwin CL, Kaushik AK, Ruvinsky A. Bovine immunogenetics. The Genetics of Cattle. In: Garrick DJ, Ruvinsky A, editors. Wallingford: International; 2015. p. 153–91.
Janeway C, Travers P, Walport M, Shlomchik MJ. Immunobiology: the immune system in health and disease, vol. 2. New York: Garland Pub; 2001.
Hughes AL, Nei M. Pattern of nucleotide substitution at major histocompatibility complex class I loci reveals overdominant selection. Nature. 1988;335(6186):167–70.
Sayers G, Good B, Hanrahan J, Ryan M, Angles J, Sweeney T. Major histocompatibility complex DRB1 gene: its role in nematode resistance in Suffolk and Texel sheep breeds. Parasitology. 2005;131(3):403–9.
Li R, Hui W, Jia B, Shi G, Zhao Z, Shen H, et al. The relationship between MHC-DRB1 gene second exon polymorphism and hydatidosis resistance of Chinese merino (Sinkiang Junken type), Kazakh and Duolang sheep. Parasite J Société Fr Parasitol. 2011;18(2):163.
McClelland EE, Penn DJ, Potts WK. Major histocompatibility complex heterozygote superiority during coinfection. Infect Immun. 2003;71(4):2079–86.
Coltman DW, O’Donoghue P, Jorgenson JT, Hogg JT, Strobeck C, Festa-Bianchet M. Undesirable evolutionary consequences of trophy hunting. Nature. 2003;426(6967):655–8.
Nagaoka Y, Kabeya H, Onuma M, Kasai N, Okada K, Aida Y. Ovine MHC class II DRB1 alleles associated with resistance or susceptibility to development of bovine leukemia virus-induced ovine lymphoma. Cancer Res. 1999;59(4):975–81.
Larruskain A, Minguijón E, Garcia-Etxebarria K, Arostegui I, Moreno B, Juste RA, et al. Amino acid signatures in the Ovar-DRB1 peptide-binding pockets are associated with Ovine Pulmonary Adenocarcinoma susceptibility/resistance. Biochem Biophys Res Commun. 2012;428(4):463–8.
Nikbakht G, Rezaii H, Stear MJ, Talebi MA, Mahmoudzadeh H. Allelic polymorphism in the second exon of Ovar-DRB1 in fat-tailed sheep. Vet J. 2012;192(3):547–9.
Yaman Y, Bay V, Aymaz R, Keleş M, Öner Y, Teferedegn EY, et al. A novel 2 bp deletion variant in Ovine-DRB1 gene is associated with increased Visna/maedi susceptibility in Turkish sheep. Sci Rep. 2021;11(1):14435.
Hedrick PW. Pathogen resistance and genetic variation at MHC loci. Evolution. 2002;56(10):1902–8.
Ellis SA, Hammond JA. The functional significance of cattle major histocompatibility complex class I genetic diversity. Annu Rev Anim Biosci. 2014;2(1):285–306.
Estrada-Reyes Z, Tsukahara Y, Goetsch A, Gipson T, Sahlu T, Puchala R, et al. Effect of Ovar-DRA and Ovar-DRB 1 genotype in small ruminants with haemonchosis. Parasite Immunol. 2018;40(6):e12534.
Liu P, Li G, Zhao N, Song X, Wang J, Shi X, et al. Neutral forces and balancing selection interplay to shape the major histocompatibility complex spatial patterns in the striped Hamster in Inner Mongolia: suggestive of broad-scale local adaptation. Genes. 2023;14(7):1500.
Atlija M, Gutíerrez-Gil B, Arranz JJ, Semmer J, Stear MJ, Buitkamp J. Major histocompatibility complex class IIB polymorphism in an ancient Spanish breed. Immunogenetics. 2015;67:531–7.
Konnai S, Nagaoka Y, Takeshima S, Onuma M, Aida Y. Sequences and diversity of 17 new Ovar-DRB1 alleles from three breeds of sheep. Eur J Immunogenet. 2003;30(4):275–82.
Boettcher PJ, Hoffmann I, Baumung R, Drucker AG, McManus C, Berg P, et al. Genetic resources and genomics for adaptation of livestock to climate change. Front Genet. 2015;5:461.
Hoffmann I. Climate change and the characterization, breeding and conservation of animal genetic resources. Anim Genet. 2010;41:32–46.
Ahmad S, Ramzan F, Aziz-ur-Rahman M, Hussain K, Umer S, Saleem MN, Thekkiniath, J. Parasite control strategies: Selective breeding. Parasitism and parasitic control in animals: Strategies for the developing world. Wallingford: CABI; 2023. p. 168–82.
Bishop SC, Woolliams JA. Genomics and disease resistance studies in livestock. Livest Sci. 2014;166:190–8.
Huntington B, Bernardo TM, Bondad-Reantaso M, Bruce M, Devleesschauwer B, Gilbert W, et al. Global burden of animal diseases: a novel approach to understanding and managing disease in livestock and aquaculture. Rev Sci Tech-Off Int Epizoot. 2021;40(2):567–83.
Dukkipati V, Blair H, Garrick D, Murray A. Ovar-Mhc’-ovine major histocompatibility complex: structure and gene polymorphisms. Genet Mol Res. 2006;5(4):581–608.
Bontrop RE, Otting N, de Groot NG, Doxiadis GG. Major histocompatibility complex class II polymorphisms in primates. Immunol Rev. 1999;167(1):339–50.
Maccari G, Robinson J, Ballingall K, Guethlein LA, Grimholt U, Kaufman J, et al. IPD-MHC 2.0: an improved inter-species database for the study of the major histocompatibility complex. Nucleic Acids Res. 2017;45(1):860–4.
Acknowledgements
We acknowledge the grant from the Ministry of Higher Education and Scientific Research (SDS 2021) and the partial financial support from Japan Society for the Promotion of Science (JSPS) KAKENHI [20KK0151 and 22H02505]. We are also grateful Sheep keepers in Sudan for their assistance and permission to sample their herds. We thank Leonidas H. Olivera for her help in data analysis. We also extend our graditude to Dr. Elfatih Mohamed for crafting the map featured in Figure 1.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Author information
Authors and Affiliations
Contributions
BS, designed, conceptualized research and supervised the work. BS, MAE, FA contributed samples. BS, RN, EC carried out the experiments. RN, OM, GG analyzed data. BS, RN funding acquisition and administered project. BS, GG, MA, OH contributed to the interpretation of the results. BS, RN, GG, OH writing original draft. All authors provided critical feedback and helped shape the research, analysis and manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
We confirm that the study is reported in accordance with ARRIVE guidelines (https://arriveguidelines.org). The study protocol was reviewed and approved by the Faculty of Veterinary Medicine, University of Khartoum. The research was conducted in accordance with their guidelines for sampling domestic animals in Sudan, ensuring ethical compliance in the collection and use of the samples.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1. Fig S1.
Neighbor-joining (NJ) tree constructed from the nucleotide sequence that only encoded the antigen-binding site (ABS) by all reported Ovar-DRB1 alleles and the four new ones (From Ovar-DRB1*Sudan1 to Ovar-DRB1*Sudan4) detected in the Sudan desert sheep breed. Numbers are bootstrap percentages that support each node. Bootstrapping was carried up with 1000 replicates to assess the reliability of individual branches. Abrag (AB), Ashgar (AS), Buze´e, Hamari (H), Kabashi (K), and Watish (W). Breeds where alleles were detected are indicated between brackets.
Additional file 2. Fig. S2.
Cumulative gene frequency plot of Ovar-DRB1 alleles in the Sudan Desert sheep breed: Abrag (AB), Ashgar (AS), Buze´e, Hamari (H), Kabashi (K), and Watish (W).
Additional file 3. Fig S3.
Venn plot of the common Ovar-DRB1 alleles shared by Sudan Desert Sheep breeds: Abrag, Ashgar, Hamari, Kabashi, and Watish breeds.
Additional file 4. Fig. S4.
a-e. Principal component analysis of Ovar-DRB1 gene using the pocket amino acid motifs frequencies in Sudan Desert Sheep (Abrag, Ashgar, Buze´e, Hamari, Kabashi, and Watish): a. Pocket 1, b. Pocket 4, c. Pocket 6, d. Pocket 7, and e. Pocket 9. Percentage of the total variance accounted for the first principal components (PC1, PC2, and PC3) were detailed.
Additional file 5. Table S1.
Genetic distance between pairs of breeds/populations estimated through Nei DA distance (above) and FST (below; p values in parentheses) in Sudan Desert Sheep breeds: Abrag (AB), Ashgar (AS), Buze´e (B), Hamari (H), Kabashi (K), and Watish (W).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Salim, B., Nakao, R., Chatanga, E. et al. Exploring genetic diversity and variation of Ovar-DRB1 gene in Sudan Desert Sheep using targeted next-generation sequencing. BMC Genomics 25, 160 (2024). https://doi.org/10.1186/s12864-024-10053-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-024-10053-3