
Abstract
Background
Insect metamorphosis from larvae to pupae is one of the most important stages of insect life history. Relatively comprehensive information related to gene transcription profiles during lepidopteran metamorphosis is required to understand the molecular mechanism underlying this important stage. We conducted transcriptional profiling of the brain and fat body of the cotton bollworm Helicoverpa armigera (Lepidoptera: Noctuidae) during its transition from last instar larva into pupa to explore the physiological processes associated with different phases of metamorphosis.
Results
During metamorphosis, the differences in gene expression patterns and the number of differentially expressed genes in the fat body were found to be greater than those in the brain. Each stage had a specific gene expression pattern, which contributed to different physiological changes. A decrease in juvenile hormone levels at the feeding stage is associated with increased expression levels of two genes (juvenile hormone esterase, juvenile hormone epoxide hydrolase). The expression levels of neuropeptides were highly expressed at the feeding stage and the initiation of the wandering stage and less expressed at the prepupal stage and the initiation of the pupal stage. The transcription levels of many hormone (or neuropeptide) receptors were specifically increased at the initiation of the wandering stage in comparison with other stages. The expression levels of many autophagy-related genes in the fat body were found to be gradually upregulated during metamorphosis. The activation of apoptosis was probably related to enhanced expression of many key genes (Apaf1, IAP-binding motif 1 like, cathepsins, caspases). Active proliferation might be associated with enhanced expression levels in several factors (JNK pathway: jun-D; TGF-β pathway: decapentaplegic, glass bottom boat; insulin pathway: insulin-like peptides from the fat body; Wnt pathway: wntless, TCF/Pangolin).
Conclusions
This study revealed several vital physiological processes and molecular events of metamorphosis and provided valuable information for illustrating the process of insect metamorphosis from larvae to pupae.
Similar content being viewed by others


Background
Insect metamorphosis from larvae to pupae occurs when insects reach a certain critical weight [1] and is regulated by the cooperation of juvenile hormone (JH) and ecdysone (E). The level of JH in the last instar larva of Lepidoptera greatly decreases to a low level before the wandering stage, remains at a low level at the initiation of the wandering stage, rises to a high level during the prepupal stage and decreases to a low level at the initiation of the pupal stage [2]. The level of E increases at the initiation of the wandering stage, rises to a high level during the prepupal stage and decreases to a low level at the initiation of the pupal stage [2]. As the titer of E rises, the old tissues of insects are separated and degraded, and new tissues are formed [3].
JH is produced by corpora allata (CA). JH acid methyltransferase (JHAMT) is the key rate-limiting enzyme controlling the titer of JH in the last instar of insects. Reduction in JHAMT transcription results in a decrease in the JH titer [4]. In the cricket, the TGF-β signaling ligands decapentaplegic (Dpp) and myoglianin regulate the transcription of JHAMT. Specifically, Dpp signaling can enhance JHAMT transcription, while myoglianin signaling can reduce JHAMT transcription by inhibiting Dpp signaling [5, 6]. Myoglianin inhibition of JHAMT transcription has been identified in crickets, cockroaches and Tribolium castaneum [6,7,8]. In Drosophila, myoglianin expression is induced by the transcription factor Max‐binding protein (MNT) [9]. It is unclear whether the regulatory role of MNT on myoglianin is also present in other insects. Juvenile hormone binding protein (JHBP) delivers JH to cells. RNA interference of JHBP causes a decrease in the JH titer in H. armigera [10, 11]. JH degradation is regulated by juvenile hormone esterase (JHE) and juvenile hormone epoxide hydrolase (JHEH) [12, 13].
After JH decreases to a certain titer, the prothoracic gland (PG) begins to secrete E under the stimulation of prothoracicotropic hormone (PTTH). A small amount of E triggers biosynthesis and secretion of a large amount of E [14]. 20-hydroxyecdysone (20E), an active metabolite of E, triggers the beginning of insect metamorphosis. Interestingly, ecdysone production is also mediated by TGF-β signaling; the TGF-β ligand Dpp inhibits ecdysone production, while activin signaling promotes ecdysone production [15,16,17].
The insulin-signaling pathway is an important pathway in insect development and metamorphosis, which interacts with JH and E. On the one hand, insulin signaling can promote the biosynthesis of E in the PG by directly promoting the expression of ecdysone biosynthetic genes and inhibiting FOXO activity [18,19,20]. On the other hand, ecdysone signaling can antagonize insulin signaling by decreasing phosphorylation levels of insulin receptor and serine-threonine protein kinase Akt [21]. In addition, E and JH form negative feedback regulation by inhibiting each other's biosynthesis [22, 23].
Autophagy, apoptosis and cell proliferation of larval tissues regulated by E are vital components of metamorphosis. Autophagy-related genes (ATGs) play vital roles in the autophagy process. Autophagy-related proteins are involved in four steps of the autophagy process, including induction, cargo recognition and selectivity, autophagosome formation, vesicle fusion and autophagosome breakdown [24]. In addition, autophagy precedes and mediates 20E-induced apoptosis [25, 26]. 20E triggers autophagy and apoptosis by changing the intracellular concentration of Ca2+ [25]. Cytochrome c released from mitochondria promotes caspase-9 and caspase-3 activation to induce apoptosis in vertebrates, but it has little effect on inducing apoptosis in C. elegans and Drosophila [27,28,29,30]. However, cytochrome c release is probably required during apoptosis in lepidopteran insects [26]. Caspases are expressed as procaspases, and the activation of caspases triggers apoptosis by degrading critical cellular substrates [31]. Caspase DRONC (the initiation caspase) and caspase 1 (the effector caspase) play vital roles in apoptosis in Drosophila and lepidopteran insects, respectively [32, 33]. 20E induces autophagy and caspase activation during metamorphosis [34].
The brain is an important organ that controls insect hormone secretion and behavior. The main functions of fat bodies include energy metabolism, nutrient storage, detoxification and the immune response [35]. Neuropeptides are extremely diverse and play an important role in many physiological processes. In addition, they often cooperate with each other to regulate certain behaviors and have multiple functions [36]. The neuropeptide SIFamide is involved in translation of hunger signals [37]. Dopamine and dopamine receptors are related to the regulation of motor behavior, food-seeking behavior, and the initiation of metamorphosis [38,39,40]. 5-Hydroxytryptamine and its receptor are involved in the modulation of feeding and gut contractions, regulation of larval locomotion [41, 42]. Octopamine is related to aggression, appetitive and aversive memory recall, and modulation of behavior and metabolism [43,44,45]. The changes in gene transcription profiles in the brain of lepidopteran insects, especially the changes in neuropeptides, during metamorphosis from larva to pupa are unknown.
Helicoverpa armigera is a serious agricultural pest worldwide. Rapid evolution of resistance to chemical pesticides and Bacillus thuringiensis toxin was observed in this pest. In recent years, great progress has been made in hormonal regulation of insect development using H. armigera as a model [46]. Understanding of the physiological processes and molecular mechanisms of development of cotton bollworm will help to find more effective control methods. Although the molecular regulatory mechanisms of hormone (JH and 20E) regulation and autophagy of insect metamorphosis have been studied, the specific physiological processes of H. armigera from larvae to pupae are not clear. Little is known about the molecular mechanism of cell proliferation in lepidopteran insects during metamorphosis from larvae to pupae. Comparative transcriptome analysis was used to analyze specific gene expression profiles in the brain and fat body of H. armigera at different stages of metamorphosis in this study. In addition, we further studied several important physiological processes of metamorphosis, including regulation of JH and ecdysone signaling pathways, autophagy, apoptosis and cell proliferation. Our primary aim was to assess the changes in crucial genes and signaling pathways during different physiological processes from larvae to pupae to provide more references for studies of insect metamorphosis.
Results
Overall transcriptome analysis of the brain and fat body of H. armigera at different physiological stages
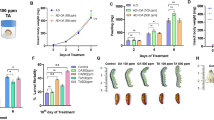
The metamorphosis of H. armigera from larvae to pupae underwent several stages. From the first day to the third day of the fifth instar, larvae fed continuously and accumulated energy to prepare for metamorphosis. On the third day of the fifth instar, the beginning of metamorphosis, larvae stopped feeding and started to wander. From the third day to the sixth day, the fifth instar larvae entered the prepupal stage, and the larval tissues underwent remodeling. From about the sixth day of the fifth instar, larvae gradually completed the transition from larvae to pupae. In this study, four key stages were chosen to study metamorphosis from larvae to pupae, including the feeding stage (L5D2: the second day of the fifth instar larvae), the initiation of the wandering stage (L5W0: the third day of the fifth instar larvae), the prepupal stage (L5D5: the fifth day of fifth instar larvae), and the initiation of the pupal stage (P0) (Fig. 1A).

Phenotypes and transcriptomic overview of Helicoverpa armigera at different metamorphosis stages. A Metamorphosis phenotypes of the last instar of H. armigera larvae at different stages. The pictures below were dissected insects revealing their midgut and fat bodies. B PCA plot of global gene expression of all samples. C Total number of DEGs at different stages. D Venn diagram exhibiting common and unique DEGs in the brain at different stages. E Venn diagram exhibiting common and unique DEGs in the fat body at different stages. L5D2: the feeding stage; L5W0: the initiation of the wandering stage; L5D5: the prepupal stage; P0: the initiation of the pupal stage
Transcriptome analysis was conducted to investigate the changes in gene expression in the brain and fat body of H. armigera at four different physiological stages, including the feeding stage (L5D2) (the brain: BL5D2; the fat body: FL5D2), the initiation of the wandering stage (L5W0) (the brain: BL5W0; the fat body: FL5W0), the prepupal stage (L5D5) (the brain: BL5D5; the fat body: FL5D5), and the initiation of the pupal stage (P0) (the brain: BP0; the fat body: FP0). A total of 174.67 Gb clean data of 24 samples was obtained, and the amount of clean data for each example exceeded 6.34 Gb with the percentage of Q30 ≥ 94.64 (Additional file 1: Table S1). For each sample, 68.8-81.3% of the clean reads were mapped to the H. armigera genome (Additional file 2: Table S2).
The different stages of metamorphosis from larva to pupa were separated by a PCA map (Fig. 1B). PCA showed that different stages of metamorphosis were significantly different. The samples from the brain at different physiological stages were clustered more closely than samples from the fat body, which indicated that the changes in gene expression in the fat body were greater than those in the brain.
|log2 FC |≥ 1 and FDR adjusted P value < 0.05 were set as thresholds, and differentially expressed genes (DEGs) were obtained from the two different tissues at four different physiological stages (Additional file 3: Table S3). A total of 928 DEGs were identified in BL5D2 vs BL5W0 (310 upregulated and 618 downregulated). A total of 1249 DEGs were identified in BL5W0 vs BL5D5 (427 upregulated and 822 downregulated). A total of 588 DEGs were identified in BL5D5 vs BP0 (405 upregulated and 183 downregulated). A total of 4094 DEGs were identified in FL5D2 vs FL5W0 (2070 upregulated and 2024 downregulated). A total of 3486 DEGs were identified in FL5W0 vs FL5D5 (1796 upregulated and 1690 downregulated). A total of 2284 DEGs were identified in FL5D5 vs FP0 (1075 upregulated and 1209 downregulated). The numbers of DEGs in the brain at different physiological stages were lower than those in the fat body (Fig. 1C). Among the comparisons of different stages, 88 and 435 shared DEGs were found in the brain and fat body, respectively (Fig. 1D and E).
GO analysis of DEGs
To investigate the functional categories of the DEGs in the brain and fat body at four different physiological stages (L5D2, L5W0, L5D5 and P0), GO enrichment analysis was carried out. The top 20 GO enrichment categories of DEGs in the brain and fat body at different stages are shown in Fig. 2.

GO enrichment analysis of DEGs of Helicoverpa armigera at different stages. A Enriched GO analysis of DEGs between BL5D2 and BL5W0. B Enriched GO analysis of DEGs between FL5D2 and FL5W0. C Enriched GO analysis of DEGs between BL5W0 and BL5D5. D Enriched GO analysis of DEGs between FL5W0 and FL5D5. E Enriched GO analysis of DEGs between BL5D5 and BP0. F Enriched GO analysis of DEGs between FL5D5 and FP0. BL5D2: the brain at the feeding stage; FL5D2: the fat body at the feeding stage; BL5W0: the brain at the initiation of the wandering stage; FL5W0: the fat body at the initiation of the wandering stage; BL5D5: the brain at the prepupal stage; FL5D5: the fat body at the prepupal stage; BP0: the brain at the initiation of the pupal stage; FP0: the fat body at the initiation of the pupal stage
Initial phase of metamorphosis
Figures 2A and B show the GO enriched categories of the last instar larvae of H. armigera from the feeding stage to the initiation of the wandering stage (L5D2 to L5W0). During the feeding stage, the larvae fed continuously, and the fat body content increased (Fig. 1A: L5D2). At the initiation of the wandering stage, the larvae stopped feeding, emptied midgut and began to wander (Fig. 1A: L5W0). GO enrichment analysis revealed that the DEGs in BL5D2 vs BL5W0 were enriched in the inner mitochondrial membrane protein complex, mitochondrial protein complex and respiratory chain complex (Fig. 2A). Mitochondria are the site of energy production in cells. The genes associated with the inner mitochondrial membrane protein complex, including NADH dehydrogenase subunits, cytochrome b-c1 complex subunit 9, cytochrome c oxidase subunit 6A1, ATP synthase-coupling factor 6, and ATP synthase subunits, were highly expressed at the feeding stage (L5D2) (Additional file 4: Table S4), indicating that the brain had high energy metabolism at the feeding stage.
The DEGs in FL5D2 vs FL5W0 were enriched in the pyruvate metabolic process, amino acid activation and tRNA aminoacylation (Fig. 2B). Aminoacyl tRNA synthetases are required for protein synthesis by catalyzing the linking of amino acids to their cognate tRNAs [47]. The genes related to amino acid activation (aminoacyl tRNA ligases) were highly expressed during the feeding stage (FL5D2) (Additional file 4: Table S4), implying that protein synthesis was more active during the feeding stage, which was associated with a continuous increase in the fat body content.
Metamorphosis progression
Figures 2C and D show the GO enriched categories of the last instar larvae of H. armigera from the initiation of the wandering stage to the prepupal stage (L5W0 to L5D5). During the prepupal stage, the larval body size shrank under the stimulation of E. The fat body was separated and degraded, and the midgut turned red (Fig. 1A: L5D5). GO enrichment analysis revealed that the DEGs in BL5W0 vs BL5D5 were enriched in amino sugar metabolic processes, neuropeptide signaling pathways and aminoglycan metabolic processes (Fig. 2C).
The DEGs in FL5W0 vs FL5D5 were enriched in the cellular amide metabolic process, amide biosynthetic process and translation (Fig. 2D). Tissues began to undergo autophagy and apoptosis during the prepupal stage. Ribosomal proteins play a vital role in protein synthesis [48]. The transcription levels of most ribosomal protein genes decreased from the initiation of the wandering stage to the prepupal stage (L5W0 to L5D5) (Additional file 4: Table S4), indicating that protein synthesis reduced during the prepupal stage.
The later phase of metamorphosis
Figures 2E and F show the GO enriched categories of the last instar larvae of H. armigera from the prepupal stage to the initiation of the pupal stage (L5D5 to P0). At the initiation of the pupal stage, the last instar larvae molted and became pupae (Fig. 1A: P0). GO enrichment analysis revealed that the DEGs in BL5D5 vs BP0 were enriched in chitin metabolic processes, aminoglycan metabolic processes and glucosamine-containing compound metabolic processes (Fig. 2E). The DEGs in FL5D5 vs FP0 were enriched in carbohydrate metabolic processes, oxoacid metabolic processes and carboxylic acid metabolic processes (Fig. 2F).
Genes related to the JH and ecdysone signaling pathways
The JH and ecdysone signaling pathways are the key factors determining insect metamorphosis from larvae to pupae. Based on previous reports [4, 10,11,12,13, 49,50,51,52], the regulatory model of JH and ecdysone signaling pathways is shown in Fig. 3A. The changes in hormone titers can be indicated by the expression levels of the early JH and ecdysone response genes. The early JH-response gene Krüppel homolog 1 (Kr-h1) in the brain and fat body was highly expressed at the prepupal stage (L5D5) compared with other stages (Figs. 3B, C, D and E; additional file 5: Table S5), implying that JH rose during the prepupal stage. The JHE and JHEH genes in the fat body were highly expressed at the feeding stage (L5D2) in comparison with the initiation of the wandering stage (L5W0) (Fig. 3D; additional file 5: Table S5), implying that they played a role in decreasing the level of JH during the feeding stage. JHBP in the fat body was expressed at lower levels at the initiation of the wandering stage (L5W0) than at other stages (Fig. 3D; additional file 5: Table S5). Broad-Complex (Br–C) and ecdysone hormone-response protein 75 (E75) were highly expressed at the prepupal stage (L5D5) compared with the initiation of the wandering stage (L5W0) (Figs. 3B, C, D and E; additional file 5: Table S5). The expression level of PTTH in the brain was high at the feeding stage (L5D2) and the initiation of the wandering stage (L5W0) and decreased at the prepupal stage (L5D5) and the initiation of the pupal stage (P0) (Figs. 3B and C; additional file 5: Table S5).

Expression of genes related to the JH and ecdysone signaling pathways during metamorphosis of Helicoverpa armigera. A The regulatory model of juvenile hormone (JH) and 20E signaling pathways. B and D Heatmap of genes in the brain (B) and fat body (D) related to the JH and ecdysone signaling pathways. C and E qPCR validation of genes in the brain (C) and fat body (E) related to regulation of JH and ecdysone signaling pathways. Histograms represent the results of qPCR (left Y axis). Line charts represent the results of RNA-Seq (right Y axis). Error bars represent mean ± SD. One-way ANOVA with Tukey’s post hoc test was used for statistical analysis of qPCR results. Different lowercase letters indicated significant differences (p < 0.05). BL5D2: the brain at the feeding stage; BL5W0: the brain at the initiation of the wandering stage; BL5D5: the brain at the prepupal stage; BP0: the brain at the initiation of the pupal stage; FL5D2: the fat body at the feeding stage; FL5W0: the fat body at the initiation of the wandering stage; FL5D5: the fat body at the prepupal stage; FP0: the fat body at the initiation of the pupal stage
Genes related to neuropeptides and hormone (or neuropeptide) receptors
Various neuropeptide genes in the brain of H. armigera were highly expressed at the feeding stage (L5D2) and the initiation of the wandering stage (L5W0) and less expressed at the prepupal stage (L5D5) and the initiation of the pupal stage (P0) (Fig. 4A and C; additional file 6: Table S5), indicating that these neuropeptides played important roles in the physiological processes during the feeding stage and the initiation of the wandering stage. The transcription levels of many hormone (or neuropeptide) receptors, including receptors of dopamine, octopamine, neuropeptide SIFamide, 5-hydroxytryptamine and somatostatin, were specifically increased at the initiation of the wandering stage (L5W0) in comparison with other stages (Fig. 4B; additional file 6: Table S6). At the initiation of the wandering stage, cotton bollworm larvae usually empty their intestines and actively crawl around to find a place to build a pupal chamber, with which they prepare for the initiation of metamorphosis. The enhanced expression levels of these hormone (or neuropeptide) receptors may be related to the series of sophisticated behaviors at this stage.

Expression of genes related to neuropeptides and hormone (or neuropeptide) receptors during metamorphosis of Helicoverpa armigera. A and B Heatmap of genes in the brain related to neuropeptides (A) and hormone (or neuropeptide) receptors (B). C qPCR validation of a gene in the brain related to neuropeptide. Histograms represent the results of qPCR (left Y axis). Line charts represent the results of RNA-Seq (right Y axis). Error bars represent mean ± SD. One-way ANOVA with Tukey's post hoc test was used for statistical analysis of qPCR results. Different lowercase letters indicated significant differences (p < 0.05). BL5D2: the brain at the feeding stage; BL5W0: the brain at the initiation of the wandering stage; BL5D5: the brain at the prepupal stage; BP0: the brain at the initiation of the pupal stage
Genes related to autophagy
The Atg1-Atg13 complex is involved in the process of autophagosome initiation [53]. In H. armigera, 14 ATGs were identified. Although the transcription levels of several ATGs (ATG5, ATG7, ATG12 and ATG13) in the brain at the initiation of the wandering stage (L5W0) were higher in comparison with other stages (Figs. 5A and B; additional file 7: Table S7), there were few differences in the expression levels (less than 1.5-fold change in pairwise comparisons) at different stages. The transcription levels of many ATGs (ATG5, ATG6, ATG7, ATG8, ATG9, ATG12) in the fat body gradually increased from the feeding stage (L5D2) to the initiation of the pupal stage (P0) (Figs. 5C and D; additional file 7: Table S7). The mRNA levels of ATG1, ATG4, and ATG18 in the fat body decreased from the initiation of the wandering stage to the prepupal stage (L5W0 to L5D5) (Fig. 5C; additional file 7: Table S7).

Expression of genes related to autophagy during metamorphosis of Helicoverpa armigera. A and C Heatmap of genes in the brain (A) and fat body (C) related to autophagy. B and D: qPCR validation of genes in the brain (B) and fat body (D) related to autophagy. Histograms represent the results of qPCR (left Y axis). Line charts represent the results of RNA-Seq (right Y axis). Error bars represent mean ± SD. One-way ANOVA with Tukey’s post hoc test was used for statistical analysis of qPCR results. Different lowercase letters indicated significant differences (p < 0.05). BL5D2: the brain at the feeding stage; BL5W0: the brain at the initiation of the wandering stage; BL5D5: the brain at the prepupal stage; BP0: the brain at the initiation of the pupal stage; FL5D2: the fat body at the feeding stage; FL5W0: the fat body at the initiation of the wandering stage; FL5D5: the fat body at the prepupal stage; FP0: the fat body at the initiation of the pupal stage
Genes related to apoptosis and proliferation
The larval tissues are removed through caspase-induced apoptosis, and new pupal-adult tissues are formed via proliferation under the control of E [3]. The transcription levels of caspase DRONC and caspase 1 in the brain and fat body were enhanced from the feeding stage to the initiation of the pupal stage (L5D2 to P0) (Figs. 6A, B, C and D; additional file 8: Table S8), indicating that apoptosis was active in cotton bollworm cells during metamorphosis. Three genes encoding matrix metalloproteinases (matrix metalloproteinase 1, 2 and 14-like) in the brain and fat body were upregulated during metamorphosis (Figs. 6A and C; additional file 8: Table S8). Cathepsin B, D, and L genes in the fat body were highly expressed during metamorphosis (Figs. 6C and D; additional file 8: Table S8). Interestingly, the transcription level of cathepsin B in the brain decreased from the initiation of the wandering stage to the prepupal stage (L5W0 to L5D5) (Figs. 6A and B; additional file 8: Table S8).
JAK/STAT [54], TGF-β [55], Wnt/β-catenin [56], and insulin signaling [57] are involved in the differentiation and proliferation of stem cells. Many genes in H. armigera related to cell differentiation and proliferation were highly expressed at the initiation of the pupal stage (P0), including jun-D, Dpp, glass bottom boat (Gbb), wntless, TCF/Pangolin and insulin-like peptides from the fat body (Figs. 6A, B, C and D, 7C and D; additional file 8: Table S8; additional file 9: Table S9).

Expression of genes related to apoptosis and proliferation during metamorphosis of Helicoverpa armigera. A and C Heatmap of genes in the brain (A) and fat body (C) related to apoptosis and proliferation. B and D qPCR validation of genes in the brain (B) and fat body (D) related to apoptosis and proliferation. Histograms represent the results of qPCR (left Y axis). Line charts represent the results of RNA-Seq (right Y axis). Error bars represent mean ± SD. One-way ANOVA with Tukey’s post hoc test was used for statistical analysis of qPCR results. Different lowercase letters indicated significant differences (p < 0.05). BL5D2: the brain at the feeding stage; BL5W0: the brain at the initiation of the wandering stage; BL5D5: the brain at the prepupal stage; BP0: the brain at the initiation of the pupal stage; FL5D2: the fat body at the feeding stage; FL5W0: the fat body at the initiation of the wandering stage; FL5D5: the fat body at the prepupal stage; FP0: the fat body at the initiation of the pupal stage

Expression of genes related to the insulin-signaling pathway during metamorphosis of Helicoverpa armigera. A and C Heatmap of genes in the brain (A) and fat body (C) related to the insulin-signaling pathway. B and D qPCR validation of genes in the brain (B) and fat body (D) related to the insulin-signaling pathway. Histograms represent the results of qPCR (left Y axis). Line charts represent the results of RNA-Seq (right Y axis). Error bars represent mean ± SD. One-way ANOVA with Tukey’s post hoc test was used for statistical analysis of qPCR results. Different lowercase letters indicated significant differences (p < 0.05). BL5D2: the brain at the feeding stage; BL5W0: the brain at the initiation of the wandering stage; BL5D5: the brain at the prepupal stage; BP0: the brain at the initiation of the pupal stage; FL5D2: the fat body at the feeding stage; FL5W0: the fat body at the initiation of the wandering stage; FL5D5: the fat body at the prepupal stage; FP0: the fat body at the initiation of the pupal stage
Insulin-signaling pathway
A total of nine insulin-like peptides (bombyxin C 1-like, B 10-like 1, B 10-like 2, A 1 homolog, B 2-like, related peptide A, G 1-like, G 1-like 2 and G 1-like 3) were identified from our transcriptome data (additional file 9: Table S9). Six insulin-like peptides (bombyxin C 1-like, B 10-like 1, B 10-like 2, A 1 homolog, B 2-like and related peptide A) were specifically expressed in the brain (Figs. 7A and B; additional file 9: Table S9), and two insulin-like peptides (bombyxin G 1-like and bombyxin G 1-like 2 [58]) were specifically expressed in the fat body (Figs. 7C and D; additional file 9: Table S9). One insulin-like peptide (bombyxin G 1-like 3 [58]) was expressed in both the brain and the fat body (Figs. 7A and C; additional file 9: Table S9). Insulin-like peptides from the brain were strongly expressed at the feeding stage (L5D2) and the initiation of the wandering stage (L5W0) in comparison with other stages (Figs. 7A and B; additional file 9: Table S9). During insect feeding, the intake of sugars and amino acids facilitates the secretion of insulin-like peptides from the brain by promoting the secretion of CCHamide 2 in the fat body [59]. The expression of CCHamide 2 in the brain (Fig. 7A; additional file 9: Table S9), rather than in the fat body, was consistent with the expression pattern of insulin-like peptides in the brain. The expression of insulin-like peptides from the brain at the initiation of the wandering stage (L5W0) may be associated with the secretion of E. Three insulin-like peptides (bombyxin G 1-like, G 1-like 2 and G 1-like 3) in the fat body highly expressed at the initiation of the pupal stage (Figs. 7C and D; additional file 9: Table S9), indicating that insulin-like peptides from the fat body were probably associated with active cell proliferation during the metamorphosis of H. armigera.
Discussion
Changes in the expression genes and signaling pathways related to hormone signaling
According to the change in JH titer in lepidopteran insect from larva to pupa [2] and the transcription level change of Kr-h1 (Figs. 3B, C, D and E; additional file 5: Table S5), the level of JH in the last instar larvae of cotton bollworm was predicted to maintain at a low level at the feeding stage and the initiation of the wandering stage, rise to a high level during the prepupal stage, and decrease to a low level at the initiation of the pupal stage. The titer of JH in the last instar of lepidopteran larvae is related to the transcription of JHAMT [4]. In Bombyx mori, the transcription level of JHAMT decreases rapidly and disappears in CA of the last instar larvae. However, other peripheral tissues still have JHAMT activity which is probably the reason causing a high level of JH in the larval hemolymph during the prepupal stage [4]. It is too hard to collect enough CA, thus we only detected JHAMT expression in the brain without corpus cardiacum and CA. The expression level of JHAMT in the CA of H. armigera was not clear in this study. Whether the expression level of JHAMT in the brain (no corpus cardiacum and CA) (Fig. 3B; additional file 5: Table S5) was related to JH titer needs to be further studied. Myoglianin regulates JHAMT transcription in CA of Gryllus bimaculatus to control JH levels to trigger metamorphosis [6]. Myoglianin is considered to triggered metamorphosis at the threshold size in Manduca sexta and T. castaneum [7]. The function of myoglianin in H. armigera needs to be investigated.
The secretion of E mainly starts at the initiation of the wandering stage according to previous studies [60]. In Drosophila, many factors control the production of E, including PTTH, insulin signaling, TGF-β/Activin signaling, nitric oxide, and prothoracicostatic factors [61]. PTTH and insulin signaling are involved in the production of E in B. mori, while insulin signaling is not required for the ecdysone production in M. sexta [19]. The transcription levels of PTTH (Figs. 3B and C; additional file 5: Table S5) and insulin-like peptides from the brain, except for bombyxin G1-like 3 (Figs. 7A and B; additional file 9: Table S9), were decreased from the feeding stage to the initiation of the wandering stage (L5D2 to L5W0), indicating that there may be other factors contributing to the biosynthesis of E in the PGs of H. armigera (Fig. 8A). JH increases again after the beginning of metamorphosis to prevent excessive metamorphosis by antagonizing 20E [50]. Kr-h1, a key JH response gene, may play an important role in preventing excessive metamorphosis in H. armigera considering its high expression level at the prepupal stage (Figs. 3B, C, D and E; additional file 5: Table S5). Overexpression of Kr-h1 reduced the transcription levels of EcR, USP and other ecdysone response genes in Drosophila [22]. The titer of 20E rises from the initiation of the wandering stage to the prepupal stage [60], while the expression levels of the ecdysone receptor EcR, USP and the ecdysone response protein 74 (E74) in the brain did not increase from the initiation of the wandering stage to the prepupal stage (L5W0 to L5D5) (Figs. 3B and C; additional file 5: Table S5), indicating that JH antagonizes ecdysone signaling by reducing the transcription of EcR, USP and E74 via Kr-h1. On the whole, 20E signaling was activated during the metamorphosis of H. armigera. JH signaling was also activated to partially block 20E signaling during the prepupal stage of H. armigera.

Putative physiological processes of Helicoverpa armigera during its metamorphosis. A The regulation of ecdysone production. B Autophagy and apoptosis induced by 20E. C Proliferation induced by 20E. The solid arrow represents direct effect. The dotted arrow represents indirect effect
Autophagy and apoptosis
When JH is below a certain limit, the secretion of E will cause autophagy and apoptosis. Generally, autophagy and apoptosis signaling were activated from the initiation of the wandering stage to the initiation of the pupal stage, implying an important role of these two signaling pathways in the metamorphosis of H. armigera.
Autophagy is a self-protection function of cells under external pressure. Ecdysone response genes induce the expression of autophagy response genes and block PI3K-TORC1 signaling to induce autophagy [53, 62]. The high transcriptional level of Br–C and E75 at the prepupal stage (L5D5) (Figs. 3B, C, D and E; additional file 5: Table S5) implied these two ecdysone response genes may play an important role in inducing autophagy and apoptosis in H. armigera. The level of 20E was low at the initiation of the pupal stage according to a previous study [60]. E93, a 20E response gene, upregulates the transcription levels of ATGs in Drosophila [34]. E93 inhibited by Kr-h1 and induced by 20E promotes autophagy in the fat body of B. mori [63, 64]. The E93 gene (Figs. 3B and D; additional file 5: Table S5) and many ATGs (Figs. 5A and C; additional file 7: Table S7) in the brain and fat body were highly expressed at the initiation of the pupal stage (P0), suggesting that upregulation of E93 was related to the induction of ATGs at the initiation of the pupal stage.
20E activates reaper and hid to induce cell apoptosis during Drosophila metamorphosis [65]. Ibm1, the reaper/grim ortholog, has been identified in B. mori and is associated with cell apoptosis [66]. The transcription of ibm1-like, the ibm1 ortholog of H. armigera, increased in the brain and fat body from the feeding stage to the initiation of the pupal stage (L5D2 to P0) (Figs. 6A and C; additional file 8: Table S8), indicating that ibm1-like was involved in the activation of apoptosis. Cathepsins are released from the lysosomes into the cytoplasm when stimulated by signals [67]. Lysosomes are involved in the process of autophagy by forming autolysosomes [24]. In addition, autophagy precedes apoptosis, and inhibition of autophagy prevents apoptosis [60]. Cathepsin L and D promote fat body degradation and programmed cell death during the metamorphosis of B. mori [68, 69]. Cathepsin L upregulated by 20E participates in the regulation of granulation of haemocytes and triggers apoptosis of midgut cells by activating caspase-1 in H. armigera [33, 70]. Mature cathepsin D triggered by autophagy promotes apoptosis via activating caspase-3 in the midgut of H. armigera [60]. Considering their high expression levels (Figs. 6C and D; additional file 8: Table S8), cathepsin B, D, and L in the fat body may play a vital role in the activation of apoptosis in H. armigera. These results implied that 20E might induce apoptosis by releasing cathepsins, which were activated by autolysosomes, to activate caspases in H. armigera. Furthermore, mitochondria are involved in the apoptosis of lepidopteran insect cells [26, 71,72,73]. However, there is no evidence that mitochondria are involved in the activation of apoptosis in Drosophila [27]. Apaf1 is essential for mitochondria-dependent apoptosis [74], and its expression level in the brain increased during metamorphosis (Fig. 6A; additional file 8: Table S8), indicating that Apaf1 is involved in the activation of apoptosis in cotton bollworm. In conclusion, many factors, including ibm1-like, autolysosome, cathepsins, mitochondria, and Apaf1, are associated with cell apoptosis of H. armigera during metamorphosis (Fig. 8B).
Cell proliferation
Cell proliferation has a crucial role in insect metamorphosis. From larvae to pupae, larval tissues undergo autophagy, apoptosis and cell proliferation under the regulation of the E. Autophagy and apoptosis are essential for the differentiation and proliferation of tissue and stem cells, which act in an ecdysone concentration-dependent manner [75, 76]. The dissociation of fat body cells is involved in the proliferation of fat bodies, which is regulated by 20E through mediating the expression of matrix metalloproteinases and cathepsins [3, 77, 78]. 20E is required for fat body cell dissociation induced by two matrix metalloproteinases in Drosophila [78]. Cathepsin L regulated by 20E is involved in the dissociation of fat body cell in H. armigera [79]. According to our data (Figs. 6C and D; additional file 8: Table S8), three matrix metalloproteinases (matrix metalloproteinase 1, 2 and 14-like) and three cathepsins (cathepsin B, D and L) in the fat body may play an important role in the dissociation of fat body cells of H. armigera. Apoptotic cells need to leave space for the proliferation of stem cells. The Dpp and Wnt signaling pathways play key roles in the differentiation and proliferation of stem cells [80,81,82]. Wingless promotes cell survival but restricts growth, which determines the final wing size during wing development in Drosophila [83]. Inhibition of caspase on apoptotic cells can promote the secretion of Dpp and wingless. Dpp can induce extra proliferation of cells, and wingless can inhibit the extra proliferation caused by Dpp in Drosophila [84]. The transcription factor TCF/Pangolin in the brain and fat body was highly expressed at the initiation of the pupal stage (P0) (Figs. 6A, B, C and D; additional file 8: Table S8), indicating that Wnt signaling was involved in cell proliferation. Gbb, a BMP ligand, plays an important role in the growth and patterning of Drosophila wings [85, 86]. The transcription level of Dpp in the fat body was enhanced from the initiation of the wandering stage to the initiation of the pupal stage (L5W0 to P0) (Figs. 6C and D; additional file 8: Table S8), and the transcription level of Gbb in the brain and fat body increased from the feeding stage to the initiation of the pupal stage (L5D2 to P0) (Figs. 6A, B, C and D; additional file 8: Table S8), implying that Dpp and Gbb were required for cell proliferation. Activation of the JNK signaling pathway promotes the expression of Dpp and wingless in Drosophila [87, 88]. The transcription level of jun-D in the brain and fat body increased from the initiation of the wandering stage to the initiation of the pupal stage (L5W0 to P0) (Figs. 6A, B, C and D; additional file 8: Table S8), implying that the JNK pathway is located upstream of Dpp and wingless signaling to activate cell proliferation. 20E activates JNK signaling to control oviposition in Anopheles gambiae [89]. In addition, 20E regulates gene expression through an interaction between EcR and Jun during Drosophila dorsal closure [90]. As an insulin-like peptide produced by the fat body, insulin-like peptide 6 induced by 20E signaling, starvation and FOXO regulates insect growth and metabolism [3, 91]. Upregulation of the three insulin-like peptides (bombyxin G1-like, G1-like 2 and G1-like 3) during metamorphosis in the fat body (Figs. 7C and D; additional file 9: Table S9) indicated their roles in cell proliferation of H. armigera. In conclusion, four pathways, including the insulin pathway, JNK pathway, TGF-β pathway, and Wnt pathway, cooperated under the regulation of 20E to promote cell proliferation during H. armigera metamorphosis from larvae to pupae (Fig. 8C).
Conclusions
We mainly studied different processes of metamorphosis by conducting a comprehensive analysis of the transcriptome at different stages in the last larval instar of H. armigera. Transcriptome analysis revealed specific gene expression patterns at different stages of metamorphosis. The expression levels of neuropeptides were downregulated during metamorphosis. Our study showed the specific process of the regulation of JH and ecdysone signaling pathways and the changes in genes related to autophagy at different stages. In addition, our study showed the levels of genes and signaling pathways related to apoptosis and cell proliferation. The molecular mechanisms of cell apoptosis and proliferation during insect metamorphosis from larvae to pupae need to be identified in the future.
Materials and methods
Insect rearing and tissue collection
H. armigera were purchased from Henan Jiyuan Baiyun Industry in China. H. armigera larvae were raised on an artificial diet and a photoperiod of 14 h light:10 h dark at a temperature of 27 ± 1 °C with 60% to 70% humidity. Three biological replicates of the brain (no corpus cardiacum and CA) (each biological replicate included 50 individuals) and the fat body (each biological replicate including 10 individuals) were collected at L5D2 (the feeding stage; the second day of the fifth instar larvae), L5W0 (the initiation of the wandering stage; the third day of the fifth instar larvae), L5D5 (the prepupal stage; the fifth day of the fifth instar larvae), and P0 (the initiation of the pupal stage) in the last instar of H. armigera larvae.
RNA extraction and library construction and sequencing
Total RNA of the brain and fat body tissues was extracted using TRIzol reagent (Invitrogen, USA) according to the manufacturer’s instructions. The RNA quality was determined using a Nanodrop ND-2000 spectrophotometer, and the RNA integrity was evaluated using an Agilent 2100 Bioanalyzer (Agilent, Santa Clara, CA, USA). A cDNA library was constructed using 1 μg of RNA following the Truseq™ RNA Sample Preparation Kit (Illumina, San Diego, CA, USA). Finally, the constructed cDNA libraries were sequenced on an Illumina HiSeq xten/NovaSeq 6000 sequencer (Majorbio, Shanghai, China).
Data analysis
Low-quality and adaptor-polluted reads and high levels of unknown base (N) reads were removed to obtain clean reads. The clean reads were aligned to the H. armigera reference genome (GCF_002156985.1) with orientation mode using HISAT2 [92]. StringTie [93] was used to assemble transcripts, and novel transcripts were identified by Cuffcompare [94]. This assembly contains 19,157 predicted protein-coding genes. The expression levels of genes were calculated by transcripts per million reads (TPM). Differentially expressed genes (DEGs) in the 24 samples were identified using the DESeq2 R package [95]. DEGs with |Log2 (fold change)|≥ 1 and FDR adjusted P value < 0.05 were identified as significant DEGs. Gene Ontology (GO) analysis was performed to assess possible gene functions [96]. DEGs were enriched in three GO groups to produce a summary of the gene groups within the transcriptome. GO terms with the FDR adjusted P value < 0.05 were considered significantly enriched. Average linkage hierarchical clustering analyses were conducted to visualize the expression profiles of metamorphosis-related genes using the RSEM software.
Quantitative reverse transcription PCR (RT–qPCR)
A total of 1.2 μg of RNA was used for cDNA synthesis using the SuperRT cDNA Kit (CWBIO, China). Quantitative real-time PCR (qPCR) was performed on a QuantStudio 12 K Flex Real-time PCR System (Applied Biosystems, USA). The ribosomal protein S3 (rpS3) gene was used as the reference gene [97]. The qPCR conditions were 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 30 s. Three biological replicates were performed for each treatment, and each biological replicate was carried out in triplicate. The expression levels of genes were calculated using the 2−ΔΔct method, and One-way ANOVA with Tukey's post hoc test was used for statistical analysis. The primers used in the qPCR are described in Additional file 10: Table S10.
Availability of data and materials
The H. armigera raw sequencing data were deposited in the Genome Sequence Archive in National Genomics Data Center, Beijing Institute of Genomics, Chinese Academy of Sciences (GSA: CRA006668) that are publicly accessible at https://ngdc.cncb.ac.cn/gsa.
Abbreviations
- DEGs:
-
Differentially expressed genes
- 20E:
-
20-Hydroxyecdysone
- JH:
-
Juvenile hormone
- ATGs:
-
Autophagy-related genes
- JHAMT:
-
Juvenile hormone acid methyltransferase
- Dpp:
-
Decapentaplegic
- Gbb:
-
Glass bottom boat
- Kr-h1:
-
Krüppel homolog 1
- JHE:
-
Juvenile hormone esterase
- JHEH:
-
Juvenile hormone epoxide hydrolase
- JHBP:
-
Juvenile hormone binding protein
- PTTH:
-
Prothoracicotropic hormone
- EcR:
-
Ecdysone receptor
- USP:
-
Ultraspiracle
- Br–C:
-
Broad-complex
- E74:
-
Ecdysone-induced protein 74
- E75:
-
Ecdysone-induced protein 75
- E93:
-
Ecdysone-induced protein 93
- IAPs:
-
Inhibitors of apoptosis
- Ibm1 like:
-
IAP-binding motif 1 like
- rpS3:
-
Ribosomal protein S3
- CA:
-
Corpora allata
- E:
-
Ecdysone
- PG:
-
Prothoracic gland
- Met:
-
Methoprene-tolerant
References
Mirth CK, Riddiford LM. Size assessment and growth control: how adult size is determined in insects. BioEssays. 2007;29:344–55.
Truman JW, Riddiford LM. Endocrine Insights into the Evolution of Metamorphosis in Insects. Annu Rev Entomol. 2002;47:467–500.
Li S, Yu X, Feng Q. Fat Body Biology in the Last Decade. Annu Rev Entomol. 2018;64(1):315–33.
Shinoda T, Itoyama K. Juvenile hormone acid methyltransferase: A key regulatory enzyme for insect metamorphosis. Proc Natl Acad Sci USA. 2003;100:11986–91.
Huang J, Tian L, Peng C, Abdou M, Wen D, Wang Y, et al. DPP-mediated TGFβ signaling regulates juvenile hormone biosynthesis by activating the expression of juvenile hormone acid methyltransferase. Development. 2011;138:2283–91.
Ishimaru Y, Tomonari S, Matsuoka Y, Watanabe T, Miyawaki K, Bando T, et al. TGF-β signaling in insects regulates metamorphosis via juvenile hormone biosynthesis. Proc Natl Acad Sci USA. 2016;113:5634–9.
He LL, Shin SH, Wang Z, Yuan I, Weschler R, Chiou A, et al. Mechanism of threshold size assessment: Metamorphosis is triggered by the TGF-beta/Activin ligand Myoglianin. Insect Biochem Mol Biol. 2020;126:103452.
Kamsoi O, Belles X. Myoglianin triggers the premetamorphosis stage in hemimetabolan insects. FASEB j. 2019;33:3659–69.
Demontis F, Patel VK, Swindell WR, Perrimon N. Intertissue Control of the Nucleolus via a Myokine-Dependent Longevity Pathway. Cell Rep. 2014;7:1481–94.
Suzuki R, Fujimoto Z, Shiotsuki T, Tsuchiya W, Momma M, Tase A, et al. Structural mechanism of JH delivery in hemolymph by JHBP of silkworm. Bombyx mori Sci Rep. 2011;1:133.
Ni M, Ma W, Wang X, Gao M, Dai Y, Wei X, et al. Next-generation transgenic cotton: pyramiding RNAi and Bt counters insect resistance. Plant Biotechnol J. 2017;15:1204–13.
Tan A, Tanaka H, Tamura T, Shiotsuki T. Precocious metamorphosis in transgenic silkworms overexpressing juvenile hormone esterase. Proc Natl Acad Sci USA. 2005;102:11751–6.
Zhang Q-R, Xu W-H, Chen F-S, Li S. Molecular and biochemical characterization of juvenile hormone epoxide hydrolase from the silkworm. Bombyx mori Insect Biochem Mol Biol. 2005;35:153–64.
Mizoguchi A, Kamimura M, Kiuchi M, Kataoka H. Positive feedback regulation of prothoracicotropic hormone secretion by ecdysteroid – A mechanism that determines the timing of metamorphosis. Insect Biochem Mol Biol. 2015;58:39–45.
Denton D, Xu T, Dayan S, Nicolson S, Kumar S. Dpp regulates autophagy-dependent midgut removal and signals to block ecdysone production. Cell Death Differ. 2019;26:763–78.
Gibbens YY, Warren JT, Gilbert LI, O’Connor MB. Neuroendocrine regulation of Drosophila metamorphosis requires TGFβ/Activin signaling. Development. 2011;138:2693–703.
Setiawan L, Pan X, Woods AL, O’Connor MB, Hariharan IK. The BMP2/4 ortholog Dpp can function as an inter-organ signal that regulates developmental timing. Life Sci Alliance. 2018;1:e201800216.
Walkiewicz MA, Stern M. Increased Insulin/Insulin Growth Factor Signaling Advances the Onset of Metamorphosis in Drosophila. PLoS One. 2009;4:e5072.
Smith WA, Lamattina A, Collins M. Insulin signaling pathways in lepidopteran ecdysone secretion. Front Physiol. 2014;5:19.
Koyama T, Rodrigues MA, Athanasiadis A, Shingleton AW, Mirth CK. Nutritional control of body size through FoxO-Ultraspiracle mediated ecdysone biosynthesis. eLife. 2014;3:e03091.
Yuan D, Zhou S, Liu S, Li K, Zhao H, Long S, et al. The AMPK-PP2A axis in insect fat body is activated by 20-hydroxyecdysone to antagonize insulin/IGF signaling and restrict growth rate. Proc Natl Acad Sci USA. 2020;117:9292–301.
Liu S, Li K, Gao Y, Liu X, Chen W, Ge W, et al. Antagonistic actions of juvenile hormone and 20-hydroxyecdysone within the ring gland determine developmental transitions in Drosophila. Proc Natl Acad Sci USA. 2018;115:139–44.
Zhang T, Song W, Li Z, Qian W, Wei L, Yang Y, et al. Krüppel homolog 1 represses insect ecdysone biosynthesis by directly inhibiting the transcription of steroidogenic enzymes. Proc Natl Acad Sci USA. 2018;115:3960–5.
He C, Klionsky DJ. Regulation Mechanisms and Signaling Pathways of Autophagy. Annu Rev Genet. 2009;43:67–93.
Li Y-B, Li X-R, Yang T, Wang J-X, Zhao X-F. The steroid hormone 20-hydroxyecdysone promotes switching from autophagy to apoptosis by increasing intracellular calcium levels. Insect Biochem Mol Biol. 2016;79:73–86.
Xie K, Tian L, Guo X, Li K, Li J, Deng X, et al. BmATG5 and BmATG6 mediate apoptosis following autophagy induced by 20-hydroxyecdysone or starvation. Autophagy. 2016;12:381–96.
Hay BA, Guo M. Caspase-Dependent Cell Death in Drosophila. Annu Rev Cell Dev Biol. 2006;22:623–50.
Kluck RM, Bossy-Wetzel E, Green DR, Newmeyer DD. The Release of Cytochrome c from Mitochondria: A Primary Site for Bcl-2 Regulation of Apoptosis. Science. 1997;275:1132–6.
Kornbluth S, White K. Apoptosis in Drosophila : neither fish nor fowl (nor man, nor worm). J Cell Sci. 2005;118:1779–87.
Jiang X, Wang X. Cytochrome C -Mediated Apoptosis. Annu Rev Biochem. 2004;73:87–106.
Cohen GM. Caspases: the executioners of apoptosis. Biochem J. 1997;326:1–16.
Cakouros D, Daish T, Martin D, Baehrecke EH, Kumar S. Ecdysone-induced expression of the caspase DRONC during hormone-dependent programmed cell death in Drosophila is regulated by Broad-Complex. J Cell Biol. 2002;157:985–96.
Yang C, Lin X-W, Xu W-H. Cathepsin L participates in the remodeling of the midgut through dissociation of midgut cells and activation of apoptosis via caspase-1. Insect Biochem Mol Biol. 2017;82:21–30.
Liu H, Wang J, Li S. E93 predominantly transduces 20-hydroxyecdysone signaling to induce autophagy and caspase activity in Drosophila fat body. Insect Biochem Mol Biol. 2014;45:30–9.
Arrese EL, Soulages JL. Insect Fat Body: Energy, Metabolism, and Regulation. Annu Rev Entomol. 2010;55:207–25.
Schoofs L, De Loof A, Van Hiel MB. Neuropeptides as Regulators of Behavior in Insects. Annu Rev Entomol. 2017;62:35–52.
Martelli C, Pech U, Kobbenbring S, Pauls D, Bahl B, Sommer MV, et al. SIFamide Translates Hunger Signals into Appetitive and Feeding Behavior in Drosophila. Cell Rep. 2017;20:464–78.
Mustard JA, Pham PM, Smith BH. Modulation of motor behavior by dopamine and the D1-like dopamine receptor AmDOP2 in the honey bee. J Insect Physiol. 2010;56:422–30.
Tsao C-H, Chen C-C, Lin C-H, Yang H-Y, Lin S. Drosophila mushroom bodies integrate hunger and satiety signals to control innate food-seeking behavior. eLife. 2018;7:e35264.
Regna K, Kurshan PT, Harwood BN, Jenkins AM, Lai C-Q, Muskavitch MAT, et al. A critical role for the Drosophila dopamine D1-like receptor Dop1R2 at the onset of metamorphosis. BMC Dev Biol. 2016;16:15.
French AS, Simcock KL, Rolke D, Gartside SE, Blenau W, Wright GA. The role of serotonin in feeding and gut contractions in the honeybee. J Insect Physiol. 2014;61:8–15.
Silva B, Goles NI, Varas R, Campusano JM. Serotonin Receptors Expressed in Drosophila Mushroom Bodies Differentially Modulate Larval Locomotion. PLoS ONE. 2014;9:e89641.
Hoyer SC, Eckart A, Herrel A, Zars T, Fischer SA, Hardie SL, et al. Octopamine in Male Aggression of Drosophila. Curr Biol. 2008;18:159–67.
Mizunami M, Unoki S, Mori Y, Hirashima D, Hatano A, Matsumoto Y. Roles of octopaminergic and dopaminergic neurons in appetitive and aversive memory recall in an insect. BMC Biol. 2009;7:46.
Roeder T, Seifert M, Kähler C, Gewecke M. Tyramine and octopamine: Antagonistic modulators of behavior and metabolism: Octopamine and Tyramine in Insects. Arch Insect Biochem Physiol. 2003;54:1–13.
Zhao X. Progress in understanding hormonal regulation during the postembryonic development of Helicoverpa armigera. J Integr Agric. 2020;19:1417–28.
Park SG, Schimmel P, Kim S. Aminoacyl tRNA synthetases and their connections to disease. Proc Natl Acad Sci USA. 2008;105:11043–9.
Naora H, Naora H. Involvement of ribosomal proteins in regulating cell growth and apoptosis: Translational modulation or recruitment for extraribosomal activity? Immunol Cell Biol. 1999;77:197–205.
Fellner SK, Rybczynski R, Gilbert LI. Ca2+ signaling in prothoracicotropic hormone-stimulated prothoracic gland cells of Manduca sexta: Evidence for mobilization and entry mechanisms. Insect Biochem Mol Biol. 2005;35:263–75.
Jindra M, Palli SR, Riddiford LM. The Juvenile Hormone Signaling Pathway in Insect Development. Annu Rev Entomol. 2013;58:181–204.
Li K, Tian L, Guo Z, Guo S, Zhang J, Gu S-H, et al. 20-Hydroxyecdysone (20E) Primary Response Gene E75 Isoforms Mediate Steroidogenesis Autoregulation and Regulate Developmental Timing in Bombyx. J Biol Chem. 2016;291:18163–75.
Yin VP, Thummel CS. Mechanisms of steroid-triggered programmed cell death in Drosophila. Semin Cell Dev Biol. 2005;16:237–43.
Tian L, Ma L, Guo E, Deng X, Ma S, Xia Q, et al. 20-hydroxyecdysone upregulates Atg genes to induce autophagy in the Bombyx fat body. Autophagy. 2013;9:1172–87.
Beebe K, Lee W-C, Micchelli CA. JAK/STAT signaling coordinates stem cell proliferation and multilineage differentiation in the Drosophila intestinal stem cell lineage. Dev Biol. 2010;338:28–37.
Watabe T, Miyazono K. Roles of TGF-β family signaling in stem cell renewal and differentiation. Cell Res. 2009;19:103–15.
Kléber M, Sommer L. Wnt signaling and the regulation of stem cell function. Curr Opin Cell Biol. 2004;16:681–7.
Choi NH, Lucchetta E, Ohlstein B. Nonautonomous regulation of Drosophila midgut stem cell proliferation by the insulin-signaling pathway. Proc Natl Acad Sci USA. 2011;108:18702–7.
Li Y-L, Yao Y-X, Zhao Y-M, Di Y-Q, Zhao X-F. The steroid hormone 20-hydroxyecdysone counteracts insulin signaling via insulin receptor dephosphorylation. J Biol Chem. 2021;296:100318.
Sano H, Nakamura A, Texada MJ, Truman JW, Ishimoto H, Kamikouchi A, et al. The Nutrient-Responsive Hormone CCHamide-2 Controls Growth by Regulating Insulin-like Peptides in the Brain of Drosophila melanogaster. PLoS Genet. 2015;11:e1005209.
Di Y-Q, Han X-L, Kang X-L, Wang D, Chen C-H, Wang J-X, et al. Autophagy triggers CTSD (cathepsin D) maturation and localization inside cells to promote apoptosis. Autophagy. 2021;17:1170–92.
Yamanaka N, Rewitz KF, O’Connor MB. Ecdysone Control of Developmental Transitions: Lessons from Drosophila Research. Annu Rev Entomol. 2013;58:497–516.
Rusten TE, Lindmo K, Juhász G, Sass M, Seglen PO, Brech A, et al. Programmed Autophagy in the Drosophila Fat Body Is Induced by Ecdysone through Regulation of the PI3K Pathway. Dev Cell. 2004;7:179–92.
Liu X, Dai F, Guo E, Li K, Ma L, Tian L, et al. 20-Hydroxyecdysone (20E) Primary Response Gene E93 Modulates 20E Signaling to Promote Bombyx Larval-Pupal Metamorphosis. J Biol Chem. 2015;290:27370–83.
Kayukawa T, Jouraku A, Ito Y, Shinoda T. Molecular mechanism underlying juvenile hormone-mediated repression of precocious larval–adult metamorphosis. Proc Natl Acad Sci USA. 2017;114:1057–62.
Yin VP, Thummel CS. A balance between the diap1 death inhibitor and reaper and hid death inducers controls steroid-triggered cell death in Drosophila. Proc Natl Acad Sci USA. 2004;101:8022–7.
Bryant B, Zhang Y, Zhang C, Santos CP, Clem RJ, Zhou L. A lepidopteran orthologue of reaper reveals functional conservation and evolution of IAP antagonists. Insect Mol Biol. 2009;18:341–51.
Repnik U, Stoka V, Turk V, Turk B. Lysosomes and lysosomal cathepsins in cell death. Biochim Biophys Acta. 2012;1824:22–33.
Yang H, Zhang R, Zhang Y, Liu Q, Li Y, Gong J, et al. Cathepsin-L is involved in degradation of fat body and programmed cell death in Bombyx mori. Gene. 2020;760:144998.
Gui ZZ, Lee KS, Kim BY, Choi YS, Wei YD, Choo YM, et al. Functional role of aspartic proteinase cathepsin D in insect metamorphosis. BMC Dev Biol. 2006;6:49.
Wang L-F, Chai L-Q, He H-J, Wang Q, Wang J-X, Zhao X-F. A cathepsin L-like proteinase is involved in moulting and metamorphosis in Helicoverpa armigera. Insect Mol Biol. 2010;19:99–111.
Sahdev S, Taneja TK, Mohan M, Sah NK, Khar AK, Hasnain SE, et al. Baculoviral p35 inhibits oxidant-induced activation of mitochondrial apoptotic pathway. Biochem Biophys Res Commun. 2003;307:483–90.
Kumarswamy R, Seth RK, Dwarakanath BS, Chandna S. Mitochondrial regulation of insect cell apoptosis: Evidence for permeability transition pore-independent cytochrome-c release in the Lepidopteran Sf9 cells. Int J Biochem Cell Biol. 2009;41:1430–40.
Huang J, Lv C, Hu M, Zhong G. The Mitochondria-Mediate Apoptosis of Lepidopteran Cells Induced by Azadirachtin. PLoS ONE. 2013;8:e58499.
Yoshida H, Kong Y-Y, Yoshida R, Elia AJ, Hakem A, Hakem R, et al. Apaf1 Is Required for Mitochondrial Pathways of Apoptosis and Brain Development. Cell. 1998;94:739–50.
Lee G, Park JH. Programmed cell death reshapes the central nervous system during metamorphosis in insects. Curr Opin Insect Sci. 2021;43:39–45.
Smagghe G, Vanhassel W, Moeremans C, De Wilde D, Goto S, Loeb MJ, et al. Stimulation of Midgut Stem Cell Proliferation and Differentiation by Insect Hormones and Peptides. Ann N Y Acad Sci. 2005;1040:472–5.
Nelliot A, Bond N, Hoshizaki DK. Fat-body remodeling in Drosophila melanogaster. Genesis. 2006;44:396–400.
Jia Q, Liu S, Wen D, Cheng Y, Bendena WG, Wang J, et al. Juvenile hormone and 20-hydroxyecdysone coordinately control the developmental timing of matrix metalloproteinase–induced fat body cell dissociation. J Biol Chem. 2017;292:21504–16.
Zhang Y, Lu Y-X, Liu J, Yang C, Feng Q-L, Xu W-H. A Regulatory Pathway, Ecdysone-Transcription Factor Relish-Cathepsin L, Is Involved in Insect Fat Body Dissociation. PLoS Genet. 2013;9:e1003273.
Xie T, Spradling AC. decapentaplegic Is Essential for the Maintenance and Division of Germline Stem Cells in the Drosophila Ovary. Cell. 1998;94:251–60.
Li H, Qi Y, Jasper H. Dpp Signaling Determines Regional Stem Cell Identity in the Regenerating Adult Drosophila Gastrointestinal Tract. Cell Rep. 2013;4:10–8.
Brock CK, Wallin ST, Ruiz OE, Samms KM, Mandal A, Sumner EA, et al. Stem cell proliferation is induced by apoptotic bodies from dying cells during epithelial tissue maintenance. Nat Commun. 2019;10:1044.
Johnston LA, Sanders AL. Wingless promotes cell survival but constrains growth during Drosophila wing development. Nat Cell Biol. 2003;5:827–33.
Perez-Garijo A, Martin FA, Struhl G, Morata G. Dpp signaling and the induction of neoplastic tumors by caspase-inhibited apoptotic cells in Drosophila. Proc Natl Acad Sci USA. 2005;102:17664–9.
Khalsa O, Yoon J, Torres-Schumann S, Wharton KA. TGF-β/BMP superfamily members, Gbb-60A and Dpp, cooperate to provide pattern information and establish cell identity in the Drosophila wing. Development. 1998;125(14):2723–34.
Bangi E, Wharton K. Dpp and Gbb exhibit different effective ranges in the establishment of the BMP activity gradient critical for Drosophila wing patterning. Dev Biol. 2006;295:178–93.
Ryoo HD, Gorenc T, Steller H. Apoptotic Cells Can Induce Compensatory Cell Proliferation through the JNK and the Wingless Signaling Pathways. Dev Cell. 2004;7:491–501.
Goberdhan DCI, Wilson C. JNK, cytoskeletal regulator and stress response kinase? A Drosophila perspective Bioessays. 1999;20:1009–19.
Peirce MJ, Mitchell SN, Kakani EG, Scarpelli P, South A, Shaw WR, et al. JNK signaling regulates oviposition in the malaria vector Anopheles gambiae. Sci Rep. 2020;10:14344.
Yoo B, Kim H, Chen X, Shen W, Jang JS, Stein SN, et al. 20-hydroxyecdysone (20E) signaling regulates amnioserosa morphogenesis during Drosophila dorsal closure: EcR modulates gene expression in a complex with the AP-1 subunit. Jun Biol Open. 2021;10:bio058605.
Ling L, Raikhel AS. Serotonin signaling regulates insulin-like peptides for growth, reproduction, and metabolism in the disease vector Aedes aegypti. Proc Natl Acad Sci USA. 2018;115:E9822–31.
Kim D, Langmead B, Salzberg SL. HISAT: a fast spliced aligner with low memory requirements. Nat Methods. 2015;12:357–60.
Pertea M, Pertea GM, Antonescu CM, Chang T-C, Mendell JT, Salzberg SL. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat Biotechnol. 2015;33:290–5.
Trapnell C, Pachter L, Salzberg SL. TopHat: discovering splice junctions with RNA-Seq. Bioinformatics. 2009;25:1105–11.
Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15:550.
Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, et al. Gene Ontology: tool for the unification of biology. Nat Genet. 2000;25:25–9.
Xiong G-H, Xing L-S, Lin Z, Saha TT, Wang C, Jiang H, et al. High throughput profiling of the cotton bollworm Helicoverpa armigera immunotranscriptome during the fungal and bacterial infections. BMC Genomics. 2015;16:321.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 32000345, 31872297).
Author information
Authors and Affiliations
Contributions
QM and XG designed the experiment. XG and RS collected the experimental tissues, and XG conducted the transcriptome analysis. XG and PW performed RT–qPCR and drafted the manuscript. JZ, QQ, HZ and QM revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Table S1. All samples for the transcriptome of cotton bollworm at different stages using RNA-seq.
Additional file 2:
Table S2. Summary of clean reads mapped to the reference H. armigera genome.
Additional file 3: Table S3.
All DEGs of H. armigera at different stages.
Additional file 4:
Table S4. DEGs (BL5D2_vs_BL5W0; FL5D2_vs_FL5W0; FL5W0_vs_FL5D5) identified by GO enrichment analysis.
Additional file 5:
Table S5. Genes related to regulation of juvenile hormone and ecdysone signaling pathways.
Additional file 6:
Table S6. Genes related to neuropeptides and hormone (or neuropeptide) receptors.
Additional file 7:
Table S7. Genes related to autophagy.
Additional file 8:
Table S8. Genes related to apoptosis and proliferation.
Additional file 9:
Table S9. Genes related to the insulin-signaling pathway.
Additional file 10:
Table S10. All primers for qPCR.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Gao, X., Zhang, J., Wu, P. et al. Conceptual framework for the insect metamorphosis from larvae to pupae by transcriptomic profiling, a case study of Helicoverpa armigera (Lepidoptera: Noctuidae). BMC Genomics 23, 591 (2022). https://doi.org/10.1186/s12864-022-08807-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-022-08807-y