
Abstract
The songs of birds are complex signals that may have several functions and vary widely among species. Different ecological, behavioural and morphological factors, as well as phylogeny, have been associated as predictors of the evolution of song structure. However, the importance of differences in development, despite their relevance, has seldom been considered. Here, we analysed the evolution of song in two families of songbirds that differ in song development, manakins (suboscines) and cardinals (oscines), with their phylogeny, morphology, and ecology. Our results show that song characteristics had higher phylogenetic signal in cardinals than in manakins, suggesting higher evolutionary lability in the suboscines. Body mass was the main predictor of song parameters in manakins, and together with habitat type, had a major effect on cardinals’ song structure. Precipitation and altitude were also associated with some song characteristics in cardinals. Our results bring unexpected insights into birdsong evolution, in which non-learners (manakins) revealed greater evolutionary lability than song learners (cardinals).
Similar content being viewed by others


Background
Birdsong is one of nature's most complex and interesting behaviours and one whose evolution is strongly influenced by sexual selection. Songs are used for many purposes, such as attracting mates or territory defence, so that greater intensity of sexual selection promotes the evolution of more exaggerated and elaborated signals [1,2,3]. Besides, as songs are directly involved in reproduction, divergence in their structure may lead to speciation, which may result from increases or decreases in complexity or performance since variation in complexity-related song traits is sufficient to promote reproductive isolation and speciation [4, 5].
Suboscines and oscines are the two major groups of passerine birds capable of song production, and they tend to differ in variability and elaboration of their songs mostly due to anatomic differences in their sound production organ, the syrinx, their brain structure, and the type of vocal development, innate vs. learned [2, 6]. Oscine birds possess complex syringeal anatomy, while suboscines have a simpler vocal apparatus [6,7,8]. Suboscine songs are considered innate and stereotyped, while oscine songs are socially learned and can be extremely variable [2, 9]. Considering these differences in development, it is expected that a higher degree of diversification and faster speciation should occur in oscines since their songs are more susceptible to variation due to copy errors and geographical variation, both consequences of song learning [2, 10], known as the ‘song learning’ hypothesis [5]. On the other hand, a slower and more limited variation of songs should be expected in suboscines, as they are more constrained in song variation than song learners. However, a comparison of speciation and discrimination rates between Central American oscines and suboscines revealed an opposite pattern, in which suboscine species appear to be more sensitive to changes in song structure than oscine birds [11].
Although most of the song is produced in the birds’ vocal organ, this is modulated throughout the vocal tract and can be affected by other morphological attributes [12, 13]. Body size is a major influence on the frequency range of sound, in which larger birds produce narrower bandwidths and lower frequencies [14,15,16,17,18,19,20]. This pattern can be explained by the physics of sound: larger birds tend to have larger syrinx and longer vocal tracts, resulting in the production of sounds with lower frequencies [14, 17, 21, 22]. So far, research on the effects of body size on song structure has been done predominantly on oscine birds due to the great variation in their song structure. Still, recent studies with a wider range of passerine species have generally confirmed the predicted relation between body size and sound frequency, including for suboscine species [23, 24].
Bill size and shape can also limit the characteristics of sound production, as smaller beaks were associated with higher-frequency songs, independently of body size [24]. However, results have been mixed so far, with some studies finding associations between beak size and song structure [16, 17, 25], and others not [16, 21, 25, 26].
Both natural and sexual selection have been shown to affect song evolution [13, 27,28,29,30,31]. Environmental characteristics such as precipitation and vegetation cover may be associated with sound propagation and, consequently, influence the evolution of sound production. The ‘acoustic adaptation’ hypothesis predicts that species that live in closed forests should produce lower song frequencies than those that live in open habitats due to the impact of vegetation on song propagation, causing attenuation, scattering and reverberation of sounds [32,33,34,35]. Although this hypothesis has been validated for some groups [33, 35], recent studies with a broad sample of passerine species have shown that such prediction cannot be generalised since there was no evidence for a correlation between sound frequency and habitat type at a wider scale [23, 36,37,38]. Similar to habitat cover, precipitation can also affect song transmission and, consequently, song structure since it relates to environmental noise and vegetation density [39].
Sexual selection has been considered to be higher in species that inhabit or breed in high latitudes and in species that present migratory behaviour [2, 30, 34]. In both situations, birds experience stronger sexual selection because of shorter mate choice periods [30, 34, 40, 41]. If sexual selection is working on the evolution of some song characteristics, it is expected that higher latitude and migration will explain some of their variations. Altitude is also related to song variables, such as syllable rate, peak frequency, song length and song elaborateness. The association between elevation gradients and song characteristics relates to differences in habitat structure, climate, and species diversity, so birds that inhabit lower altitudes have longer and louder songs with higher syllable rates [42,43,44].
While comparative evolutionary studies on birdsong in suboscine and oscine species are rare, the differences between these two major groups can provide useful insights into song's evolution and its role in speciation [8, 13, 31, 45]. We aim to contribute to filling this gap and identify the factors that may drive the evolution of birdsong and their putative dependence on neuro-anatomical and developmental constraints. We analysed the song evolution of two groups of New World passerines: Pipridae (manakins), a group of suboscine species, and Cardinalidae, a group of oscine birds. Manakins inhabit mostly tropical rainforests of Central and South America [46, 47], while cardinals have a broader distribution from North to South America. These families vary widely in song structure (e.g. Figure 1 and Supporting information, Figures 4 and 5), and ecological and life-histories characteristics [48, 49]. At the same time, both are speciose monophyletic groups with a similar number of species, and the availability of recently published molecular phylogenies allows for the phylogenetic testing of birdsong evolution [46, 50, 51]. Given the enormous effort that would be necessary for comparing all representatives of oscines and suboscines with statistical resolution at this level, we chose to analyse manakins and cardinals as they differ in many aspects related to song production and evolution as a first step for an understanding of the weight of differences in sound motor control and development, together with morphology, ecology, life history and phylogeny.

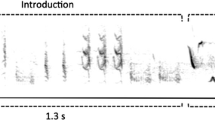
Song spectrograms of cardinal and manakin males reveal the contrast between two of the heaviest species of both families, with larger beak sizes (above) and two of the smallest species (below), with distinct song structures. Images from Birds of the World
By analysing these two distinct families in a comparative approach, we evaluated different assumptions for song evolution: (a) Acoustic adaptation hypothesis (AAH)—we predict that species found in closed habitats, with highest amount of annual precipitation will produce lower frequencies and fewer syllables per song due to differences in song propagation between open and closed vegetation; (b) Morphological constraints on song—we expect to find a negative relation between body mass and high frequency and syllable rate, as vocal tract and beak size correlates positively with body size in manakins and cardinals, affecting sound production; (c) Sexual selection hypothesis—we predict that stronger sexual selection will occur in species with shorter breeding seasons, that is, those breeding in higher altitudes and those that are migratory, resulting in more complex songs; (d) Phylogenetic effect on song structure and AAH—we predict that manakins should have a more limited song evolution and as a consequence present lower evolutionary lability in song characteristics in comparison with cardinals.
Materials and Methods
Song measurements
Recordings were obtained from the Macaulay Library of Natural Songs, the National Sound Archive (British Library), and the Jacques Vielliard Neotropical Music Library of the State University of Campinas (FNJV). Only males were considered, as the female song is present in just a few species. We used a maximum number of 20 recordings per species (each from a different bird), and three songs from each record were considered for measuring sound parameters. Songs were defined and selected in the recordings as being stereotyped sequences of syllables repeated in series, separated by larger time intervals. We then averaged the results per individual and then per species. Song parameters were measured in Raven Pro 1.6.3 [52] with an established spectrogram set: window type Hann, Fourier transform length of 512 samples, 50% overlap, and window overlap size of 256 samples. Each song was selected manually, and then several acoustic parameters were automatically extracted: peak frequency (frequency with maximum power within the song), bandwidth (calculated by the difference between the log10-transformed maximum and minimum frequencies in a selected song) [22], and song length. The number of syllables per song was counted manually, and they were defined as vocal elements tied together in a short time (s) window [53]. Syllable rate (number of syllables per song/song length (s), was calculated from there. In total, 2,433 songs from 979 individuals of 89 species were analysed (1,273 songs from 542 individuals and 44 species in Cardinalidae, and 1,160 songs from 437 individuals and 45 species in Pipridae). Peak frequency was log-transformed according to Weber’s law of song perception for both families, while song length and number of syllables for cardinals, syllable rate, number of syllables and song length for manakins were log-transformed to achieve data normality (Supporting Information). We tested for the correlation between all song variables to avoid overlapping or crossing information.
Molecular data and phylogenetic inference
The species selection of Pipridae and Cardinalidae was performed according to available molecular data and phylogenies (Pipiridae: Leite et al. 2021; Cardinalidae: Barker et al. 2013), together with acoustic, morphological, and ecological information for these species [46]. For Pipridae, we used the recently published phylogenomic topology available in Leite et al. (2021), which includes 51 species of manakins (out of the 55 species recognized [54], and three outgroup species (Pyroderus scutatus, Onychorhynchus coronatus, and Pachyramphus minor). Leite et al. (2021) analysed different datasets, and for our work, we used the consensus topology that resulted from the Maximum Likelihood (ML) analysis of the concatenated dataset of UCE data with 95% complete loci [51]. This topology is available in the Supporting Information.
To obtain a topology for Cardinalidae inferred with similar methods (ML) as the ones used for the Pipridae dataset, we compiled a supermatrix of available DNA sequences for 6 loci and 44 ingroup taxa (out of the 49 species currently recognized by Winkle et al. 2020) and three outgroups (Mitrospingus oleagineus, Lamprospiza melanoleuca, and Orthogonys chloricterus). All molecular data were retrieved from GenBank. The complete list of taxa, voucher information and Genbank accession numbers of all analysed samples and genes are provided in the Supporting Information. The loci comprised segments of two mitochondrial genes: cytochrome b (Cytb, 47 sequences) and NADH dehydrogenase subunit II (ND2, 46 sequences); and segments of four nuclear genes: recombination activating protein 1 (Rag1, 31 sequences), intron 9 of the aconitase 1 gene (ACO-I9, 30 sequences), intron 5 of the beta-fibrinogen gene (FGB-I5, 31 sequences), intron 2 of the myoglobin gene (MB-I2, 30 sequences). We tried to retrieve sequences from the same individual; when this was not possible, sequences of a different individual from the same species were used. We used PipeLogeny in R 4.0.2 [55, 56] to automatise the preparation of the input files for several phylogenetic tools, which were then run in their separate software applications. We aligned sequences with MAFFT 7.310 [57] and used PartitionFinder2 2.1.1 [58] on the CIPRES Science Gateway [59] to identify the best-fit partitioning scheme and models of sequence evolution, which were tested under Bayesian Information Criterion (BIC). The input configuration file included 12 partitions, corresponding to one partition for every intronic region (ACO-I9, FGB-I5, MB-I2) and individual codon positions for each of the Cytb, ND2 and Rag1 markers. This represents the most finely partitioned scheme for our dataset. We used the ‘greedy’ algorithm (heuristic search) with branch lengths estimated as ‘unlinked’. We performed ML analyses using RAxMLHPC2- HYBRID-AVX (Stamatakis 2014) in CIPRES, setting 1,000 replicates of rapid bootstrap and 20 initial random trees. We assessed nodal support via the autoMRE option to generate bootstrap replicates until convergence was reached and to draw bipartitions onto the best-scoring ML tree. The resulting consensus topology is available in the Supporting Information.
For comparative analyses, we imported the consensus trees of both families (with branch lengths based on genetic distances) using the ape package in R [56, 60].
Ecological, life-history and morphological traits
We assembled life-history, ecological and morphological data for each species from the literature. The predictor variables selected were habitat cover, mean altitude (m), midpoint latitude, precipitation, body mass and beak size for both families and breeding latitude and migration for cardinals [23, 54, 61,62,63,64]. Habitat cover was categorised into (0) open, (1) semi-open, and (2) closed environments; breeding latitude was divided into (0) tropical, (1) tropical to temperate, (2) temperate, and (3) temperate to polar regions; migratory movements were defined as (0) sedentary, (1) short-distance migrant, and (2) medium/long distances migrant [62, 65]. Annual precipitation and midpoint latitude were obtained according to range maps available at NatureServe and Birdlife International [66].
Body mass (g) information has been mostly retrieved from Dunning [67] which this measurement is used for most species and each sex. Additional data regarding the body mass of the remaining species (four of 45 manakin species and three of 44 cardinal species) was obtained from three other sources [23, 62, 65]. Beak size measurements consist of the first principal component score that explains 84% of the variation of several bill measurements from a Principal Component Analysis, namely beak width and depth at the nares and two beak lengths at the nares and culmen [61]. Since body mass and beak size were highly correlated (Pearson coefficient (r) = 0.90, p = 0.00, see Figure 6 in Supporting information), only body mass was included as a predictor in the models since we can account for sex differences in this trait, and because body mass is a significant trait to consider when analysing song characteristics in birds. Migration and breeding latitude were not included in the analysis of manakins since all species are sedentary, and most breed in tropical regions. Matching information about predictors and Pipridae and Cardinalidae phylogenies resulted in a sample of 37 manakin and 38 cardinal species evaluated in this study.
Phylogenetic comparative analysis
To test the association between song characteristics and morphological, ecological and life-history parameters, we conducted a phylogenetic generalised least-squares (PGLS) regression [68] using the caper package [69] in R [56]. To avoid multi-collinearity among the predictors, we tested for the variation inflation factor (VIF) in our models, using the performance package [70] which resulted in a low correlation (VIF < 1.6) between our predictors. In each regression, we evaluated the phylogenetic signal, considering a maximum likelihood estimate (Pagel’s lambda) to correct the intensity of the phylogenetic signal in the regression model [68]. Multiple PGLS regression models were performed, including the complete dataset of predictors according to each group (manakins and cardinals) and each song parameter as the response variable. To determine the degree of the phylogenetic signal of acoustic and morphological parameters, individual analyses were performed to calculate Pagel’s λ, measured with the package phytools, function phylosig in R (version 4.1.3). Moreover, for variables that presented high phylogenetic signal but no significance (p > 0.05), we performed an additional test to compare different models of evolution (Brownian motion, Ornstein-Uhlenbec and Early-burst models) using AIC Geiger package, function fitContinuous [71].
Results
Phylogenetic inference of cardinalidae
The alignment consisted of a total of 7,290 bp. The optimal partitioning scheme that yielded the lowest score for our analyses included eight partitions (see details in the Supplementary Information). The GTR model was determined to be the best-fitting model of substitution for all partitions. The resulting topology largely agrees with the phylogenetic hypothesis proposed by Barker et al. (2013).
Phylogenetic signal
Phylogenetic signals of song (Pagel’s λ—Table 1) were significant for more variables in Cardinals than in Manakins. Peak frequency and song length in piprids had a high and significant phylogenetic signal. In contrast, the number of syllables per song and syllable rate evolved more independently from phylogenetic history. Although bandwidth presented a high lambda (0.891), this parameter was not significant (p = 0.066), which led us to perform an additional test to compare different models of evolution that revealed great evidence for a Brownian motion model. In cardinals, five out of six acoustic parameters had a high phylogenetic signal: peak frequency, frequency bandwidth, number of syllables per song, and syllable rate, indicating they evolved more following the evolutionary history of the group. Only song length had a non-significant phylogenetic signal, suggesting an evolution independent from phylogeny for this trait. This is contrary to manakins, for which song length is precisely the trait with a higher phylogenetic signal. The morphological traits, beak size and body mass, revealed a high phylogenetic signal in both families (Table 1, Fig. 2, and Figures 7–10 in Supporting information).

Phylogenetic trees of manakins (above) and cardinals (below) represent a strong phylogenetic signal (λ) of body mass (left) and beak size (right) in both families. Colour gradients represent the trait values. Images and taxonomic nomenclature from Birds of the World
Manakins
Body mass was the only significant predictor for song parameters in manakins (Fig. 3 and Table 2), being negatively associated with syllable rate (partial βst = − 0.528, P = 0.005; model λ = 0.00) and positively associated with song length (partial βst = 0.561, P = 0.019; model λ = 0.947). Thus, heavier manakin species sing longer songs with a slower syllable rate than smaller birds. Both peak frequency and bandwidth were not affected by body size. There was also no effect of habitat cover, mean altitude, midpoint latitude, or precipitation on any song parameters in manakins (Supporting Information).

Scatterplot with regression lines illustrating associations predictors and song traits in manakins (green) and cardinals (red); these are linear associations that do not consider the effects of other variables. For a better comparative illustration, the cardinal’s syllable rate has been log transformed to be represented in the same scale as manakins in the two families' graph sets. Significative results are highlighted in bold
Cardinals
In cardinals, body mass and habitat cover were the main predictors of song characteristics (Fig. 3 and Table 2). Body mass was negatively associated with peak frequency (partial βst = − 0.556, P = 0.006; model λ = 1.00) and syllable rate (partial βst = − 0.439, P = 0.012; model λ = 0.334) and positively associated with song length (partial βst = 0.442 P = 0.031; model λ = 0.749). Smaller cardinal species (according to body mass) tend to sing at higher peak frequencies and with faster syllable rates and sing shorter songs than heavier species.
Cardinals inhabiting closed environments showed lower peak frequencies (partial βst = − 0.427, P = 0.008; model λ = 1.00), slower syllable rates (partial βst = − 0.381, P = 0.018; model λ = 0.344), and longer songs (partial βst = 0.513, P = 0.004; model λ = 0.749) than species living in open and semi-open habitats. Moreover, the mean altitude showed a significant positive effect on syllable rate (partial βst = 0.367, P = 0.012; model λ = 0.344), while precipitation was positively related to peak frequency (partial βst = 0.284, P = 0.046; model λ = 1.00), and negatively associated with bandwidth (partial βst = -0.336, P = 0.022; model λ = 1.00). Cardinal species that inhabit areas with lower rain rates sing at lower peak frequencies and wider bandwidths than cardinals living in areas with higher rain rates. Migratory movements and breeding latitude did not show significative results as predictors of song parameters in cardinals.
Discussion
Various evolutionary forces can influence bird song evolution in diverse ways. The Pipridae and Cardinalidae constitute two speciose families representing suboscine and oscine birds, which greatly differ in their ability to evolve and produce songs. We found that the evolution of song characteristics had a higher phylogenetic signal in cardinals than in manakins. The evolution of manakins’ songs was not influenced by ecological factors, with body weight being the only factor explaining the existing variation. In cardinals, both body size and several ecological factors were predictors of the evolution of their songs.
Effects of phylogeny on song evolution
We found that phylogenetic relationships explain a considerable amount of song characteristic variation in both groups. That was more the case for cardinals, where peak frequency, bandwidth, number of syllables, and syllable rate all had high phylogenetic signal, meaning songs evolved following phylogeny. Two evaluated song parameters had a significantly high phylogenetic signal in manakins: peak frequency and song length, while syllable rate, bandwidth, and number of syllables in song did evolve more independently from phylogeny. If natural or sexual selection acts strongly upon song, then the phylogenetic signal is expected to be high [72].
It is not possible to discern whether song contributed to speciation, as would be required for testing the song learning hypothesis [73], which proposes a major role of song in speciation in oscine species due to being learnt. Still, our findings do not align with what one would expect from the song learning hypothesis [67], which proposes that song should have a major role in speciation in oscine species due to being learnt. If that was the case, a lower phylogenetic signal should be observed in oscines (resulting from higher evolutionary lability) compared to suboscines since [1] song can diverge in several directions (speed, frequency, complexity) in song learners and [2] a significant divergence is needed for the pre-mating isolation processes of speciation [2, 10]. However, mixed results have been found regarding the strength of phylogenetic signal in song parameters in oscines and suboscines [11, 74,75,76,77,78]. Recent findings considering song learners and vocal non-learners indicate that high phylogenetic signals exist for both song and call structure [74]. Moreover, shared ancestry is reflected in other determinant components of song production, such as the syringeal constitution and vocal tract. Our results reinforce the argument that oscines and suboscines can have high phylogenetic signals in song structure besides being learned or innate.
Song constrained by morphology (Magic Traits)
We found that body mass is strongly associated with song characteristics in manakins and cardinals. This finding aligns well with other studies that report an association between body mass and song characteristics, particularly in the frequency domain [16, 22,23,24, 79, 80]. In both families, syllable rate and song length were related to body mass, and heavier birds sang longer songs with a slower syllable rate, concordant with previous findings for other groups of birds [14, 20, 25, 80]. Also, the relation between syllable rate and body mass might reflect the association between beak size and syllable rate, as both morphological parameters are positively related in manakins and cardinals. The limitation imposed by beak size in song has been shown for other groups, as larger beaks will face a trade-off between force and speed at the skeleton and muscle levels, and so larger beaks are associated with slower frequency changes and trill rates [12, 14, 18,19,20].
In cardinals, body mass was also negatively related to peak frequency. Smaller and less heavy species sing at higher frequencies than heavier ones. This is expected considering the physical properties of resonator size and frequency output. Also, larger birds with longer and wider vocal tract may have difficulty reaching high frequencies or, simply, those are not the frequencies for an optimal use of the energy they can produce vocally [80, 81]. It is also worth mentioning that body mass was not related to peak frequency in manakins. The range of frequencies is very similar between the two groups despite the different structural morphology of their syrinxes [82, 83]. Two complementary explanations can be given for this. It is possible that frequency modulation is not dependent on the syrinx in suboscines. However, it is unknown how the variation in the air sac pressure is controlled [84, 85], which can be less dependent on body size than in oscines. Besides, the range of body mass variation is very small in manakins compared with cardinals, which can prevent a possible tendency to be discernible. Our results support the importance of considering diverse groups such as oscine and suboscine birds to understand better the constraints of distinct song development on birdsong evolution [23, 86].
Ecology and behaviour (AAH hypothesis & Sexual selection)
The ecological and behavioural predictors we considered were only associated with song characteristics in cardinals. Habitat cover significantly predicted peak frequency, in which species living in closed habitats with lower peak frequencies than those inhabiting open or semi-open areas. This agrees with the sensory drive (acoustic adaptation) hypothesis [32, 35, 37, 87] and aligns with findings from other studies on the environmental effects of sound propagation and animal communication [34, 88, 89]. Also, cardinals living in open habitats had higher syllable rates and sang shorter songs. Species that inhabit closed forests face greater signal transmission challenges due to physical obstacles, being more efficient in those conditions to sing at lower frequencies, with slower syllable rates, and longer songs. In manakins, there is no sufficient variation in habitat cover when considering the type of habitat (open and closed) to allow for an effective test of its effect on song evolution [62].
Precipitation also influenced cardinals’ songs. Species living in rainier areas sang with narrower bandwidths and higher peak frequencies. These results also agree with the acoustic adaptation hypothesis since species inhabiting areas with more noise (rain) face greater challenges for signal transmission [34]. Cardinals may have facilitated song transmission in these environments by singing with higher peak frequencies and narrower bandwidths.
In birds, shorter breeding periods were associated with more intense sexual selection due to greater breeding synchrony, and it was predicted that sexual selection would increase with latitude and elevation [90, 91]. Following this hypothesis, a positive relation was found between elevation and sexual selection in several studies [91,92,93], while the opposite was revealed in cardueline finches [44, 94]. Our results for Cardinals agree with the above hypothesis since species that inhabit higher altitudes sing with faster syllable rates. Species breeding at higher altitudes will also experience shorter breeding seasons, which could explain its effects on song structure.
This association of elevation and latitude could not be tested in manakins, as most of the species breed in lowland tropical regions. The absence of significant effects of ecological factors associated with song structure in manakins may be due to the reduced variation in ecology since all members are tropical species. It would be interesting to widen this analysis by considering suboscines with broader distribution and varied ecology and life histories, as it has rarely been attempted for suboscine species [26, 95].
Conclusion
We found that the evolution of song in manakins and cardinals was strongly influenced by body weight, indicating that size is a major constraining factor in song production [20, 38, 96]. Peak frequency decreased with size in cardinals; syllable rate also decreased with size in both groups, while song length increased with body mass in manakins and cardinals. A different pattern is apparent when we consider ecological variables. The cardinal’s song evolution was also influenced by several ecological factors, which was not the case for manakins’ whose song was not affected by ecology. Habitat cover and precipitation affected the frequency characteristics of songs. Syllable rate decreased with habitat cover, as an adaptive response to scattering and reverberation, and increased with altitude. This increase of a performance variable with altitude agrees with previous findings for eleven species of island birds [42]. Both migration and breeding latitude had no influence on song characteristics, indicating that cardinals’ songs appear not to have been affected by sexual selection, at least for the variables we considered.
Cardinals’ songs were influenced by ecological factors, but not manakins, which may be explained by the greater ability to adjust sounds to better transmission conditions in song learners compared with sub-oscine species with no or very limited learning [97]. Suboscines are also more responsive to slight variations in songs, which may be important for species recognition [33].
In addition, manakins have particularly elaborated dances and colouration, while songs are comparatively much simpler, so it is possible that sexual selection acted more in traits that could suffer greater elaboration. Supporting this, brain size has been associated with display complexity in manakins [98, 99]. It would be worth widening the research on the evolution of song in groups of birds differing in song development to better understand vocal constraints in oscine and suboscine species [100, 101]. Besides ecological and morphological traits, the phylogenetic relationship also determines song structure in manakins and cardinals. Opposite from what would be expected according to the learning hypothesis, cardinals, as representative of song-learning species, showed higher phylogenetic signal among their song structure traits compared to manakins with innate songs.
Our findings show the importance of evaluating evolutionary constraints and modulators at a smaller taxonomic scale in parallel to wider research. More extensive studies can hide patterns that exist in some taxonomic groups, which may have specific and unique evolutionary histories that would be overridden by large-scale patterns. Furthermore, we highlight that more studies on the evolution of song in suboscines are needed since they constitute a very diverse and poorly studied group.
Availability of data and materials
All data generated or analysed during this study are included in this published article (and its supplementary information files).
References
Nowicki S, Searcy WA. Song function and the evolution of female preferences: why birds sing, why brains matter. Ann N Y Acad Sci. 2004;1016:704–23.
Catchpole CK, Slater PJ. Bird Song Biological Themes and Variations. New York: Cambridge University Press; 2008.
Robinson CM, Creanza N. Species-level repertoire size predicts a correlation between individual song elaboration and reproductive success. Ecol Evol. 2019;9(14):8362–77.
Cardoso GC, Mota PG. Speciational evolution of coloration in the genus Carduelis. Evolution. 2008;62(4):753–62.
Cardoso GC, Mamede AT, Atwell JW, Mota PG, Ketterson ED, Price TD. Song frequency does not reflect differences in body size among males in two oscine species. Ethology. 2008;114(11):1084–93.
Suthers RA, Zollinger SA. Producing song: the vocal apparatus. Ann N Y Acad Sci. 2004;1016:109–29.
Goller F, Suthers RA. Role of syringeal muscles in controlling the phonology of bird song. J Neurophysiol. 1996;76(1):287–300.
Goller F, Love J, Mindlin G. Different frequency control mechanisms and the exploitation of frequency space in passerines. Ecol Evol. 2021;11(11):6569–78.
Beecher MD, Brenowitz EA. Functional aspects of song learning in songbirds. Trends Ecol Evol. 2005;20(3):143–9.
Kroodsma DE. Geographic variation in songs of the Bewick’s wren: a search for correlations with avifaunal complexity. Behav Ecol Sociobiol. 1985;16(2):143–50.
Freeman BG, Montgomery GA, Schluter D. Evolution and plasticity: Divergence of song discrimination is faster in birds with innate song than in song learners in Neotropical passerine birds. Evolution. 2017;71(9):2230–42.
Podos J, Nowicki S. Beaks, Adaptation, and Vocal Evolution in Darwin’s Finches. Bioscience. 2004;54(6):501–10.
Mejias MA, Roncal J, Imfeld TS, Boisen S, Wilson DR. Relationships of song structure to phylogenetic history, habitat, and morphology in the vireos, greenlets, and allies (Passeriformes: Vireonidae). Evolution. 2020;74(11):2494–511.
Podos J. Correlated evolution morphology and vocal signal structure in Darwin’s finches. Nature. 2001;409:185–8.
Fletcher NH. A simple frequency-scaling rule for animal communication. J Acoust Soc Am. 2004;115(5 Pt 1):2334–8.
Podos J, Southall JA, Rossi-Santos MR. Vocal mechanics in Darwin’s finches: correlation of beak gape and song frequency. J Exp Biol. 2004;207(Pt 4):607–19.
Derryberry EP, Seddon N, Claramunt S, Tobias JA, Baker A, Aleixo A, et al. Correlated evolution of beak morphology and song in the neotropical woodcreeper radiation. Evolution. 2012;66(9):2784–97.
Friedman NR, Harmackova L, Economo EP, Remes V. Smaller beaks for colder winters: Thermoregulation drives beak size evolution in Australasian songbirds. Evolution. 2017;71(8):2120–9.
Porzio NS, Repenning M, Fontana CS. Do beak volume and bite force influence the song structure of sympatric species of seedeaters (Thraupidae:Sporophila)? Emu - Austral Ornithology. 2018;119(1):71–8.
Demery A-JC, Burns KJ, Mason NA. Bill size, bill shape, and body size constrain bird song evolution on a macroevolutionary scale. Ornithology. 2021;138(2):1–11.
Porzio NS, Repenning M, Fontana CS. Evolution of beak size and song constraints in neotropical seedeaters (Thraupidae: Sporophila). Acta Ornithologica. 2019;53(2):173–80.
Cardoso GC, Abreu JM, Archer J, Crottini A, Mota PG. Independent evolution of song diversity and song motor performance in canaries, goldfinches and allies indicates clade-specific trade-offs in birdsong. Evolution. 2020;74(6):1170–85.
Mikula P, Valcu M, Brumm H, Bulla M, Forstmeier W, Petruskova T, et al. A global analysis of song frequency in passerines provides no support for the acoustic adaptation hypothesis but suggests a role for sexual selection. Ecol Lett. 2021;24(3):477–86.
Friis JI, Sabino J, Santos P, Dabelsteen T, Cardoso GC, Jennions MD. Ecological adaptation and birdsong: how body and bill sizes affect passerine sound frequencies. Behav Ecol. 2022;33(4):798–806.
García NC, Tubaro PL. Dissecting the roles of body size and beak morphology in song evolution in the “blue” cardinalids (Passeriformes: Cardinalidae). Auk. 2018;135(2):262–75.
Derryberry EP, Seddon N, Derryberry GE, Claramunt S, Seeholzer GF, Brumfield RT, et al. Ecological drivers of song evolution in birds: Disentangling the effects of habitat and morphology. Ecol Evol. 2018;8(3):1890–905.
Ballentine B. Vocal performance influences female response to male bird song: an experimental test. Behav Ecol. 2004;15(1):163–8.
Cardoso GC, Mota PG, Depraz V. Female and male serins (Serinus serinus) respond differently to derived song traits. Behav Ecol Sociobiol. 2007;61(9):1425–36.
Holveck M-J, Riebel K. Preferred songs predict preferred males: consistency and repeatability of zebra finch females across three test contexts. Anim Behav. 2007;74(2):297–309.
Cardoso GC, Hu Y, Mota PG. Birdsong, sexual selection, and the flawed taxonomy of canaries, goldfinches and allies. Anim Behav. 2012;84(1):111–9.
Mason NA, Shultz AJ, Burns KJ. Elaborate visual and acoustic signals evolve independently in a large, phenotypically diverse radiation of songbirds. Proc Biol Sci. 2014;281(1788):20140967.
Morton ES. Ecological Sources of Selection on Avian Sounds. Am Nat. 1975;109(965):17–34.
Seddon N. Ecological adaptation and species recognition drives vocal evolution in neotropical suboscine birds. Evolution. 2005;59(1):200–15.
Weir JT, Wheatcroft DJ, Price TD. The role of ecological constraint in driving the evolution of avian song frequency across a latitudinal gradient. Evolution. 2012;66(9):2773–83.
Vélez A, Gall MD, Fu J, Lucas JR, Patek S. Song structure, not high-frequency song content, determines high-frequency auditory sensitivity in nine species of New World sparrows (Passeriformes: Emberizidae). Funct Ecol. 2014;29(4):487–97.
Bertelli ST, Tubaro PL. Body mass and habitat correlates of song structure in a primitive group of birds. Biol J Lin Soc. 2002;77(4):423–30.
Boncoraglio G, Saino N. Habitat structure and the evolution of bird song: a meta-analysis of the evidence for the acoustic adaptation hypothesis. Funct Ecol. 2006;21(1):134–42.
Friis JI, Dabelsteen T, Cardoso GC. Contingency and determinism in the evolution of bird song sound frequency. Sci Rep. 2021;11(1):11600.
DeWalt S, Schnitzer S, Chaves JA, Bongers F, Burnham R. Annual rainfall and seasonality predict pan-tropical patterns of liana density and basal area. Biotropica. 2010;43(3):309–17.
Wei C, Price TD, Liu J, Alström P, Zhang Y. The evolutionary origin of variation in song length and frequency in the avian family Cettiidae. J Avian Biol. 2017;48(10):1295–300.
Singh P, Price TD. Causes of the latitudinal gradient in birdsong complexity assessed from geographical variation within two Himalayan warbler species. Ibis. 2015;157:511–27.
Robert A, Lengagne T, Melo M, Gomez D, Doutrelant C. Evolution of vocal performance and song complexity in island birds. J Avian Biol. 2022;1:1–12.
Peters MK, Hemp A, Appelhans T, Behler C, Classen A, Detsch F, et al. Predictors of elevational biodiversity gradients change from single taxa to the multi-taxa community level. Nat Commun. 2016;7:13736.
Snell-Rood EC, Badyaev AV. Ecological gradient of sexual selection: elevation and song elaboration in finches. Oecologia. 2008;157(3):545–51.
Gomes ACR, Funghi C, Soma M, Sorenson MD, Cardoso GC. Multimodal signalling in estrildid finches: song, dance and colour are associated with different ecological and life-history traits. J Evol Biol. 2017;30(7):1336–46.
Klicka J, Burns K, Spellman GM. Defining a monophyletic Cardinalini: a molecular perspective. Mol Phylogenet Evol. 2007;45(3):1014–32.
Heindl M, Winkler H. Interacting effects of ambient light and plumage color patterns in displaying Wire-tailed Manakins (Aves, Pipridae). Behav Ecol Sociobiol. 2003;53(3):153–62.
Gomes AC, Sorenson MD, Cardoso GC. Speciation is associated with changing ornamentation rather than stronger sexual selection. Evolution. 2016;70(12):2823–38.
Shultz AJ, Burns KJ. The role of sexual and natural selection in shaping patterns of sexual dichromatism in the largest family of songbirds (Aves: Thraupidae). Evolution. 2017;71(4):1061–74.
Barker FK, Burns KJ, Klicka J, Lanyon SM, Lovette IJ. New insights into New World biogeography: An integrated view from the phylogeny of blackbirds, cardinals, sparrows, tanagers, warblers, and allies. Auk. 2015;132(2):333–48.
Leite RN, Kimball RT, Braun EL, Derryberry EP, Hosner PA, Derryberry GE, et al. Phylogenomics of manakins (Aves: Pipridae) using alternative locus filtering strategies based on informativeness. Mol Phylogenet Evol. 2021;155:107013.
Bioacoustics KLYCfC. Raven Pro: Interactive Sound Analysis Software (Version 1.6.3). Ithaca: The Cornell Lab of Ornithology; 2021. https://doi.org/10.2173/bow.cardin1.01.
Kawaji T, Fujibayashi M, Abe K. Goal-directed and flexible modulation of syllable sequence within birdsong. Nat Commun. 2024;15(3419):1–12.
Winkler DW, Billerman SM, Lovette IJ. Cardinals and Allies (Cardinalidae), version 1.0. In: Birds of the World. Billerman SM, Keeney BK, Rodewald PG, Schulenberg TS, Eds. Ithaca, NY, USA: Cornell Lab of Ornithology; 2020. https://doi.org/10.2173/bow.cardin1.01.
Muñoz-Pajarez AJ, Belluardo F, Cocca W, Crottini A. PipeLogeny: An automated pipeline for phylogenetic reconstruction. https://sites.google.com/site/pipelogenydownload/2019.
R Core Team. R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing; 2022.
Katoh KK, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 2013;30:772–80.
Lanfear R, Frandsen PB, Wright AM, Senfeld T, Calcott B. PartitionFinder 2: new methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol Biol Evol. 2017;34(3):772–3.
Miller MA, Pfeiffer W, Schwartz T. Creating the CIPRES science gateway for inference of large phylogenetic trees. Gateway Computing Environments Workshop (GCE). 2010:1–8.
Paradis E, Claude J, Strimmer K. APE: analyses of phy- logenetics and evolution in R language. Bioinformatics. 2004;20:289–90.
Pigot AL, Sheard C, Miller ET, Bregman TP, Freeman BG, Roll U, et al. Macroevolutionary convergence connects morphological form to ecological function in birds. Nat Ecol Evol. 2020;4(2):230–9.
Winkler DW, Billerman SM, Lovette IJ. Manakins (Pipridae), version 1.0. In: S. M. Billerman BKK, P. G. Rodewald, and T. S. Schulenberg, editor. Birds of the World. Ithaca: Cornell Lab of Ornithology; 2020.
IUCN. The IUCN Red List of Threatened Species 2022.
Dunning JBJ. CRC handbook of avian body masses. Boca Raton: CRC Press; 1993.
Winkler DW, Billerman SM, Lovette IJ. Cardinals and Allies (Cardinalidae), version 1.0. In: S. M. Billerman BKK, P. G. Rodewald, and T. S. Schulenberg, editor. Birds of the World. Ithaca: Cornell Lab of Ornithology; 2020.
Cooney CR, Varley ZK, Nouri LO, Moody CJA, Jardine MD, Thomas GH. Sexual selection predicts the rate and direction of colour divergence in a large avian radiation. Nat Commun. 2019;10(1):1773.
Price T. Speciation in Birds. Greenwood Village (Colorado): Roberts & Co; 2008. p. 2008.
Freckleton RP, Harvey PH, Pagel M. Phylogenetic analysis and comparative data: a test and review of evidence. Am Nat. 2002;160(6):712–26.
Orne D, Freckleton RP, Thomas G, Petzoldt T, Fritz S, Issac N, et al. The Caper package: comparative analysis of phyloge-netics and evolution in R. 2013.
Lüdecke D, Ben-Shachar M, Patil I, Waggoner P, Makowski D. performance: An R Package for Assessment, Comparison and Testing of Statistical Models. J Open Source Soft. 2021;6(60):3139.
Harmon LJ, Weir JT, Brock CD, Glor RE, Challenger W. GEIGER: investigating evolutionary radiations. Bioinformatics. 2008;24(1):129–31.
Revell LJ, Harmon LJ, Collar DC. Phylogenetic signal, evolutionary process, and rate. Syst Biol. 2008;57(4):591–601.
Price T, Young S, Paul ER. Speciation in birds. 1st ed. Greenwood Village: Roberts and Co; 2008. x, 470 p. : ill. (some col.), maps p.
Arato J, Fitch WT. Phylogenetic signal in the vocalizations of vocal learning and vocal non-learning birds. Philos Trans R Soc Lond B Biol Sci. 1836;2021(376):20200241.
Päckert M, Martens J, Kosuch J, Nazarenko AA, Veith M. Phylogenetic signal in the song of crests and kinglets (Aves: Regulus). Evolution. 2003;57(3):616–29.
Price JJ, Lanyon SM. A robust phylogeny of the oropendolas: polyphyly revealed by mitochondrial sequence data. Auk. 2002;119:335–48.
Price JJ, Lanyon SM. Reconstructing the evolution of complex bird song in the oropendolas. Evolution. 2002;56(7):1514–29.
Mason NA, Burns KJ, Tobias JA, Claramunt S, Seddon N, Derryberry EP. Song evolution, speciation, and vocal learning in passerine birds. Evolution. 2017;71(3):786–96.
Ryan MJ, Brenowitz EA. The role of body size, phylogeny, and ambient noise in the evolution of bird song. Am Nat. 1985;26(1):87–100.
Mason NA, Burns K. The effect of habitat and body size on the evolution of vocal displays in Thraupidae (tanagers), the largest family of songbirds. Biol J Lin Soc. 2015;114:538–51.
Nowicki S, Marler P. How Do Birds Sing? Music Percept. 1988;5(4):391–426.
Prum RO. Syringeal morphology, phylogeny, and evolution of the neotropical manakins (Aves, Pipridae). New York: American Museum of Natural History; 1992.
Ames PL. The morphology of the syrinx in Passerine birds. New Haven: Peabody Museum of Natural History, Yale University; 1971.
Amador A, Goller F, Mindlin GB. Frequency modulation during song in a suboscine does not require vocal muscles. J Neurophysiol. 2008;99(5):2383–9.
Beckers GJ, Suthers RA, ten Cate C. Mechanisms of frequency and amplitude modulation in ring dove song. J Exp Biol. 2003;206(Pt 11):1833–43.
Pearse WD, Morales-Castilla I, James LS, Farrell M, Boivin F, Davies TJ. Global macroevolution and macroecology of passerine song. Evolution. 2018;72(4):944–60.
Hansen P. Vocal learning: its role in adapting sound structures to long-distance propagation and a hypothesison its evolution. Anim Behav. 1979;27:1270–1.
Blumstein DT, Turner AC. Can the acoustic adaptation hypothesis predict the structure of Australian birdsong? Acta Ethologica. 2005;8(1):35–44.
Tobias JA, Aben J, Brumfield RT, Derryberry EP, Halfwerk W, Slabbekoorn H, et al. Song divergence by sensory drive in Amazonian birds. Evolution. 2010;64(10):2820–39.
Spottiswoode C, Møller AP. Extrapair paternity, migration, and breeding synchrony in birds. Behav Ecol. 2004;15:41–57.
Albrecht T, Schnitzer J, Kreisinger J, Exnerová A, Bryja J, Munclinger P. Extrapair paternity and the opportunity for sexual selection in long-distant migratory passerines. Behav Ecol. 2007;18:477–86.
Bonier F, Eikenaar C, Martin PR, Moore IT. Extrapair paternity rates vary with latitude and elevation in emberizid sparrows. Am Nat. 2014;183(1):54–61.
LaBarbera K, Hayes KRR, Langhans KE, Lacey EA. Elevation affects extra-pair paternity but not a sexually selected plumage trait in dark-eyed juncos. Behavior Ecol Sociobiol. 2019;73(6):1–12.
Badyaev AV. Altitudinal variation in sexual dimorphism: a new pattern and alternative hypotheses. Behav Ecol. 1997;8(6):675–90.
Acero-Murcia AC, Raposo do Amaral F, de Barros FC, da Silva Ribeiro T, Miyaki CY, Maldonado-Coelho M. Ecological and evolutionary drivers of geographic variation in songs of a Neotropical suboscine bird: The Drab-breasted Bamboo Tyrant (Hemitriccus diops, Rhynchocyclidae). Ornithology. 2021;138(2):1–15.
Ryan MJ, Brenowitz EA. The role of body size, phylogeny, and ambient noise in the evolution of bird song. Am Nat. 1985;126:87–100.
Kroodsma DE, Hamilton D, Sanchez JE, Byers BE, Fandino-Marino H, Stemple DW, et al. Behavioral evidence for song learning in the Suboscine Bellbirds (Procnias spp.; Cotingidae). Wilson J Ornithol. 2013;125:1–14.
Day LB, Helmhout W, Pano G, Olsson U, Hoeksema JD, Lindsay WR. Correlated evolution of acrobatic display and both neural and somatic phenotypic traits in Manakins (Pipridae). Integr Comp Biol. 2021;61(4):1343–62.
Day LB, Lindsay WR. Associations between Manakin display complexity and both body and brain size challenge assumptions of allometric correction: a response to Gutierrez-Ibanez et al. (2016). Brain Behav Evol. 2016;87(4):227–31.
Goller F, Riede T. Integrative physiology of fundamental frequency control in birds. J Physiol Paris. 2013;107(3):230–42.
Goller F. Vocal athletics – from birdsong production mechanisms to sexy songs. Anim Behav. 2022;184:173–84.
Acknowledgements
We are grateful to Portuguese National Funds, through FCT (Foundation for Science and Technology), which support N.S.P. (PhD. Grant 2020.06170.BD) and AC (Research contract: 2020.00823.CEECIND/CP1601/CT0003). We would also like to thank Matthew Medler from Macaulay Library, who shared over 1,000 songs for this study. We are also grateful to the National Sound Archive (British Library) and Fonoteca Neotropical Jacques Vielliard (FNJV) for providing access to their databases. Work supported by National Funds through FCT-Fundação para a Ciência e a Tecnologia in the scope of the project UIDP/50027/2020.
Funding
The Portuguese National Funds, through FCT (Foundation for Science and Technology) supported N.S.P. PhD scholarship (PhD. Grant 2020.06170.BD) and AC research funding (contract: 2020.00823.CEECIND/CP1601/CT0003).
Author information
Authors and Affiliations
Contributions
N.S.P. and P.G.M. conceived and designed the study. N.S.P. analysed the acoustic data and the statistical models. A.C. and R.N.L. conducted the phylogenetic simulations. N.S.P. and P.G.M. drafted the initial version of the manuscript and all authors contributed substantially to later versions of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
12862_2024_2298_MOESM3_ESM.tiff
Supplementary Material 3: Figure 6. Scatterplot reveals the positive correlation between body mass and beak size for manakins and cardinals, showing Pearson’s coefficient and significance.
12862_2024_2298_MOESM4_ESM.tiff
Supplementary Material 4: Figure 7. Pipridae phylogenetic trees represent the phylogenetic signalof peak frequencyand bandwidth. Colour gradients represent the trait values.
12862_2024_2298_MOESM5_ESM.tiff
Supplementary Material 5: Figure 8. Pipridae phylogenetic trees represent the phylogenetic signalof syllable rateand song length. Colour gradients represent the trait values.
12862_2024_2298_MOESM6_ESM.tiff
Supplementary Material 6: Figure 9. Cardinalidae phylogenetic trees represent the phylogenetic signalof peak frequencyand bandwidth. Colour gradients represent the trait values.
12862_2024_2298_MOESM7_ESM.tiff
Supplementary Material 7: Figure 10. Cardinalidae phylogenetic trees represent the phylogenetic signalof syllable rateand song length. Colour gradients represent the trait values.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Porzio, N.S., Crottini, A., Leite, R.N. et al. Song determined by phylogeny and body mass in two differently constrained groups of birds: manakins and cardinals. BMC Ecol Evo 24, 109 (2024). https://doi.org/10.1186/s12862-024-02298-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12862-024-02298-z