
Abstract
Background
Species and structural diversity are important for understanding the formation of forest communities, key ecological processes, and improving forest ecological functions and services, but their spatial characteristics have received little attention. Based on the spatial relationships among neighbouring trees, we proposed to divide trees within a structural unit into 15 structural types, and used the univariate distributions of the uniform angle index (W), mingling (M), and dominance (U), along with four common species diversity indices, to analyse the diversity of structural types in natural forests near the Tropic of Cancer.
Results
Only a portion of clumped class maintained aggregation, most exhibited a random pattern. Species mixture increased exponentially across distribution classes, and abundance and richness exhibited an initial increase followed by a slight decrease. The distribution patterns of mixture classes varied from highly clustered to random, and M distributions gradually shifted from an inverted J-shaped curve to a J-shaped curve. Abundance and richness exhibited an exponential distribution, whereas the Shannon–Wiener index increased linearly. The W distribution of differentiation classes approximated a normal distribution, whereas M distributions exhibited a J shape. The U distribution of each structure type was approximately 0.2.
Conclusions
These results reveal the species and structural diversity characteristics of trees at the structural type level and expand our knowledge of forest biodiversity. The new method proposed here should significantly contribute to biodiversity monitoring efforts in terrestrial ecosystems, and suggests that higher standards for the simulation and reconstruction of stand structure, as well as thinning in near-natural forests, is warranted.
Similar content being viewed by others

Introduction
Forests provide primary habitats for a variety of species, as well as essential ecosystem services to humans [1]. Climate change, environmental pollution, frequent natural disasters (e.g. drought, freezing, and wildfires), human disturbance, resource exploitation (e.g. logging, grazing, and changing vegetation types), and land use have resulted in the fragmentation and degradation of many forests [2,3,4,5]. This has significantly impacted the distribution patterns, species composition, growth, and diversity of forests, thus reducing ecological stability and resistance to disturbance [1, 6]. Forest environment (e.g., soil, microclimate), fauna and subsurface components (e.g. litter) associated with this process have also been substantially impacted [7,8,9,10]. Rapid losses in forest biodiversity have attracted much attention and generated considerable concern [6], and numerous nature reserves, among other protected areas, have been established to protect biodiversity and provide models for forest management [11]. An accurate understanding of the quantitative characteristics of biodiversity at different scales would facilitate assessments of forest conservation status, management efforts, and monitoring conservation and restoration efforts.
Biodiversity refers to forest components (species, functional traits, and genes), structure, and functions/processes [12, 13]. Among them, species and structural diversity have emerged as the most important aspects of forest biodiversity [14,15,16,17], and play functional roles (e.g. dead wood) in forest ecosystems. Species are the primary component of forest ecosystems, and species composition largely determines forest productivity, spatial allocation, and carbon storage, among other characteristics [18]. Forest structure plays an important role in regulating species composition, seed dispersal, establishment, interactions between neighbouring trees, habitat type, resource use (e.g. light and nutrition), and other ecological processes related to forest growth [16, 19,20,21]. Forest structure also strongly affects ecological functions (e.g. biomass, carbon storage, and productivity), stress resistance, successional trajectories, the suitability of silvicultural methods, and stand microenvironments [2, 9, 22, 23]. While on a small scale, species and structure, together with other environmental factors (e.g. local climate and soil), directly or indirectly influence forest stability [1, 2]. Most biodiversity studies focus on one or the other, and rarely address both factors simultaneously [9]. Studies considering both species and structural diversity may improve our understanding of the processes and mechanisms underlying forest community development [24].
Both species and structural diversity are related to spatial scale [17, 25]. From small to large, species diversity can be divided into α, β, γ, and δ levels, and each of them has been well documented. α diversity refers to species diversity at the community, stand, or quadrat level [22]. β diversity describes species turnover between communities, whereas γ and δ describe broader scale patterns [26]. The studies of structural diversity mainly focus on the non-spatial properties of diameter class/ontogenetic stages, lifeforms, chronosequences and quadrat [1, 2, 4, 9, 17, 24, 26]. To date, however, few spatial analyses involving species have been applied in biodiversity studies [27], and few studies have highlighted the utility of analysing spatial diversity [14]. Not to mention the relationship between species and structures below the quadrat level.
The introduction of stand spatial structure parameters (SSSPs), such as the uniform angle index (W), mingling (M), and dominance (U), has enhanced analyses of spatial structure [17, 28,29,30,31,32]. These metrics describe the distributional attributes of the four nearest neighbouring trees in a structural unit or frame around reference tree i, including the degree of species mixing and size differentiation (Fig. 1). They are independent of each other and each index has the same range values [29], which just meets the conditions for tree classification. That is, trees in any quadrat can be further divided into five distribution classes, five mixture classes and five differentiation classes, which represent species groups with different aggregation, mixing and differentiation, respectively. We called them structural types in this study. Obviously, structural type level is smaller than quadrat and allows the analysis of species and structural diversity in space.

Explanations for stand spatial structural parameters. Structural unit contains a reference trees i and four nearest neighbors and their spatial relationships can be described by uniform angle index (W), mingling (M) and dominance (U), respectively. In the top panels, patterns change from regularity to clump with increasing W value. In the medium panels, reference tree i has more heterospecific neighbors with increasing M value. In the bottom panels, reference tree is less dominant in structural unit with increasing U value
Since the properties of structural types have yet to be explored. This study mainly focused on the structural diversity (i.e. tree point distribution, species mixing, and size differentiation) and species diversity of structural types in natural forests. Conspecific aggregation is prevalent in natural forests [33,34,35,36,37], we assumed that the aggregation of conspecifics increases with increasing W value (Fig. 1), that is, the aggregation and interspecific isolation of distribution classes increase while species diversity and size differentiation decrease (Hypothesis 1). While other types of interactions (e.g. facilitative or neutral) in forest ecosystems cannot be ignored, intraspecific exclusion is thought to be more prevalent [33, 38]. Strong intraspecific competition will lead to size differentiation, self-thinning and promotes the formation of species and structural diversity [39]. As a result, the distribution pattern of highly mixed trees may become more uniform and their differentiation is more obvious (Hypothesis 2). Similarly, highly differentiated trees might have a greater mixture and a more uniform pattern (Hypothesis 3). We used dataset from dozens of natural forest plots near the Tropic of Cancer (Guangxi, China) to test these hypotheses and interpreted the results.
Materials and methods
Study sites
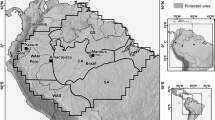
The study sites were located in the primary national nature reserves of Guangxi Zhuang Autonomy Region, China. From south to north, these include the Shiwandashan (SWDS), Damingshan (DMS), Dayaoshan (DYS), Cenwanglaoshan (CWLS), Mulun (ML), Yachang (YC), Jiuwanshan (JWS), and Huaping (HP) reserves. The reserves are located near or on The Tropic of Cancer (23° 26ʹ N) (Fig. 2), and thus represent the northern tropics and southern and mid-subtropics. The region is characterised by abundant rainfall and long, hot summers that last from May to September. Autumn (October–November), winter (December–January), and spring (February–April) are all short and relatively warm. The reserves are distributed throughout the mountains of Guangxi, and support montane forest ecosystems. They are composed of evergreen broad-leaved mixed forests, broad-leaved mixed forests, and evergreen and deciduous broad-leaved mixed forests (Table S1). These forests are highly structurally complex and rich in species, making them a global biodiversity hotspot [1, 17]. The vegetation is mainly composed of secondary forests, with some plantations and a small amount of primary and mature forests. All of the study reserves are dominated by broadleaf trees. Due to strict regulations, there has been little anthropogenic disturbance to the vegetation since the reserves were established.

Locations of the study sites. Green dots represent quadrats and Arabic numeral represents the number of quadrat in each reserve. SWDS = Shiwandashan nature reserve, DMS = Damingshan nature reserve, DYS = Dayaoshan nature reserve, CWLS = Cenwanglaoshan nature reserve, ML = Mulun nature reserve, YC = Yachang nature reserve, JWS = Jiuwanshan nature reserve, and HP = Huaping nature reserve
Plot establishment
Between 2016 and 2022, 34 standard plots were established in the study reserves. Quadrat locations were selected based on topographical and forest characteristics, including species composition, structure, and growth status (Table 1). All quadrats were established in natural forests representing different developmental stages, including early, mid-, and late-successional. Some stands have a known date of origin, whereas others are poorly documented (Table 1). Nevertheless, all stands represent locally common forest types and have not recently been impacted by disturbance. The quadrats were randomly located on mountains. With the aid of a total station, we first delineated the perimeters of the quadrats and then divided each quadrat into n 20 × 20 m quadrats. We measured the coordinates of all trees in each quadrat with a DBH ≥ 1 cm. Each tree was recorded, identified to species (by doctor Lei Wang from College of Forestry, Guangxi University non-doposited), and marked. Furthermore, we noted the growth status of each tree, including characteristics such as stem form, vitality, disease, and insect damage. Information about non-biological habitat attributes (e.g. soil type and thickness, litter, rock, and coarse woody debris), disturbance, and stand age was also collected. Finally, a global positioning system was used to determine the geographical position of each quadrat.
Data analyses
We calculated the parameters for each tree in all quadrats [29], and divided the results into 15 structural types (5 each for W, M, and U) using thresholds of 0.0, 0.25, 0.50, 0.75, and 1.00 to delineate the classes (Fig. 1). Correspondingly, structural types refer to the various levels of Wi, Mi and Ui, which we refer to as distribution classes, mixture classes, and differentiation classes, respectively (Fig. 1, Table 2). We then calculated the parameters for each structural type, and considered their univariate distributions as metrics of spatial diversity [28, 29]. We used the wilcox.test function to test for significant differences among the univariate distributions. Because the mean value of W is substantially influenced by the number of individuals (N) per quadrat, we calculated 95% confidence intervals for each structural type using the formula \({{\text{y}}}_{0.05}=0.5\pm 1.96\upsigma \times 0.21034{N}^{-0.48872}\) [40], and considered the result to represent the range of random distribution. We then determined whether trees were distributed in an aggregated, random, or regular fashion. For better explain and compare the results with other studies, we further considered Wi = 0.00 and 0.25 as regular class (ReC), Wi = 0.50 as random class (RaC), and Wi = 0.75 and 1.00 as clumped class (CC). Similarly, we considered Mi = 0.00 and 0.25 to be low-mixture class (LMC), Mi = 0.50 to be medium-mixture class (MMC), and Mi = 0.75 and 1.00 to be highly mixed class (HMC), and Ui = 0.00 and 0.25 as dominant class (DC), Ui = 0.50 as moderately dominant class (MDC), and Ui = 0.75 and 1.00 as weak (non-dominant) class (WC).
We quantified the species diversity of each structural type using two basic diversity indices: species richness (SR) and N. (number of individual) Furthermore, we calculated an index that describes the relationship between SR and N (i.e. the Shannon–Wiener index; Hʹ) and a second index that characterises the consistency of N among species (i.e. the Pielou evenness index; EH) (Table 2). These indices were calculated using the ‘vegan’ package in R [42]. The wilcox.test function was used to test for significant differences in species diversity among structural types, and Jaccard’s similarity coefficient and Venn diagrams were used to describe correlations in species composition (Table 2).
Results
Spatial diversity of distribution classes
The W distribution of ReC, RaC, and half of CC (W = 0.75) exhibited unimodal patterns, that is, the values first increased with increasing W and then decreased (Fig. 3a–d). The univariate distributions of the other CC group exhibited an initial increase, then decreased before increasing again, and values were relatively variable in the mid and high W grades (Fig. 3e). Across all five distribution classes, trees had significantly smaller values when W was low (p < 0.001). M distributions had a similar pattern that increased exponentially, and their mean values gradually decreased from 0.83 to 0.63 (Fig. 3f–j). Distribution classes had similar U distributions, with a mean value of approximately 0.2. ReC exhibited broader distributions than RaC and CC (Fig. 3k–o).

W distribution (a-e), M distribution (f-j) and U distribution (k-o) of distribution classes. ReC = regular class, RaC = random class, CC = clumped class, W = uniform angle index, M = mingling, U = dominance. ***, **, * and NS dominate p < 0.001, p < 0.01, p < 0.05 and p ≥ 0.05, respectively
Spatial diversity of mixture classes
The W values of the low-mixture class exhibited a positive trend, peaking at 0.93 at W = 1.0, with a coefficient of variation as high as 1.22 (Fig. 4a). In the remaining classes, W distributions gradually approximated a normal distribution, and distributions became increasingly narrow (Fig. 4b–e). The M distribution changed substantially among different mixture classes, shifting from an inverted J-shaped to J-shaped distribution, and the mean value increased from 0.05 to 0.97 (Fig. 4f–j). The five mixture classes exhibited similar U distributions, with mean values of approximately 0.2 (Fig. 4k–o).

W distribution (a-e), M distribution (f-j) and U distribution (k-o) of mixture classes. LCM = low-mixture class, MMC = medium-mixture class, HMC = highly mixed class, W = uniform angle index, M = mingling, U = dominance.***, **, * and NS dominate p < 0.001, p < 0.01, p < 0.05 and p ≥ 0.05, respectively
Spatial diversity of differentiation classes
All differentiation classes exhibited similar, narrow W distributions that approximated a normal distribution. Mean values increased from 0.01–0.03 at W = 0.0 to 0.20–0.30 at W = 0.25 and 0.50–0.57 at W = 0.50, and then decreased to 0.14–0.22 at W = 0.75 and 0.00–0.09 at W = 1.00 (Fig. 5a–e). M distributions exhibited a slow initial increase followed by a rapid increase, and values increased from around 0.02 to 0.56 (Fig. 5f–j). M grades were similar across differentiation classes, but values differed significantly between M grades (p < 0.01). Aside from the U values of WC, which increased from 0.17 to 0.25, other differentiation classes exhibited relatively stable U values of approximately 0.2 (Fig. 5k–o).

W distribution (a-e), M distribution (f-j) and U distribution (k-o) of differentiation classes. DC = dominant class, MDC = moderately dominant class, WC = clumped class, W = uniform angle index, M = mingling, U = dominance. ***, **, * and NS dominate p < 0.001, p < 0.01, p < 0.05 and p ≥ 0.05, respectively
Species diversity among structural types
As W increased, the mean value of N increased from 22.6 to 2,764.3 and then decreased to 695.03; the differences among structural types were significant. RaC accounted for a much higher proportion of trees than the other classes (Fig. 6a). SR exhibited a similar trend, with a mean that increased from 12.21 to 88.09 and then decreased to 55.24 (Fig. 6b). The mean value of Hʹ increased from 2.17 to 3.16 for ReC and remained high in the remaining structural types (Fig. 6c), whereas EH exhibited the opposite trend (Fig. 6d). N and SR exhibited exponential increases with increased mixture, whereas Hʹ exhibited a significant linear increase (Fig. 6e–g). The EH of HMC was slightly higher and more narrowly distributed than that of the other classes (Fig. 6h). With respect to differentiation classes, the mean values of N, SR, Hʹ, and EH were approximately 1,000, 67, 3.0, and 0.73, respectively (Fig. 6i–l). We observed little variation in SR among differentiation classes, except that the regular and low-mixed classes were both characterised by relatively low SR (Fig. 7a–c). Jaccard similarity coefficients for distribution classes and mixture classes ranged from 0.15–0.99 to 0.35–0.52, while those for the differentiation classes were < 0.16 (Fig. 7d–f).

Species diversity of distribution classes (a-d), mixture classes (e-h) and differentiation classes (i-l). ReC = regular class, RaC = random class, CC = clumped class, LMC = low-mixture class, MMC = medium-mixture class, HMC = highly mixed class, DC = dominant class, MDC = moderately dominant class, WC = weak class, N = number of individual, SR = species richness, Hʹ = Shannon-Wiener index, EH = Pielou’ evenness index. ***, **, * and NS dominate p < 0.001, p < 0.01, p < 0.05 and p ≥ 0.05, respectively

Venn diagrams (a-c) and Jaccard similarity coefficients (d-f) of structural types. ReC = regular class, RaC = random class, CC = clumped class, LMC = low-mixture class, MMC = medium-mixture class, HMC = highly mixedclass, DC = dominant class, MDC = moderately dominant class, WC = weak class
Discussion
Although the concept and characterisation of biodiversity have been widely discussed [12, 13, 16, 43], surprisingly little is known about the spatial characteristics of biodiversity at forest stand scales [31, 44]. In many montane tropical and subtropical forests, even small forest patches may exhibit high structural complexity and a range of growing conditions [45], and biodiversity at each scale is very high [15, 25, 36, 38].
Distribution patterns at the structural type level
In undisturbed or lightly disturbed natural forests, the univariate distribution of uniform angle index is unimodal, which represents a stable pattern [21, 39, 46]. Similar patterns were observed when we classified distribution classes and mixture classes into different structural types (Figs. 3a-d and 4d-e), thereby highlighting stable pattern occur at the stand and sub-stand levels. Random framework (framework when Wi = 0.5) comprises the majority of basal area and abundance in this pattern, and has a competitive advantage compared with other frameworks and is considered crucial for community stability [27]. Residual classes maintained an evident clumped pattern (Table S2), and most clumps comprised the same species, contributed substantially to stand-level aggregation. This implies that, in stands with high tree density, the average scale of aggregation is much larger than a single structural unit. Various biological and abiotic factors, such as habitat heterogeneity, habitat filtering, gap dynamics, niche differentiation, resource management, stress, and human disturbance, can induce aggregation together [4, 34, 36, 37, 47,48,49]. The changes of aggregation with distribution classes and mixture classes (Figs. 3a-d and 4a-e) are consistent with Hypothesis 1 and Hypothesis 2, suggesting that distribution patterns can be predicted by species mixture [50], which provides a new approach for the study of distribution patterns.
Species mixture at the structural type level
The mingling distribution of distribution classes is consistent with that of other natural forests [51], suggesting that hierarchical prediction can be applied to species mixture. In general, species mixture was negatively correlated with aggregation (Fig. 3f-j), while it has a linearly positive correlation with tree sizes [39], indicating that small trees preferred intraspecific aggregation, while large trees preferred interspecific mixing and dispersion, consistent with the mingling-size hypothesis [48, 52, 53]. Many other point pattern studies have shown that aggregation is higher among small and understorey trees compared to large and canopy trees [15, 54, 55], supporting Hypothesis 1. The relationships among distribution pattern, species mixture and tree size can be explained by dispersal limitation and conspecific negative density effects. The way of seeding and regeneration in natural forests largely determines the aggregation of small, conspecific trees, and then intraspecific exclusion and host (disease and insect) invasion lead to mortality and xenogenetic settlement, thus pushing the community towards a more regular pattern comprising multiple species [4, 21, 48]. These effects are important mechanisms by which species coexistence and diversity are maintained in many temperate, tropical, and subtropical forests [38, 49, 56, 57]. In particular, significant habitat heterogeneity near the Tropic of Cancer may enhance aggregation, asymmetric competition and negative density-dependent mortality [37, 47].
The species mixture of mixture classes was consistent with that of the quadrat. Low-mixture class means reference tree i and its neighbours are, by definition, conspecifics (Fig. 1). The frequency distribution of this class was opposite to that of other natural forests [32, 39, 51, 58], implies that tree populations are distributed in patches. Highly mixed class, on the contrary, dominated the number of plants in quadrants, and was often surrounded by heterospecifics (Fig. 4i-j), which may be the direct reason for the high mixing of natural forests near the Tropic of Cancer. Its mixture distribution was similar to that of other species-rich natural forests, but with a larger means, indirectly reflecting the study forests were in a highly mixed state [17]. Medium-mixture class is a transitional type, its neighbour could be conspecific or heterospecific (Figs. 1 and 4h). The distribution of mixture classes shows the change of species relationship in detail.
Size differentiation at the structural type level
There is a negative exponential relationship between dominance and different DBH classes [39], therefore, dominance values represent the advantage of differentiation classes at the stand level. Differentiation classes divided forests near the Tropic of Cancer into groups of trees that were very close to each other in abundance and structural diversity (Fig. 5), but completely different in dominance (Fig. 1) and species composition (Fig. 7f). Their structural diversity approximated those of typical species-rich natural forests [21, 32, 46]. This indicates that trees of different sizes have similar distribution patterns and mixtures at the structural type level, and that these characteristic can be extrapolated to the stand level. The results are contrary to Hypothesis 3, because tree sizes within structural units are relative, and differentiation occurs frequently. Differentiation may occur between conspecifics or heterospecifics, among trees of a particular size class, or between small and large trees, separating the distribution pattern and species mixture of trees of different sizes almost evenly. Distribution classes and mixture classes had similar dominance distributions (Figs. 3k-o, and 4k-o), further emphasizes the fact. Most populations and trees of different DBHs in pine-oak mixed forests and Koran pine broadleaved forests are nearly randomly distributed, and the dominance is only weakly related to uniform angle index [39, 58, 59], which strongly supports our results. Without obvious disturbance, a balanced size differentiation emerges very early [39], which undoubtedly reduce direct competition, thus maximizing the use of three-dimensional spatial resources by the stand. This strategy may facilitate maintenance of species diversity in line with niche differentiation theory [39, 60]. However, large differences in the size of neighbours may obscure the relationship between tree size and spatial structure [61], and stands with different DBH distributions may have similar values of dominance [30]. Dominance and DBH distribution quantify the sizes of tree within different scales, driving forces and community differences may lead to inconsistent results.
Species diversity at the structural type level
Random class includes the major species of the stand, has high diversity, highlighting its critical role in maintaining community stability [27]. Clumped class also includes numerous species and exhibits high diversity, consistent with the biodiversity characteristics of tropical and subtropical forests [15]. Especially, these natural forests typically contain numerous rare species that have small tree sizes and tend to aggregate [34, 36, 37], thereby enhancing community diversity, evenness and aggregation [17]. Natural forests near the Tropic of Cancer contain karst and non-karst sites that are rich in microhabitats [17] and may promote the formation and aggregation of species diversity [41, 45]. Along bioclimatic gradients across Europe, increased SR alters interspecific and intraspecific relationships, and generally increases the intensity of tree aggregation at small scales [4], which supports our results. Higher functional diversity allows individuals to grow in close proximity, thus promoting the complementary use of resources [4]. The low species diversity of regular class suggests that it plays a minor role in our study stands (Fig. 6a-c). In fact, regular distributions can only occur within a small range, as habitats and germplasm resources are variable in time and space. Few reports of regular distributions in natural forests are available.
Most species and individuals were highly mixed and exhibited high evenness, consistent with findings for tropical broadleaf forests in central Vietnam [32]. Some old-growth forests in temperate regions also exhibit similar characteristics [62]. A high degree of isolation reduces intraspecific competition and is beneficial for species coexistence and the maintenance of biodiversity [4, 33, 48]. By contrast, low-mixture class was characterized by variable evenness and low abundance, richness, and Shannon–Wiener diversity, suggesting intraspecific aggregation. Potential conspecific competition drives community succession [39]. The relationship between species diversity and mixture can be modelled using exponential or linear functions. [63] integrated the number of tree species per structural unit into simple mingling formula (Table 2), and [31] integrated the stand spatial structure parameters into diversity indices to evaluate tree spatial diversity. Together with our results, these studies demonstrate that species diversity is closely related to the structure of neighbouring trees.
Differentiation classes had similar levels of species abundance and richness, which explains the high similarity in structural diversity. Hʹ and EH were evenly separated; however, the variability among dominant class was significantly greater than that of weak class. This may be related to the species composition and successional stage of the community. Some old-growth forests have plenty of dominant populations (e.g., YC-8), while early successional communities have low species richness with respect to dominant canopy species (e.g., YC-2, YC-4 YC-5), and slow growing trees require a considerable amount of time to reach the canopy. Conversely, this pattern also indicates that weak class promotes community diversity [6, 15, 64], which in turn suggests that small trees may decrease diversity as they grow into large trees. Close relationship between species diversity and tree size has been found in natural forests around the Tropic of Cancer [17], which strongly supports our results. At the regional scale, however, the relationship between tree size and species diversity may become increasingly random or inverse [65, 66]. Moreover, evenness is much less important than richness for maintaining biodiversity [2]. It is less stable than other diversity indices, suggesting that the interspecific and individual relationships in natural forests are never in equilibrium.
Conclusion
A new method based on the spatial relationship of location, mixture and differentiation of neighbouring trees, is proposed for the classification of tree groups. The method classifies quadrats into 15 structural types to enable spatial analysis of species and structural diversity, different from the way of tree attributes (e.g., population, diameter, stratification, life history stage) or combinations of the spatial structure parameters. We further analysed the biodiversity of structural types in main forest ecosystems near the Tropic of Cancer, and found that differentiation classes contribute equally to species and structural diversity. The diversity approaches to quadrat level and are almost independent of tree size and species. The structural diversity of distribution classes and mixture classes exhibits a clear pattern in which small size of conspecifics favour aggregation while big size of heterospecifics favour random distributions. Relationships between tree size, mixture and distribution pattern are close, and can be well explained by popular forest ecology theories in current. The positive relationship between species diversity and mixture is also very close, and could be predicted by generalized linear model. The new method proposed here greatly contributes to biodiversity monitoring and assessment, and suggest that higher standards for the simulation and reconstruction of stand structure, as well as the thinning of near-natural forests, are warranted. With respect to methods, structural types can be divided into smaller groups, and the characteristics of tree species groups after multiple classifications should be explored in future.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- W:
-
Uniform angle index
- M:
-
Mingling
- U:
-
Dominance
- SSSPs:
-
Stand spatial structure parameters
- SWDS:
-
Shiwandashan nature reserve
- DMS:
-
Damingshan nature reserve
- DYS:
-
Dayaoshan nature reserve
- CWLS:
-
Cenwanglaoshan nature reserve
- ML:
-
Mulun nature reserve
- YC:
-
Yachang nature reserve
- JWS:
-
Jiuwanshan nature reserve
- HP:
-
Huaping nature reserve
- DBH:
-
Diameter at breast height
- N:
-
Number of individuals
- ReC:
-
Regular class
- RaC:
-
Random class
- CC:
-
Clumped class
- LMC:
-
Low-mixture class
- MMC:
-
Medium-mixture class
- HMC:
-
Highly mixed class
- DC:
-
Dominant class
- MDC:
-
Moderately dominant class
- WC:
-
Weak class
- SR:
-
Species richness
- Hʹ:
-
Shannon-Wiener index
- EH :
-
Pielou evenness index
References
Ouyang S, Xiang W, Gou M, Chen L, Lei P, Xiao W, Deng X, Zeng L, Li J, Zhang T, Peng C, Forrester DI, Meyer C. Stability in subtropical forests: The role of tree species diversity, stand structure, environmental and socio-economic conditions. Global Ecol Biogeogr. 2020;30(2):500–13.
Guo Z, Wang X, Fan D. Ecosystem functioning and stability are mainly driven by stand structural attributes and biodiversity, respectively, in a tropical forest in Southwestern China. Forest Ecol Manag. 2021;481:118696.
Arroyo-Rodriguez V, Melo FP, Martinez-Ramos M, Bongers F, Chazdon RL, Meave JA, Norden N, Santos BA, Leal IR, Tabarelli M. Multiple successional pathways in human-modified tropical landscapes: new insights from forest succession, forest fragmentation and landscape ecology research. Biol Rev Camb Philos Soc. 2017;92(1):326–40.
Bastias CC, Truchado DA, Valladares F, Benavides R, Bouriaud O, Bruelheide H, Coppi A, Finér L, Gimeno TE, Jaroszewicz B, Scherer-Lorenzen M, Selvi F, De la Cruz M. Species richness influences the spatial distribution of trees in European forests. Oikos. 2019;129(3):380–90.
Wright SJ. The future of tropical forests. Ann N Y Acad Sci. 2010;1195:1–27.
Li Y, Ye S, Bai W, Zhang G. Species diversity patterns differ by life stages in a pine-oak mixed forest. Dendrobiology. 2022;88:138–49.
Gardner TA, Hernández MIM, Barlow J, Peres CA. Understanding the biodiversity consequences of habitat change: the value of secondary and plantation forests for neotropical dung beetles. J Appl Ecol. 2007;45(3):883–93.
Hume AM, Chen HYH, Taylor AR, Kardol P. Intensive forest harvesting increases susceptibility of northern forest soils to carbon, nitrogen and phosphorus loss. J Appl Ecol. 2018;55(1):246–55.
Ruiz-Jaén MC, Aide TM. Vegetation structure, species diversity, and ecosystem processes as measures of restoration success. Forest Ecol Manag. 2005;218(1–3):159–73.
Cesarz S, Craven D, Auge H, Bruelheide H, Castagneyrol B, Gutknecht J, Hector A, Jactel H, Koricheva J, Messier C, Muys B, O’Brien MJ, Paquette A, Ponette Q, Potvin C, Reich PB, Scherer-Lorenzen M, Smith AR, Verheyen K, Eisenhauer N, Xu X. Tree diversity effects on soil microbial biomass and respiration are context dependent across forest diversity experiments. Global Ecol Biogeogr. 2022;31(5):872–85.
Bílek L, Remeš J, Podrázský V, Rozenbergar D, Diaci J, Zahradník D. Gap regeneration in near-natural European beech forest stands in Central Bohemia – the role of heterogeneity and micro-habitat factors. Dendrobiology. 2013;71:59–71.
Ferris R, Humphrey JW. A review of potential biodiversity indicators for application in British forests. Forestry. 1999;72(4):313–28.
Noss RF. Indicators for monitoring biodiversity: A hierarchical approach. Concervation biology. 1990;4(4):355–64.
Hui G, Pommerening A. Analysing tree species and size diversity patterns in multi-species uneven-aged forests of Northern China. Forest Ecol Manag. 2014;316:125–38.
Feroz S, Mamun A, Kabir ME. Composition, diversity and distribution of woody species in relation to vertical stratification of a tropical wet evergreen forest in Bangladesh. Glob Ecol Conserv. 2016;8:144–53.
McElhinny C, Gibbons P, Brack C, Bauhus J. Forest and woodland stand structural complexity: Its definition and measurement. Forest Ecol Manag. 2005;218(1–3):1–24.
Li Y, Ye S, Luo Y, Yu S, Zhang G. Relationship between species diversity and tree size in natural forests around the Tropic of Cancer. J Forestry Res. 2023;34:1735–45.
Forrester DI, Bauhus J. A review of processes behind diversity—productivity relationships in forests. Curr For Rep. 2016;2(1):45–61.
Fraver S, D’Amato AW, Bradford JB, Jonsson BG, Jönsson M, Esseen P-A, Turnbull L. Tree growth and competition in an old-growth Picea abies forest of boreal Sweden: influence of tree spatial patterning. J Veg Sci. 2014;25(2):374–85.
Jucker T, Koricheva J, Finér L, Bouriaud O, Iacopetti G, Coomes DA, Chen H. Good things take time—Diversity effects on tree growth shift from negative to positive during stand development in boreal forests. J Eco. 2020;108(6):2198–211.
Zhang G, Hui G, Zhao Z, Hu Y, Wang H, Liu W, Zang R. Composition of basal area in natural forests based on the uniform angle index. Ecol Inform. 2018;45:1–8.
Ehbrecht M, Schall P, Ammer C, Seidel D. Quantifying stand structural complexity and its relationship with forest management, tree species diversity and microclimate. Agr Forest Meteorol. 2017;242:1–9.
Giesbrecht IJW, Saunders SC, MacKinnon A, Lertzman KP. Overstory structure drives fine-scale coupling of understory light and vegetation in two temperate rainforest floodplains. Can J Forest Res. 2017;47(9):1244–56.
Mensah S, Salako VK, Assogbadjo AE, Glèlè Kakaï R. Differential responses of taxonomic, structural, and functional diversity to local-scale environmental variation in afromontane forests in South Africa. Trop Conserv Sci. 2018;11:1–13.
He F, Lafrankie JV, Song B. Scale dependence of tree abundance and richness in a tropical rain forest. Malaysia Landscape Ecol. 2002;17(6):559–68.
Dupuy JM, Hernández-Stefanoni JL, Hernández-Juárez RA, Tetetla-Rangel E, López-Martínez JO, Leyequién-Abarca E, Tun-Dzul FJ, May-Pat F. Patterns and correlates of tropical dry forest structure and composition in a highly replicated chronosequence in Yucatan. Mexico Biotropica. 2012;44(2):151–62.
Zhang G, Hui G. Random trees are the cornerstones of natural forests. Forests. 2021;12(8):1046.
Hui G, Zhang G, Zhao Z, Yang A. Methods of forest structure research: a review. Curr For Rep. 2019;5(3):142–54.
Li Y, Hui G, Zhao Z, Hu Y. The bivariate distribution characteristics of spatial structure in natural Korean pine broad-leaved forest. J Veg Sci. 2012;23(6):1180–90.
Petritan AM, Biris IA, Merce O, Turcu DO, Petritan IC. Structure and diversity of a natural temperate sessile oak (Quercus petraea L.) – European Beech (Fagus sylvatica L.) forest. Forest Ecol Manag. 2012;280:140–9.
Zhao Z, Hui G, Liu W, Hu Y, Zhang G. A novel method for calculating stand structural diversity based on the relationship of adjacent trees. Forests. 2022;13:134.
Nguyen HH, Erfanifard Y, Petritan I. Nearest neighborhood characteristics of a tropical mixed broadleaved forest stand. Forests. 2018;9:33.
Adler PB, Smull D, Beard KH, Choi RT, Furniss T, Kulmatiski A, Meiners JM, Tredennick AT, Veblen KE. Competition and coexistence in plant communities: intraspecific competition is stronger than interspecific competition. Ecol Lett. 2018;21(9):1319–29.
Condit R, Ashton PS, Baker P, Bunyavejchewin S, Gunatilleke S, Gunatilleke N, Hubbell SP, Foster Rb, Itoh A, LaFrankie JV, Lee HS, Losos E, Manokaran N, Sukumar R, Yamakura T. Spatial patterns in the distribution of tropical tree species. Science. 2000;288(5470):1414–8.
Getzin S, Dean C, He F, Trofymow JA, Wiegand K, Wiegand T. Spatial patterns and competition of tree species in a Douglas-fir chronosequence on Vancouver Island. Ecography. 2006;29(5):671–82.
Lin YC, Chang LW, Yang KC, Wang HH, Sun IF. Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation. Oecologia. 2011;165(1):175–84.
Li L, Huang Z, Ye W, Cao H, Wei S, Wang Z, Lian J, Sun IF, Ma K, He F. Spatial distributions of tree species in a subtropical forest of China. Oikos. 2009;118(4):495–502.
Kalyuzhny M, Lake JK, Wright SJ, Ostling AM. Pervasive within-species spatial repulsion among adult tropical trees. Science. 2023;381:563–8.
Li Y, He J, Yu S, Wang H, Ye S. Spatial structures of different-sized tree species in a secondary forest in the early succession stage. Eur J Forest Res. 2020;139(5):709–19.
Zhao Z, Hui G, Hu Y, Wang H, Zhang G, von Gadow K. Testing the significance of different tree spatial distribution patterns based on the uniform angle index. Can J Forest Res. 2014;44(11):1419–25.
Do TV, Sato T, Saito S, Kozan O, Yamagawa H, Nagamatsu D, Nishimura N, Manabe T. Effects of micro-topographies on stand structure and tree species diversity in an old-growth evergreen broad-leaved forest, southwestern Japan. Glob Ecol Conser. 2015;4:185–96.
Oksanen J, Blanchet F, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin P, O’Hara R, Simpson G, Solymos P, Stevens M, Szoecs E, Wagner H. vegan: Community Ecology Package. CRAN R package; 2009.
Kouba Y, Martínez-García F, de Frutos Á, Alados CL. Plant β-diversity in human-altered forest ecosystems: the importance of the structural, spatial, and topographical characteristics of stands in patterning plant species assemblages. Eur J Forest Res. 2014;133(6):1057–72.
Pinzon J, Wu L, He F, Spence JR. Fine-scale forest variability and biodiversity in the boreal mixedwood forest. Ecography. 2018;41(5):753–69.
Wulf M, Kolk J, Zobel M. Plant species richness of very small forests related to patch configuration, quality, heterogeneity and history. J Veg Sci. 2014;25(5):1267–77.
Yang M, Cai T, Ju C, Zou H. Evaluating spatial structure of a mixed broad-leaved/Korean pine forest based on neighborhood relationships in Mudanfeng National Nature Reserve. China J Forestry Res. 2019;30(4):1375–81.
Getzin S, Wiegand T, Wiegand K, He F. Heterogeneity influences spatial patterns and demographics in forest stands. Jf Ecol. 2008;96(4):807–20.
Wang H, Peng H, Hui G, Hu Y, Zhao Z. Large trees are surrounded by more heterospecific neighboring trees in Korean pine broad-leaved natural forests. Sci Rep. 2018;8(1):9149.
D’Amen M, Mod HK, Gotelli NJ, Guisan A. Disentangling biotic interactions, environmental filters, and dispersal limitation as drivers of species co-occurrence. Ecography. 2017;41(8):1233–44.
Hu Y, Hui G. A new method for measuring population distribution patterns of forest trees based on the mingling degree. J Beijing Forestry Univer. 2015;37(1):9–14.
Keren S, Diaci J, Motta R, Govedar Z. Stand structural complexity of mixed old-growth and adjacent selection forests in the Dinaric Mountains of Bosnia and Herzegovina. Forest Ecol Manag. 2017;400:531–41.
Pommerening A, Uria-Diez J. Do large forest trees tend towards high species mingling? Ecol Inform. 2017;42:139–47.
Pommerening A, Wang H, Zhao Z. Global woodland structure from local interactions: new nearest-neighbour functions for understanding the ontogenesis of global forest structure. Forest Ecosyst. 2020;7:22.
Omelko A, Ukhvatkina O, Zhmerenetsky A, Sibirina L, Petrenko T, Bobrovsky M. From young to adult trees: How spatial patterns of plants with different life strategies change during age development in an old-growth Korean pine-broadleaved forest. Forest Ecol Manag. 2018;411:46–66.
Hao Z, Zhang J, Song B, Ye J, Li B. Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest. Forest Ecol Manag. 2007;252(1–3):1–11.
LaManna JA, Walton ML, Turner BL, Myers JA, Rejmanek M. Negative density dependence is stronger in resource-rich environments and diversifies communities when stronger for common but not rare species. Ecol Lett. 2016;19(6):657–67.
Umaña MN, Zipkin EF, Zhang C, Cao M, Lin L, Swenson NG, Norden N. Individual-level trait variation and negative density dependence affect growth in tropical tree seedlings. J Ecol. 2018;106(6):2446–55.
Xu H, Hui G, Hu Y, Li C, Lin T, Zhang X, Wu X. Analysis of spatial distribution characteristics of trees with different diameter classes in natural Korean pine broad leaved forest. Forest Res. 2006;19(6):687–91.
Ni R, Baiketuerhan Y, Zhang C, Zhao X, Gadow Kv. Analysing structural diversity in two temperate forests in northeastern China. Forest Ecol Manag. 2014;316:139–47.
Franklin J, Keppel G, Webb EL, Seamon JO, Rey SJ, Steadman DW, Wiser SK, Drake DR, Gillman LN. Dispersal limitation, speciation, environmental filtering and niche differentiation influence forest tree communities in West Polynesia. J Biogeogr. 2013;40(5):988–99.
MacFarlane DW, Kobe RK. Selecting models for capturing tree-size effects on growth–resource relationships. Can J Forest Res. 2006;36(7):1695–704.
Chai Z, Sun C, Wang D, Liu W, Zhang C. Spatial structure and dynamics of predominant populations in a virgin old-growth oak forest in the Qinling Mountains. China Scandinavian J Forest Rese. 2016;32(1):19–29.
Hui G, Zhao X, Zhao Z, Gadow Kv. Evaluating tree species spatial diversity based on neighborhood relationships. Forest Sci. 2011;57(4):292–300.
Shankar U. Phytosociology of stratification in a lowland tropical rainforest occurring north of the Tropic of Cancer in Meghalaya. India Plant Divers. 2019;41(5):285–99.
Ostertag R, Inman-Narahari F, Cordell S, Giardina CP, Sack L. Forest structure in low-diversity tropical forests: a study of Hawaiian wet and dry forests. PLoS ONE. 2014;9(8): e103268.
Bohlman SA. Species diversity of canopy bersus understory trees in a neotropical forest: Implications for forest structure, function and monitoring. Ecosystems. 2015;18(4):658–70.
Acknowledgements
Employees from Guangxi Forest Resources and Environment Monitoring Center helped us to collect data.
Funding
This work was supported the National Natural Science Foundation of China (Grant No. 32060340) and Scientific Research Capacity Building Project for Laibin Jinxiu Dayaoshan Forest Ecosystem Observation and Research Station of Guangxi (Grant No. 22–035-130–01).
Author information
Authors and Affiliations
Contributions
Yuanfa Li conceived the idea and collected dataset from wild; Yuanfa Li and Liting Wei wrote and reviewed the manuscript; Liting Wei analyzed the dataset and prepared all figures.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Since dataset comes from fixed plots, no specimen is collected and no permission is necessary to design the study. All figures are our own and have not been published elsewhere.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The English in this document has been checked by at least two professional editors, both native speakers of English. For a certificate, please see: https://www.textcheck.com/certificate/gYQqhU.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, Y., Wei, L. Species and structural diversity of trees at the structural type level. BMC Ecol Evo 24, 40 (2024). https://doi.org/10.1186/s12862-024-02229-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12862-024-02229-y