
Abstract
Background
Understanding genomic and phenotypic diversity among cryptic pest taxa has important implications for the management of pests and diseases. The diamondback moth, Plutella xylostella L., has been intensively studied due to its ability to evolve insecticide resistance and status as the world’s most destructive pest of brassicaceous crops. The surprise discovery of a cryptic species endemic to Australia, Plutella australiana Landry & Hebert, raised questions regarding the distribution, ecological traits and pest status of the two species, the capacity for gene flow and whether specific management was required. Here, we collected Plutella from wild and cultivated brassicaceous plants from 75 locations throughout Australia and screened 1447 individuals to identify mtDNA lineages and Wolbachia infections. We genotyped genome-wide SNP markers using RADseq in coexisting populations of each species. In addition, we assessed reproductive compatibility in crossing experiments and insecticide susceptibility phenotypes using bioassays.
Results
The two Plutella species coexisted on wild brassicas and canola crops, but only 10% of Plutella individuals were P. australiana. This species was not found on commercial Brassica vegetable crops, which are routinely sprayed with insecticides. Bioassays found that P. australiana was 19-306 fold more susceptible to four commonly-used insecticides than P. xylostella. Laboratory crosses revealed that reproductive isolation was incomplete but directionally asymmetric between the species. However, genome-wide nuclear SNPs revealed striking differences in genetic diversity and strong population structure between coexisting wild populations of each species. Nuclear diversity was 1.5-fold higher in P. australiana, yet both species showed limited variation in mtDNA. Infection with a single Wolbachia subgroup B strain was fixed in P. australiana, suggesting that a selective sweep contributed to low mtDNA diversity, while a subgroup A strain infected just 1.5% of P. xylostella.
Conclusions
Despite sympatric distributions and the capacity to hybridize, strong genomic and phenotypic divergence exists between these Plutella species that is consistent with contrasting colonization histories and reproductive isolation after secondary contact. Although P. australiana is a potential pest of brassicaceous crops, it is of secondary importance to P. xylostella.
Similar content being viewed by others


Background
Cryptic species can show remarkable diversity in aspects of their ecology, behaviour, and at the level of the genome. They exist across metazoan taxa [1], including globally important arthropod pest taxa, such as whiteflies [2], disease-vectoring mosquitoes [3], fruit flies [4], thrips [5, 6] and mites [7, 8], some of which are characterised by cryptic species complexes. Discovering cryptic diversity has important consequences for estimates of global biodiversity, conservation planning, and the management of pests and diseases. Morphologically similar species can vary in pest status due to differences in genotypic and/or phenotypic traits that influence their host range and specificity, geographic distribution, the ability to vector diseases, or insecticide resistance [8–10]. Therefore, recognising cryptic species and the differences in their biology and ecology are essential for effective management, with important implications for public health, agriculture and trade.
The diamondback moth, Plutella xylostella, is the major pest of brassicaceous crops worldwide, costing an estimated US$4 to US$5 billion annually in direct losses and management costs [11, 12]. Insecticide resistance is widespread in P. xylostella populations around the world, fuelling wide-ranging research to develop alternative management tactics [11, 13]. Plutella xylostella was initially recorded in Australia in the late 1800s and rapidly became a widespread pest of Brassica vegetables, and then canola following its expanded production from the 1990s [14, 15]. Recently, Landry and Hebert [16], through mtDNA barcoding, identified a cryptic lineage of Plutella in Australia not detected in previous molecular studies of P. xylostella [14, 17–21]. Although external morphology was indistinguishable from P. xylostella, deep mtDNA divergence (8.6%), differences in genital morphology and endemism in Australia led them to describe a new species, Plutella australiana Landry & Hebert. Plutella australiana was originally collected together with P. xylostella in light trap samples in eastern Australia, suggesting at least some ecological overlap [16], but its biology, ecology and pest status were unknown.
The management of P. xylostella in Australian Brassica crops has been a significant challenge for decades [15, 22], but the discovery of P. australiana has made the relative abundance and pest status of both species in these crops uncertain. With rare exception, P. xylostella and allied species feed on plants in the order Brassicales, mainly within the family Brassicaceae [16, 23, 24], implying that the host range of P. australiana may include cultivated brassicas. Widespread resistance to pyrethroid and organophosphate insecticides has been attributed to Australian populations of P. xylostella from all vegetable and canola production regions, which has led to ineffective control during outbreaks [22, 25]. Plutella xylostella is well known as a migratory insect with a high capacity for gene flow [11, 13], facilitating the rapid spread of resistance alleles. Australian P. xylostella are thought to disperse frequently, based on indirect evidence from ecological and genetic studies [14, 15, 26]. Most studies have found a lack of genetic differentiation at microsatellite loci and low sequence variation in mitochondrial DNA markers among Australian and New Zealand populations of P. xylostella, consistent with high gene flow and/or recent ancestry [14, 15, 17, 18]. While species identification was not in question in these studies, somewhat inconsistent findings in two studies from eastern Australia using allozymes or SSR markers [19, 20] might reflect the confounding presence of P. australiana samples [16]. Given these considerations, future management of Plutella in Australian crops will require thorough understanding of the ecological requirements, genetic traits and pest status of the two Plutella species. In addition, reproductive isolation between these two species is unknown but has implications for evolutionary inference and the potential for gene flow. The capacity for hybridization and introgression could lead to the exchange of insecticide resistance or other adaptive alleles [27, 28].
Although mtDNA markers are widely used in studies of species identity and population structure [29–31], mitochondrial variation within or between species can be influenced by direct and/or indirect selection, or introgressive hybridization [32, 33]. One factor that can confound mtDNA-based inference is interaction with inherited bacterial symbionts [34, 35]. Wolbachia is a widespread endosymbiont thought to infect at least half of arthropod [36] and 80% of lepidopteran [37] species. It is mainly transmitted vertically from infected females to their offspring through the egg cytoplasm, and inheritance is therefore linked with mtDNA. To facilitate its spread, Wolbachia manipulates host reproductive biology to favour the fitness of infected females by inducing host phenotypes that distort sex ratios (male-feminization, male-killing or induction of parthenogenesis) or cause sperm-egg cytoplasmic incompatibility (CI) [38, 39]. In the simple case involving a single CI-inducing strain, crosses with infected females are fertile but crosses between uninfected females and infected males fail to produce offspring. If maternal transmission is efficient and infected females have a reproductive advantage, Wolbachia infection can spread rapidly through an insect population [40], driving a selective sweep of a single haplotype and reducing mtDNA diversity [41]. Limited surveys to date have identified Wolbachia strains infecting P. xylostella at low frequency in populations from North America, Africa, Asia and Europe [18, 42, 43]. Because symbionts can contribute to reproductive isolation and influence mtDNA diversity [34, 44], assessing their role can provide important insights into host evolution and population structure [35, 45–47].
Here we investigated the biology, ecology and population genetic structure of two cryptic Plutella species by collecting Plutella from brassicaceous plants throughout Australia and screening individuals to identify mtDNA lineages and Wolbachia infections. For a subset of populations, we examined genetic diversity using thousands of nuclear SNPs from across the genome. In addition, we assessed reproductive compatibility in laboratory crosses and determined the susceptibility of each species to commercial insecticides.
Methods
Sample collection
Plutella larvae (rarely, eggs or pupae) were collected from canola crops, Brassica vegetable crops, forage brassicas and wild brassicas throughout Australia between March 2014 and December 2015 (Table 1). The wild species included wild radish, Raphanus raphanistrum, turnip weed, Rapistrum rugosum, sea rocket, Cakile maritima, Ward’s weed, Carrichtera annua, African mustard, Brassica tournefortii, and mixed stands of sand rocket, Diplotaxis tenuifolia, and wall rocket, D. muralis. At each location, at least 25 individuals were collected from randomly selected plants to achieve a representative sample. Insect samples were collected from Brassica vegetables by hand, from sea rocket by beating plants over a collection tray and from other hosts using a sweep net. Each population sample was separately reared in ventilated plastic containers on leaves of the original host material for 1–2 days and thereafter on cabbage leaves. Non-parasitised pupae or late-instar larvae were fresh frozen at −80 °C.
DNA isolation and COI genotyping
For each population sample, we aimed to genotype a minimum of 16 individuals where possible after removing parasitized individuals. Individual pupae (but not larvae) were sexed under a dissecting microscope, then genomic DNA was isolated by homogenising whole individuals followed by two phenol and one chloroform extractions according to Zraket et al. [48]. DNA was treated with RNase A, then precipitated and re-suspended in TE buffer. Plutella lineages were distinguished using a PCR-RFLP assay [49]. A 707 bp COI region was amplified using a combination of two primer pairs: (i) PxCOIF (5′-TCAACAAATCATAAAGATATTGG- 3′) and PxCOIR (5′-TAAACTTCAGGGTGACCAAAAAATCA- 3′), and (ii) PaCOIF (5′-TCAACAAATCATAAGGATATTGG- 3′) and PaCOIR (5 ′-TAAACCTCTGGATGGCCAAAAAATCA- 3′). Ten microliter reactions were run with 2 µL of MyTaq 5x buffer, 0.2 µL of each primer (10mM stocks), 1 µL of DNA (approx. 5 ng) and 0.05 µL of MyTaq polymerase (Bioline). Samples were amplified at 95 °C for 2 min, then 35 cycles at 95 °C for 10 s, 52 °C for 20 s, 72 °C for 30 s followed by a 5 min final extension at 72 °C. PCR products were digested at 37 °C for 1 h with 1 unit of AccI (NEB) restriction enzyme with 2 µL Cutsmart Buffer in a 20 µL reaction. Following digestion, products were separated using agarose gel electrophoresis (1.5%). Plutella xylostella products are approximately 516 bp and 191 bp and P. australiana products are 348 bp and 359 bp [49]. To examine mtDNA haplotypes, sequencing of the 707 bp COI amplicon was performed for 44 P. xylostella and 37 P. australiana individuals at the Australian Genome Research Facility (AGRF). In addition, we downloaded sequence trace files from Landry and Hebert [16] (dx.doi.org/10.5883/DS-PLUT1) and re-analysed, aligned and trimmed all sequences in GENEIOUS version 10.0.6 [50]. Haplotype networks were constructed using R package pegas version 0.9 [51].
Wolbachia screening and phylogenetics
Wolbachia infection was detected using two separate PCR assays of the 16S rRNA gene (16S-2 and 16S-6) according to Simoes et al. [52]. To identify Wolbachia strains, the Wolbachia surface protein (wsp) gene was sequenced in a subset of individuals. Amplification was performed using wsp81F and wsp691R sequence primers [53]. Amplicons were sequenced using the reverse primer and aligned in GENEIOUS version 10.0.6 [50]. We used a 493 bp alignment to construct a maximum likelihood phylogeny in RAxML version 8.2.4 [54] using a general time reversal substitution model [55] with 1000 bootstrap replicates.
RADseq library preparation and sequencing
Libraries were prepared for restriction-site-associated DNA sequencing (RADseq) according to a protocol modified from Baird et al. [56]. Genomic DNA was quantified using a Qubit 2.0 fluorometer (Invitrogen) and 200 ng digested with 10 units of high fidelity SbfI in Cutsmart Buffer (NEB) for 1 h at 37 °C, then heat inactivated at 80 °C for 20 min. One microlitre of P1 adapter (100nM) with a 6-base molecular identifier (MID) (top strand 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGxxxxxxTGCA- 3′, bottom strand 5′-[P]xxxxxxCTGTCTCTTATACACATCTGACGCTGCCGACGA- 3′, x represents sites for MIDs) were then added using 0.5 µL T4 DNA ligase (Promega), 1 nM ATP and Cutsmart buffer. Library pools were sheared using a Bioruptor sonicator (Diagenode), then DNA fragments end-repaired using a Quick Blunting Kit (NEB), adenine overhangs added then P2 adapters (top strand 5′-[P]CTGTCTCTTATACACATCTCCAGAATAG- 3′, bottom strand 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGT- 3′) ligated. DNA purification between steps was performed using a MinElute PCR purification kit (Qiagen). Libraries were amplified using KAPA HiFi Hotstart Readymix (Kapa Biosystems) and Nextera i7 and i5 indexed primers with PCR conditions: 95 °C for 3 min, two cycles of 98 °C for 20 s, 54 °C for 15 s, 72 °C for 1 min, then 15 cycles of 98 °C for 20 s, 65 °C for 15 s, 72 °C for 1 min followed by a final extension of 72 °C for 5 min. Libraries were size-selected (300-700 bp) on 1–1.5% agarose gel and purified using a minElute Gel Extraction Kit (Qiagen), then Illumina paired-end sequencing was performed using HiSeq2500 (100 bp) or NextSeq500 (75 bp) at the AGRF.
Read filtering and variant calling
Sequence reads were demultiplexed using RADtools version 1.2.4 [57] allowing one base MID mismatch, then TRIMMOMATIC version 0.32 [58] was used to remove restriction sites, adapter sequences and a thymine base from reverse reads introduced by the P2 adapter, and quality filter using the ILLUMINACLIP tool with parameters: TRAILING:10 SLIDINGWINDOW:4:15 MINLEN:40. Paired reads were aligned to the P. xylostella reference genome (accession number: GCF_000330985.1) using STAMPY version 1.0.21 [59] with --baq and --gatkcigarworkaround options and expected substitution rate set to 0.03 for P. xylostella and 0.05 for P. australiana to reflect expected levels of sequence divergence relative to the P. xylostella reference genome. Duplicate reads were removed using PICARD version 1.71 [60]. Genotypes were called using the Genome Analysis Toolkit (GATK) version 3.3-0 [61, 62] HaplotypeCaller tool. We determined that base quality score recalibration using bootstrapped SNP databases was inappropriate for this dataset as it globally reduced quality scores. For downstream comparisons between species, we joint-genotyped P. australiana and P. xylostella individuals using the GATK GenotypeGVCFs workflow. To examine finer scale population structure, we also joint-genotyped the P. australiana individuals alone. All variant callsets were hard-filtered with identical parameters using VCFtools version 0.1.12a [63]: We removed indels and retained confidently-called biallelic SNPs (GQ\(\geqslant \)30) genotyped in at least 70% of individuals with a minimum genotype depth of 5, minQ\(\geqslant \)400, average site depth of 12–100, minimum minor allele frequency of 0.05, in Hardy-Weinberg equilibrium at an alpha level of 0.05. To avoid linked sites, we used the VCFtools --thin function to retain only SNPs separated by a minimum of 2000 bp. To estimate genetic diversity, we generated a set of all confidently-called variant and invariant sites (GQ\(\geqslant \)30), and hard filtered to remove sites within repetitive regions and retain sites genotyped in at least 70% of individuals with an average site depth of 12–100. Sites from the mitochondrial genome were excluded from all datasets.
Genetic diversity and population structure
Genetic diversity was calculated for Plutella populations of both species from five locations. The R package hierfstat [64] was used to calculate observed heterozygosity, gene diversity and the inbreeding coefficient, FIS, according to Nei [65]. Population means for site depth and number of SNPs, indels and private sites were calculated using the --depth function and vcfstats module in VCFtools version 0.1.12a [63]. The number of heterozygous sites within individuals was determined from all confidently-called sites excluding indels using a custom python script parseVCF.py [66] and visualised using R [67].
To examine population structure in P. australiana, a global estimate of FST [68] with bootstrapped 99% confidence intervals (104 bootstrap replicates) was calculated in R package diveRsity [69]. Pairwise FST values for all population pairs were calculated and significance determined using exact G tests (104 mc burnins, 103 batches, and 104 iterations per batch) in GENEPOP version 4.6 [70] after Bonferroni correction for multiple comparisons. Separate analysis of population structure was performed using the Bayesian clustering program STRUCTURE version 2.3.4 [71], first for all individuals of co-occurring Plutella species, and second for P. australiana alone. For all runs, we used a burnin length of 5×105 followed by a run length of 106 MCMC iterations and performed ten independent runs for each K value from 1 to 10, where K is the number of genotypic clusters, using a different random seed for each run, assuming the locprior model with correlated allele frequencies and λ set to 1. The optimal value of K was determined using the delta K method [72] implemented in STRUCTURE HARVESTER [73] and inspection of the likelihood distribution for each model. Q-matrices were aligned across runs using CLUMPP version 1.1.2 [74] and visualised using DISTRUCT version 1.1 [75].
Laboratory cultures of Plutella species
Laboratory cultures of P. australiana and P. xylostella were established from field populations and used for crossing experiments and insecticide bioassays. Plutella adults were collected at light traps at Angle Vale and Urrbrae, South Australia, in October–November 2015. Females were isolated and allowed to lay eggs, then identified using PCR-RFLP and progeny pooled to produce separate cultures of each species. A laboratory culture of the Waite Susceptible P. xylostella strain (S) has been maintained on cabbage without insecticide exposure for approximately 24 years (≈310 generations) and was used as a bioassay reference strain. All cultures were maintained in laboratory cages at 26 ± 2.0°C and a 14:10 (L:D) hour photoperiod at the South Australian Research and Development Institute, Waite Campus, Adelaide, South Australia. The P. australiana culture was maintained on sand rocket, Diplotaxis tenuifolia, and the P. xylostella culture was maintained on cabbage, Brassica oleracea var. capitata. The purity of cultures was assessed regularly using PCR-RFLP.
Crossing experiments
Plutella australiana and P. xylostella pupae were sexed under a stereo microscope, then placed into individual 5 mL clear polystyrene tubes with fine mesh lids and gender visually confirmed after eclosion. Enclosures used for crossing experiments were 850 mL polypropylene pots (Bonson Pty Ltd) modified with lateral holes covered with voile mesh for ventilation. Crosses of single mating pairs were performed on laboratory benches at 26 ± 2.0 °C and 14:10 (L:D) photoperiod using 3-week old D. tenuifolia seedlings as the host plant. After seven days, adults were collected into a 1.5 mL tube and fresh frozen at −80 °C for species confirmation using PCR-RFLP. Seedlings were examined and eggs counted under a stereo microscope, then returned to enclosures to allow egg hatch. Larvae were provided with fresh 3–4 week old seedlings until pupation, then pupae were individually collected into 5 mL tubes. Hybrid F1xF1 crosses and back-crosses were then performed as above. The presence of egg and adult offspring was recorded for all replicates, and for the majority of replicates (> 80%), the numbers of offspring were counted and used to calculate a mean.
Insecticide bioassays
Insecticide susceptibility of field-collected Plutella strains was compared to the susceptible P. xylostella (S) reference in dose-response assays using four commercial insecticides: Dominex (100 g L−1 alpha-cypermethrin), Proclaim (44 g kg−1 emamectin benzoate), Coragen (200 g L−1 chlorantraniliprole) and Success Neo (120 g L−1 spinetoram). Bioassays were performed by placing 3rd instar larvae onto inverted leaf discs embedded in 1% agar in 90 mm Petri dishes. Cabbage leaves, Brassica oleracea. var. capitata were used for P. xylostella and canola leaves, B. napus var. ‘ATR Stingray’, were used for P. australiana. Eight concentrations and a water-only control were evaluated for each insecticide using four replicates of ten larvae. A 4 mL aliquot of test solution was applied directly to leaves using a Potter Spray Tower (Burkard Manufacturing Co. Ltd.) calibrated to deliver an aliquot of 3.52 ± 0.09 mg cm-1. After application, each dish was placed in a controlled temperature room at 25 ± 0.5 °C, then mortality was assessed after 48 h (Dominex, Success Neo) or 72 h (Proclaim, Coragen). Dose-response analysis was performed using log-logistic regression in R package drc [76] and the fitted models were used to estimate the lethal concentration predicted to cause 50% (LC50) and 99% (LC99) mortality of the test population. Resistance ratios were calculated by dividing the LC50 and LC99 estimates for field strains by the corresponding LC estimates for the P. xylostella (S) reference strain.
Results
Geographic distribution and host associations
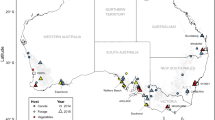
Plutella larvae were collected from brassicaceous plants at 75 locations in Australia and 1477 individuals were genotyped at the COI locus using PCR-RFLP to identify species. Of these, 88% (n=1300) were genotyped as P. xylostella, 10% (n=147) as P. australiana and 2% (n=30) were unresolved (Table 1). Plutella australiana was identified in around one quarter (n=20/75) of collections distributed across southern Australia, while P. xylostella occurred at all locations except Cunnamulla, Queensland, in a collection from wild African mustard, Brassica tournefortii (Table 1). The sex ratio was not different from 1:1 for P. xylostella (481 females, 517 males, χ2=1.2986, p=0.2545) or P. australiana (63 females, 55 males, χ2=0.5424, p=0.4615). The relative incidence and abundance of P. australiana was >2-fold higher in the eastern state of New South Wales than in other states (Fig. 1). Plutella australiana larvae were detected in 29% (n=5/17) of collections from wild brassicas and from species including wild radish, Raphanus raphanistrum, wild turnip, Rapistrum rugosum, African mustard, B. tournefortii, and mixed stands of sand rocket, D. tenuifolia and wall rocket, D. muralis (Table 2). Among cultivated crops, P. australiana larvae occurred in 36% (n=14/39) of samples from canola, consisting of 11% of total Plutella individuals from those crops, but were not identified from commercial Brassica vegetable farms (Table 2). However, P. australiana eggs were collected from kale at one farm.

The geographic distribution of P. xylostella (light grey) and P. australiana individuals (black) in larval collections from brassicaceous plants in Australia during 2014 and 2015. Pie diagrams show the relative proportion of each species at each location. Overlapped pies represent locations with 100% P. xylostella. Green highlighted circles indicate five locations from which individuals of each species were RAD sequenced
Wolbachia infections
Plutella individuals (n=1447) were screened for Wolbachia infection using 16S rRNA PCR assays. Only 1.5% (n=19/1300) of P. xylostella collected from eight different locations were infected (Table 1). In contrast, all 147 P. australiana individuals were infected with Wolbachia across the 20 locations where this species occurred. To identify Wolbachia strains, a Wolbachia surface protein (wsp) amplicon was sequenced from 14 P. xylostella and 30 P. australiana individuals. Each species was infected with a different strain. The wsp sequence for Australian P. xylostella showed 100% identity to a Wolbachia supergroup A isolate infecting P. xylostella from Malaysia, plutWA1 [18]. For P. australiana, the wsp sequence showed 100% identity to a Wolbachia supergroup B isolate infecting a mosquito, Culex pipiens, from Turkey and the winter moth, Operophtera brumata, from the Netherlands (Fig. 2).

Maximum likelihood phylogeny of 493 bp of Wolbachiawsp amplicons for Plutella and other arthropods. The strain infecting P. australiana (wAus) was identical to a Wolbachia supergroup B strain reported from Culex pipiens and Operophtera brumata. The strain infecting Australian P. xylostella was identical to a supergroup A strain (plutWA1) reported from Malaysian P. xylostella. Labels include the Wolbachia strain, host species and GenBank accession number. Labels in bold denote strains sequenced in this study. The scale bar shows the mean number of nucleotide substitutions per site
Crossing experiments
Inter-species single pair mating experiments showed that hybridization between P. australiana and P. xylostella was possible, yet less successful than intra-species crosses. While most intra-species crosses produced adult offspring, the fecundity of P. xylostella was >2-fold higher than P. australiana (Table 3). Both reciprocal inter-species crosses produced F1 adult offspring, but success was asymmetric and notably higher in the pairs with P. australiana females. In this direction, there was a strong male bias in the F1 progeny: from 76 cross replicates, 16 collectively produced 9 female and 80 male adults, a ratio of 8.9. Hybrid F1xF1 crosses for both parental lines produced F2 adult offspring (Table 4). For the P. australiana maternal line, parental back-crosses using F1 hybrid males successfully produced offspring, while parental back-crosses with F1 hybrid females were sterile. For the P. xylostella maternal line, low fitness allowed only a single parental back-cross replicate, which involved a hybrid female and was sterile.
Mitochondrial haplotype diversity
Mitochondrial haplotype networks of Australian Plutella were constructed using a 613 bp COI alignment that included 81 sequences from this study and 108 from Landry and Hebert [16]. We found low haplotype diversity within Australian P. xylostella, consistent with previous reports [17, 18, 77]. Only five haplotypes were identified among 102 individuals, including three identified by Saw et al. [17] and three occurring in single individuals (Fig. 3a). The most common haplotype, PxCOI01, occurred at high frequency and differed by a single base mutation from other haplotypes (Fig. 3a, Additional file 1: Table S1). Nine closely related haplotypes were identified in 87 P. australiana individuals with seven occurring in single individuals (Fig. 3b). The most common haplotype, PaCOI01, occurred at high frequency and differed by 1-2 base mutations from other haplotypes (Fig. 3b, Additional file 1: Table S2).

Mitochondrial DNA haplotype network for aP. xylostella (n=102,44 from this study, 58 from [16]) and bP. australiana (n=87, 37 from this study, 50 from [16]) individuals from Australia based on a 613 bp COI sequence alignment. Haplotypes shared by more than one individual are shown in circles with a grey border with the number of individuals indicated inside the circle. Haplotypes connected by a line differ by a single mutation
Nuclear diversity and population structure
At five collection locations, P. australiana co-occurred with P. xylostella in sufficient numbers to enable comparison of nuclear genomes, though the relative abundance of species varied between locations. To ensure representation from the south-west region of Australia, the Esperance population (n=5) was formed by including one P. australiana individual from Boyup Brook. Despite only two P. xylostella individuals at Gilgandra, this population had 17 P. australiana individuals and was included. To generate nuclear SNP markers, we performed RADseq for a total of 52 P. australiana and 47 P. xylostella individuals.
Illumina sequencing and demultiplexing using RADtools [57] yielded 276.8 million raw sequence reads. Following read quality filtering and mapping, genotypes were called for 99 individuals from the two species. Hard filtering retained 300,241 confidently-called variant and invariant nuclear sites at a mean depth >36 per individual, and a subset of 689 widely-dispersed nuclear SNP variants (to avoid linkage bias) at a mean depth >36 per individual, for comparative analyses of genetic diversity and population structure. The dataset with all confidently-called sites was used to estimate population-level genetic diversity.
Estimates of nuclear genetic diversity across 300,241 variant and invariant sites revealed a striking contrast between Plutella species, with notably higher diversity within populations of P. australiana than co-occurring populations of P. xylostella (Table 5). The mean observed heterozygosity within populations ranged from 0.013–0.016 for P. australiana and 0.009–0.010 for P. xylostella. Similarly, the average numbers of SNPs, indels and private alleles were considerably higher within P. australiana populations. As P. australiana may have fixed nucleotide differences relative to the P. xylostella reference genome that may affect population level statistics, we also removed indels from this dataset and directly compared the heterozygosity among individuals using 289,347 sites. Plutella australiana individuals had on average a 1.5-fold higher proportion of heterozygous sites than P. xylostella individuals (Fig. 4).

Boxplot showing the proportion of heterozygous sites across 289,347 confidently-called nuclear sites for individuals of P. xylostella (light grey boxes, n=47) and P. australiana (dark grey boxes, n=52) from five locations. Heterozygosity was consistently higher in P. australiana
Genetic structure among co-occurring populations of Plutella species was investigated using 689 widely-dispersed nuclear SNPs in the program STRUCTURE. The delta K method predicted a strong optimal at K=2 genotypic clusters. Plutella australiana and P. xylostella individuals were clearly separated into distinct genotypic clusters in accordance to their species identified through mtDNA genotypes regardless of geographic location (Fig. 5, left panel). Five individuals across four locations showed greater than 1% admixture as shown by sharing of colored bars.

Proportional assignment of Plutella individuals to genotypic clusters, K, based on STRUCTURE analysis. Individuals are represented by vertical bars and genotypic clusters are represented by different colors. Left panel: Analysis at K=2 for 52 P. australiana and 47 P. xylostella individuals sorted left-to right by proportion of cluster membership. The predominantly red bars correspond to P. australiana individuals and the predominantly blue bars correspond to P. xylostella individuals identified through mtDNA genotypes. Locations are labelled for five individuals showing >1% genotypic admixture. Right panel: Analysis at K=3 for 52 P. australiana individuals sorted left-to-right by proportion of cluster membership within geographic locations, showing a high degree of genotypic admixture among individuals across locations
Assessing population structure from datasets with multiple species can mask heirachical structure [78]. To address this, genotypes were separately called for 52 P. australiana individuals, and hard filtering retained a set of 974 widely-dispersed SNP variants at a mean depth >33 per individual for examination of finer scale structure among five populations. The delta K method predicted a weak modal signal at K=3, but the highest likelihood value occurred at K=1. Bar plots for K=3 showed a high degree of admixture among individuals across the five populations, consistent with high levels of gene flow across Australia (Fig. 5, right panel). Pairwise FST was then calculated for the five P. australiana populations using 974 SNPs. The global estimate of FST was not significantly different from zero, indicating the populations are not differentiated (FST=0.0002, 99% CI = -0.0274–0.0387). Further, pairwise FST values were low, ranging from –0.0041 to 0.0038, suggesting substantial gene flow among populations separated by distances of between 341 and 2700 kilometres (Table 6).
Insecticide susceptibility
Bioassays revealed highly contrasting responses to insecticide exposure in P. xylostella and P. australiana field strains (Fig. 6). Plutella australiana showed extremely high susceptibility to all four insecticides evaluated: resistance ratios at the LC50 and LC99 estimates were less than 1.0 and showed that this strain was 1.5-fold to 7.4-fold more susceptible than the laboratory P. xylostella (S) reference (Additional file 1: Table S3). In contrast, resistance ratios at the LC50 for the field P. xylostella strain ranged from 2.9 for Success Neo to 41.4 for Dominex, indicating increased tolerance to all insecticides. Comparison of the LC99 estimates with commercial field doses for each insecticide implies differences in field efficacy between species. The commercial field rate of Dominex was >8-fold lower than the LC99 for P. xylostella, suggesting likely poor field control of this strain, but was >17-fold higher than the LC99 for P. australiana (Fig. 6). Control mortality was similar for the field and reference strains, averaging 3.1 to 4.4% across all bioassays.

Insecticide bioassay dose-response curves for P. australiana (dotted line) and P. xylostella (dashed line) field strains collected from Angle Vale and Urrbrae, South Australia, and a susceptible P. xylostella (S) reference strain (solid line), exposed to four commercial insecticides: Dominex, Coragen, Proclaim and Success Neo. Points are the mean observed response across 4 bioassay replicates of 10 larvae each and lines are the fitted log-logistic response curves with 95% confidence intervals shown in grey shading. The vertical red line represents the approximate commercial field dose for each insecticide and vertical black lines represent the estimated LC99 for the corresponding Plutella strain
Discussion
Cryptic species arise when divergence does not lead to morphological change [79]. The recent discovery of a cryptic ally, P. australiana, to the diamondback moth, P. xylostella, was unexpected given the breadth of previous molecular studies of this insect. Several factors may have contributed to this discovery, including the use of light traps for specimen collection, rather than limiting sampling to Brassica vegetable farms. Landry and Hebert [16] also isolated DNA from legs, keeping most of each specimen intact and providing a morphological reference for examining unexpected genotypes. It is also possible that P. australiana was previously overlooked from nuclear DNA studies due to biases in amplification of divergent alleles. Here, we sought to determine whether P. australiana is an agricultural pest, and to understand its ecological and genetic differences from P. xylostella.
Extensive larval sampling from wild and cultivated brassicaceous plants revealed that P. australiana co-occurs widely with P. xylostella throughout southern Australia and utilizes some of the same host plants. The relative abundance of P. australiana was on average 9-fold lower than P. xylostella. We observed higher proportions of P. australiana in larval collections from the eastern state of New South Wales, similar to the light trap samples from Landry and Hebert [16], possibly reflecting habitat suitability. Although we did not detect P. australiana in limited sampling from the island state of Tasmania, the presence of brassicas in the region and evidence from light traps that wind currents can transport Plutella moths across Bass Strait (Lionel Hill, Pers. Comm.) suggest it is likely to occur there.
Our study confirms that the host range of P. australiana includes canola crops and wild brassicaceous species. In laboratory rearing, P. australiana completed development on sand rocket, D. tenuifolia, and canola, B. napus, and was also collected from several other wild species, though without rearing to confirm host status. Our sampling focused on relatively few introduced brassicaceous species common in agricultural areas, yet the Australian Brassicales is represented by 11 plant families [80], including several non-Brassicaceae on which P. xylostella and its allies have been documented feeding, such as Capparaceae [24], Cleomaceae [16] and Tropaeolaceae [23]. The Australian Brassicaceae has records for 61 genera and 205 species [80], including many introduced species but also a diversity of native genera, such as Lepidium, Blennodium, and Arabidella, that occur over vast areas of Australia. Wider sampling of native Brassicales may identify other suitable hosts for P. australiana.
Plutella australiana larvae were not identified among samples from sixteen commercial Brassica vegetable crops despite the high suitability of these crops for P. xylostella [81], however eggs were collected from kale. It is possible that extreme insecticide susceptibility prevents juvenile P. australiana populations from establishing, as commercial Brassica vegetable crops are typically sprayed multiple times per crop cycle [22]. Our data show that P. australiana is far more susceptible than P. xylostella to four commonly used insecticides. At commercial application rates, these insecticides are likely to provide high-level control of P. australiana in Australian Brassica crops, but some products may provide marginal or poor control against P. xylostella due to insecticide resistance (Fig. 6) [22, 25]. Alternatively, some vegetable cultivars may not be attractive for oviposition or suitable for larval survival in P. australiana. We noted that P. australiana cultures provided with cabbage seedlings failed to produce viable eggs over seven days, but after replacing cabbage with Diplotaxis seedlings, egg-laying then occurred within 24 h. Exposure to host plants stimulates reproductive behaviour in P. xylostella [82], but olfactory cues for host recognition or oviposition [83–85] may differ between these Plutella species. Host preference and performance studies are required to test these hypotheses.
Insecticide bioassays have been conducted routinely on Australian P. xylostella to monitor levels of insecticide resistance in field populations [22, 25]. This method appears unlikely to be affected by the presence of P. australiana under typical conditions, as a period of laboratory rearing is usually necessary to multiply individuals prior to screening. In our experience, laboratory rearing of the two Plutella species on cabbage plants selects against P. australiana individuals when competing with P. xylostella in cages, causing the complete loss of P. australiana within a few generations. The reasons for this are unknown but may include differences in host preference or development rate, or direct competition.
Crossing experiments revealed that hybridization can occur between P. australiana and P. xylostella under controlled conditions and is most likely to occur in crosses involving Wolbachia-infected P. australiana females. Hybridization occurs in around 10% of animal species, particularly in captivity [86], but asymmetric reproductive isolation is commonly observed in reciprocal crosses between taxa [87]. In our experiments, a strong male bias in the offspring of interspecific crosses and failure to back-cross hybrid females both follow Haldane’s rule [88], which predicts greater hybrid inviability or sterility in the heterogametic sex (female, in Lepidoptera). This pattern can arise from epistatic interactions between sex-linked and/or autosomal genes that result in genetic incompatibilities [89, 90]. Although the back-crosses with F1 hybrid females were sterile, the back-crosses with hybrid males (to both species) were viable, which could enable the transfer of genes between hybrid and/or parental species. However, it is unclear whether hybridization occurs in the wild.
Although P. australiana and P. xylostella show deep divergence (8.6%) in mtDNA [16], the sole use of mtDNA can be unreliable for inference of evolutionary history and should be corroborated using evidence from nuclear markers [34]. Our analysis revealed striking differences in nuclear diversity across the genome between co-existing populations of each Plutella species collected at the same locations and times, and from the same host plant species. Plutella xylostella populations from Australia and New Zealand have low levels of genetic diversity compared with populations from other continents, thought to reflect the recent introduction of this species from a small founding population [14, 17, 77]. Consistent with this view, we found a remarkable 1.5-fold reduction in heterozygosity across >300,000 sites in P. xylostella compared with sympatric P. australiana populations. However, both species showed limited mtDNA diversity with a single predominant haplotype. While outgroups from other continents were not available, comparative analysis of these closely-related Australian Plutella species suggested that patterns of mitochondrial and nuclear diversity are concordant in P. xylostella and consistent with a demographic bottleneck [17, 18], but discordant in P. australiana.
Sequence variation in mitochondrial DNA can be strongly influenced by Wolbachia infection [41]. Extensive Wolbachia screening showed that each Plutella species was infected with a different strain at contrasting frequencies, and fit a ‘most-or-few’ pattern whereby species infection rates are often very low (<10%) or very high (>90%) [91]. Infection incidence in P. xylostella was lower in Australia (1%) than previously reported across global samples (5%) [18]. Our finding of a single supergroup A strain showing 100% sequence similarity to a strain reported in P. xylostella from Malaysia, plutWA1 [18], provides some support of an Asian origin for Australian P. xylostella [17], though does not preclude this strain also occurring elsewhere.
Fixation of infection in P. australiana suggests that Wolbachia manipulates the reproductive biology of this species. We found no evidence of sex-ratio distortion, which has been associated with a Wolbachia strain, plutWB1, in P. xylostella [18]. High infection can be driven by cytoplasmic incompatibility (CI) [40]. The high frequency (87%) of a single mtDNA haplotype among P. australiana individuals implies that the spread of Wolbachia infection has driven a selective sweep of co-inherited mtDNA through the population, causing a loss of mtDNA diversity [41]. High nuclear diversity (relative to sympatric P. xylostella) supports this hypothesis, because a demographic bottleneck should reduce diversity across the entire genome [34].
Plutella australiana and P. xylostella have co-existed in Australia for at least 125 years (\(\geqslant \)1300 generations), yet have strongly divergent mitochondrial and nuclear genomes, Wolbachia infections and insecticide susceptibility phenotypes. Our observations during laboratory rearing and crossing experiments also suggested that interspecific differences in host plant use may exist. What explains such strong divergence between the two Plutella species, given sympatry and the capacity to hybridize? Endemism of P. australiana [16] implies an ancient evolutionary history in Australia, and our data provide support for existing views that Australian P. xylostella were recently introduced from a small ancestral source population, possibly from Asia [17, 18, 77]. Therefore, the two Plutella species may have diverged in allopatry and recently come into secondary contact. Maintenance of divergence suggests strong continuing reproductive isolation, which can evolve as a side-effect of allopatric divergence [44]. All 99 individuals that were RAD sequenced showed concordance in nuclear multilocus genotypes and mtDNA genotypes identified through PCR-RFLP regardless of geographic location, as shown by STRUCTURE analysis. Cryptic species in sympatry provides strong evidence of limited genetic exchange [79]. A small degree of genotypic admixture evident for a few individuals in the STRUCTURE plots might be explained by ancestral polymorphism or introgressive hybridization [28], or alternatively, could be an artefact if our dataset is not representative of the entire genetic background [33]. The level of hybridization that may be occurring between these species is unknown. Isolation may not be uniform across the genome [92, 93], and scans of larger genomic regions may be required to identify introgression and detect hybrids.
The factors leading to reproductive isolation between the two Plutella species in nature are unknown but could include a range of pre- or post-mating isolation mechanisms, such as assortive mating or hybrid fitness costs. Behavioural mating choices are often the main isolating factor in sympatric animals [86]. Does Wolbachia cause a reproductive barrier? The contrast in infection status creates the potential for cytoplasmic incompatibility between species [94]. Interspecific crosses showed a pattern of asymmetric isolation consistent with the expected effects of unidirectional CI, where 21% crosses involving infected P. australiana females produced viable offspring, while the reciprocal CI-cross direction (uninfected P. xylostella females crossed with infected P. australiana males) was nearly sterile. However, this pattern was not continued in the F1 generation: infected hybrid males (derived from the P. australiana maternal line) produced offspring at comparable rates when back-crossed to either uninfected P. xylostella or infected P. australiana female parents. The role of Wolbachia-induced postzygotic isolation between the two Plutella species requires further study, though our results suggest it could be more important in the F0 generation. Wolbachia can contribute to post-zygotic genetic isolation after speciation by complementing hybrid incompatibilities [94, 95]. Symbiont infections could also influence mating behaviour and contribute to pre-mating isolation [96].
Conclusions
The discovery of cryptic pest species introduces complexities for their management and also exciting opportunities for understanding ecological traits. We found strong genomic and phenotypic divergence in two cryptic mitochondrial Plutella lineages co-existing in nature, supporting their status as distinct species [16] despite the capacity to hybridize. Reproductive isolation is likely to have evolved during allopatric speciation, and genome-wide sequence data suggest it has been maintained following secondary contact. Variation in Wolbachia infections might be one factor reinforcing reproductive barriers.
Plutella australiana co-occurs with P. xylostella throughout agricultural regions of southern Australia, but made up only 10% of Plutella juveniles collected from cultivated and wild brassicaceous plants. A lack of population structure across neutral SNP markers suggests that P. australiana populations are linked by high levels of gene flow, and also that P. australiana is a highly mobile species, which is supported by light trap collections [16] and seasonal colonization of canola crops. Future molecular analysis of Australian Plutella should include a species identification step using a molecular diagnostic assay. For ecological studies, it may be possible to perform molecular species identification to confidently distinguish a representative sub-sample of individuals or pooled samples. Our study has shown that while P. australiana can attack canola crops, there is no evidence of pest status in commercial Brassica vegetables crops, and bioassays suggested that field populations should be easily controlled with insecticides. Though P. australiana is a potential pest of some Australian Brassica crops, it is of secondary importance to the diamondback moth, P. xylostella.
References
Pfenninger M, Schwenk K. Cryptic animal species are homogeneously distributed among taxa and biogeographical regions. BMC Evol Biol. 2007; 7:121. https://doi.org/10.1186/1471-2148-7-121.
De Barro PJ, Liu SS, Boykin LM, Dinsdale AB. Bemisia tabaci: A statement of species status. Annu Rev Entomol. 2011; 56:1–19. https://doi.org/10.1146/annurev-ento-112408-085504.
Coetzee M, Hunt RH, Wilkerson R, Della Torre A, Coulibaly MB, Besansky NJ. Anopheles coluzzii and Anopheles amharicus, new members of the Anopheles gambiae complex. Zootaxa. 2013; 3619(3):246–74. https://doi.org/10.11646/zootaxa.3619.3.2.
Hendrichs J, Teresa Vera M, De Meyer M, Clarke AR. Resolving cryptic species complexes of major tephritid pests. ZooKeys. 2015; 540:5–39. https://doi.org/10.3897/zookeys.540.9656.
Rugman-Jones PF, Hoddle MS, Stouthamer R. Nuclear-mitochondrial barcoding exposes the global pest Western flower thrips (Thysanoptera: Thripidae) as two sympatric cryptic species in its native California. J Econ Entomol. 2010; 103(3):877–86. https://doi.org/10.1603/EC09300.
Jacobson AL, Nault BA, Vargo EL, Kennedy GG. Restricted gene flow among lineages of Thrips tabaci supports genetic divergence among cryptic species groups. PLoS ONE. 2016;11(9). https://doi.org/10.1371/journal.pone.0163882.
Skoracka A, Kuczynski L, Szydlo W, Rector B. The wheat curl mite Aceria tosichella (Acari: Eriophyoidea) is a complex of cryptic lineages with divergent host ranges: Evidence from molecular and plant bioassay data. Biol J Linn Soc. 2013; 109(1):165–80. https://doi.org/10.1111/bij.12024.
Miller AD, Skoracka A, Navia D, de Mendonca RS, Szydlo W, Schultz MB, Smith CM, Truol G, Hoffmann AA. Phylogenetic analyses reveal extensive cryptic speciation and host specialization in an economically important mite taxon. Mol Phylogenet Evol. 2013; 66(3):928–40. https://doi.org/10.1016/j.ympev.2012.11.021.
Umina P, Hoffmann A, Weeks A. Biology, ecology and control of the Penthaleus species complex (Acari: Penthaleidae). Exp Appl Acarol. 2004; 34(3-4):211–37. https://doi.org/10.1007/s10493-004-1804-z.
Ashfaq M, Hebert PDN, Mirza MS, Khan AM, Mansoor S, Shah GS, Zafar Y. DNA barcoding of Bemisia tabaci complex (Hemiptera: Aleyrodidae) reveals southerly expansion of the dominant whitefly species on cotton in Pakistan. PLoS ONE. 2014; 9(8):e104485. https://doi.org/10.1371/journal.pone.0104485.
Furlong MJ, Wright DJ, Dosdall LM. Diamondback moth ecology and management: Problems, progress, and prospects. Annu Rev Entomol. 2013; 58:517. https://doi.org/10.1146/annurev-ento-120811-153605.
Zalucki MP, Shabbir A, Silva R, Adamson D, Shu-Sheng L, Furlong MJ. Estimating the economic cost of one of the world’s major insect pests, Plutella xylostella (Lepidoptera: Plutellidae): Just how long is a piece of string?J Econ Entomol. 2012; 105(4):1115–29. https://doi.org/10.1603/EC12107.
Li Z, Feng X, Liu SS, You M, Furlong MJ. Biology, ecology, and management of the diamondback moth in China. Annu Rev Entomol. 2016; 61:277–96. https://doi.org/10.1146/annurev-ento-010715-023622.
Endersby NM, McKechnie SW, Ridland PM, Weeks AR. Microsatellites reveal a lack of structure in Australian populations of the diamondback moth, Plutella xylostella (L.)Mol Ecol. 2006; 15(1):107–18. https://doi.org/10.1111/j.1365-294X.2005.02789.x.
Furlong MJ, Spafford H, Ridland PM, Endersby NM, Edwards OR, Baker GJ, Keller MA, Paull CA. Ecology of diamondback moth in Australian canola: Landscape perspectives and the implications for management. Aust J Exp Agr. 2008; 48(12):1494–505. https://doi.org/10.1071/EA07413.
Landry JF, Hebert PDN. Plutella australiana (Lepidoptera, Plutellidae), an overlooked diamondback moth revealed by DNA barcodes. ZooKeys. 2013; 327:43–63. https://doi.org/10.3897/zookeys.327.5831.
Saw J, Endersby NM, McKechnie SW. Low mtDNA diversity among widespread Australian diamondback moth Plutella xylostella (L.) suggests isolation and a founder effect. Insect Sci. 2006; 13(5):365–373.
Delgado AM, Cook JM. Effects of a sex-ratio distorting endosymbiont on mtDNA variation in a global insect pest. BMC Evol Biol. 2009;9:49. https://doi.org/10.1186/1471-2148-9-49.
Roux O, Gevrey M, Arvanitakis L, Gers C, Bordat D, Legal L. ISSR-PCR: Tool for discrimination and genetic structure analysis of Plutella xylostella populations native to different geographical areas. Mol Phylogenet Evol. 2007; 43(1):240–50. https://doi.org/10.1016/j.ympev.2006.09.017.
Pichon A, Arvanitakis L, Roux O, Kirk A, Alauzet C, Bordat D, Legal L. Genetic differentiation among various populations of the diamondback moth, Plutella xylostella (Lepidoptera : Yponomeutidae). B Entomol Res. 2006; 96(2):137–44. https://doi.org/10.1079/BER2005409.
Endersby NM, Viduka K, Baxter SW, Saw J, Heckel DG, McKechnie SW. Widespread pyrethroid resistance in Australian diamondback moth, Plutella xylostella (L.), is related to multiple mutations in the para sodium channel gene. B Entomol Res. 2011; 101(4):393–405. https://doi.org/10.1017/S0007485310000684.
Baker G. Crucifer vegetable insecticide resistance management strategies and issues in Australia In: Srinivasan R, Shelton AM, Collins HL, editors. The Sixth International Workshop on Management of the Diamondback Moth and Other Crucifer Insect Pests. Tainan, Taiwan: AVRDC – The World Vegetable Center: 2011. p. 21–5.
Sarfraz M, Dosdall L, Keddie B. Diamondback moth-host plant interactions: Implications for pest management. Crop Prot. 2006; 25(7):625–39. https://doi.org/10.1016/j.cropo.2005.09.011.
Robinson GS, Sattler K. Plutella in the Hawaiian Islands: Relatives and host-races of the diamondback moth (Lepidoptera: Plutellidae). Bishop Museum Occasional Papers. 2001; 67:1–27.
Endersby NM, Ridland PM, Hoffmann AA. The effects of local selection versus dispersal on insecticide resistance patterns: Longitudinal evidence from diamondback moth (Plutella xylostella (Lepidoptera: Plutellidae)) in Australia evolving resistance to pyrethroids. B Entomol Res. 2008; 98(2):145–57. https://doi.org/10.1017/S0007485307005494.
Ridland P, Endersby N. The Management of Diamondback Moth and Other Crucifer Pests: Proceedings of the Fifth International Workshop In: Shelton, Anthony M, Collins, Hilda L, Zhang Y, Wu Q, editors. Beijing, China: China Agricultural Science and Technology Press: 2008. p. 90–101.
Clarkson CS, Weetman D, Essandoh J, Yawson AE, Maslen G, Manske M, Field SG, Webster M, Antao T, MacInnis B, Kwiatkowski D, Donnelly MJ. Adaptive introgression between Anopheles sibling species eliminates a major genomic island but not reproductive isolation. Nat Commun. 2014;5. https://doi.org/10.1038/ncomms5248.
Hedrick PW. Adaptive introgression in animals: Examples and comparison to new mutation and standing variation as sources of adaptive variation. Mol Ecol. 2013; 22(18):4606–18. https://doi.org/10.1111/mec.12415.
Hebert P, Penton E, Burns J, Janzen D, Hallwachs W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. P Natl Acad Sci USA. 2004; 101(41):14812–7. https://doi.org/10.1073/pnas.0406166101.
Ashfaq M, Hebert PDN. DNA barcodes for bio-surveillance: Regulated and economically important arthropod plant pests. Genome. 2016; 59(11, 2):933–45. https://doi.org/10.1139/gen-2016-0024.
Smith MA, Bertrand C, Crosby K, Eveleigh ES, Fernandez-Triana J, Fisher BL, Gibbs J, Hajibabaei M, Hallwachs W, Hind K, Hrcek J, Huang DW, Janda M, Janzen DH, Li Y, Miller SE, Packer L, Quicke D, Ratnasingham S, Rodriguez J, Rougerie R, Shaw MR, Sheffield C, Stahlhut JK, Steinke D, Whitfield J, Wood M, Zhou X. Wolbachia and DNA barcoding insects: Patterns, potential, and problems. PLoS ONE. 2012; 7(5):e36514. https://doi.org/10.1371/journal.pone.0036514.
Whitworth TL, Dawson RD, Magalon H, Baudry E. DNA barcoding cannot reliably identify species of the blowfly genus Protocalliphora (Diptera: Calliphoridae). P Roy Soc B-Biol Sci. 2007; 274(1619):1731–9. https://doi.org/10.1098/rspb.2007.0062.
Dupont L, Porco D, Symondson WOC, Roy V. Hybridization relics complicate barcode-based identification of species in earthworms. Mol Ecol Resour. 2016; 16(4):883–94. https://doi.org/10.1111/1755-0998.12517.
Hurst G, Jiggins F. Problems with mitochondrial DNA as a marker in population, phylogeographic and phylogenetic studies: The effects of inherited symbionts. P Roy Soc B-Biol Sci. 2005; 272(1572):1525–34. https://doi.org/10.1098/rspb.2005.3056.
Ritter S, Michalski SG, Settele J, Wiemers M, Fric ZF, Sielezniew M, Sasic M, Rozier Y, Durka W. Wolbachia infections mimic cryptic speciation in two parasitic butterfly species, Phengaris teleius and P. nausithous (Lepidoptera: Lycaenidae). PLoS ONE. 2013; 8(11). https://doi.org/10.1371/journal.pone.0078107.
Weinert LA, Araujo-Jnr EV, Ahmed MZ, Welch JJ. The incidence of bacterial endosymbionts in terrestrial arthropods. P Roy Soc B-Biol Sci. 2015; 282(1807):20150249. https://doi.org/10.1098/rspb.2015.0249.
Ahmed MZ, Breinholt JW, Kawahara AY. Evidence for common horizontal transmission of Wolbachia among butterflies and moths. BMC Evol Biol. 2016; 16:118. https://doi.org/10.1186/s12862-016-0660-x.
Werren JH, Baldo L, Clark ME. Wolbachia: Master manipulators of invertebrate biology. Nat Rev Microbiol. 2008; 6(10):741–51. https://doi.org/10.1038/nrmicro1969.
Engelstaedter J, Hurst GDD. The ecology and evolution of microbes that manipulate host reproduction. Annu Rev Ecol Evol Syst. 2009; 40:127–49. https://doi.org/10.1146/annurev.ecolsys.110308.120206.
Jiggins FM. The spread of Wolbachia through mosquito populations. PLoS Biol. 2017; 15(6):e2002780. https://doi.org/10.1371/journal.pbio.2002780.
Shoemaker D, Dyer K, Ahrens M, McAbee K, Jaenike J. Decreased diversity but increased substitution rate in host mtDNA as a consequence of Wolbachia endosymbiont infection. Genetics. 2004; 168(4):2049–58. https://doi.org/10.1534/genetics.104.030890.
Batista PD, Keddie BA, Dosdall LM, Harris HL. Phylogenetic placement and evidence for horizontal transfer of Wolbachia in Plutella xylostella (Lepidoptera: Plutellidae) and its parasitoid, Diadegma insulare (Hymenoptera: Ichneumonidae). Can Entomol. 2010; 142(1):57–64. https://doi.org/10.4039/n09-050.
Jeyaprakash A, Hoy M. Long PCR improves Wolbachia DNA amplification: wsp sequences found in 76% of sixty-three arthropod species. Insect Mol Biol. 2000; 9(4):393–405. https://doi.org/10.1046/j.1365-2583.2000.00203.x.
Telschow A, Hilgenboecker K, Hammerstein P, Werren JH. Dobzhansky-Muller and Wolbachia-induced incompatibilities in a diploid genetic system. PLoS ONE. 2014; 9(4):e95488. https://doi.org/10.1371/journal.pone.0095488.
Dumas E, Atyame CM, Milesi P, Fonseca DM, Shaikevich EV, Unal S, Makoundou P, Weill M, Duron O. Population structure of Wolbachia and cytoplasmic introgression in a complex of mosquito species. BMC Evol Biol. 2013;13. https://doi.org/10.1186/1471-2148-13-181.
Munoz AG, Baxter SW, Linares M, Jiggins CD. Deep mitochondrial divergence within a Heliconius butterfly species is not explained by cryptic speciation or endosymbiotic bacteria. BMC Evol Biol. 2011:358. https://doi.org/10.1186/1471-2148-11-358.
Sun XJ, Xiao JH, Cook JM, Feng G, Huang DW. Comparisons of host mitochondrial, nuclear and endosymbiont bacterial genes reveal cryptic fig wasp species and the effects of Wolbachia on host mtDNA evolution and diversity. BMC Evol Biol. 2011; 11:86. https://doi.org/10.1186/1471-2148-11-86.
Zraket C, Barth J, Heckel D, Abbott A. Genetic Linkage Mapping with Restriction Fragment Length Polymorphisms in the Tobacco Budworm, Heliothis virescens: Springer; 1990. pp. 13–20. http://dx.doi.org/10.1007/978-1-4899-3668-4_2.
Perry KD, Pederson SM, Baxter SW. Genome-wide SNP discovery in field and laboratory colonies of Australian Plutella species. bioRxiv. 2017. https://doi.org/10.1101/141606. http://www.biorxiv.org/content/early/2017/05/24/141606.full.pdf.
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, Thierer T, Ashton B, Meintjes P, Drummond A. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012; 28(12):1647–9. https://doi.org/10.1093/bioinformatics/bts199.
Paradis E. pegas: An R package for population genetics with an integrated-modular approach. Bioinformatics. 2010; 26(3):419–20. https://doi.org/10.1093/bioinformatics/btp696.
Simoes PM, Mialdea G, Reiss D, Sagot MF, Charlat S. Wolbachia detection: An assessment of standard PCR Protocols. Mol Ecol Resour. 2011; 11(3):567–72. https://doi.org/10.1111/j.1755-0998.2010.02955.x.
Zhou W, Rousset F, O’Neill S. Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences. P Roy Soc B-Biol Sci. 1998; 265(1395):509–15. https://doi.org/10.1098/rspb.1998.0324.
Stamatakis A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014; 30(9):1312–3. https://doi.org/10.1093/bioinformatics/btu033.
Tavaré S. Some probabilistic and statistical problems in the analysis of DNA sequences. Lect Math Life Sci. 1986; 17:57–86.
Baird NA, Etter PD, Atwood TS, Currey MC, Shiver AL, Lewis ZA, Selker EU, Cresko WA, Johnson EA. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE. 2008; 3(10):e3376. https://doi.org/10.1371/journal.pone.0003376.
Baxter SW, Davey JW, Johnston JS, Shelton AM, Heckel DG, Jiggins CD, Blaxter ML. Linkage mapping and comparative genomics using next-generation RAD sequencing of a non-model organism. PLoS ONE. 2011; 6(4):e19315. https://doi.org/10.1371/journal.pone.0019315.
Bolger AM, Lohse M, Usadel B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics. 2014; 30(15):2114–20. https://doi.org/10.1093/bioinformatics/btu170.
Lunter G, Goodson M. Stampy: A statistical algorithm for sensitive and fast mapping of Illumina sequence reads. Genome Res. 2011; 21(6):936–9. https://doi.org/10.1101/gr.111120.110.
Broad Institute. http://broadinstitute.github.io/picard/. Accessed 10 December 2017.
McKenna A, Hanna M, Banks E, Sivachenko A, Cibulskis K, Kernytsky A, Garimella K, Altshuler D, Gabriel S, Daly M, DePristo MA. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010; 20(9):1297–303. https://doi.org/10.1101/gr.107524.110.
DePristo MA, Banks E, Poplin R, Garimella KV, Maguire JR, Hartl C, Philippakis AA, Del Angel G, Rivas MA, Hanna M. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat Genet. 2011; 43(5):491–8. https://doi.org/10.1038/ng.806.
Danecek P, Auton A, Abecasis G, Albers CA, Banks E, DePristo MA, et al.The variant call format and VCFtools. Bioinformatics. 2011; 27(15):2156–8. https://doi.org/10.1093/bioinformatics/btr330.
Goudet J, Jombart T. hierfstat: Estimation and Tests of Hierarchical F-statistics. 2015. R package version 0.04-22. https://CRAN.R-project.org/package=hierfstat.
Nei M. Molecular Evolutionary Genetics: Columbia University Press; 1987.
Martin S. https://github.com/simonhmartin. Accessed 30 March 2018.
R Core Team. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2017. https://www.R-project.org/.
Weir B, Cockerham C. Estimating F-statistics for the analysis of population-structure. Evolution. 1984; 38(6):1358–70. https://doi.org/10.2307/2408641.
Keenan K, McGinnity P, Cross TF, Crozier WW, Prodoehl PA. diveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors. Methods Ecol Evol. 2013; 4(8):782–8. https://doi.org/10.1111/2041-210X.12067.
Rousset F. GENEPOP ‘007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol Ecol Resour. 2008; 8(1):103–6. https://doi.org/10.1111/j.1471-8286.2007.01931.x.
Pritchard J, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data. Genetics. 2000; 155(2):945–59.
Evanno G, Regnaut S, Goudet J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol Ecol. 2005; 14(8):2611–20. https://doi.org/10.1111/j.1365-294X.2005.02553.x.
Earl DA, vonHoldt BM. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour. 2012; 4(2):359–61. https://doi.org/10.1007/s12686-011-9548-7.
Jakobsson M, Rosenberg NA. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics. 2007; 23(14):1801–6. https://doi.org/10.1093/bioinformatics/btm233.
Rosenberg N. DISTRUCT: A program for the graphical display of population structure. Mol Ecol Notes. 2004; 4(1):137–8. https://doi.org/10.1046/j.1471-8286.2003.00566.x.
Ritz C, Baty F, Streibig JC, Gerhard D. Dose-response analysis using R. PLoS ONE. 2015; 10(12):e0146021. https://doi.org/10.1371/journal.pone.0146021.
Juric I, Salzburger W, Balmer O. Spread and global population structure of the diamondback moth Plutella xylostella (Lepidoptera: Plutellidae) and its larval parasitoids Diadegma semiclausum and Diadegma fenestrale (Hymenoptera: Ichneumonidae) based on mtDNA. B Entomol Res. 2017; 107(2):155–64. https://doi.org/10.1017/S0007485316000766.
Kalinowski ST. The computer program STRUCTURE does not reliably identify the main genetic clusters within species: Simulations and implications for human population structure. Heredity. 2011; 106(4):625–32. https://doi.org/10.1038/hdy.2010.95.
Bickford D, Lohman DJ, Sodhi NS, Ng PKL, Meier R, Winker K, Ingram KK, Das I. Cryptic species as a window on diversity and conservation. Trends Ecol Evol. 2007; 22(3):148–55. https://doi.org/10.1016/j.tree.2006.11.004.
Australasian Virtual Herbarium. https://avh.chah.org.au/. Accessed 20 July 2017.
Talekar N, Shelton A. Biology, ecology, and management of the diamondback moth. Annu Rev Entomol. 1993; 38:275–301. https://doi.org/10.1146/annurev.en.38.010193.001423.
Pivnick K, Jarvis B, Gillott C, Slater G, Underhill E. Daily patterns of reproductive activity and the influence of adult density and exposure to host plants on reproduction in the diamondback moth (Lepidoptera, Plutellidae). Environ Entomol. 1990; 19(3):587–93. https://doi.org/10.1093/ee/19.3.587.
Sun JY, Sonderby IE, Halkier BA, Jander G, de Vos M. Non-volatile intact indole glucosinolates are host recognition cues for ovipositing Plutella xylostella. J Chem Ecol. 2009; 35(12):1427–36. https://doi.org/10.1007/s10886-009-9723-4.
Renwick J, Haribal M, Gouinguene S, Stadler E. Isothiocyanates stimulating oviposition by the diamondback moth, Plutella xylostella. J Chem Ecol. 2006; 32(4):755–66. https://doi.org/10.1007/s10886-006-9036-9.
Justus K, Mitchell B. Oviposition site selection by the diamondback moth, Plutella xylostella (L) (Lepidoptera: Plutellidae). J Insect Behav. 1996; 9(6):887–98. https://doi.org/10.1007/BF02208976.
Mallet J. Hybridization as an invasion of the genome. Trends Ecol Evol. 2005; 20(5):229–37. https://doi.org/10.1016/j.tree.2005.02.010.
Turelli M, Moyle LC. Asymmetric postmating isolation: Darwin’s corollary to Haldane’s rule. Genetics. 2007; 176(2):1059–88. https://doi.org/10.1534/genetics.106.065979.
Haldane J. Sex ratio and unisexual sterility in hybrid animals. J Genet. 1922; 12(2):101–9. https://doi.org/10.1007/BF02983075.
Jiggins C, Linares M, Naisbit R, Salazar C, Yang Z, Mallet J. Sex-linked hybrid sterility in a butterfly. Evolution. 2001; 55(8):1631–8. https://doi.org/10.1111/j.0014-3820.2001.tb00682.x.
Turelli M, Orr H. Dominance, epistasis and the genetics of postzygotic isolation. Genetics. 2000; 154(4):1663–79.
Hilgenboecker K, Hammerstein P, Schlattmann P, Telschow A, Werren J. H. How many species are infected with Wolbachia? - a statistical analysis of current data. Fems Microbiol Lett. 2008; 281(2):215–20. https://doi.org/10.1111/j.1574-6968.2008.01110.x.
Harrison RG, Larson EL. Hybridization, introgression, and the nature of species boundaries. J Hered. 2014; 105(1):795–809. https://doi.org/10.1093/jhered/esu033.
Harrison RG, Larson EL. Heterogeneous genome divergence, differential introgression, and the origin and structure of hybrid zones. Mol Ecol. 2016; 25(11, SI):2454–66. https://doi.org/10.1111/mec.13582.
Jaenike J, Dyer KA, Cornish C, Minhas MS. Asymmetrical reinforcement and Wolbachia infection in Drosophila. PLoS Biol. 2006; 4(10):1852–62. https://doi.org/10.1371/journal.pbio.0040325.
Gebiola M, Kelly SE, Hammerstein P, Giorgini M, Hunter MS. “Darwin’s corollary" and cytoplasmic incompatibility induced by Cardinium may contribute to speciation in Encarsia wasps (Hymenoptera: Aphelinidae). Evolution. 2016; 70(11):2447–58. https://doi.org/10.1111/evo.13037.
Shropshire JD, Bordenstein SR. Speciation by symbiosis: The microbiome and behavior. MBIO. 2016; 7(2):e01785-15. https://doi.org/10.1128/mBio.01785-15.
Acknowledgements
We thank colleagues who collected Plutella samples: Adam Hancock, Adam Pearce, Adam Quade, Alan Lord, Andrew McMahen, Andy Bates, Andy Ryland, Brenton Spriggs, Chris Davey, Chris Teague, Craig James, Dustin Berryman, Grant Hudson, Guy Westmore, James McKee, Jessica Smith, Joanne Holloway, Josh Andrews, Josh Hollitt, Karina Bennett, Laura Archer, Levon Cookson, Lisa Ohlson, Louise Flohr, Melina Miles, Michael Collins, Monica Field, Nigel Myers, Peter Cole, Peter Ellison, Peter Gregg, Peter Mangano, Richard Saunders, Sarina Macfadyen, Stewart Learmonth. We thank Lynn Dieckow and Birte Albrecht for assistance with genotyping and library preparation.
Funding
KDP was supported by the University of Adelaide (UA00146) and the Grains Research and Development Corporation (GRDC) (DAS00094), GJB, KJP and JKK were supported by GRDC (DAS00155), and SWB was supported by the Australian Research Council (DP120100047, FT140101303). The funding bodies had no role in the design of the study, the collection, analysis, and interpretation of data or writing the manuscript.
Availability of data and materials
The DNA sequence data are available from GenBank under the accessions MF804301-MF804314 (Wolbachia wsp) and MF151826-MF151906 (mitochondrial COI). The raw RADseq fastq files will be submitted to the NCBI Sequence Read Archive.
Author information
Authors and Affiliations
Contributions
All authors conceived and designed experiments. KDP collected Plutella samples, and KDP and SWB performed RADseq, COI genotyping, data analysis and wrote the manuscript. SWB and CMW genotyped Wolbachia and sequenced wsp. KJP, JKK and GJB cultured Plutella strains and performed insecticide bioassays and crossing experiments. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1
Table S1 The four variable nucleotide sites among the five P. xylostella 613 bp COI haplotypes identified in 102 individuals from Australia. Table S2 The eight variable nucleotide sites among the nine P. australiana 613 bp COI haplotypes identified in 87 individuals from Australia. Table S3 Log-logistic regression statistics for dose-response bioassays on P. australiana and P. xylostella field strains and the P. xylostella reference strain exposed to four commercial insecticides. (PDF 91 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License(http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver(http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Perry, K., Baker, G., Powis, K. et al. Cryptic Plutella species show deep divergence despite the capacity to hybridize. BMC Evol Biol 18, 77 (2018). https://doi.org/10.1186/s12862-018-1183-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12862-018-1183-4