
Abstract
Background
The increasing prevalence of heated tobacco products (HTPs) has heightened concerns regarding their potential health risks. Previous studies have demonstrated the toxicity of cigarette smoke extract (CSE) from traditional tobacco’s mainstream smoke, even after the removal of nicotine and tar. Our study aimed to investigate the cytotoxicity of CSE derived from HTPs and traditional tobacco, with a particular focus on the role of reactive oxygen species (ROS) and intracellular Ca2+.
Methods
A human oral squamous cell carcinoma (OSCC) cell line, HSC-3 was utilized. To prepare CSE, aerosols from HTPs (IQOS) and traditional tobacco products (1R6F reference cigarette) were collected into cell culture media. A cell viability assay, apoptosis assay, western blotting, and Fluo-4 assay were conducted. Changes in ROS levels were measured using electron spin resonance spectroscopy and the high-sensitivity 2ʹ,7ʹ-dichlorofluorescein diacetate assay. We performed a knockdown of calcium/calmodulin-dependent protein kinase kinase 2 (CaMKK2) by shRNA lentivirus in OSCC cells.
Results
CSE from both HTPs and traditional tobacco exhibited cytotoxic effects in OSCC cells. Exposure to CSE from both sources led to an increase in intracellular Ca2+ concentration and induced p38 phosphorylation. Additionally, these extracts prompted cell apoptosis and heightened ROS levels. N-acetylcysteine (NAC) mitigated the cytotoxic effects and p38 phosphorylation. Furthermore, the knockdown of CaMKK2 in HSC-3 cells reduced cytotoxicity, ROS production, and p38 phosphorylation in response to CSE.
Conclusion
Our findings suggest that the CSE from both HTPs and traditional tobacco induce cytotoxicity. This toxicity is mediated by ROS, which are regulated through Ca2+ signaling and CaMKK2 pathways.
Graphical Abstract

Similar content being viewed by others


Background
Tobacco is responsible for over 8 million deaths annually worldwide, with over 8 million deaths attributed to direct tobacco use [1]. Tobacco smoke, a blend of over 5000 harmful and carcinogenic chemicals, includes 98 particularly dangerous substances [2]. Smokers have a fivefold higher risk of oral cancer [3].
Cigarette smoke is composed of two phases: the particulate phase and the gas phase. The particulate phase mainly consists of nicotine, tar, and water, while the gas phase contains a number of carbonyl compounds, such as formaldehyde, acrolein, and tobacco-specific nitrosamines. While tar and nicotine are often highlighted regarding the harm of traditional tobacco, reports indicate that not only the particulate phase of tobacco, but also its gas phase has various effects on the body [4, 5]. Research indicates that the gas phase of tobacco activates nicotinamide adenine dinucleotide phosphate (NADPH) and induces cytotoxicity. Furthermore, the tobacco gas phase increases reactive oxygen species (ROS) in human umbilical vein endothelial cells (HUVECs), and inhibits thromboxane A2, thereby affecting platelet aggregation [6, 7]. In recent years, there has been a global surge in demand for heated tobacco products (HTPs). These products heat processed tobacco leaves at temperatures far below the combustion temperature of combustible cigarettes to generate nicotine-containing aerosols, which users inhale into their lungs. In the United States, brands of HTPs that are permitted for sale include IQOS (‘I-Quit-Ordinary-Smoking’; Philip Morris International [PMI]), Eclipse (R. J. Reynolds [RJR]) and Glo (British American Tobacco [BAT]), while Ploom (Japan Tobacco Inc [JTI]) is sold in Japan. The global HTPs market was valued at 13.6 billion USD in 2021 and is expected to grow to 947.8 billion USD by 2030 [8]. Although HTPs are purported to reduce the amounts of harmful and potentially harmful substances generated, further research is necessary to understand the short-term and long-term health impacts of HTPs.
The toxicity of traditional tobacco is known to be associated with the involvement of reactive oxygen ROS and Ca2+ [9, 10]. On the other hand, there is little knowledge of these in the context of HTPs. ROS act as second messengers and play a role in regulating normal physiological processes and cellular homeostasis [11]. To regulate ROS levels, various mechanisms exist, including localized generation, detoxification by antioxidant factors. In human cells, there are many enzymes, such as NAPDH oxidases, that generate hydrogen peroxide and superoxide [12]. However, when ROS are produced excessively, they can induce oxidative stress, resulting in damage to biomolecules and contributing to the development of various diseases, including those affecting the respiratory, neurodegenerative, and digestive systems. The production of ROS in smoking is well-known, and the presence of ROS in tobacco smoke is a significant factor in the oxidative stress and damage caused by smoking [9].
Ca2+ is also a versatile second messenger that is involved in multiple functions in eukaryotic cells, such as signal transduction, muscle contraction, apoptosis, cell cycle progression, cell migration and cell proliferation [13]. They function as a signal transduction pathway, in the release of neurotransmitters from neurons, in the contraction of all types of muscle cells, and as a second messenger in fertilization. Many enzymes, including several coagulation factors, require Ca2+ as a cofactor. However, high intracellular Ca2+ concentrations can become toxic, leading to cell death either through necrosis or apoptosis. This is often due to sustained high levels of Ca2+ in mitochondria, which can trigger the release of cytochrome c, followed by the activation of caspases and subsequent death signaling. While Ca2+ has important functions in the body, there are multiple reports that conventional tobacco affects intracellular calcium levels. When HL60 cells were stimulated with particulate phase of traditional tobaccos, an increase in intracellular Ca2+ was observed, which was not seen with nicotine alone [6]. Furthermore, it has been reported that whole particulate phase from traditional tobacco causes an increase in cytosolic Ca2+ concentration, and components of tobacco such as 1-NH2-naphthalene, formaldehyde, nicotine, and nicotine-derived nitrosamine ketone may be involved [10].
The manufacturers claim that HTPs have a reduced toxicity compared to traditional tobacco [14]. PMI reported that the levels of 40 out of the 93 harmful and potentially harmful constituents (HPHC) listed by the FDA are lower in IQOS aerosol than in traditional tobacco smoke [15]. On the other hand, an additional 56 other substances not included in PMI’s list or the FDA’s HPHC list were found at higher levels in IQOS emissions. Of these, 22 types were reported to be over 200% higher and 7 types were over 1000% higher compared to traditional tobacco smoke. Recently, it has become understood that heated tobacco also has toxicity. However, many aspects, such as the specific causes of this toxicity, remain poorly understood. Therefore, FDA does not recognize the reduction of health risks of heated tobacco. Due to differences in the chemicals produced, HTPs may elicit a different pharmacological action of cytotoxicity compared to traditional tobacco.
In the current study, we present our findings on the cytotoxic effects of nicotine- and tar-free cigarette smoke extract (CSE) from heat-not-burn tobacco products (HTPs) and traditional tobacco on oral squamous cell carcinoma (OSCC) cell lines (HSC-3, SCC-25 and OSC-19), as well as on some normal cell lines.
Materials and methods
CSE
The HTPs composing of a heating device (IQOS ILUMA, PMI) and tobacco sticks (TEREA Regular, PMI) was commercially obtained in Japan. Combustible reference cigarettes (1R6F) were purchased from the Center for Tobacco Reference Products at the University of Kentucky College of Agriculture (Lexington, KY, USA). The mainstream smoke from HTPs and traditional tobacco was generated according to the Cooperation Centre for Scientific Research Relative to Tobacco (CORESTA) technical report (HTP-259-CTR) smoking regime for high-temperature HTPs (55 mL puff volume, 30 s puff interval, 2 s puff duration, bell-shaped puff profile, and no blocking of the filter ventilation holes) and the Health Canada Intense (HCI) smoking regime (55 mL puff volume, 30 s puff interval, 2 s puff duration, bell-shaped puff profile, and 100% blocked filter ventilation holes), respectively, by using an analytical vaping machine (LM5E; Körber Technologies Instruments GmbH, Hamburg, Germany) [16, 17]. The particulate phase in the mainstream smoke was collected on a Cambridge filter pad (Körber Technologies Instruments GmbH). The resulting gas phase of ten cigarettes (eight puffs from one cigarette per trial) was bubbled into 5 mL of ice-cold, fetal bovine serum (FBS)-free cell culture media to prepare 80 puffs of CSE solution. The concentration of each CSE solution was considered 100%.
Cell line
Human oral squamous cell carcinoma cell line HSC-3 was purchased from Health Science Research Resources Bank (Japan Health Sciences Foundation). Cell was cultured in High glucose Dulbecco’s modified Eagle’s medium (DMEM; Wako) supplemented with 10% FBS, and 5% penicillin/streptomycin (Thermo Scientific, IL, USA) in an incubator containing 5% CO2 at 37 °C. SCC-25 was obtained from American Type Culture Collection (USA). The OSC-19 used in the supplement data was obtained from the Health Science Research Resources Bank. Normal human astrocyte (NHA) was purchased from Lonza (Basel, Switzerland). Human gingival fibroblasts (HGnF) and oral keratinocytes were purchased from ScienCell Research Laboratories (Carlsbad, CA, USA) HUVEC was purchased from PromoCell (Heidelberg, Germany).
Inhibitor
N-Acetylcysteine (NAC) was purchased from Wako (Osaka, Japan). In this experiment, NAC was used at a concentration of 1250 μM.
Cell viability assay
Cell Counting Kit-8 (CCK-8; Dojin, Kumamoto, Japan) assay was used for cell proliferation viability. Briefly, cells were seeded in a 96-well plate at 5,000 cells per well and cultured for 24 h. In some experiments, we considered for a longer time (24, 48 and 72 h). We added 10 μl of reagent after stimulation, incubated for 2 h, and then measured the absorbance at 450 nm [18].
Apoptosis assay
HSC-3 cells were incubated with or without CSE for 18 h. Flow cytometry using BD fluorescence-activated cell sorting (FACS) Canto II (BD Biosciences, CA, USA) were carried out as previously described. FITC Annexin V Apoptosis Detection Kit with 7-AAD (BioLegend, SD, USA) was used to stain the cells. Cells were divided into four sections. Q1, Q2, Q3, and Q4 marked in each panel indicate the necrosis, late apoptosis, early apoptosis, and lived cells, respectively.
Western blotting
Western blot analyses were carried out as previously described. Briefly, cells were lysed and sonicated in RIPA buffer (Thermo Scientific, IL, USA). The following primary antibodies were used for immunoblotting: phosphor-p38, p38 obtained from CST (Cell Signaling Technology, Danvers, MA, USA), GAPDH obtained from Santa Cruz Biotechnology (Dallas, TX, USA). The calcium/calmodulin-dependent protein kinase kinase 2 (CaMKK2) antibody used to confirm the knockdown efficiency of CaMKK2 was purchased from CST (Supplementary Fig. 3). Chemiluminescence detection was performed using ECL reagent (Bio-Rad Laboratories, CA, USA) and high-sensitivity ECL reagent (Thermo Scientific). Signals were visualized using a LuminoGraph II imaging system (ATTO, Tokyo, Japan). The signal intensities of the bands were quantified using ATTO CS Analyzer 4 software (ATTO).
ROS assay
To investigate the production of ROS, high-sensitive 2ʹ,7ʹ-dichlorofluorescein diacetate assay (HS-DCFH: Dojin) and Electron Spin Resonance (ESR) was performed.
HS-DCFH assay
Cells were incubated with Hanks’ Balanced Salt Solution (HBSS+; Wako) buffer containing of HS-DCFH working solution, followed by washing and incubation with HBSS+. After the CSE stimulation, measurements were performed using a microplate reader (Nivo, PerkinElmer, MA, USA). Fluorescence signals were measured with a microscopy system using an excitation wavelength of 485 nm and an emission of 535 nm.
ESR
ROS production was evaluated using an EMX-8/2.7 ESR spectrometer (Bruker Biospin, Billerica, MA, USA) with/without CSE. 5,5-dimethyl-pyrroline N-oxide (DMPO) was purchased from Labotec (Tokyo, Japan). The sample was injected into a capillary tube and inserted into an ESR measurement device and measured. Testing parameters were set as follows: 20 mW microwave power, 9.85 GHz microwave frequency, 30 s conversion time, five times scans, 2 G field modulation, and 100 G scan range.
Fluorescence imaging of intracellular Ca2+
Cells were incubated with Hanks’ Balanced Salt Solution (HBSS+;Wako) buffer containing 4 µmol/l of Fluo-4AM (Dojin. Kumamoto, Japan), followed by washing and incubation with HEPES-buffered saline containing 2.0 mmol/l of CaCl2. An Eclipse Ti (Nikon Corporation, Tokyo, Japan) was used to monitor fluorescence changes. Full images were collected every 5 s. Fluo-4 fluorescence was excited at 488 nm, and data were expressed as normalized changes in background-corrected fluorescence emission (F/F0). Fluorescence signals were measured at room temperature using an excitation wavelength of 488 nm and an emission of 522.5 nm. Representative Ca2+ signals averaged from 50 individual cells were analyzed.
Short-hairpin RNA transduction
HSC-3 cells were subjected to transduction with CaMKK2 shRNA and scramble control shRNA. Lentiviral transduction by VectorBuilder (Chicago, USA), Sigma‒Aldrich (MO, USA) and SCBT (Santa Cruz Biotechnology, CA, USA) was performed as previously described [19]
The transfection efficiency of the shRNAs was evaluated by Western blotting. Transductions with lentivirus were carried out as previously described. The sequences are shown in Supplementary Table 1.
Data analysis and statistics
Statistical analysis was performed using GraphPad Prism 9 software (GraphPad Software Inc., San Diego, CA, USA). Statistical comparisons between two groups were performed using Student’s t test. Comparisons among more than two groups were performed using one-way analysis of variance (ANOVA) followed by Tukey’s test or by two-way ANOVA followed by the Bonferroni post hoc test. The criterion for statistical significance was set at p < 0.05.
Large language model (LLM)
ChatGPT (GPT-4) was used only for English proofreading.
Results
Substances in the CSE derived from both HTPs and traditional tobacco exhibited cytotoxicity in OSCC cells. Additionally, CSE derived from HTPs had the potential to induce cell proliferation.
To evaluate the effects of substances in the CSE derived from HTPs and traditional tobacco, we conducted a CCK-8 assay in OSCC cells (HSC-3 and SCC-25). Substances in the CSE derived from HTPs suppressed cell viability in a dose-dependent manner (Fig. 1a and c). Similarly, substances in the CSE derived from traditional tobacco also suppressed cell viability in a dose-dependent manner (Fig. 1b and d). Furthermore, we confirmed that substances derived from both HTPs and traditional tobacco products in the CSE also exhibited cytotoxicity in a range of cell types including OSC-19, HGnF, HUVEC, and NHA (Supplementary Fig. 1, a–h).

Substances derived from both HTPs and traditional tobacco products in the CSE exhibited cytotoxicity in HSC-3 cells. a Change in cell proliferation rate of HSC-3 cells after a 24-h exposure to various concentrations of CSE derived from HTPs. b Change in the cell proliferation rate of HSC-3 cells after a 24-h exposure to various concentrations of CSE derived from traditional tobacco. c Change in the cell proliferation rate of SCC-25 cells after a 24-h exposure to various concentrations of CSE derived from HTPs. d Change in the cell proliferation rate of SCC-25 cells after a 24-h exposure to various concentrations of CSE derived from traditional tobacco. e Long-term change in the cell proliferation rate after a 5% CSE derived from HTPs exposure. f Long-term change in the cell proliferation rate after a 15% CSE derived from HTPs exposure. g Long-term change in the cell proliferation rate after a 2.5% CSE derived from traditional tobacco exposure. Statistics: one-way ANOVA (and nonparametric), n = 4, ns; not significant, *; p < 0.05, **; p < 0.01, ***; p < 0.001, ****; p < 0.0001
Interestingly, the CSE from HTPs at low concentrations (5%) appeared to facilitate cell proliferation, while at high concentrations (15%) they demonstrated time-dependent cytotoxic effects (Fig. 1e and f). Furthermore, substances in the 2.5% CSE derived from traditional tobacco suppressed cell proliferation of HSC-3 in a time-dependent manner (Fig. 1g). Our results indicated that high concentrations of CSE from HTPs exhibit cytotoxicity similar to CSE from traditional tobacco, but low concentrations of CSE from HTPs have shown the potential to induce cell proliferation.
Substances in the CSE derived from both HTPs and traditional tobacco promoted cell apoptosis and phosphorylated p38 in OSCC cells
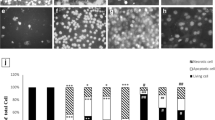
In our subsequent research, we examined how tobacco affects cell apoptosis in OSCC cells. We found that CSE derived from HTPs, specifically IQOS, and traditional tobacco products, like 1R6F (the each amount that shows the same effect of each CSEs on cell viability), induced late-stage apoptosis in HSC-3 cells (Fig. 2a–e).

Substances derived from both HTPs and traditional tobacco in the CSE enhanced cytotoxicity and induced p38 phosphorylation. a Apoptosis assay in the control group. Q1, Q2, Q3, and Q4 marked in each panel indicate the necrosis, late apoptosis, early apoptosis, and lived cells, respectively. b Apoptosis in HSC3 cells 18 h after exposure to 15% CSE derived from HTPs exposure. c Apoptosis in HSC3 cells 18 h after exposure to 2.5% CSE derived from traditional tobacco exposure. d Late apoptosis (percentage of Q2) 18 h after exposure to 15% CSE derived from HTPs exposure. e Late apoptosis (percentage of Q2) 18 h after exposure to 2.5% CSE derived from traditional tobacco exposure. f Western blot (WB) analysis showing p38 phosphorylation following exposure to 15% CSE derived from HTPs. g WB analysis showing p38 phosphorylation following exposure to 2.5% CSE derived from traditional tobacco. Statistics: one-way ANOVA (and nonparametric), Tukey’s multiple comparisons test; d and e (n = 4), f (n = 5), g (n = 6), ns; not significant, *; p < 0.05, **; p < 0.01, ***; p < 0.001
p38 protein belongs to the mitogen-activated protein kinases family that link extracellular stimuli with intracellular responses participating in a number of fundamental cell processes [20]. Gkouveris et al. have reported that the expression of p38 in OSCC cells may be involved in the proliferation of these cells through the mediation of signal transducer and activator of transcription 3.
Therefore, our investigation focused on this process to understand the underlying mechanism of tobacco-induced cytotoxicity in HSC-3 cells. Our findings confirmed that substances from CSE of both HTPs and traditional tobacco products indeed cause p38 phosphorylation in HSC-3 cells (as shown in Fig. 2f and g).
Substances in the CSE derived from both HTPs and traditional tobacco increased ROS production in OSCC cells
Our study found that substances from both HTPs and traditional tobacco products increased the production of ROS in OSCC cells. This was determined by measuring the HS-DCFH assay one hour after stimulation (Fig. 3a, b). When measured for one hour with CSE of HTPs or traditional tobacco at the same time, the ROS production was higher in 2.5% CSE of traditional tobacco than 15% CSE of HTPs (Supplemental Fig. 2). ESR spectroscopy, a highly regarded method for such analysis, indicated an increase in ROS production one hour post-stimulation with both types of tobacco. ESR spectroscopy exhibits characteristic four-line spectra when free radicals bind to DMPO (Fig. 3c–e).

Substances derived from both HTPs and traditional tobacco in the CSE induced ROS production. a ROS measurement following exposure to 15% CSE derived from HTPs exposure, as determined by HS-DCFH assay. b ROS measurement following exposure to 2.5% CSE derived from traditional tobacco exposure, as determined by HS-DCFH assay. c Representative ESR result for ROS measurement in unstimulated cells. d Representative ESR result for ROS measurement following exposure to 15% CSE derived from HTPs. exposure. e Representative ESR result for ROS measurement following exposure to 2.5% CSE derived from traditional tobacco exposure. f WB analysis illustrating p38 phosphorylation following exposure to 15% CSE derived from HTPs in the presence of NAC (1250 μM). g WB analysis illustrating p38 phosphorylation following exposure to 2.5% CSE derived from traditional tobacco in the presence of NAC (1250 μM). h Cell proliferation under NAC (1250 μM) treatment24 h after exposure to 15% CSE derived from HTPs exposure. i Cell proliferation under NAC (1250 μM) treatment 24 h after exposure to 2.5% CSE derived from traditional tobacco exposure. Statistics: one-way ANOVA (and nonparametric), ns; not significant, *; p < 0.05, **; p < 0.01, ***; p < 0.001, ****; p < 0.0001, a and b (n = 4), f and g (n = 5), h and i (n = 6)
It is well-established that ROS production induces apoptosis in cells via the p38 pathway [21]. Given these results showing increased ROS production by tobacco substances, we next evaluated the impact of NAC, a known antioxidant, on tobacco-induced cytotoxicity and p38 phosphorylation. Our findings revealed that NAC reduced the p38 phosphorylation triggered by both tobacco substances (Fig. 3f and g). Furthermore, NAC mitigated the cytotoxic effects induced by these tobacco substances (Fig. 3h and i). Interestingly, HSC-3 stimulated with CSE of HTPs combined with NAC led to cell proliferation compared to the control.
Substances in the CSE derived from both HTPs and traditional tobacco increased intracellular Ca2+ concentration in OSCC cells
Intracellular Ca2+, a ubiquitous second messenger, is vital in regulating numerous cellular processes, such as proliferation, apoptosis and cell migration, including in cancer cells [13]. Wylam et al. previously reported that substances derived from traditional tobacco products increased intracellular Ca2+ concentration [22]. To observe the effect of substances derived from both HTPs and traditional tobacco products on intracellular Ca2+ concentration, we measured the intracellular Ca2+ levels using the Fluo-4 assay, a Ca2+-sensitive dye, immediately after stimulation with these tobacco product ups to 7 min post-stimulation. Similarly, our study found that substances in the CSE derived from both HTPs and traditional tobacco products also elevated intracellular Ca2+ concentration in HSC-3 cells as well (Fig. 4a–c).

Substances derived from both HTPs and traditional tobacco in the CSE increased intracellular Ca2+ concentration. a Changes in intracellular Ca2+ concentration following stimulation with either 5% or 15% CSE derived from HTPs exposure. b Changes in intracellular Ca2+ concentration following stimulation with either 1.25% or2.5% CSE derived from traditional tobacco exposure. c Representative fluorescence image of Ca2+ concentration before and 7 min after exposure to CSE derived from either HTPs or traditional tobacco. Statistics: one-way ANOVA (and nonparametric), ****; p < 0.0001, 50 cells were randomly selected for analysis from the microscope images
Knockdown of CaMKK2 attenuated the CSE derived from HTPs and traditional tobacco substance-induced cytotoxicity, p38 phosphorylation and ROS production
Following our finding that tobacco substances quickly elevate intracellular Ca2+ levels, we shifted our focus to CaMKK2, a calcium-associated protein. Initially, we developed two types of HSC-3 cells with CaMKK2 knockdown (#1 and #2) using shRNA lentivirus and assessed the effects on cytotoxicity, ROS production, and p38 phosphorylation induced by these tobacco substances. We conducted western blotting to investigate the efficiency of CaMKK2 knockdown. It was confirmed that CaMKK2 was significantly suppressed in comparison to the control shRNA (Supplemental Fig. 4). Our results showed that CaMKK2 knockdown reduced cytotoxicity in HSC-3 cells caused by tobacco substances (Fig. 5a and b). Subsequent experiments only used #2, which showed more pronounced suppression of CaMKK2, for comparison with Sh-CTRL. Additionally, CaMKK2 knockdown cells decreased ROS production induced by both tobacco substances (Fig. 5c and d). Furthermore, CaMKK2 knockdown cells also mitigated p38 phosphorylation triggered by both types of tobacco substances (Fig. 5e and f).

Knockdown of CaMKK2 mitigates cytotoxicity, ROS production and p38 phosphorylation in HSC3 cells induced by substances derived from both HTPs and traditional tobacco in the CSE. a Change in cell proliferation rate in CaMKK2 knockdown HSC-3 cells (#1 and #2) following a 24-h exposure to 15% CSE derived from HTPs. b Change in cell proliferation rate in CaMKK2 knockdown HSC-3 cells (#1 and #2) following a 24-h exposure to 2.5% CSE derived from traditional tobacco. c Measurement of ROS in CaMKK2 knockdown HSC-3 cells (#2) following exposure to 15% CSE derived from HTPs exposure, as determined by the HS-DCFH assay. d Measurement of ROS in CaMKK2 knockdown HSC-3 cells (#2) following exposure to 2.5% CSE derived from traditional tobacco exposure, as determined by the HS-DCFH assay. e WB analysis illustrating p38 phosphorylation in CaMKK2 knockdown HSC-3 cells (#2) following exposure to 15% CSE derived from HTPs. f WB analysis illustrating p38 phosphorylation in CaMKK2 knockdown HSC-3 cells (#2) following exposure to 2.5% CSE derived from traditional tobacco. Statistics: one-way ANOVA (and nonparametric), n = 4, ns; not significant, *; p < 0.05, **; p < 0.01, ***; p < 0.001
Discussion
In this research, we compared and verified the known mechanisms of toxicity in traditional tobacco with those in HTPs and investigated whether there are any unidentified signaling pathways. Our study is the first to reveal that the substances in the CSE derived from both HTPs and traditional tobacco promoted cell apoptosis and suppressed cell proliferation through the ROS/p38 pathway and CaMKK2 in OSCC cells. This experiment’s findings are presented in a schematic diagram (Fig. 6). Additionally, CSE derived from HTPs, at certain concentrations, induced cell proliferation, during this process enhanced phosphorylation of p38 and increased intracellular Ca2+ concentration occurred.

Hypothetical pathway of apoptosis stimulation by CSE derived from both HTPs and traditional tobacco. The findings indicate that, similar to traditional tobacco, HTPs induce apoptosis via ROS/p38 signaling pathways. These pathways could potentially be regulated by CaMKK2
Several reports indicate that extracts from the CSE derived from IQOS exhibit lower cytotoxicity than traditional tobacco [23,24,25]. In addition, there are a few reports that heated tobacco caused cell proliferation, and there is also a report that CSE contributed to proliferation [26,27,28]. However, the mechanisms of cytotoxicity and cell proliferation brought about by HTPs were not yet fully understood.
In our study, whether at concentrations that induce cell proliferation or those that cause apoptosis, phosphorylation of p38 was enhanced with CSE derived from HTPs. Phosphorylation of p38 can induce apoptosis through various downstream signaling pathways [21]. Additionally, multiple studies have reported that p38 may be involved in cell proliferation [20, 29, 30]. There are several reports suggesting that traditional tobaccos cause phosphorylation of p38, which is implicated in carcinogenesis and toxicity [31, 32]. Furthermore, the apoptosis mechanism in traditional tobacco is reported to involve the ROS/p38 pathway [33]. Although it has been previously reported that IQOS aerosols can enhance p38 in rat liver cells, our findings suggest that this effect might be attributed to the gas phase of the aerosol [34]. There are also reports that the CSE of HTPs has overactivated p53 [35].
Previous reports have acknowledged that the toxicity of traditional tobacco involves the generation of ROS and an increase in intracellular Ca2+ concentration [4, 22, 36, 37]. Similarly, in our study, CSEs derived from HTPs and traditional tobacco were found to increase ROS activity and intracellular Ca2+ concentration in OSCC cells. For both types of tobacco, suppressing ROS production also reduced cytotoxicity and inhibited the phosphorylation of p38. Notably, combining CSE from HTPs with NAC resulted in cell proliferation. Since combining CSE from HTPs with NAC also resulted in the suppression of phosphorylation of p38, there is a possibility that cell proliferation was induced by other mechanisms, such as other Ca2+-related signals.
It is reported that in traditional tobacco, stimulating human pulmonary artery smooth muscle cells (PASMCs) and endothelial cells (PAECs) leads to COX-2 production, which in turn drives cell proliferation [38]. This cell proliferation is considered abnormal remodeling, suggesting that excessive cell growth can lead to damage in lung tissue. There are multiple reports suggesting that the gaseous components of cigarette smoke can accelerate the cell cycle, leading to uncontrolled cell proliferation, which ultimately may result in carcinogenesis [39]. Similarly, it has been reported that in the case of IQOS, stimulation can lead to accelerated aging of vascular smooth muscle cells [40].
In our study, we observed an increase in intracellular Ca2+ concentration in OSCC cells. To evaluate the toxicity of tobacco extracts on cells, we knocked down CaMKK2, which is associated with Ca2+ elevation, using lentivirus. The results showed that knockdown of CaMKK2 reduced cytotoxicity and decreased ROS production in cells exposed to both HTPs and traditional tobacco. These findings suggest that CaMKK2 may play a role upstream of ROS production.
In this study, the equivalent of 80 puffs of IQOS was extracted into 5 ml of medium. Other literature adjusts IQOS to about 25 puffs/5 ml [24], and 2.5 puffs/5 ml [27], indicating that our study used a more concentrated condition for creating CSEs. For traditional tobacco, a 2.5% concentration involves dissolving the equivalent of 0.4 puffs in 1 ml of solution, whereas for HTPs (IQOS), a 15% concentration involves dissolving 2.4 puffs in 1 ml of solution. These smoking frequencies do not greatly deviate from the actual smoking frequencies of humans. However, it is challenging to directly correlate these CSE concentrations with in vivo conditions. Further research is required to understand how much of the gas-phase components of HTPs and traditional tobacco are actually absorbed and metabolized by body fluids and cells. While HTPs are marketed as safer, it can only be said that if the number of puffs is high, they may have cytotoxicity similar to traditional tobacco. Other in vitro experiments, like those spraying gas directly into the medium instead of using extracts [25], also exist, and differences in stimulation methods on cells must be considered.
Previous reports have acknowledged that the toxicity of traditional tobacco involves the generation of ROS and an increase in intracellular Ca2+ concentration [4, 22, 36, 37]. Similarly, in our study, CSEs derived from HTPs and traditional tobacco were found to increase ROS activity and intracellular Ca2+ concentration in OSCC cells. For both types of tobacco, suppressing ROS production also reduced cytotoxicity and inhibited the phosphorylation of p38. Notably, combining CSE from HTPs with NAC resulted in cell proliferation. Since combining CSE from HTPs with NAC also resulted in the suppression of phosphorylation of p38, there is a possibility that cell proliferation was induced by other mechanisms, such as other Ca2+-related signals.
A limitation of our study is that we only used the CSE from HTPs and traditional tobacco, without analyzing the individual substances in the extracts. Although we standardized the mass of each extract, the specific chemicals in them remain unidentified. While a comparison of toxicity is necessary, the different concentration conditions of HTPs and traditional tobacco mean that the efficacy of directly comparing changes in cellular signaling or ROS production is not necessarily guaranteed. In this study, we did not conduct in vivo experiments due to the challenges associated with testing how the extracts affect tumor cells in a living organism. However, it seems necessary for future research to examine the effects of extracts from HTPs and traditional tobacco on the body, especially on cancer cells.
In our study, we found that HTPs, like traditional tobacco, possess cytotoxicity, but their toxicity might be reduced as claimed by the manufacturers. The introduction of HTPs is touted as a 'harm reduction' strategy for individuals unable to quit nicotine addiction. However, there are reports suggesting that this strategy may be a rationale to circumvent regulations imposed on traditional tobacco products [41].
Substances like acrolein and formaldehyde, which generate ROS and cause toxicity, may be reduced in heated tobacco. However, there might be entirely different substances in heated tobacco that could promote cancer cell proliferation. While traditional tobacco, due to combustion, might produce a wider range of substances, heated tobacco could potentially control the chemicals generated by maintaining a constant temperature. Instead of simply considering heated tobacco as a lower-risk alternative as suggested by manufacturers, further research is needed focusing on the nature and characteristics of individual chemicals produced when using heated tobacco.
Conclusion
Our findings suggest that the CSE from both HTPs and traditional tobacco induce cytotoxicity. This toxicity is mediated by ROS, which are regulated through Ca2+ signaling and CaMKK2 pathways. On one hand, CSE derived from HTPs causes cell proliferation at low concentrations, and identifying its mechanism remains a challenge for future research.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
References
World Health Organization (2023) WHO report on the global tobacco epidemic, 2023: protect people from tobacco smoke.
Talhout R, Schulz T, Florek E, Van Benthem J, Wester P, Opperhuizen A (2011) Hazardous compounds in tobacco smoke. Int J Environ Res Public Health 8:613–628
Chaturvedi P, Singh A, Chien C-Y, Warnakulasuriya S (2019) Tobacco related oral cancer. BMJ 365:l2142
Chen S, Wang Y, Zhang H, Chen R, Lv F, Li Z, Jiang T, Lin D, Zhang H, Yang L, Kong X (2019) The antioxidant MitoQ protects against CSE-induced endothelial barrier injury and inflammation by inhibiting ROS and autophagy in human umbilical vein endothelial cells. Int J Biol Sci 15:1440–1451
Mainali P, Pant S, Rodriguez AP, Deshmukh A, Mehta JL (2015) Tobacco and cardiovascular health. Cardiovasc Toxicol 15:107–116
Asano H, Horinouchi T, Mai Y, Sawada O, Fujii S, Nishiya T, Minami M, Katayama T, Iwanaga T, Terada K, Miwa S (2012) Nicotine- and Tar-free cigarette smoke induces cell damage through reactive oxygen species newly generated by PKC-dependent activation of NADPH oxidase. J Pharm Sci 118:275–287
Kashiwagi H, Yuhki KI, Imamichi Y, Kojima F, Kumei S, Higashi T, Horinouchi T, Miwa S, Narumiya S, Ushikubi F (2017) Cigarette smoke extract inhibits platelet aggregation by suppressing cyclooxygenase activity. TH Open 1:e122–e129
Global Heated Tobacco Products Market. SKYQUEST: 157; 2023
Valavanidis A, Vlachogianni T, Fiotakis K (2009) Tobacco smoke: involvement of reactive oxygen species and stable free radicals in mechanisms of oxidative damage, carcinogenesis and synergistic effects with other respirable particles. Int J Environ Res Public Health 6:445–462
Sassano MF, Ghosh A, Tarran R (2017) Tobacco smoke constituents trigger cytoplasmic calcium release. Appl In Vitro Toxicol 3:193–198
Bardaweel SK, Gul M, Alzweiri M, Ishaqat A, HA AL, Bashatwah RM, (2018) Reactive oxygen species: the dual role in physiological and pathological conditions of the human body. Eurasian J Med 50:193–201
Jha Jay C, Watson Anna MD, Mathew G, de Vos LC, Jandeleit-Dahm K (2017) The emerging role of NADPH oxidase NOX5 in vascular disease. Clin Sci 131:981–990
Berridge MJ, Bootman MD, Roderick HL (2003) Calcium signalling: dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol 4:517–529
McKelvey K, Baiocchi M, Halpern-Felsher B (2020) PMI’s heated tobacco products marketing claims of reduced risk and reduced exposure may entice youth to try and continue using these products. Tob Control 29:e18–e24
St Helen G, Jacob Iii P, Nardone N, Benowitz NL (2018) IQOS: examination of Philip Morris International’s claim of reduced exposure. Tob Control 27:s30–s36
(2020) Heated Tobacco Products (HTPs): Standardized terminology and recommendations for the generation and collection of emissions. Heated Tobacco Products Task Force Technical Report HTP-259-CTR
(1999) Determination of “Tar”, nicotine and carbon monoxide in mainstream tobacco smoke. Health Canada T-115
Wan L, Jinyi L, Weiqi F, Xiangjin Z, Liwen R, Shiwei L, Jinhua W, Tengfei J, Guanhua D (2018) 3-O-acetyl-11-keto-β-boswellic acid exerts anti-tumor effects in glioblastoma by arresting cell cycle at G2/M phase. J Exp Clin Cancer Res 37:132
Ishikawa S, Umemura M, Nakakaji R, Nagasako A, Nagao K, Mizuno Y, Sugiura K, Kioi M, Mitsudo K, Ishikawa Y (2024) EP4-induced mitochondrial localization and cell migration mediated by CALML6 in human oral squamous cell carcinoma. Commun Biol 14:567
Gkouveris I, Nikitakis N, Sklavounou A (2020) p38 expression and modulation of STAT3 signaling in oral cancer. Pathol Oncol Res 26:183–219
Yue J, López JM (2020) Understanding MAPK signaling pathways in apoptosis. Int J Mol Sci 21:2346
Wylam ME, Sathish V, VanOosten SK, Freeman M, Burkholder D, Thompson MA, Pabelick CM, Prakash YS (2015) Mechanisms of cigarette smoke effects on human airway smooth muscle. PLoS ONE 10:e0128778
Scharf P, da Rocha GHO, Sandri S, Heluany CS, Pedreira Filho WR, Farsky SHP (2021) Immunotoxic mechanisms of cigarette smoke and heat-not-burn tobacco vapor on Jurkat T cell functions. Environ Pollut 268:115863
Davis B, To V, Talbot P (2019) Comparison of cytotoxicity of IQOS aerosols to smoke from Marlboro Red and 3R4F reference cigarettes. Toxicol In Vitro 61:104652
Dusautoir R, Zarcone G, Verriele M, Garçon G, Fronval I, Beauval N, Allorge D, Riffault V, Locoge N, Lo-Guidice J-M, Anthérieu S (2021) Comparison of the chemical composition of aerosols from heated tobacco products, electronic cigarettes and tobacco cigarettes and their toxic impacts on the human bronchial epithelial BEAS-2B cells. J Hazard Mater 401:123417
Esteban-Lopez M, Perry MD, Garbinski LD, Manevski M, Andre M, Ceyhan Y, Caobi A, Paul P, Lau LS, Ramelow J, Owens F, Souchak J, Ales E, El-Hage N (2022) Health effects and known pathology associated with the use of E-cigarettes. Toxicol Rep 9:1357–1368
Pagano S, Negri P, Coniglio M, Bruscoli S, Di Michele A, Marchetti MC, Valenti C, Gambelunghe A, Fanasca L, Billi M, Cianetti S, Marinucci L (2021) Heat-not-burn tobacco (IQOS), oral fibroblasts and keratinocytes: cytotoxicity, morphological analysis, apoptosis and cellular cycle. An in vitro study. J Periodontal Res 56:917–928
Hirata N, Horinouchi T, Kanda Y (2022) Effects of cigarette smoke extract derived from heated tobacco products on the proliferation of lung cancer stem cells. Toxicol Rep 9:1273–1280
Chen L, Mayer JA, Krisko TI, Speers CW, Wang T, Hilsenbeck SG, Brown PH (2009) Inhibition of the p38 kinase suppresses the proliferation of human ER-negative breast cancer cells. Cancer Res 69:8853–8861
Leelahavanichkul K, Amornphimoltham P, Molinolo AA, Basile JR, Koontongkaew S, Gutkind JS (2014) A role for p38 MAPK in head and neck cancer cell growth and tumor-induced angiogenesis and lymphangiogenesis. Mol Oncol 8:105–118
Xie C, Zhu J, Wang X, Chen J, Geng S, Wu J, Zhong C, Li X (2019) Tobacco smoke induced hepatic cancer stem cell-like properties through IL-33/p38 pathway. J Exp Clin Cancer Res 38:39
Low B, Liang M, Fu J (2007) p38 Mitogen-activated protein kinase mediates sidestream cigarette smoke- induced endothelial permeability. J Pharmacol Sci 104:225–231
Lin X-X, Yang X-F, Jiang J-X, Zhang S-J, Guan Y, Liu Y-N, Sun Y-H, Xie Q-M (2014) Cigarette smoke extract- induced BEAS-2B cell apoptosis and anti-oxidative Nrf-2 up-regulation are mediated by ROS-stimulated p38 activation. Toxicol Mech Methods 24:575–583
Granata S, Canistro D, Vivarelli F, Morosini C, Rullo L, Mercatante D, Rodriguez-Estrada MT, Baracca A, Sgarbi G, Solaini G, Ghini S, Fagiolino I, Sangiorgi S, Paolini M (2023) Potential harm of IQOS smoke to rat liver. Int J Mol Sci 24:12462
Nishimura A, Zhou L, Kato Y, Mi X, Ito T, Ibuki Y, Kanda Y, Nishida M (2023) Supersulfide prevents cigarette smoke extract-induced mitochondria hyperfission and cardiomyocyte early senescence by inhibiting Drp1-filamin complex formation. J Pharmacol Sci 154(2):127–135
Fearon IM, Acheampong DO, Bishop E (2012) Modification of smoke toxicant yields alters the effects of cigarette smoke extracts on endothelial migration: an in vitro study using a cardiovascular disease model. Int J Toxicol 31:572–583
Arimilli S, Makena P, Prasad GL (2019) Combustible cigarette and smokeless tobacco product preparations differentially regulate intracellular calcium mobilization in HL60 cells. Inflammation 42:1641–1651
Alqarni AA, Brand OJ, Pasini A, Alahmari M, Alghamdi A, Pang L (2022) Imbalanced prostanoid release mediates cigarette smoke-induced human pulmonary artery cell proliferation. Respir Res 23:136
Pezzuto A, Citarella F, Croghan I, Tonini G (2019) The effects of cigarette smoking extracts on cell cycle and tumor spread: novel evidence. Future Sci OA 5:FAO394
Poussin C, van der Toorn M, Scheuner S, Piault R, Kondylis A, Savioz R, Dulize R, Peric D, Guedj E, Maranzano F, Merg C, Morelli M, Egesipe A-L, Johne S, Majeed S, Pak C, Schneider T, Schlage WK, Ivanov NV, Peitsch MC, Hoeng J (2021) Systems toxicology study reveals reduced impact of heated tobacco product aerosol extract relative to cigarette smoke on premature aging and exacerbation effects in aged aortic cells in vitro. Arch Toxicol 95:3341–3359
Bialous SA, Galntz SA (2018) Heated tobacco products: another tobacco industry global strategy to slow progress in tobacco control. Tob Control 27:s111–s117
Acknowledgements
The authors are grateful to Yuto Mizuno, Fumina Suzuki, Wakana Fukae, Chihiro Hayashi, Junko Arai, Makiko Yamada and Mieko Niwa for their assistance with technical aspects. Graphical abstract was created with biorender.com.
Funding
This study was supported in part by the Smoking Research Foundation of Japan.
Author information
Authors and Affiliations
Contributions
K.N., M.U., T. Y., and Y.I. designed the whole study. T.H. prepared the CSE. S.I., Y.I., S.T., R.N., A.N., K.N., and Y.M. conducted the pharmacological and molecular-biological studies. M.U, and K.N. prepared the manuscript. T.A., and M.O. revised the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Kagemichi, N., Umemura, M., Ishikawa, S. et al. Cytotoxic effects of the cigarette smoke extract of heated tobacco products on human oral squamous cell carcinoma: the role of reactive oxygen species and CaMKK2. J Physiol Sci 74, 35 (2024). https://doi.org/10.1186/s12576-024-00928-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12576-024-00928-1