
Abstract
This study provides an updated catalogue of non-indigenous species (NIS) of polychaetes reported from the continental coasts of the Iberian Peninsula based on the available literature. A list of 23 introduced species were regarded as established and other 11 were reported as casual, with 11 established and nine casual NIS in the Atlantic coast of the studied area and 14 established species and seven casual ones in the Mediterranean side. The most frequent way of transport was shipping (ballast water or hull fouling), which according to literature likely accounted for the introductions of 14 established species and for the presence of another casual one. To a much lesser extent aquaculture (three established and two casual species) and bait importation (one established species) were also recorded, but for a large number of species the translocation pathway was unknown. About 25% of the reported NIS originated in the Warm Western Atlantic region, followed by the Tropical Indo West-Pacific region (18%) and the Warm Eastern Atlantic (12%). In the Mediterranean coast of the Iberian Peninsula, nearly all the reported NIS originated from warm or tropical regions, but less than half of the species recorded from the Atlantic side were native of these areas. The effects of these introductions in native marine fauna are largely unknown, except for one species (Ficopomatus enigmaticus) which was reported to cause serious environmental impacts. In other cases, the displacement of native species or economic damages are suspected.
Similar content being viewed by others

Background
Non-indigenous species (NIS) are defined as species introduced outside of their native distribution range (past or present) and outside of their natural dispersal potential by intentional or unintentional human activities [1]. Their translocation is one of the major drivers of biodiversity changes [2]. Global trade and the construction of land or water corridors to facilitate trade or human migration are causing the breakdown of biogeographic barriers leading to an increasing homogenization of the biota [3] in such a way that the occurrence of NIS is altering marine ecosystems all over the world. Some NIS may even become invasive in the recepient environment and when it happens they might displace native species and reduce biodiversity, thus modifying community structures, physical properties of the habitat, food webs and ecosystem function [4, 5]. In extreme cases, this process can lead to financial losses due to fishery decreases and to irreversible damage to vessels, water pipes, piers and other port and canalization infrastructures by fouling organisms [6].
Unfortunately, there is still a lack of information about marine NIS on a global scale and their ecological impacts on native biota as well as their current distribution ranges are not well known [7]. Any NIS that is released into a new recipient environment can potentially start an invasion process that consists of three progressive stages: the release and introduction of the species outside its native range of distribution through human activities, the establishment of a self-sustaining population in the new ecosystem, and the dispersal of this species outside of its site of introduction [8, 9]. It is thus important to asses in which phase of an invasion process an evaluation of the effects on the native biota is performed [10] and to make a follow up of the evolution of already established NIS and newly recorded ones to detect eventual sudden outbursts or blooms [11]. Since once NIS are established, their eradication is almost impossible [12, 13], early interception by controlling potential pathways is probably the most efficient method to prevent future impacts on native ecosystems [14]. This implies a correct taxonomic identification of new arriving NIS and distinguishing them from the native biota, which is sometimes problematic—misidentifications with native species or other NIS frequently occur [11, 15]—and knowledge of the pathways and vectors for NIS introductions with a high level of certainty. Although there are a few exceptions (i.e. [16]), the most common introduction mechanisms for marine NIS are (1) movement of fouling organisms on the hull of ships, (2) release of organisms in solid or liquid ship ballast, (3) movement or intentional release of aquaculture target species along with their associated organisms, (4) connection of water bodies through canals, and (5) release associated with pet industries or bait species [17].
In Europe, the European Strategy on Invasive Alien Species encouraged the implementation of a pan-European inventory of NIS [18]. Within this framework, marine NIS catalogues have been elaborated and published for several countries: France [19], Norway [20], United Kingdom [21, 22], Turkey [23, 24], Denmark [25], Greece [26], the Netherlands [27], Germany [4], Belgium [28], Ireland [29], Cyprus [30], Italy [31], and Portugal [32]. The situation in Europe encompasses a lively, yet fragmented, field of investigation [31]. Their results have been reviewed by a number of research projects funded by the European Commission such as DAISIE: Delivering Alien Invasive Species Inventories for Europe [33] or EASIN: European Alien Species Information Network [34]. From these datasets, the number of marine NIS recorded is estimated to be about 1400 [9]. When comparing to the nearly 33,000 registered European marine species [35], the share of NIS is slightly over 4%. This may represent an insignificant proportion, but the presence of NIS is strongly biased towards environments affected by natural disturbances and/or alterations caused by human activities like estuaries, lagoons, harbours and canals, where native species richness is low [10, 36, 37]. In Europe, the most altered environments, be it due to chemical or organic pollution, physical disturbances or a combination of both, seem to be the most favourable for NIS settlement [38, 39].
The class Polychaeta is the most important taxonomic group within Annelida in terms of species richness, with about 100 families and over 12,000 valid species [40]. They inhabit almost every benthic ecosystem, ranging from littoral environments down to bathyal and abyssal depths. In addition to their great biodiversity, polychaetes play a key role in benthic communities as they take part in the decay of organic sediment matter as well as in the transfer of nutrients to the water column [41]. Although some species are sensitive to high or medium levels of pollution and are useful for environmental monitoring programs [42], many of them are opportunistic species that can withstand high levels of perturbation [43]. Besides environmental tolerance [22], there are a series of biological traits that allow NIS to overcome the negative genetic effects of small founder populations therefore promoting their successful establishment and spread. These include a high genetic variability [44], short generation time, early sexual maturity, high reproductive capacity, and a broad diet [45]. Asexual reproduction [46], hermaphroditism and gregariousness [47] also seem to favour the successful establishment of NIS, as these sexual and behavioural strategies avoid or mitigate the mate finding Allee effect in situations of low population density [48]. Many opportunistic polychaetes show these features that justify them as the second most often detected group of marine animal NIS in Europe, just behind arthropods [9].
In this study, our goals were firstly to elaborate a reliable updated inventory of NIS belonging to Polychaeta using data on their introduced distribution range and the invasion stage they have reached in the studied region, the Atlantic and Mediterranean coast of the Iberian Peninsula, following the unifying quality criteria proposed by Blackburn et al. [8] and Marchini et al. [49], and secondly to reveal eventual regional trends in their introduction pathways and biogeographic origins. The results obtained will provide a baseline to develop regional and national strategies to prevent new introductions of NIS of polychaetes and/or to control the demographic growth and spread of already introduced species.
Methods
For this work, the literature was thoroughly reviewed in search of polychaete NIS records all around the continental coast of Iberian Peninsula, including the Balearic and Chafarinas islands. The Canary, Madeira, and Azores Islands were excluded from this study. Following the quality criteria for establishing normalized inventories [49], only reliable sources that allowed for the validation of the taxonomic identity of the NIS, their time and spatial records and their status in the invasion process were considered for data extraction and analyses. Especial attention was paid to publications in scientific journals and academic works, such as PhD theses. Unpublished reports from monitoring programs, workshops and conferences were only considered when publicly available and their results testable through detailed descriptions or deposition of voucher specimens. Newly recorded species in the time period considered were regarded as true NIS following the published criteria [15]. Thus, species previously known to be absent from the region based on convincing archaeological or historical evidence and whose new record could be directly linked in space and time with a human mediated vector was considered NIS. In the absence of direct evidence of a human-mediated introduction and provided that the natural distribution range was known, the species were considered NIS only if their new records were located clearly beyond their current known distribution limit and outside their natural dispersal potential, rendering intentional or unintentional human mediated introductions as the only translocation mechanisms. Consequently, deep-water and interstitial species whose occurrence cannot be linked to any human activity and those that might be able to reach the Atlantic and/or Mediterranean coasts of the Iberian Peninsula by natural dispersal from neighbouring areas in which they are native or cryptogenic species sensu Carlton [50] were excluded. For nomenclatural purposes, names were updated according to the World Register of Marine Species [51], but in some instances older names or misidentifications were also mentioned. The records were cited in strict chronological order and registered separately for the Atlantic and the Mediterranean coasts of the Iberian Peninsula in order to detect a biogeographic effect, if present. In addition, they were assigned to administrative divisions such as Spanish Marine Demarcations [52] and Portugal (Fig. 1). For each area, the stage of the invasion process for the recorded species was categorized according to the conceptual framework proposed by Blackburn et al. [8], which is summarized in Table 1. Species were considered as established in one location when they were recorded with certainty in the same site or close locations successively over a reasonably long period, assuming that the short life-span of most species implies the presence of self-sustaining populations in these cases. Otherwise, the species was recorded as casual. When vouchers and/or complete descriptions were given with the bibliographic report allowing corroboration of the taxonomic identity, it was clearly mentioned. The casual records not fulfilling this requirement should be considered questionable and were excluded from the checklist, as well as doubtful (cf.) ones. The native distribution of all NIS was assigned to broad oceanic regions according to the classification proposed by Briggs and Bowen [53]. It must be stressed here that unlike animals with hard endo or exoskeletons, such as molluscs, polychaetes hardly ever fossilize and leave remains in archaeological findings, so their native or non-indigenous status is often difficult to assess with certainty [15]. Thus, the present list may grow or shrink when more information on the life cycle and larval development, ecology and evolutionary biology of the polychaetes inhabiting the studied region becomes available. The introduction pathway to the Iberian Peninsula of each species was cited from literature when available, considering it the most likely mechanims. When it was not stated for the Iberian Peninsula, reported pathways to nearby countries were proposed.

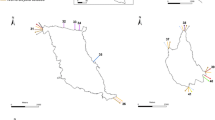
Map of Iberian Peninsula showing the limits between administrative marine demarcations. NOR North-Atlantic Demarcation (Demarcación Marina Noratlántica); PT Portuguese mainland; SUD South-Atlantic Demarcation (Demarcación Marina Sudatlántica); ESAL Gibraltar and Alborán Sea Demarcation (Demarcación Marina del Estrecho y Alborán); LEBA Levantine and Balearic Demarcation (Demarcación Marina Levantino-balear). Derivative of “Positionkarte von der Iberische Halbinseln” © NordNordWest/Wikimedia Commons/CC BY-SA 3.0
Results
According to literature, a total of 23 polychaete NIS have been able to establish in the Iberian coasts (Table 2), with an additional 11 casual records (Table 3). The Atlantic coast of the Iberian Peninsula accounted for 11 established and nine casual NIS of polychaetes, whereas in the Mediterranean coast they numbered 14 and seven, respectively (Fig. 2). Not only were the numbers different, but also the species. Only Ficopomatus enigmaticus and Amphicorina pectinata were established NIS in both the Atlantic and Mediterranean coasts; another three species (Novafabricia infratorquata, Polydora cornuta and Pista unibranchia) were reported from the two coasts, but were only established in the Mediterranean. Two species (Metasychis gotoi and Syllis pectinans) were cited as casual records in both of the two areas. Considering administrative divisions, in Spain 21 established NIS of polychaetes and nine casual ones were found, whereas five established and five casual species have been reported from Portugal.

Number of polychaete NIS recorded from each administrative demarcation of the Iberian Peninsula and from the Atlantic and Mediterranean coasts
The historical evolution of new records of NIS of polychaetes shows that the reports of new polychaete species were just incidental prior to 1980, but their number increased dramatically in the following 10 years. After this point in time, the new records of established NIS have remained stable (Fig. 3). Regarding their origin (Fig. 4), slightly above 25% of the NIS reported for the Iberian Peninsula originated in the Warm Western Atlantic region, followed by the Tropical Indo-West Pacific region (nearly 18%) and the Warm Eastern Atlantic region (about 12%), The remainder of the species are native to an array of biogeographical regions and it must be noted that none were translocated from the Polar Arctic or Antarctic regions. Furthermore, the species originating in Warm Western Atlantic are seemingly the most successful ones, considering that six of the established species came from this region. The Warm Eastern Atlantic region and the Tropical Indo-West Pacific region provided four and three established species, respectively. If the two slopes are considered separately, a clear difference can be noticed. Almost all the species reported from the Mediterranean coast were native from warm or tropical areas, with the Warm Western Atlantic (five established and one casual) and the Tropical Indo-West Pacific (three established and three casual) regions as the main sources. However, the origin of the NIS reported from the Atlantic coast of the Iberian Peninsula is more diverse, since seven established and two casual species are native from cold-temperate regions while four established and seven casual ones originated from warm or tropical areas.

Numbers of new records of polychaete NIS reported from the Iberian Peninsula in each decade of 20th and 21st centuries

Native distribution of NIS of Polychaeta introduced to the Iberian Peninsula
According to availbale literature, shipping was the most common way of translocation as it was the most likely pathway for 14 species (nine with ballast water and five as part of the hull fouling community), although no experimental studies have been conducted. Aquaculture related activities accounted for five introductions and bait industry for another one. Finally, one species is considered a Lessepsian migrant that spread along the Mediterranean basin and eventually arrived to the Spanish coasts. However, for more than one third of the reported species (four established and eight casual) the exact mechanism that facilitated their transport remains unknown (Fig. 5). The most remarkable difference between the Atlantic and Mediterranean coasts is the higher number of fouling species in the latter. In addition, there were a Lessepsian migrant and an introduction related to the fish bait commerce in the Mediterranean, pathways which are to date still unreported from the Iberian Atlantic slope (Tables 2, 3).

Introduction pathway of polychaete NIS to the Iberian Peninsula
Discussion
Is the situation in Spain and Portugal worse than in other European countries?
From the literature examined, we found that 30 polychaete NIS have been recorded from continental Spanish and ten from mainland Portuguese coasts, of which about two thirds were established in one or more locations. Taking into account the Atlantic coast of the Iberian Peninsula, where 20 species were cited, the number was higher than that of other Europeans countries of the Atlantic slope, such as the United Kingdom [22] and France [19, 54] with ten NIS of polychaetes, Germany [4] with nine, Belgium [28, 55] with five, or Ireland [29] with just three. Not only can differences among the countries in the quantity be mentioned, but also in the species involved, with Ficopomatus enigmaticus as the only one present in all these countries. Other shared Iberian polychaetous NIS were Desdemona ornata and Goniadella gracilis, which were also recorded from the United Kingdom.
The number of NIS in the Mediterranean side of the Iberian Peninsula was similar to those from other countries from the Central and Eastern Mediterranean, where 19 NIS of polychaetes were recorded from Cyprus [30], 16 were reported from Greece [26] and 37 are established along Italian coasts [31, 56,57,58]. With just two NIS, the inventory is much less in Malta [59], but it is a much smaller country, whereas the list from Turkey of 66 species is by far the largest, [24]. In fact, compilations for the Mediterranean Sea are more complete as 132 NIS polychaete species [60, 61] have been recorded, of which at least 26 are known to be established in the western area of this sea [62]. Some of the species herein recorded as established are present in all the aforementioned countries except Malta, namely S. marioni, L. collaris and Notomastus aberans, whereas D. ornata and F. enigmaticus have been reported from Turkey, Greece and Italy; Pista unibranchia, Branchiomma luctuosum and B. bairdi from Turkey, Cyprus and Italy; Metasychis gotoi from Turkey, Cyprus and Italy; and Syllis pectinans from Turkey and Italy. The presence of S. marioni, D. ornata, and F. enigmaticus in several Atlantic countries points to their great colonization ability.
The great difference between the Mediterranean and Atlantic national inventories might suggest that the former is under a stronger invasion pressure. In fact, the Mediterranean Sea is among the most affected marine areas in the world where about 820 NIS have been recorded [61], accounting for at least 4.8% of its total biodiversity. This figure might be an underestimation, since a number of species belonging to the smallest fraction of plankton or benthos might have passed overlooked [63]. However, when comparing our data for the Mediterranean and Atlantic Iberian coasts, it can be observed that the difference is not so great, indicating that the number of polychaete NIS is more or less equal when sampling efforts are similar and that lower number in the Atlantic countries can be caused by poorer knowledge of the current situation of polychaete fauna.
Chronological trends in NIS reports
The temporal dynamics of new NIS records of polychaetes shows that prior to 1970 the number of reports of new polychaete species was negligible. Rather than concluding an acceleration of the introduction rates, this fact reflects the poor knowledge of the polychaete fauna in the Iberian coasts at the time. After the most important polychaetologist in Spain, Enrique Rioja was exiled to Mexico at the end of the Spanish Civil War [64], just a handful of local publications were devoted to this group and the main effort in data gathering relied on the work of French scientists [65,66,67,68,69,70]. With the coming of a new generation of researchers, in the 70’s and 80’s, a number of PhD theses and their associated papers [71,72,73,74,75,76] resumed the study of these animals in an exhaustive way. This accounts for the high number of newly recorded species in the following years (Fig. 3). Subsequently, the investigations lead by some of these scientists and the students they trained, together with an increasing concern on environmental issues has maintained the number of new records more or less stable every year.
To what extent are these species harmful?
Thus far, the effects of the settlement of most of these new polychaete species are very poorly understood, mostly as a consequence of the insufficient development of research projects, although the lack of investigation does not justify the assumption that these species have no impact [31]. Streftaris and Zenetos [77] gave a list of invasive species in the Mediterranean biota and established which ones were the worst on account of their known impact. Five of these species have been recorded in our compilation, namely B. luctuosum, F. enigmaticus, Hydroides dirampha, Polydora cornuta and S. marioni.
The NIS belonging to the genus Branchiomma Kölliker, 1858 are relatively large sized tubiculous fan-worms (Sabellidae). They present a very high risk of invasion because they are hermaphrodites with a very short pelagic phase that may produce high population densities starting from only a few individuals [78]. Branchiomma luctuosum was first recorded in Europe from Italy [79], where it was found in littoral lagoons, marinas and other significantly disturbed habitats. In these environments, it seemingly displaced native species [80]. A well-established population is also known from the port of Valencia, but no negative effect was observed [81]. In Spain, B. bairdi was first recorded from Mar Menor Lagoon, where it was especially abundant in meadows of Caulerpa prolifera (Forsskål) J.V. Lamouroux [82], and within a few years it was present in several localities of south-eastern Spain [83] as well as in other parts of the western Mediterranean basin [56, 84]. In Portugal, the species has been reported only from the island of Madeira [85], but not from the mainland coasts. Competition with native suspensions feeders and interfernce with recruitment processes were observed for B. luctuosum [80] and B. bairdi [86] in the Mediterranean Sea. For D. ornata no effect on native species has been observed in British coasts [22], although it has been considered a potential competitor of the native Manayunkia aestuarina (Bourne, 1883) in Northern Spain because they have similar ecological requirements [87]. Albeit no impact is described for Amphicorina pectinata, it is by far the dominant sabellid in the area [88], indicating a probable replacement of native small species of the family.
Serpulid tubeworms settle and build their calcareous tubes on various natural substrates, as well as on artificial ones, forming an important part of the biofouling pests on submerged marine structures with a significant economic impact [77, 89]. In this respect, F. enigmaticus is considered a real conservation threat, as it is the only annelid included in the Spanish official catalogue of invasive species (Real Decreto 630/2013. BOE, 185: 56764). It is a eurihaline species dwelling in estuaries and coastal lagoons. Its highest population densities are achieved in locations with salinities below 30 ppm [90], mild hydrodynamic conditions and high content of organic matter [91]. Under these conditions, it forms large circular reefs that can exceed 1 m in diameter and they have been observed to alter natural water circulation in tidal marshes of southern Atlantic coast of Spain [92]. When populations are very dense, their filter feeding can disturb the natural concentration of nutrients and phytoplankton [93], although the very same effect has also been considered to be beneficial, acting as natural bioremediation in high organic pollution situations [94]. The reefs are structurally stable over time, providing complex three-dimensional habitat in a soft sediment environment, and this biogenic habitat hosts an abundant associated fauna, distinct from nearby mudflats in species composition and relative abundance [95]. NIS belonging to the genus Hydroides also form dense aggregations that cause economic problems [96] and can constitute a major part of the fouling communities in ports and marinas where they displace native species [38, 96]. Spirorbis marioni also build calcareous tubes, but being a minute species its physical impact is much lesser. Nevertheless, alterations on native biodiversity have been reported from the Mediterranean Sea [77].
A third major group of polychaetous NIS is formed by members of the family Spionidae Grube, 1850. In some cases, the colonization of the Iberian coasts has been rapid. For instance, Prionospio pulchra quickly spread along the Atlantic Iberian coast [87, 97] after being reported from its first location in Spain [98] and Pseudopolydora paucibranchiata has also been steadily spreading to the Bay of Biscay [99] and Portugal [97, 100] after its first cite from NW Spain [101]. The successful colonizations by these species seem to be related to their tolerance to severe pollution, as is the case of P. paucibranchiata in some Mediterranean [102] and Atlantic [87] locations, as well as the Spanish population of P. cornuta [103], or of P. pulchra, both within its Japanese native range [104] and in its Mediterranean locations [105]. Regarding the effect on native biota, the extent of the impact differs between species. In Aegean Turkish [106] and Black Sea [107] harbours, where P. cornuta constitutes one of the most conspicuous NIS, massive colonization and replacement of native species with the same ecological requirements have been reported. Similarly, large populations of P. paucibranchiata were reported from locations in NW Spain [101] and southern Portugal [97], where it might replace native species. However, coexistence without replacement of autochthonous taxa has been reported for P. pulchra from colonized locations in California [108]. Thus far, only one shell-boring species has been reported from Spanish coast, Dipolydora socialis [99] and no economic damage was recorded. However, the species is a well-known pest of mollusc aquaculture, both within its native range and in new locations [109].
What is their origin and how did they arrive?
The majority of NIS introduced to European Atlantic waters were found to originate from the North Pacific, particularly the western area, followed by the North-West Atlantic. This suggests that introductions from regions with similar temperature regimes are more likely to become established and widespread, as they would be physiologically adapted to the environmental conditions experienced in the recipient waters [22]. Nevertheless, introductions in the Iberian Peninsula as a whole showed a different pattern. Less than 25% of the established NIS originated in cold temperate regions and the region that provided the higher number of species was the Warm Western Atlantic, where the tropical component is also important, in addition to nine species originating from other warm areas. However, the patterns in the Atlantic and Mediterranean coasts were noticeably different. The Atlantic NIS were a mixture of species with warm and cold-temperate origins, although species from cold-temperate regions were more successful in terms of establishment. It pointed to a condition not so divergent from that described above for the northern European countries. On the contrary, nearly all the reported NIS from the Mediterranean side were native from warm or tropical regions. In this respect, the situation in the Mediterranean coast was more similar to that in some Mediterranean countries, such as Italy, where marine NIS of tropical origin account for half of the total number [31], or to that of the easternmost area of this sea, where 49 out of 52 established polychaete NIS are of tropical origin [62]. Thus, it can be concluded that the Iberian Peninsula is divided in two clear biogrographic units regarding NIS polychaetes. This fact should be considered in order to implement future research and control procedures.
Based on the present records, the principal pathway for the introduction of the established NIS of polychaetes in the Iberian coastal waters was being transported by shipping activities. It involves translocation via commercial or recreational vessel hulls and ship sea-chests (used during ballast water exchange), either as sessile (fouling), boring, vagile, or clinging species [22]. Most of these species were first cited in ports and then expanded to other environments or in some cases even stayed restricted to such localities. Branchiomma bairdi [78], B. luctuosum [81], F. enigmaticus, H. dirampha [110] and S. marioni [31] have been considered as belonging to the hull fouling community and having this mechanism as their more likely way of introduction. Ballast water has also been highlighted in numerous studies as an important transmission vector for NIS, particularly plankton [22], thus being especially important for those species with life cycles involving long planktonic larval stages. This vector is proposed for several members of Spionidae, namely the established Polydora colonia [63], P. cornuta [62], P. paucibranchiata and Boccardia proboscidea [99], but also by some small sabellids, such as A. pectinata and D. ornata [31], and members of other families, including the capitellids N. aberans and Leiochrides australis, and the terebellid P. unibranchia [81]. Thus, introduction via ballast water seems to be the most common pathway. Again, some differences arose between the coasts of the Iberian Peninsula, since the number of introductions related to hull fouling was proportionately fewer in the Atlantic coasts.
Aquaculture was identified as the second major pathway for the importation of marine NIS to European coastal waters [19, 22]. Introductions have occurred, either through the deliberate movement of a target species for culture or unintentional introduction as a ‘hitchhiker’. Considering they have no commercial value, the latter seems the more likely pathway for polychaetes. Only three of the 39 species recorded in this report as established have been related with aquaculture for their introduction: P. pulchra [98], Novafabricia infratorquata [31] and D. socialis [109]. Another two casual ones have also been related to aquaculture activities: Lepidonotus carinulatus [111] and Fauveliopsis glabra [112]. It is much lesser than expected since roughly 20% of the polychaete NIS introduced in Europe as a whole arrived via vectors related to aquaculture in some way [9]. Another unintentional way of introduction is the accidental release of imported bait worms, which apparently caused the establishment of Perinereis linea in SE Spain [113]. Finally, there are possible transmission routes that might include secondary spread from neighboring countries where the NIS arrived via the Suez Canal, as seems to be case of Hesionura serrata [114]. Thus far, the two last mechanisms have been reported only from the Mediterranean coast. However, for the remaining NIS it is still unclear how they arrived.
Some cryptogenic species
During the last 20 years, more than 70 species of polychaetes belonging to different biogeographical regions were newly cited in the Iberian Peninsula. Sometimes difficulties were faced when trying to assess their stages in the invasion process and even when trying to determine if new reports were real introductions of NIS. For several new records of polychaete species found in the literature, it was uncertain whether the extension of their biogeographic distribution and their presence in new localities beyond their known distribution limits was caused by a human mediated dispersal or represented a natural range expansion.
A good example of this is D. marocensis, whose status as NIS is somewhat controversial. The species was first described from the Atlantic coast of Morroco and was subsequently found in at least two separated locations in Portugal [115, 116] and three estuaries from Northen Spain [117], where it can be found sympatrically with the native Diopatra neapolitana Delle Chiaje, 1841. However, D. marocensis is absent from the Atlantic coast of SW Spain [116], the closest Iberian estuarine region to the Moroccan coast. The species has also been recently recorded in the Aegean and Levantine coasts of Turkey [118], but is currently unknown to any other location from the Meditterranean. Berke et al. [116] considered the presence of D. marocensis in the Iberian Peninsula as the result of a human-mediated introduction, probably through the fishing bait trade, since harvesting and selling Diopatra species as a bait is a common resource in Atlantic areas of the Iberian Peninsula [119]. Berke et al. [116] based their statement in the tube brooding of the larvae and the lack of planktonic stages that might facilitate dispersal between locations several hundred kilometers away, rendering all the Iberian populations as virtually isolated as long as only natural means of transport are involved. However, Gil [120] considered these statements mainly speculative and the species was not included in the recently published list of NIS marine species in Portugal [32]. In addition, the populations from the Iberian Peninsula might be relicts from a previous larger population with a wider and more continuous distribution range cannot be excluded. Prospections in suitable locations between the known ones coupled with genetic analyses of representative samples from populations covering the current range of distribution, a powerful tool that helps in unravelling the source and history of human mediated introductions [15, 121], might clarify this issue.
For some cases with a remarkably disjunct distribution and a still poorly known natural geographic distribution, it is difficult to assess whether the species is native or not and therefore they must be considered as cryptogenic, following the definition of the term by Carlton [50]. This may be the case of 15 out of the 17 new records of species belonging to Syllidae reported since 1980 and summarized by San Martín [122], which apart from their type localities are widespread around the Mediterranean basin and Atlantic coast of the Iberian Peninsula. All these records might belong to undescribed crytogenic species that might actually be native of the supposedly colonized habitats. The existence in Syllidae of complexes of sibling species that are morphologically identical and occupy disjunct or extremely wide distribution areas has been demonstrated a number of times by more detailed morphological studies [123] or molecular methods [124]. However, Erinaceusyllis serratosetosa and S. pectinans, which are known from a few localities of the Iberian Peninsula, seem to be true NIS [24, 62], although no human-mediated pathway has been identified for these species. The family Syllidae is not unique in this respect. Some species, newly reported from the Iberian Peninsula and with disjunct distribution, belonging to Pilargidae, such as Ancistrosyllis hamata (Hartman, 1960) and Sigambra parva (Day, 1963), and to Capitellidae, such as Mediomastus capensis Day, 1961, might fall into this category, provided that cryptic species complexes have been detected within both families [125, 126]. The spionid Paraprionospio coora Wilson, 1990, described from eastern Australia, has been considered as cryptogenic in the Mediterranean basin [127] and accordingly we consider the reports from Portugal [120] and the Bay of Biscay [128] in the same way.
Some problems arise when the identity of the species remains unclear. Harmothoe notochaetosa López & San Martín, 1996, a member of Polynoidae, was described based on material from the southern Alborán Sea [129]. The authors stated that the taxon was morphologically very similar to the South-African species H. serrata Day, 1963, but they considered the anatomical differences enough to erect a new taxon. Subsequently, Barnich and Fiege [130] revised the type material of the two taxa and regarded the observed differences of lesser importance, dismissing the new specific name. Nevertheless, because of the extreme distance between the two known localities and the absence of any introduction pathway, the species is herein treated as cryptogenic, awaiting further analyses that might resolve the uncertainty. Another case of likely misidentification is that of Namanereis littoralis (Grube, 1871), a species originally described from Brazil. In the Iberian Peninsula, it has been reported as Namalycastis brevicornis from the Tagus estuary [131] and as N. littoralis from the Basque Country [132]. However, in his revision of the subfamily Namanereidinae, Glasby [133] realized that specimens collected worldwide and referred to as this species and collected worldwide showed great variation in morphological characters. An informal group was created for all these taxa, but a thorough revision of material from the various locations is much needed to assess the number of species forming it. Taking this into account, the species is herein treated as cryptogenic. Lysidice collaris was first reported from the Eastern Mediterranean [134] and was considered a Lessepsian migrant. Some years later it was recorded from Spain [135] and subsequently from a number of Mediterranean countries, being listed as a NIS from Turkey [23], Greece [26], Cyprus [30], and Italy [31]. A comparison of material of the genus with the type series of L. collaris [136] proved that specimens from the Eastern Mediterranean actually belonged to the native Mediterranean L. margaritacea (Claparède, 1868), whereas those of the Iberian Peninsula actually corresponded to L. collaris. Furthermore, an evaluation based on genetic features [137] proved that specimens from Spain and Italy (including Adriatic Sea) were virtually identical, indicating a recent colonization. According to these studies, the species is herein reported as a real NIS for the Iberian Peninsula although it is probably absent from countries of the Eastern Mediterranean, so its Lessepsian introduction is rejected.
Species considered elsewhere as NIS and excluded from the present inventory
In some cases, NIS records result from misidentifications. This is especially frequent for polychaetes [138], resulting in an overestimation of the number of NIS. Nereis jacksoni Kinberg, 1866 is a clear example of this. The species, native of the Southern Australia region, was first recorded from the Mediterranean coast of France [139] and later from Spanish coast near Gibraltar Strait [140]. It has also been reported from Turkey [24, 86], where it is considered a Lessepsian migrant. As a result, Zenetos et al. [62] assumed the species as established in both eastern and western extremes of the Mediterranean Sea. However, a thorough revision of material from the Iberian Peninsula [141] pointed out that the specimens from the western Mediterranean Sea were juveniles of the native Nereis funchalensis (Langerhans, 1880). The real identity of the specimens from the Levantine Sea is still to be clarified, but for Spain N. jacksoni must be excluded from NIS catalogues.
The presence of the South-African hesionid Podarkeopsis capensis (Day, 1963) in the European coasts has been questioned. First reported from the British Isles [142], it has been recorded several times from the Spanish Atlantic coast [143, 144] and Portugal [69, 145]. Some years after its first European report, it was cited from Italy [146] and in the subsequent years was reported from several Mediterranean locations, including the Catalonian coast [147]. Zenetos et al. [62] and Çinar et al. [120] catalogued it as NIS, although some authors consider the identification questionable stating that specimens belonging to the native P. galangaui Laubier, 1961 were erroneously referred to P. capensis [24]. A thorough revision of material from each locality is needed in order to resolve that question. With the same disjunct distribution, Neanthes agulhana (Day, 1963) was reported as a single specimen from the area of the Gibraltar Strait [148] and later as an abundant species from the Balearic Islands [149, 150]. More recently it has been reported from Italy [31]. Apart from the type locality and the Mediterranean ones, the species has never been recorded elsewhere, but when comparing the original description [151] with that of Iberian specimens it can be observed that posterior parapodia are represented with distinctly short neuropodial lobes in the latter, while they are longer in South African specimens [141]. It probably indicates the presence of a cryptogenic taxon instead of a true NIS.
In some other cases, misidentifications are due to a lack of knowledge of the actual native fauna. For instance, Onuphis geophiliformis (Moore, 1903), a Japanese species, was recorded from SW Spain as early as the 70’s [152] and subsequently it was reported as a NIS in the Bay of Biscay [99]. However, a careful examination of material from southern Portugal identical to that described by the former author, demonstrated that it belonged to a new species [120, 153], Onuphis farensis Gil & Machado 2014. This might also be the case of Dispio uncinata (Hartman, 1951), a species native of the Gulf of Mexico. After its first record from NW Spain [154], it has been cited many times from most of the Spanish demarcations: NOR [155], SUD [156, 157] and LEBA [158], although it has not been cited from Portugal. In addition, it has been recorded as a NIS polychaete in a number of Mediterranean countries [62]. However, a recent revision of the genus states that D. uncinata, as currently considered, hides a complex of sibling cryptogenic species; therefore, records from localities different of the type one, including specifically the Iberian Peninsula, should be re-examined [159]. The cirratulid Kirkegaardia dorsobranchialis (Kirkegaard, 1959), described from Southern Africa, was first reported from the Spanish Mediterranean coast in 1972 [70]. Subsequently, it has been recorded several times as a NIS from other locations in the Mediterranean basin [81, 160] although, other authors consider it as a native species [24, 96]. A complete revision of the genus including an examination of the type-series of this species [161] suggests that records of K. dorsobranchialis out of the type locality are likely to belong to a number of new undescribed taxa.
The case of Hydroides dianthus (Verrill, 1873) deserves special attention. It is one of the most documented invasive foulers worldwide and can form dense populations in polluted areas [62], even able to build reef-like aggregations similar to those of F. enigmaticus, although within a narrower range of salinity [91]. The species was originally described from Massachusetts and it has been considered native of the eastern coast of North America, from the type locality to Florida and the Caribbean [162]. In Europe, it was reported from Mediterranean localities of France [163], Spain [164], Cyprus [30], Greece [26], and Turkey [24], and from the French Atlantic coast [165] and the British Isles [22]. Due to its fouling abilities, it was listed within the most harmful invasive species in the Mediterranean [77]. Nevertheless, some authors questioned the identity of the Mediterranean specimens and Zenetos et al. [96] treated the species as a questionable alien. In a recent study, Sun et al. [121] studied the genetic structure of a number of populations of the species and concluded that it encompassed several clades and that the Mediterranean one (excluding populations from the Black Sea) was autochthonous, pending the evaluation of the status of the populations inhabiting the Atlantic European coasts. It must be stressed that some catalogues of NIS are very locally focused, narrowing the concept of “native” species. Spain and France are the only European countries having Atlantic and Mediterranean coasts and in some instances species that are native to the Mediterranean Sea have been recorded as NIS in the Atlantic Ocean. For example, Boccardia semibranchiata Guérin, 1990, that was described from the Gulf of Lyon, in the Mediterranean coast of France, was reported as a NIS from Arcachon Bay [19] and San Sebastián [166], both in the Bay of Biscay. This is also the case of the catalogues for the Basque Country and Bay of Biscay [87, 99] in which as many as another 10 species (Arichlidon reyssi (Katzmann, Laubier & Ramos, 1974), Dasybranchus gajolae Eisig, 1887, Dialychone longiseta (Giangrande, 1992), Microphtahlmus similis Bobretzky, 1870, Nereiphyla pusilla (Claparède, 1870), Oriopsis eimeri (Langerhans, 1881), Paleanotus chrysolepis Schmarda, 1861, Paralacydonia paradoxa Fauvel, 1913, Sabella spallanzanii (Gmelin, 1791) and Syllis westheidei San Martín, 1984) that are native to the Mediterranean coast of Spain are listed as NIS. However, it is well known that the Bay of Biscay, especially in its inner part, shares many species with the Mediterranean biota [167]. Thus, considering Mediterranean species present in the Bay of Biscay a priori as NIS is incorrect and the term cryptogenic (or just native) is more adequate to describe them. Therefore, the taxa mentioned above should be excluded from NIS inventories in the absence of other evidence. In addition, these regional catalogues mention Paranaitis speciosa (Webster, 1879), a phyllodocid, and Asclerocheilus ashworthi Blake, 1981, belonging to Scalibregmatidae, without providing an adequate description or voucher specimens. Thus, they are considered as questionable records. Some of the species of syllids herein described as cryptogenic, namely P. brevicirra, P. wolfi, S. edentatus, and E. belizensis, appear in these documents. The six aforementioned species have been excluded from our list.
Conclusions
This study is the first exhaustive compilation of new records for Spain of polychaete species that are native from different biogeographical areas and updates the previous compilations made for Portugal. According to the current literature, Spain is the third European country in terms of the number of polychaete NIS, with 21 established and another nine casual ones, whereas Portuguese mainland accounts for five established and five casual species. Considering the Iberian distribution of these species, the Mediterranean coast was the area where more NIS have been found, with 14 established and seven casual species, whereas the figure was slightly lower in the Atlantic coast, numbering 20 NIS (11 established and nine casual). The translocation pathway was unknown for a third of the species, but in those for which a vector has been proposed, shipping was the most frequent way of transport, accounting for 14 introductions of established species and for the presence of another casual one (10 in ballast water and five as a part of the fouling fauna). Other known recorded introduction pathways were aquaculture (three established and two casual species) and bait importation (one established species). Approximately 25% of the reported NIS originated in the the Warm Western Atlantic region and nearly 18 and 12% in Tropical Indo-West Pacific and Warm Eastern Atlantic regions, respectively. However, the situation is different if the two sides of the Iberian Peninsula are compared; the species from warm regions constitute the bulk of the Mediterranean NIS, whereas they are approximately half of the taxa reported from the Atlantic coast. It indicates the existence of clearly distinct biogeographical patterns of colonization. The effects of these introductions in native marine fauna are largely unknown, and the displacement of native species in heavily disturbed environments or at least slight economic damages are suspected. Nevertheless, more severe environmental impacts were recorded in SE Spain for Ficopomatus enigmaticus, which causes flow alteration in tidal creeks of brackish marshes.
Abbreviations
- NIS:
-
non-indigenous species
- PhD:
-
Philosophiæ doctor
- NOR:
-
North-Atlantic Spanish Demarcation (Demarcación Marina Noratlántica)
- PT:
-
Portuguese mainland coast
- SUD:
-
South-Atlantic Spanish Demarcation (Demarcación Marina Sudatlántica)
- ESAL:
-
Gibraltar and Alborán Sea Spanish Demarcation (Demarcación Marina del Estrecho y Alborán)
- LEBA:
-
Levantine and Balearic Spanish Demarcation (Demarcación Marina Levantino-balear)
- cf.:
-
Confer (doubtful species level identification)
- BOE:
-
Official Bulletin of the State (Boletín Oficial del Estado)
- NW:
-
north-western
- SW:
-
south-western
- SE:
-
south-eastern
- Aq:
-
aquaculture
- Bt:
-
bait commerce
- BW:
-
ship ballast water
- HF:
-
ship hull fouling
- Ls:
-
Lessepsian migrant
- Uk:
-
unknown
- Cr:
-
cryptogenic species
- DW:
-
deep-water species
- Mi:
-
misidentification
- Qu:
-
questionable identification
References
Olenin S, Alemany F, Cardoso C, Gollasch S, Goulletquer P, Lehtiniemi M, McCollin T, Minchin D, Miossec L, Occhipinti-Ambrogi A, Ojaveer H, Jensen KR, Stankiewicz M, Wallentinus I, Aleksandrov B. Marine strategy framework directive—task group 2 report. Non-indigenous species. EUR24342EN. Luxembourg: Office for Official Publications of the European Communities; 2010.
Halpern BS, Walbridge S, Selkoe KA, Kappel CV, Micheli F, Agrosa CD, Bruno JF, Casey KS, Ebert C, Fox HE, Fujita R, Heinemann D, Lenihan HS, Madin EMP, Perry MT, Selig ER, Spalding M, Steneck R, Watson R. A Global Map of human impact on marine ecosystems. Science. 2008;319:948–52.
Rahel FJ. Biogeographic barriers, connectivity and homogenization of freshwater faunas: it’s a small world after all. Freshw Biol. 2007;52:696–710.
Gollasch S, Nehring S. National checklist for aquatic alien species in Germany. Aquat Invasions. 2006;1:245–69.
Byers JE, Gribben PE, Yeager C, Sotka EE. Impacts of an abundant introduced ecosystem engineer within mudflats of the southeastern US coast. Biol Invasions. 2012;12:2587–600.
Molnar JL, Gamboa RL, Revenga C, Spalding MD. Assessing the global threat of invasive species to marine biodiversity. Front Ecol Environ. 2008;6:485–92.
Galil BS. Loss or gain? Invasive aliens and biodiversity in the Mediterranean Sea. Mar Pollut Bull. 2007;55:314–22.
Blackburn TM, Pysek P, Bacher S, Carlton JT, Duncan RP, Jarosik V, Wilson JR, Richardson DM. A proposed unified framework for biological invasions. Trends Ecol Evol. 2011;26:333–339.
Nunes AL, Katsanevakis S, Zenetos A, Cardoso AC. Gateways to alien invasions in the European seas. Aquat Invasions. 2014;9:133–44.
Reise K, Olenin S, Thieltges DW. Are aliens threatening European aquatic coastal ecosystems? Helgol Mar Res. 2006;60:77–83.
Ojaveer H, Galil BS, Gollasch S, Marchini A, Minchin D, Occhipinti-Ambrogi A, Olenin S. Indentifying top issues of marine invasive alien species in Europe. Manag Biol Invasions. 2014;5:81–4.
Wasson K, Zabin CJ, Bedingen L, Diaz MC, Pearsen JS. Biological invasions of estuaries without international shipping: the importance of intraregional transport. Biol Cons. 2001;102:143–53.
Thresher RE, Kuris AM. Options for managing invasive marine species. Biol Invasions. 2004;6:295–300.
Carlton JT, Ruiz GM. (2005) Vector science and integrated vector management in bioinvasion ecology: conceptual frameworks. In: Mooney HA, Mack RN, McNeely JA, et al., editors. Invasive alien species: a new synthesis. Washington DC: Island Press; 2005. p. 195–212.
Geller JB, Darling JA, Carlton JT. Genetic perspectives on marine biological invasions. Ann Rev Mar Sci. 2010;2:367–93.
Luque Á, Barrajón A, Remón JM, Moreno D, Moro L. Marginella glabella (Mollusca: Gastropoda: Marginellidae): a new alien species from tropical West Africa established in southern Mediterranean Spain through a new introduction pathway. Mar Biodivers Rec. 2012;5:e17. doi:10.1017/S1755267212000012.
Ruiz GM, Carlton JT, Grosholtz ED, Hines AH. Global invasions of marine and estuarine habitats by non-indigenous species: mechanisms, extent and consequences. Am Zool. 1997;37:621–32.
Commision European. Towards an EU strategy on invasive species COM/2008/789. Brussels: European Commision; 2008.
Goulletquer P, Lenz J, Sauria PG, Noel P. Open Atlantic coast of Europe—a century of introduced species into French waters. In: Leppaäkoski E, Gollasch S, Olenis S, editors. Invasive aquatic species of Europe. Distribution, impacts, and management. Dordrecht: Kluwer Academic Publisher; 2002. p. 276–90.
Hopkins CCE. Introduced marine organisms in Norwegian waters, including Svalbard. In: Leppaäkoski E, Gollasch S, Olenis S, editors. Invasive aquatic species of Europe. Distribution, impacts, and management. Dordrecht: Kluwer Academic Publisher; 2002. p. 116–9.
Minchin D, Eno C. Exotics of coastal and inland waters of Ireland and Britain. In: Leppaäkoski E, Gollasch S, Olenis S, editors. Invasive aquatic species of Europe. Distribution, impacts, and management. Dordrecht: Kluwer Academic Publisher; 2002. p. 267–75.
Minchin D, Cook EJ, Clark PF. Alien species in British brackish and marine waters. Aquat Invasions. 2013;8:3–19.
Çinar ME, Bilecenoglu M, Öztürk B, Katagan T, Aysel V. Alien species on the coasts of Turkey. Medit Mar Sci. 2005;6:63–118.
Çinar ME, Dagli E. Kurt Şahin G checklist of Annelida from the coasts of Turkey. Turk J Zool. 2014;38:734–64.
Jensen KR, Knudsen J. A summary of alien marine benthic invertebrates in Danish waters. Hydrobiol Stud. 2005;34:137–62.
Pancucci-Papadopoulos MA, Zenetos A, Corsini-Foka M, Politou C-Y. Update of marine alien species in Hellenic waters. Medit Mar Sci. 2005;6:147–58.
Wolff WJ. Non-indigenous marine and estuarine species in the Netherlands. Zool Meded. 2005;79:1–116.
Kerckhof F, Haelters J, Gollasch S. Alien invertebrates in the marine and brackish ecosystem: the situation in Belgian waters. Aquat Invasions. 2007;2:243–57.
Minchin D. A checklist of alien and cryptogenic aquatic species in Ireland. Aquat Invasions. 2007;2:341–66.
Katsanevakis S, Tsiamis K, Ioannou G, Michailidis N, Zenetos A. Inventory of alien marine species of Cyprus. Medit Mar Sci. 2009;10:109–33.
Occhipinti-Ambrogi A, Marchini A, Cantone G, Castelli A, Chimenz C, Cormaci M, Froglia C, Furnari G, Gambi MC, Giaccone G, Giangrande A, Gravili C, Mastrototaro F, Mazziotti C, Orsi-Relini L, Piraino S. Alien species along the Italian coasts: an overview. Biol Invasions. 2011;13:215–37.
Chainho P, Fernandes A, Amorin A, Ávila SP, Canning-Clode J, Castro JJ, Costa AC, Costa JL, Cruz T, Gollasch S, Grazziotin-Soares C, Melo R, Micael J, Parente MI, Semedo J, Silva T, Sobral D, Sousa M, Torres P, Veloso V, Costa MJ. Non-indigenous species in Portuguese coastal areas, coastal lagoons, estuaries and islands. Estuar Coast Shelf Sci. 2015;167:199–211.
Hulme PE, Roy DB, Cunha T, Larsson T-B. A pan-European inventory of alien species: rationale, implementation and implications for managing biological invasions. In: DAISIE, editor. Handbook of alien species in Europe. Dordrecht: Springer; 2009. p. 1–14.
Tsiamis K, Gervasini E, D’Amico F, Deriu I, et al. The EASIN Editorial Board: quality assurance, exchange and sharing of alien species information in Europe. Manag Biol Invasions. 2016;7:321–8.
MarBEF European Marine Biodiversity Datasets. http://www.marbef.org/data/imis.php?module=dataset. Accessed 26 March 2017.
Nehring S. Four arguments why so many alien species settle into estauries, with special reference to the German river Elbe. Helgol Mar Res. 2006;60:127–34.
Occhipinti-Ambrogi A, Galil B. Marine alien species as an aspect of global change. AIOL. 2010;1:199–218.
Koçak F, Ergen Z, Çinar ME. Fouling organisms and their developments in a polluted and an unpolluted marina in the Aegean Sea (Turkey). Ophelia. 1999;50:1–20.
Occhipinti-Ambrogi A, Savini D. Biological invasions as a component of global change in stressed marine ecosystems. Mar Pollut Bull. 2003;46:542–51.
Appeltans W, Ahyong ST, Anderson G, Angel MV, et al. The magnitude of global marine species diversity. Curr Biol. 2012;22:2189–202.
Hutchings P. Biodiversity and functioning of polychaetes in benthic sediments. Biodivers Conserv. 1998;7:1133–45.
Olsgard F, Brattegard T, Holthe T. Polychaetes as surrogates for marine biodiversity: lower taxonomic resolution and indicator groups. Biodivers Conserv. 2000;12:1033–49.
Dauvin JC, Ruellet T. Polychate/Amphipod ratio revisited. Mar Pollut Bull. 2007;55:215–24.
Rius M, Turon X, Bernardi G, Volckaert FA, Viard F. Marine invasion genetics: from spatio-temporal patterns to evolutionary outcomes. Biol Invasions. 2015;17:869–85.
Essink K, Dekker R. General patterns in invasion ecology tested in the Dutch Wadden Sea: the case of a brackish-marine polychaetous worm. Biol Invasions. 2002;4:359–68.
Roman J, Darling JA. Paradox lost: genetic diversity and the success of aquatic invasions. Trends Ecol Evol. 2007;22:454–64.
Dupont L, Richard J, Paulet YM, Thouzeau G, Viard F. Gregariousness and protandry promote reproductive insurance in the invasive gastropod Crepidula fornicata: evidence from assignment of larval paternity. Mol Ecol Evol. 2006;15:3009–21.
Gascoigne J, Berec L, Gregory S, Courchamps F. Dangerously few liaisons: a review of mate-finding Allee effects. Popul Ecol. 2009;51:355–72. doi:10.1007/s10144-009-0146-4.
Marchini A, Galil B, Occhipinti-Ambrogi A. Recommendations on standardizing lists of marine alien species: lessons from the Mediterranean Sea. Mar Pollut Bull. 2015;101:267–73.
Carlton JT. Biological invasions and cryptogenic species. Ecology. 1996;77:1653–5.
Costello MJ, Bouchet P, Boxshall G, Fauchald K, Gordon D, Hoeksema BW, Poore GCB, van Soest RBM, Stöhr S, Walter TC, Vanhoome B, Decock W, Appeltans W. Global coordination and standardisation in marine biodiversity through the World Register of Marine Species (WoRMS) and related databases. PLoS ONE. 2013;8(1):e51629. doi:10.1371/journal.pone.0051629.
Bellas J. The implementation of the Marine Strategy Framework Directive: shortcomings and limitations from the Spanish point of view. Mar Policy. 2014;50:10–7.
Briggs JC, Bowen BW. A realignment of marine biogeographic provinces with particular reference to fish distributions. J Biogeogr. 2012;39:12–30.
Le Garrec V, Grall J, Chevalier C, Guyonet B, Jourde J, Lavesque N, Bonifácio P, Blake JA. Chaetozone corona (Polychaeta, Cirratulidae) in the Bay of Biscay: a new alien species for the North-east Atlantic waters). J Mar Biol Assoc UK. 2017;97:433–45.
Kerckhof F, Faase MA. Boccardia proboscidea and Boccardiella hamata (Polychaeta: Spionidae: Polydorinae), introduced mud worms new for the North Sea and Europe, respectively. Mar Biodivers Rec. 2014;7:e76. doi:10.1017/S1755267214000803.
Giangrande A, Cosentino A, Lo Presti C, Licciano M. Sabellidae (Annelida) from the Faro coastal lake (Messina, Inonian Sea), with the first recors of the Invasive species Branchiomma bairdi along the Italian coast. Medit Mar Sci. 2012;13:283–93.
Atzori G, López E, Addis P, Sabatini A, Cabiddu S. First record of the alien polychaete Naineris setosa (Scolecida; Orbiniidae) in Tyrrhenian Sea (Western Mediterranean). Mar Biodivers Rec. 2016;9:5. doi:10.1186/s41200-016-0017-6.
Lipej L, Acevedo I, Akel EHK, Anastasopoulou A, et al. New Mediterranean Biodiversity Records (March 2017). Medit Mar Sci. 2017;18:179–201.
Evans J, Barbara J, Schembri PJ. Updated review of marine alien species and other “newcomers” recorded from the Maltese Island (Central Mediterranean). Medit Mar Sci. 2015;16:225–44.
Zenetos Α, Gofas S, Morri C, Rosso A, Violanti D, Garcia Raso JE, Cinar ME, Almogi-Labin A, Ates AS, Azzurro E, Ballesteros E, Biachi CN, Bilecenoglu M, Gambi MC, Giangrande A, Gravili C, Hyams-Kaphzan O, Karachle PK, Katsanevakis S, Lipej L, Mastrototaro F, Mineur F, Pancucci-Papadopoulou MA, Ramos Espla A, Salas C, San Martin G, Sfriso A, Streftaris N, Verlaque M. Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Medit Mar Sci 2012;13:328–52.
Zenetos A, Çinar ME, Crocetta F, Golani D, Rosso A, Servello G, Shnekar N, Turon X, Verlaque M. Uncertainties and validation of alien species: the Mediterranean as an example. Estuar Coast Shelf Sci. 2017;191:171–87.
Zenetos A, Gofas S, Verlaque M, Çinar ME, García Raso JE, Bianchi CN, Morri C, Azzurro E, Bilecenoglu M, Froglia C, Siokou I, Violanti D, Sfriso A, San Martín G, Giangrande A, Katagan T, Ballesteros E, Ramos-Esplá A, Mastrototaro F, Ocaña O, Zingone A, Gambi MC, Streftaris N. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Medit Mar Sci 2010;11:381–493.
Çinar ME. Alien polychaete species worlwide: current status and their impacts. J Mar Biol Assoc UK. 2013;93:1257–78.
Dosil Mancilla FJ, Cremades Ugarte J. El zoólogo Enrique Rioja (1895–1963). Datos sobre su vida y su contribución a la ciencia y a la cultura en España y México. Actas VIII Congreso de la Sociedad Española de Historia de las Ciencias y de las Técnicas. 2004. p. 497–516.
Bellan G. Campagnes de la “Calypso”: Mer d’Alboran. 2. Annélides Polychètes. Ann Inst Océanogr. 1959;37(1):315–42.
Amoureux L. Annélides Polychètes recueillies sur les pentes du talus continental, au large de la Galice (Espagne). Campagnes 1967 et 1968 de la “Thalassa”. Cah Biol Mar. 1972;3:63–89.
Amoureux L. Annélides Polychètes recueillies sur les pentes du talus continental au nord de la côte espagnole, Campagne 1970 de la “Thalassa”. Cah Biol Mar. 1973;14:429–52.
Amoureux L. Annélides Polychètes recueillies sur les pentes du talus continental au nord-ouest de l’Espagne et du Portugal (Campagne 1972 de la “Thalassa”). Cuad Cienc Biol. 1974;3:121–54.
Annélides Amoureux L. Polychètes du talus continental européen. Données complémentaires concernant les campagnes 1970-1971-1972 de la “Thalassa”. Données globales sur l’ensemble des six campagnes (1967 à 1973). Cah Biol Mar. 1987;28:569–77.
Desbruyères D, Guille A, Ramos J. Bionomie benthique du plateau continental de la côte catalane espagnole. Vie Milieu. 1972;23:335–63.
Ibáñez M. Catálogo de los Anélidos Poliquetos citados en las costas españolas. Cuad Cienc Biol. 1973;2:121–40.
Saldanha L. Estudo do povoamento dos horizontes superiors da rocha litoral da costa da Arrabida (Portugal). Arquiv Mus Bocage Ser 2. 1974;5:1–382.
Campoy A. Fauna de España. Fauna de Anélidos Poliquetos de la Península Ibérica. Pamplona: EUNSA/Publicaciones de Biología de la Universidad de Navarra; 1982.
San Martín G. Estudio biogeográfico, faunístico y sistemático de los Poliquetos de la familia Sílidos (Syllidae: Polychaeta) en Baleares. Madrid: Publicaciones de la Universidad Complutense de Madrid, Madrid; 1984.
Sardà R. Contribución al conocimiento de los anélidos poliquetos litorales ibéricos. Estudio sobre la fauna de anélidos poliquetos de las zonas mediolitoral e infralitoral en la región del estrecho de Gibraltar. Ph.D. Thesis. Universidad de Barcelona; 1984.
Ariño A. Optimización de los estudios faunísticos: Ensayo sobre poliquetos sedentarios ibéricos. Ph.D. Thesis. Universidad de Navarra; 1987.
Streftaris N, Zenetos A. Alien Marine Species in the Mediterranean—the 100 ‘Worst Invasives’ and their Impact. Medit Mar Sci. 2006;7:87–118.
Tovar-Hernández MA, Yáñez-Rivera B, Bortolini-Rosales JL. Reproduction of the invasive fan worm Branchiomma bairdi (Polychaeta: Sabellidae). Mar Biol Res. 2011;7:710–8.
Giangrande A. Censimento dei policheti dei mari italiani: Sabellidae Malmgren, 1867. Atti Soc Toscana Sci Nat Mem Ser B. 1989;96:153–89.
Licciano M, Stabili L, Giangrande A, Cavallo RA. Bacterial accumulation by Branchiomma luctuosum (Annelida: Polychaeta). A tool for biomonitoring marine systems and restoring polluted waters. Mar Environ Res. 2007;63:291–302.
El Haddad M, Assadi C, Tasso V, Villaroya L, Gallardo FJ, Capaccioni Azzati R, García-Carrascosa AM, Sáez J, Monforte F. Catálogo preliminar de especies no indígenas de la biota marina del Puerto de Valencia (Mediterráneo Occidental) y su potencial invasivo. Actas II Congreso sobre Especies Exóticas Invasoras “EEI 2006”. GEIB, serie técnica. 2007. p. 202–24.
Román S, Pérez-Ruzafa A, López E. First record in the Western Mediterranean Sea of Branchiomma boholense (Grube, 1878) (Polychaeta: Sabellidae), an alien species of Indo-Pacific origin. Cah Biol Mar. 2009;50:241–50.
Arias A, Giangrande A, Gambi MC, Anadón N. Biology and new records of the invasive species Branchiomma bairdi (Annelida; Sabellidae) in the Mediterranean Sea. Medit Mar Sci. 2013;14:162–71.
Khedhri I, Tovar-Hernández MA, Bonifacio P, Ahmed A, Aleya L. First report of the invasive species Branchiomma bairdi McIntosh, 1885 (Annelida: Sabellidae) along the Tunisian coast (Mediterranean Sea). BioInvasions Rec. 2017;6:139–45.
Ramalhosa P, Camacho-Cruz K, Bastida-Zavala R, Canning-Clode J. First record of Branchiomma bairdi Mc Intosh, 1885 (Annelida: Sabellidae) from Madeira Island, Portugal (northeastern Atlantic Ocean). BioInvasions Rec. 2014;3:235–9.
Çinar ME. Alien polychaete species (Annelida: Polychaeta) on the southern coast of Turkey (Levantine Sea, eastern Mediterranean), with 13 new records for the Mediterranean Sea. J Nat Hist. 2009;43:2283–328.
Zorita I, Solaun O, Galparsoro I, Borja A. Especies exóticas en el medio marino del País Vasco, en relación con el cambio global. Informe para la Dirección de Biodiversidad de la Viceconsejería de Medio Ambiente. Pasaia: Gobierno Vasco; 2009.
Serrano A. Ecología de las poblaciones de poliquetos del entorno de la isla de Mouro (Santander, mar Cantábrico). Ph.D. Thesis. Universidad Autónoma de Madrid; 2002.
Ben-Eliahu MN, ten Hove HA. Serpulidae (Annelida: Polychaeta) from the Suez Canal: from a Lessepsian migration perspective (a monograph). Zootaxa. 2011;2848:1–147.
Fornós JJ, Fortez V, Martínez-Taberner A. Modern polychaete reefs in Western Mediterranean lagoons: Ficopomatus enigmaticus (Fauvel) in the Albufera of Menorca, Balearic Islands. Palaeo. 1997;128:175–86.
Bianchi CN, Morri C. The battle is not to the strong: serpulid reefs in the lagoon of Obetello (Tuscany, Italy). Estuar Coast Shelf Sci. 2001;53:215–20.
Moreno Lampreave D (2010) Flora y fauna alóctona del medio marino. In: Cobos Aguirre J. et al. Especies Exóticas Invasoras en Andalucía. Talleres provinciales 2004–2006. Seville: Consejería de Medio Ambiente/Junta de Andalucía; 2010. p. 214–29.
Bruschetti M, Luppi T, Fanjul E, Rosenthal A, Iribarne O. Grazing effect of the invasive reef-forming polychaete Ficopomatus enigmaticus (Fauvel) on phytoplankton biomass in a SW Atlantic coastal lagoon. J Exp Mar Biol Ecol. 2008;354:212–9.
Hughes DJ, Cook EJ, Sayer MDJ. Biofiltration and biofouling on artificial structures in Europe: the potential for mitigating organic impacts. Oceanogr Mar Biol Ann Rev. 2005;43:123–72.
Heiman KW, Micheli F. Non-native ecosystem engineer alters estuarine communities. Integr Comp Biol. 2010;50:226–36.
Zenetos A, Çinar ME, Pancucci-Papadopoulou MA, Harmelin JG, Furnari G, Andaloro F, Bellou N, Streftaris N, Zibrowius H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Medit Mar Sci. 2005;6:63–118.
Carvalho S, Constantino R, Cerqueira M, Pereira F, Subida MD, Drake P, Gasper MB. Short-term impact of bait digging on intertidal macrozoobenthic assemblages of two Iberian Atlantic systems. Estuar Coast Shelf Sci. 2013;132:65–76.
Moreira J, Parapar J, Troncoso JS. On the presence of Prionospio pulchra (Polychaeta: Spionidae) in the Atlantic Ocean. Cah Biol Mar. 2000;41:233–9.
Martínez J, Adarraga I. Programa de vigilancia y control de la introducción de especies invasoras en los ecosistemas litorales de la costa vasca. 1. Costa de Gipuzkoa. 2. Costa de Bizkaia. Vitoria-Gasteiz: Dirección de Biodiversidad del Departamento de Medio Ambiente y Ordenación del Territorio del Gobierno Vasco; 2006.
.
López-Jamar E, Francesch O, Dorrío AV, Parra S. Long-term variation of the infaunal benthos of La Coruña Bay (NW Spain): results from a 12-year study (1982-1993). Sci Mar. 1995;59(Suppl. 1):49–61.
Dagli E, Çinar ME. Invasion of polluted soft substratum of Izmir Bay (Aegean Sea, eastern Mediterranean) by the spionid worm, Pseudopolydora paucibranchiata (Polychaeta: Spionidae). Cah Biol Mar. 2008;49:87–96.
Tena J, Capaccioni-Azzati R, Porras R, Torres-Gavilá FJ. Cuatro especies nuevas para las costas Mediterráneas en los sedimentos del antepuerto de Valencia. Misc Zool. 1991;15:29–41.
Yokoyama H, Inoue M, Katsuyuki A. Estimation of the assimilative capacity of fish-farm environments based on the current velocity measured by plaster balls. Aquaculture. 2004;240:233–47.
Çinar ME, Katagan T, Öztürk B, Bakir K, Açik S, Dogan A, Bitlis B. Spatio-temporal distribution of zoobenthos in soft substratum of Izmir Bay (Aegean Sea, eastern Mediterranean), with special emphasis on alien species and ecological quality status. J Mar Biol Assoc UK. 2012;9:1457–77.
Çinar ME, Ergen Z, Dağlı E, Petersen ME. Alien species of spionid polychaetes (Streblospio gynobranchiata and Polydora cornuta) in Izmir Bay, eastern Mediterranean. J Mar Biol Assoc UK. 2005;85:821–7.
Radashevsky VI, Selifonova ZP. Records of Polydora cornuta and Streblospio gynobranchiata (Annelida, Spionidae) from the Black Sea. Medit Mar Sci. 2013;14:261–9.
Ranasinghe A, Mikel TK, Velarde RG, Weisberg SB, Montagne DE, Cadien DB, Dalkey A. The prevalence of non-indigenous species in southern California embayments and their effects on benthic macroinvertebrate communities. Biol Invasions. 2005;7:679–86.
Simon CA, Sato-Okoshi W. Polydorid polychaetes on farmed molluscs: distribution, spread and factors contributing to their success. Aquacult Environ Interact. 2015;7:147–66.
Çinar ME, Bilecenoglu M, Öztürk B, Katagan T, Yokes MB, Aysel V, Dagli E, Açik S, Ozcan T, Erdogan H. An updated review of alien species on the coasts of Turkey. Medit Mar Sci. 2011;12:257–315.
Barnich R, Fiege D. The Aphroditoidea (Annelida: Polychaeta) of the Mediterranean Sea. Abh senckenberg naturforsch Ges. 2003;559:1–167.
López E. Occurrence of Fauveliopsis glabra (Fauveliopsida: Annelida: Polychaeta) in a rhodolith bed off the south-eastern coast of Spain (Western Mediterranean). Mar Biodivers Rec. 2011;4:e38. doi:10.1017/S1755267211000418.
Arias A, Richter A, Anadón N, Glasby CJ. Revealing polychaete invasion patterns: identification, reproduction and potential risks of the Korean ragworm, Perinereis linea (Treadwell), in the Western Mediterranean. Estuar Coast Shelf Sci. 2013;131:117–28.
Alós MC. Familia Phyllodocidae Örsted, 1843. In: Viéitez JM et al. Fauna Ibérica. Volumen 25. Annelida, Polychaeta I. Madrid: Museo Nacional de Historia Natural/CSIC; 2004. p. 105–209.
Rodrigues AM, Pires A, Mendo S, Quintino V. Diopatra neapolitana and D. marocensis from the Portuguese coast: morphological and genetic comparison. Estuar Coast Shelf Sci. 2009;85:609–17.
Berke SK, Mahon AR, Lima FP, Halanych KM, Wethey DS, Woodin SA. Range shifts and species diversity in marine ecosystem engineers: patterns and predictions for European sedimentary habitats. Global Ecol Biogeogr. 2010;19:223–32.
Arias A, Anadón N, Paxton H. New records of Diopatra marocensis (Annelida: Onuphidae) from northern Spain. Zootaxa. 2010;2691:67–8.
Çinar ME, Fauchald K, Dagli E. Occurrence of Diopatra marocensis (Annelida, Onuphidae) in the eastern Mediterranean. ZooKeys. 2014;445:1–11. doi:10.3897/zookeys.445.8464.
Cunha T, Hall A, Queiroga H. Estimation of the Diopatra neapolitana annual harvest resulting from digging activity in Canal de Mira, Ria de Aveiro. Fisheries Res. 2005;76:56–66.
Gil J. The European fauna of Annelida Polychaeta. Ph.D. Thesis. Universidade Lisboa; 2011.
Sun Y, Wong E, Keppel E, Williamson JE, Kupriyanova E. A global invader or a complex of regionally distributed species? Clarifying the status of an invasive calcareous tubeworm Hydroides dianthus (Verrill, 1873) (Polychaeta: Serpulidae) using DNA barcoding. Mar Biol. 2017;164:28. doi:10.1007/s00227-016-3058-9.
San Martín G. Familia Syllidae Ehlers, 1864. In: Ramos MA et al., editor. Fauna Ibérica, vol. 21. Annelida, Polychaeta II. Madrid: Museo Nacional de Historia Natural/CSIC; 2003. p. 1–554.
Lattig P, Martin D. A taxonomic revision of the genus Haplosyllis Langerhans, 1887 (Polychaeta; Syllidae: Syllinae). Zootaxa. 2009;2220:1–40.
Álvarez-Campos P, Giribet G, Riesgo A. The Syllis gracilis complex: a molecular approach to a difficult taxonomic problem (Annelida, Syllidae). Mol Phyl Evol. 2017;109:138–50.
Glasby CJ, Marks S. Revision of the genus Synelmis Chamberlin, 1919 (Annelida: Phyllodocida: Pilargidae) in Australia. Zootaxa. 2013;3646:561–74.
Tomioka S, Kondoh T, Sato-Okoshi W, Ito K, Kakui K, Kajihara H. Cosmopolitan or cryptic species? A case study of Capitella teleta (Annelida: Capitellidae). Zool Sci. 2016;33:545–54.
Yokoyama H, Dagli E, Çinar ME. First record of Paraprionospio coora Wilson, 1990 (Polychaeta: Spionidae) from the Mediterranean Sea. Medit Mar Sci. 2010;11:133–41.
Martínez J, Adarraga I. First record of Paraprionospio coora Wilson, 1990 (Polychaeta: Spionidae) in the Atlantic Ocean. BioInvasions Rec. 2013;2:271–80.
López E, San Martín G. A new species of Harmothoe (polychaeta: Polynoidae) from the Chafarinas Islands (Alborán Sea, western Mediterranean). Cah Biol Mar. 1996;37:183–7.
Barnich R, Fiege D. Revision of the Mediterranean species of Harmothoe Kinberg, 1856 and Lagisca Malmgren, 1865 (Polychaeta: Polynoidae: Polynoinae), with descriptions of a new genus and a new species. J Nat Hist. 2000;34:1889–938.
Amoureux L, Calvário J. Annélides Polychètes du Portugal. Données nouvelles. Arquiv Mus Bocage, ser B. 1981;1:145–56.
Gibbs PE, Saiz-Salinas JI. The occurrence of the estuarine polychaete Lycastopsis littoralis (Namanereidinae; Nereididiae) in the Ría de Bilbao, Northern Spain. J Mar Biol Assoc UK. 1996;76:617–23.
Glasby CJ. The Namanereididae (Polycheta: Nereididae). Part 1, Taxonomy and Phylogeny. Rec Aust Mus, Suppl. 1999;25:1–129.
Ben-Eliahu MN. Polychaeta errant of the Suez Canal. Isr J Zool. 1972;21:189–237.
Martin D. Anélidos poliquetos asociados a las concreciones calcáreas del litoral catalán. Misc Zool. 1987;11:61–75.
Kurt Şahin G, Çinar ME. Eunicidae (Polychaeta) species in and around Iskenderun Bay (Levantine Sea, Eastern Mediterranean) with a new alien species for the Mediterranean Sea and re-description of Lysidice collaris. Turk J Zool. 2009;32:331–47.
Ianotta MA, Patti FA, Ambrosino M, Procaccini G, Gambi MC. Phylogeography of two species of Lysidice (Polychaeta, Eunicidae) associated to the seagrass Posidonia oceanica in the Mediterranean Sea. Mar Biol. 2007;150:1115–26.
Galil BS. Taking stock: inventory of alien species in the Mediterranean Sea. Biol Invasions. 2009;11:359–72.
Harmelin J-G. Etude de l’endofaune des “mattes” d’herbiers de Posidonia oceanica Delile. Rec Trav Stat Mar Endoume. 1964;35:43–105.
San Martín G, Acero MI, Contonente M, Gómez JJ. Una colección de Anélidos Poliquetos de las costas mediterráneas andaluzas. Actas II Simposio Ibérico de Estudio del Bentos Marino. 1982;3:171–82.
Núñez J. Familia Nereididae Savigny, 1822. In: Viéitez JM et al., editor. Fauna Ibérica, vol. 25. Annelida, Polychaeta I. Madrid: Museo Nacional de Historia Natural/CSIC; 2004. p. 293–390.
Gibbs PE, Probert K. Notes on Gyptis capensis and Sosane sulcata (Annelida: Polychaeta) from the benthos off the south coast of Cornwall.). J Mar Biol Assoc UK. 1973;53:397–401.
Mora J. Consideraciones generales sobre la macrofauna bentónica de la ría de Arosa. Oecologia Aquatica. 1982;6:41–9.
Moreira J, Parapar J. Hesionidae y Pilargidae (Annelida, Polychaeta) del proyecto “Fauna Ibérica” (Campañas “Fauna II, III y IV”), recogidos a lo largo de las costas de la península Ibérica. Bol R Soc Esp Hist Nat (Sec Biol). 2003;98:47–55.
Martins R, Sampaio L, Rodrigues AM, Quintino V. Soft-bottom Portuguese continental shelf polychaetes: diversity and distribution. J Mar Syst. 2013;123(124):41–54.
Gravina MF, Giangrande A. A new record for the Mediterranean Sea: Podarkeopsis capensis (Polychaeta, Hesionidae). Atti Soc Tosc Sci Nat Mem Ser B. 1988;95:155–60.
Méndez N, Cardell MJ. Littoral annelid polychaetes inhabiting soft bottoms of the Barcelonès (Catalonia, NE Spain). Misc Zool. 1996;19:119–47.
Sardà R. Anélidos poliquetos del Estrecho de Gibraltar. I. Amphinomida, Spintherida y Phyllodocida. Misc Zool. 1985;9:65–78.
Soler A, Ballesteros M, Turón X. Poliquetos del Estany des Peix (Formentera, Baleares). Aproximación al estudio faunístico y ecológico. Historia Animalium, 1997;3:9–23.
Box A, Martin D, Deudero S. Changes in seagrass polychaete assemblages after invasion by Caulerpa racemosa var, cylindracea (Chlorophyta: Caulerpales): community structure, trophic guilds and taxonomic distinctness. Sci Mar. 2010;74:317–29.
Day JH. The polychaete fauna of South Africa. Part 8. New species and records from grab samples and dredgings. Bull Brit Mus (Nat Hist), Zool. 1963;10:381–445.
Ibáñez M. Notas sobre algunas especies de Anélidos Poliquetos nuevas para las costas de España, con especial referencia a Nereis (Neanthes) oxypoda Marenzeller y Onuphis (Nothria) geophiliformis Moore. Bol R Soc Esp Hist Nat (Sec Biol). 1972;70:23–31.
Gil J, Machado M. A new species of Onuphis (Polychaeta: Onuphidae) from Southern Portugal, with comments on the validity of O. pancerii Claparède, 1868. Zootaxa. 2014;3860:343–60.
Ibáñez M, Viéitez JM. Presencia en la costa cantábrica del anélido poliqueto Dispio uncinata: contribución a su estudio sistemático y ecológico. Bol R Soc Esp Hist Nat (Sec Biol). 1973;71:5–8.
Junoy J, Castellanos C, Viéitez JM, de la Huz MR, Lasta M. The macroinfauna of the Galician Sandy beaches (NW Spain) affected by the Prestige oil-spill. Mar Pollut Bull. 2005;50:526–36.
Sardà R. Fauna de anélidos poliquetos de la región del estrecho de Gibraltar. V-Comunidades localizadas en sustratos blandos. Misc Zool. 1987;11:77–86.
Mayoral MA, López-Serrano L, Viéitez JM. Benthic macrofauna of three intertidal sandy beaches of the Río Piedras mouth (Huelva, Spain). Bol R Soc Esp Hist Nat (Sec Biol). 1994;91:231–40.
Martí E, Torres-Gavilá J, Tena J, Rodilla M, Sánchez-Arcilla A, Mestres M, Mösso C. Temporal Changes in Mollusk and Polychaete Communities in the Soft Bottom of Cullera Bay (Western Mediterranean). J Coastal Res Sp Iss. 2007;47:76–83.
Delgado-Blas VH, Díaz-Díaz O. Redescription of two species and five new species of Dispio Hartman, 1951 (Spionidae: Polychaeta) from the eastern Pacific Coast and Caribbean Sea, with a review of the genus. Zootaxa. 2016;4178(1):151–81.
Streftaris N, Zenetos A, Papathanassiou E. Globalisation in marine ecosystems: the story of non-indigenous marine species across European seas. Oceanogr Mar Biol Ann Rev. 2005;43:419–53.
Blake JA. Kirkegaardia (Polychaeta, Cirratulidae), new name for Monticellina Laubier, preoccupied in the Rhabdocoela, together with new records and descriptions of eight previously known and sixteen new species from the Atlantic, Pacific, and Southern Seas. Zootaxa. 2016;4166:1–93.
Bastida-Zavala R, ten Hove HA. Revision of Hydroides Gunnerus, 1768 (Polychaeta: Serpulidae) from the Western Atlantic region. Beaufortia. 2002;52:103–78.
Zibrowius H. Remarques sur trois espèces de Serpulidae acclimatées en Méditerranée: Hydroides dianthus (Verrill, 1873), Hydroides dirampha Mörch, 1863, et Hydroides elegans (Haswell, 1883). Rapp Comm Int Mer Medit. 1973;21:683–6.
Capaccioni-Azzati R. Anélidos Poliquetos de la ensenada de los Alfaques (delta del Ebro, Mediterráneo Occidental). Ph.D. Thesis, Universitat de València; 1987.
Dauvin JC, Dewarumez JM, Gentil F. Liste actualisée des espèces d’Annélides Polychètes présentes en Manche. Cah Biol Mar. 2003;44:67–95.
Martínez J, Adarraga I, López E. Nuevos datos del género Boccardia Carazzi, 1893 (Polychaeta: Spionidae) para la península Ibérica y el oceáno Atalntico. Bol Inst Esp Oceanogr. 2006;22:53–64.
Casamajor M-N, Lalanne Y. Intérét biogéographique de la côte basque rocheuse. Bull Soc Zool Fr. 2016;141:3–13.
Ravara A, Moreira MH. Polychaeta (Annelida) from the continental shlef off Aveiro (NW Portugal): species composition and community structure. Check List. 2013;9:533–9.
Capaccioni-Azzati R. Prionospio multibranchiata (Polychaeta, Spionidae), Notomastus aberans y N. formianus (Polychaeta, Capitellidae) en el litoral de la península Ibérica. Misc Zool. 1988;12:47–56.
Torres-Gavilá FJ. Estudio faunístico, ecológico y ambiental de la fauna de Anélidos Poliquetos de sustratos sueltos de las islas Chafarinas (Mar de Alborán, S.W. Mediterráneo). Valencia: Servei de Publicacions, Universitat de València; 2008.
López E. Anélidos poliquetos de sustratos duros de las islas Chafarinas. Ph.D. Thesis. Universidad Autónoma de Madrid; 1995.
Bick A. Redescription of Fabriciola tonerella Banse, 1959, and a new record of Novafabricia infratorquata (Fitzhugh, 1983) from the Mediterranean Sea, with a key for the Fabriciinae (Annelida: Polychaeta) of the Mediterranean Sea and the north-east Atlantic. Zool Anz. 2005;244:137–52.
Cepeda D, Lattig P. A new species of Polycirridae (Annelida: Terebellida) and three new reports for Cantabrian and Mediterranean Seas. Cah Biol Mar. 2016;57:371–87.
Martin D, Cuesta JA, Drake P, Gil J, Nygren A, Pleijel F. The symbiotic hesionid Parasyllidea humesi Pettibone, 1961 (Annelida: Polychaeta) hosted by Scrobicularia plana (da Costa, 1778) (Mollusca: Bivalvia: Semelidae) in European waters. Org Divers Evol. 2012;12:145–53.
Cardell MJ, Méndez N. First record of Hesionura serrata (Hartmann-Schröder, 1996) (Polychaeta: Phyllodocidae) in the Mediterranean Sea (littoral zone of Barcelona, NE Spain). Sci Mar. 1996;60:423–6.
Tena J. Faunística y ecología de los Anélidos Poliquetos de los fondos de sustrato duro del archipiélago de las Chafarinas (SW mar de Alborán). Ph.D. Thesis. Universitat de València; 1996.
Ceberio A, Martínez J, Aguirrezabalaga F. Presencia de Desdemona ornata Banse, 1957 (Polychaeta, Sabellidae) en las costas de la Península Ibérica, Golfo de Vizcaya. Munibe. 1988;50:37–41.
Machado M, Cancela da Fonseca L. Nota sobre o macrozoobentos de uma instalação de piscicultura intensiva (Olhão, Portugal). Actas do 9º Congresso do Algarve. 1997. p. 907–19.
La Rioja E. Mercierella enigmatica Fauvel, serpúlido de agua salobre, en España. Bol R Soc Esp Hist Nat. 1924;24:160–9.
Margalef R. Materiales para la hidrobiología de la isla de Menorca. P Inst Biol Appl. 1952;11:1–112.
Cancela da Fonseca L. Estudo da influencia da “abertura ao mar” sobre um sistema lagunar costeiro: a lagoa de Santo André. Ph.D. Thesis. Universidade Lisboa; 1989.
Rioja E. Estudio de los Poliquetos de la Península Ibérica. Mem Acad Cienc Ex Fís Nat ser Zool. 1931;2:1–471.
Zibrowius H, Bianchi CN. Spirorbis marioni et Pileolaria berkeleyana, Spirorbidae exotiques dans les ports de la Mediteranée nord-occidentale. Rapp Commun Int Mer Medit. 1981;27:163–4.
Aguirre O, San Martín G, Baratech L. Presencia de la especie Polydora colonia Moore, 1907 (Polychaeta, Spionidae) en las costas españolas. Misc Zool. 1986;10:375–7.
Ravara A. Estudo da comunidade de poliquetas da plataforma continental aolargo de Aveiro. Master Thesis. Universidade de Aveiro; 1997.
Redondo MS, San Martín G. Anélidos poliquetos de la costa comprendida entre el cabo de San Antonio y el puerto de Valencia (Mediterráneo occidental). Publ Espec Inst Esp Oceanogr. 1997;23:225–33.
Capaccioni-Azzati R, Torres-Gavilá FJ, Tena J. Dos nuevos géneros de anélidos poliquetos para la península Ibérica: Neopseudocapitella Rullier and Amoureux, 1979 y Demonax Kinberg, 1867. Thalassas. 1993;10:115–21.
Martínez J, Adarraga I. Nuevas citas de anélidos poliquetos y de un anfípodo marino en la Península Ibérica. Bol R Soc Esp Hist Nat (Sec Biol). 2001;96:137–50.
Baratech L, Aguirre O, San Martín G. Primera cita de la especie Lepidonotus carinulatus (Grube, 1870) (Polychaeta: Polynoidae) en aguas españolas. Bol Inst Esp Oceanogr. 1986;3:1–4.
San Martín G, González G, López-Jamar E. Aspectos sistemáticos y ecológicos sobre algunas especies de Sílidos (Polychaeta. Syllidae) en las costas gallegas. Bol Inst Esp Oceanogr. 1985;2:27–36.
López E, San Martín G, Jiménez M. Syllinae (Syllidae, Annelida, Polychaeta) from Chafarinas Islands (Alborán Sea, W Mediterranean). Misc Zool. 1996;19:105–18.
Authors’ contributions
EL conducted the main literature review on Polychaeta, compiled the data, and led writing of the paper. AR initiated the study, reviewed the general literature on biological invasions, and co-wrote the paper. Both authors read and approved the final manuscript.
Acknowledgements
We are grateful to J. Moreira and G. San Martín, from Universidad Autónoma de Madrid, for their advice and loan of bibliography, and to Mary J. MacKenty, for her thorough revision of the English language. The useful comments of two anonymous reviewers as well as that by the journal editor are acknowledged for greatly improving the scientific quality of this work.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
Consent for publication
In this study, human participants whose consent for publication was needed were not involved.
Ethics approval and consent to participate
Since neither live animals were used for experimentation nor biological material (be it genetically modified or not) were used for the current study, approval from the “Cómite de Ética en la Investigación” (Ethics Committee for Research) of the Universidad Autónoma de Madrid was not required, as established in the University, regional and national normatives. In no case human participants whose consent to participate was needed were involved.
Funding
This work was partially funded by the project “Polychaeta VI: Palpata-Canalipalpata I (CGL2014-53332-C5-3-P)”.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
López, E., Richter, A. Non-indigenous species (NIS) of polychaetes (Annelida: Polychaeta) from the Atlantic and Mediterranean coasts of the Iberian Peninsula: an annotated checklist. Helgol Mar Res 71, 19 (2017). https://doi.org/10.1186/s10152-017-0499-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s10152-017-0499-6